


Neuroprotective Effects of Ethanol Extract of Polyscias fruticosa (EEPF) against Glutamate-Mediated Neuronal Toxicity in HT22 Cells
 , ,
, ,
Abstract
:1. Introduction
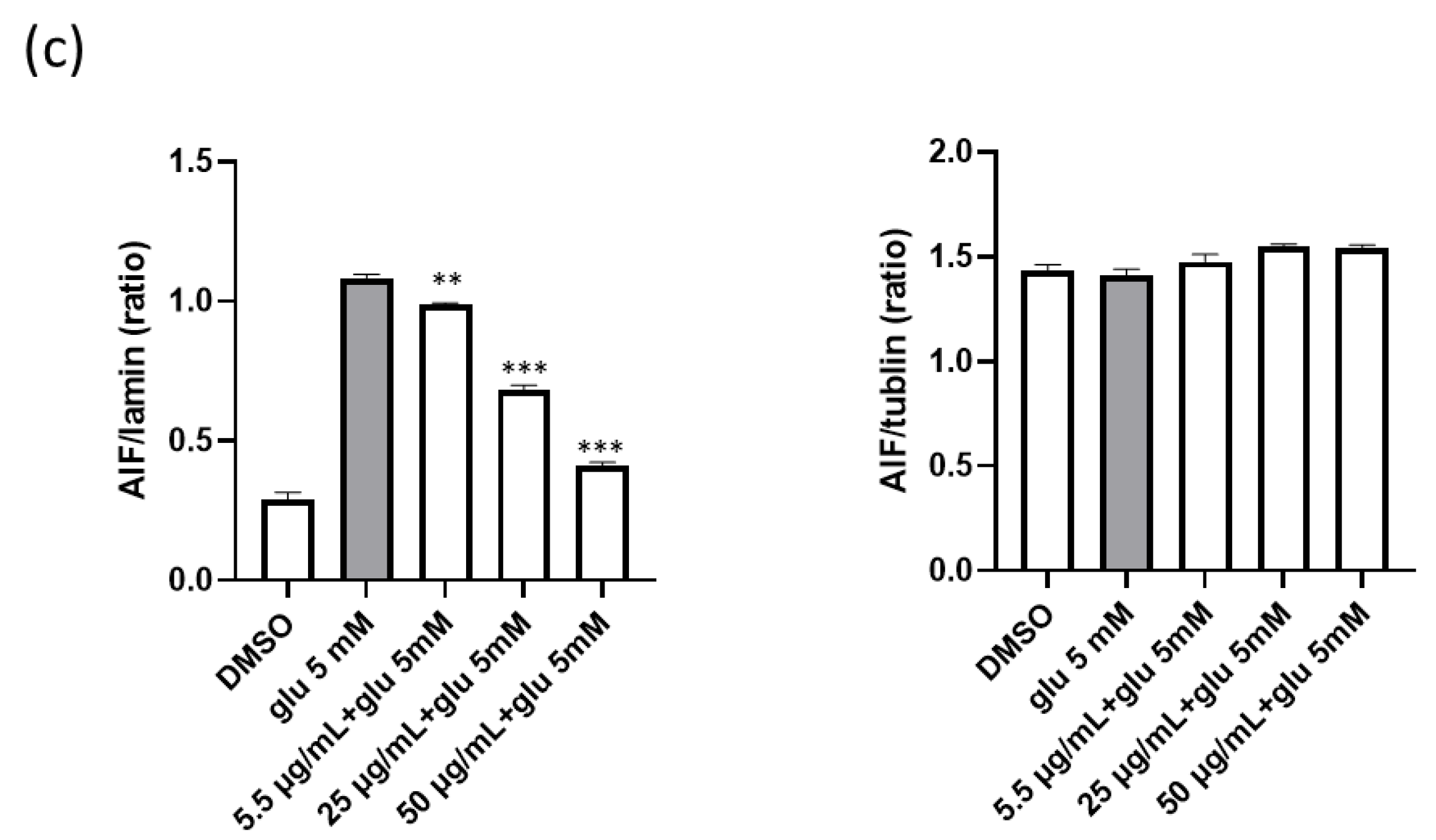
2. Results
2.1. Neuroprotection of P. fruticosa Extracts against Glutamate-Induced Cell Death
2.2. The 75% EEPF Prevents ROS Generation and Ca2+ Influx
2.3. The 75% EEPF Inhibits Glutamate-Induced Apoptotic Cell Death
2.4. The 75% EEPF Inhibits the MAPK and Activates AKT/BDNF/CREB Signal Pathway
2.5. The 75% EEPF Reverses Glutamate-Mediated Altered Ratio of Bcl-2 and Bax Proteins
2.6. Neuroprotective Effects of 75% EEPF against Ischemic Brain Injury
2.7. Identification of Novel Compounds in EEPF
3. Discussion
4. Materials and Methods
4.1. General Experimental Procedures
4.2. Preparation of EEPF (Ethanol Extract of Polyscias fruticosa)
4.3. Extraction and Isolation
4.4. Cell Culture
4.5. Calcein-AM Live-Cell Assay
4.6. DCF-DA ROS Assay
4.7. Fluo-3 AM Calcium Assay
4.8. Western Blot Analysis
4.9. Apoptosis Analysis by Flow Cytometry
4.10. Pretreatment of EEPF In Vivo Study
4.11. Induction of Transient Cerebral Ischemia
4.12. Histology
4.13. Cresyl Violet Staining
4.14. Staining with Fluoro-Jade C
4.15. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Bean, A.R. A conspectus of Polyscias J.R.Forst. & G.Forst. (Araliaceae) in Queensland, Australia. Austrobaileya 2015, 9, 445–456. [Google Scholar]
- Do, T.L. Medicinal Plants and Remedy of Vietnam; Publisher of Medicine: Hanoi, Vietnam, 2004. [Google Scholar]
- Vo, V.C. Dictionary of Vietnamese Medicinal Plants; Publisher of Medicine: Hanoi, Vietnam, 2012. [Google Scholar]
- Ly, H.T.; Nguyen, T.T.H.; Le, V.M.; Lam, B.T.; Mai, T.T.T.; Dang, T.P.T. Therapeutic potential of Polyscias fruticosa (L.) Harms leaf extract for parkinson’s disease treatment by Drosophila melanogaster model. Oxid. Med. Cell. Longev. 2022, 2022, 5262677. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, T.-T.-D.; Nguyen, Q.-D.; Nguyen, T.-V.-L. Kinetic Study on Chlorophyll and Antioxidant Activity from Polyscias fruticosa (L.) Harms Leaves via Microwave-Assisted Extraction. Molecules 2021, 26, 3761. [Google Scholar] [CrossRef] [PubMed]
- Bernard, B.M.; Pakianathan, N.; Divakar, M.C. On the antipyretic, anti-inflammatory, analgesic and molluscicidal properties of Polyscias fruticosa (L.) Harms. Anc. Sci. Life 1998, 17, 313–319. [Google Scholar]
- Koffuor, G.A.; Boye, A.; Kyei, S.; Ofori-Amoah, J.; Asiamah, E.A.; Barku, A.; Acheampong, J.; Amegashie, E.; Awuku, A.K. Anti-asthmatic property and possible mode of activity of an ethanol leaf extract of Polyscias fruticose. Pharm. Biol. 2016, 54, 1354–1363. [Google Scholar] [CrossRef] [Green Version]
- Tran, P.T.; Dang, N.H.; Kim, O.; Cuong, P.V.; Dat, N.T.; Hwang bo, C.; Minh, C.V.; Lee, J.-H. Ethanol extract of Polyscias fruticosa leaves suppresses RANKL-mediated osteoclastogenesis in vitro and LPS-induced bone loss in vivo. Phytomedicine 2019, 59, 152908. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Xilouri, M.; Bruban, J.; Shao, Z.; Papazolou, J.; Vekrellis, K.; Robakis, N.K. Extracellular progranulin protects cortical neurons from toxic insults by activating survival signaling. Neurobiol. Ageing 2011, 32, 2326.e5-16. [Google Scholar] [CrossRef] [Green Version]
- Duchen, M.R. Mitochondria and calcium: From cell signaling to cell death. J. Physiol. 2000, 529, 57–68. [Google Scholar] [CrossRef]
- Tobaben, S.; Grohm, J.; Seiler, A.; Conrad, M.; Plesnila, N.; Culmsee, C. Bid-mediated mitochondrial damage is a key mechanism in glutamate-induced oxidative stress and AIF-dependent cell death in immortalized HT-22 hippocampal neurons. Cell Death Differ. 2011, 18, 282–292. [Google Scholar] [CrossRef] [Green Version]
- Jung, H.; Kim, S.Y.; Cecen, F.S.C.; Cho, Y.; Kwon, S.-K. Dysfunction of mitochondrial Ca2+ regulatory machineries in brain aging and neurodegenerative diseases. Front. Cell Dev. Biol. 2020, 18, 599792. [Google Scholar] [CrossRef]
- Pereira, C.F.; de Oliveria, C.R. Oxidative glutamate toxicity involves mitochondrial dysfunction and perturbation of intracellular Ca2+ homeostasis. Neurosci. Res. 2000, 37, 227–236. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Landshamer, S.; Hoehn, M.; Barth, N.; Duvezin-Caubet, S.; Schwake, G.; Tobaben, S.; Kazhdan, I.; Becattini, B.; Zahler, S.; Vollmar, A.; et al. Bid-induced release of AIF from mitochondria causes immediate neuronal cell death. Cell Death Differ. 2008, 15, 1553–1563. [Google Scholar] [CrossRef] [PubMed]
- Garrido, C.; Galluzzi, L.; Brunet, M.; Puig, P.E.; Didelot, C.; Kroemer, G. Mechanisms of cytochrome c release from mitochondria. Cell Death Differ. 2006, 13, 1423–1433. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, D.H.; Kim, D.W.; Jung, B.H.; Lee, J.H.; Lee, H.; Hwang, G.S.; Kang, K.S.; Lee, J.W. Ginsenoside Rb2 suppresses the glutamate-mediated oxidative stress and neuronal cell death in HT22 cells. J Ginseng Res. 2009, 43, 326–334. [Google Scholar] [CrossRef]
- Son, Y.; Cheong, Y.-K.; Kim, N.-H.; Chung, H.-T.; Kang, D.G.; Pae, H.-O. Mitogen-Activated Protein Kinases and Reactive Oxygen Species: How Can ROS Activate MAPK Pathways? J. Signal Transduct. 2011, 2011, 792639. [Google Scholar] [CrossRef] [Green Version]
- Barr, R.K.; Bogoyevitch, M.A. The c-Jun N-terminal protein kinase family of mitogen-activated protein kinases (JNK MAPKs). Int. J. Biochem. Cell Biol. 2001, 33, 1047–1063. [Google Scholar] [CrossRef]
- Yue, J.; López, J.M. Understanding MAPK signaling pathways in apoptosis. Int. J. Mol. Sci. 2020, 21, 2346. [Google Scholar] [CrossRef] [Green Version]
- Lavoie, H.; Gagnon, J.; Therrien, M. ERK signaling: A master regulator of cell behavior, life and fate. Nat. Rev. Mol. Cell Biol. 2020, 21, 607–632. [Google Scholar] [CrossRef]
- Sánchez-Alegría, K.; Flores-León, M.; Avila-Muñoz, E.; Rodríguez-Corona, N.; Arias, C. PI3K signaling in neurons: A central node for the control of multiple functions. Int. J. Mol. Sci. 2018, 19, 3725. [Google Scholar] [CrossRef] [Green Version]
- Nakaso, K.; Tajima, N.; Horikoshi, Y.; Nakasone, M.; Hanaki, T.; Kamizaki, K.; Matsura, T. The estrogen receptor beta-PI3K/Akt pathway mediates the cytoprotective effects of tocotrienol in a cellular Parkinson’s disease model. Biochim. Biophys. Acta 2014, 1842, 1303–1312. [Google Scholar] [CrossRef] [Green Version]
- Qina, R.; Li, X.; Li, G.; Tao, L.; Li, Y.; Sun, J.; Kang, X.; Chen, J. Protection by tetrahydroxystilbene glucoside against neurotoxicity induced by MPP+: The involvement of PI3K/Akt pathway activation. Toxicol. Lett. 2011, 202, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Walton, M.R.; Dragunow, M. Is CREB a key to neuronal survival? Trends Neurosci. 2000, 23, 48–53. [Google Scholar] [CrossRef]
- Elonze, B.; Ginty, D.D. Function and regulation of CREB family transcription factors in the nervous system. Neuron 2002, 35, 605–623. [Google Scholar]
- Almeida, R.D.; Manadas, B.J.; Melo, C.V.; Gomes, J.R.; Mendes, C.S.; Grâos, M.M.; Carvalho, F.R.; Carvalho, A.P.; Duarte, C.B. Neuroprotection by BDNF against glutamate-induced apoptotic cell death is mediated by ERK and PI3-kinase pathways. Cell Death Differ. 2005, 12, 1329–1343. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahn, S.M.; Kim, H.N.; Kim, Y.R.; Choi, Y.W.; Kim, C.M.; Shin, H.K.; Cho, B.T. Emodin from Polygonum multiflorum ameliorates oxidative toxicity in HT22 cells and deficits in photothrombotic ischemia. J. Ethnopharm. 2016, 188, 13–20. [Google Scholar] [CrossRef]
- Wu, L.; Xiong, X.; Wu, X.; Ye, Y.; Jian, Z.; Zhi, Z.; Gu, L. Targeting oxidative stress and inflammation to prevent ischemia reperfusion injury. Front. Mol. Neurosci. 2020, 13, 28. [Google Scholar] [CrossRef] [Green Version]
- Fukui, M.; Song, J.-H.; Choi, J.; Choi, H.J.; Zhu, B.T. Mechanism of glutamate-induced neurotoxicity in HT22 mouse hippocampal cells. Eur. J. Pharmacol. 2009, 617, 1–11. [Google Scholar] [CrossRef]
- Lee, Y.B.; Du, S.; Rhim, H.; Lee, E.B.; Markelonis, G.J.; Oh, T.H. Rapid increase in immunoreactivity to GFAP in astrocytes in vitro induced by acidic pH is mediated by calcium inflex and calpain I. Brain Res. 2000, 864, 220–229. [Google Scholar] [CrossRef]
- Quideau, S.; Grabber, J.H.; Hatfieida, R.D. Identification and synthesis of new ferulic acid dehydrodimers present in grass cell walls. J. Chem. Soc. 1994, 602, 3485–3498. [Google Scholar]
- Funk, C.; Ralph, J.; Steinhart, H.; Bunzel, M. Isolation and structural characterization of 8-O-4/8-O-4- and 8-8/8-O-4-coupled dehydrotriferulic acids from maize bran. Phytochemistry 2005, 66, 363–371. [Google Scholar] [CrossRef]
- Chemam, Y.; Benayache, S.; Marchinoni, E.; Zhao, M.; Mosset, P.; Benayache, F. On-line screening, isolation and identification of antioxidant compounds of Helianthemum ruficomum. Molecules 2007, 8, 239. [Google Scholar] [CrossRef] [PubMed]
- Huang, W.-Y.; Sheu, S.-J. Separation and identification of the organic acids in Angelicae Radix and Ligustici Rhizoma by HPLC and CE. J Sep. Sci. 2006, 29, 2616–2624. [Google Scholar] [CrossRef] [PubMed]
- Kato, H.; Takahashi, A.; Itoyama, Y. Cell cycle protein expression in proliferating microglia and astrocytes following transient global cerebral ischemia in the rat. Brain Res Bull. 2003, 60, 215–221. [Google Scholar] [CrossRef] [PubMed]
- Park, C.W.; Lee, T.-K.; Cho, J.H.; Kim, I.H.; Lee, J.-C.; Shin, B.-N.; Ahn, J.H.; Kim, S.K.; Shin, M.C.; Ohk, T.G.; et al. Rufinamide pretreatment attenuates ischemia-reperfusion injury in the gerbil hippocampus. Neurol. Res. 2017, 39, 941–952. [Google Scholar] [CrossRef]
- Ko, I.-G.; Jin, J.J.; Hwang, L.; Kim, S.H.; Kim, C.J.; Jeon, J.W.; Chung, J.Y.; Han, J.H. Adenosine A2A receptor agonist polydeoxyribonucleotide ameliorates shortterm memory impairment by suppressing cerebral ischemia-induced inflammation via MAPK pathway. PLoS ONE 2021, 16, e0248689. [Google Scholar] [CrossRef]
- Yano, S.; Morioka, M.; Fukunaga, K.; Kawano, T.; Hara, T.; Kai, Y.; Hamada, J.; Miyamoto, E.; Ushio, Y. Activation of Akt/Protein Kinase B Contributes to Induction of Ischemic Tolerance in the CA1 Subfield of Gerbil Hippocampus. J. Cereb. Blood Flow Metab. 2001, 21, 351–360. [Google Scholar] [CrossRef] [Green Version]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| 1H (Methanol-d4, 500 MHz) | 13C(Methanol-d4, 125 MHz) | ||||
|---|---|---|---|---|---|
| 1 | 2 | 1 | 2 | ||
| benzene ring A | 1 | 126.5 | 126.9 | ||
| 2 | 7.57 (1H, d, J = 8.45) | 7.44 (1H, s) | 134.5 | 115.0 | |
| 3 | 6.74 (1H, d, J = 8.45) | 117.9 | 150.2 | ||
| 4 | 161.6 | 151.1 | |||
| 5 | 6.74 (1H, d, J = 8.50) | 6.76 (1H, d, J = 8.25) | 117.9 | 117.5 | |
| 6 | 7.57 (1H, d, J = 8.45) | 7.11 (1H, d, J = 8.30) | 134.5 | 127.7 | |
| 7 | 7.37 (1H, s) | 7.41 (1H, s) | 129.8 | 130.5 | |
| 8 | 140.5 | 140.0 | |||
| 9 | 168.2 | 168.0 | |||
| OCH3 | 3.70 (3H, s) | 57.2 | |||
| benzene ring B | 1 | 118.8 | 119.0 | ||
| 2 | 7.33 (1H, s) | 7.33 (1H, s) | 114.1 | 113.9 | |
| 3 | 151.9 | 151.8 | |||
| 4 | 150.7 | 150.5 | |||
| 5 | 6.79 (1H, d, J = 8.25) | 6.79 (1H, d, J = 8.40) | 116.3 | 115.9 | |
| 6 | 7.08 (1H, d, J = 8.30) | 7.08 (1H, d, J = 8.40) | 124.4 | 124.4 | |
| 7 | 7.62 (1H, d, J = 15.9) | 7.62 (1H, d, J = 15.9) | 147.3 | 147.2 | |
| 8 | 6.39 (1H, d, J = 15.9) | 6.40 (1H, d, J = 15.9) | 118.9 | 118.9 | |
| 9 | 171.9 | 171.8 | |||
| OCH3 | 3.99 (3H, s) | 3.99 (3H, s) | 58.0 | 58.0 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Selvaraj, B.; Le, T.T.; Kim, D.W.; Jung, B.H.; Yoo, K.-Y.; Ahn, H.R.; Thuong, P.T.; Tran, T.T.T.; Pae, A.N.; Jung, S.H.; et al. Neuroprotective Effects of Ethanol Extract of Polyscias fruticosa (EEPF) against Glutamate-Mediated Neuronal Toxicity in HT22 Cells. Int. J. Mol. Sci. 2023, 24, 3969. https://doi.org/10.3390/ijms24043969
Selvaraj B, Le TT, Kim DW, Jung BH, Yoo K-Y, Ahn HR, Thuong PT, Tran TTT, Pae AN, Jung SH, et al. Neuroprotective Effects of Ethanol Extract of Polyscias fruticosa (EEPF) against Glutamate-Mediated Neuronal Toxicity in HT22 Cells. International Journal of Molecular Sciences. 2023; 24(4):3969. https://doi.org/10.3390/ijms24043969
Chicago/Turabian StyleSelvaraj, Baskar, Tam Thi Le, Dae Won Kim, Bo Hyun Jung, Ki-Yeon Yoo, Hong Ryul Ahn, Phuong Thien Thuong, Thi Thu Thuy Tran, Ae Nim Pae, Sang Hoon Jung, and et al. 2023. "Neuroprotective Effects of Ethanol Extract of Polyscias fruticosa (EEPF) against Glutamate-Mediated Neuronal Toxicity in HT22 Cells" International Journal of Molecular Sciences 24, no. 4: 3969. https://doi.org/10.3390/ijms24043969