
Physiological and Biochemical Responses of Solanum lycopersicum L. to Benzo[a]pyrene Contaminated Soils

 ,
,  , , , , and
, , , , and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Uptake of BaP in Plant Tissues and Impact on Morphometric Parameters
2.2. Assessment of Photosynthetic Pigments
2.3. Assessment of Localization of Superoxide in Situ
2.4. Enzymatic Activity in Plants of Model Experience
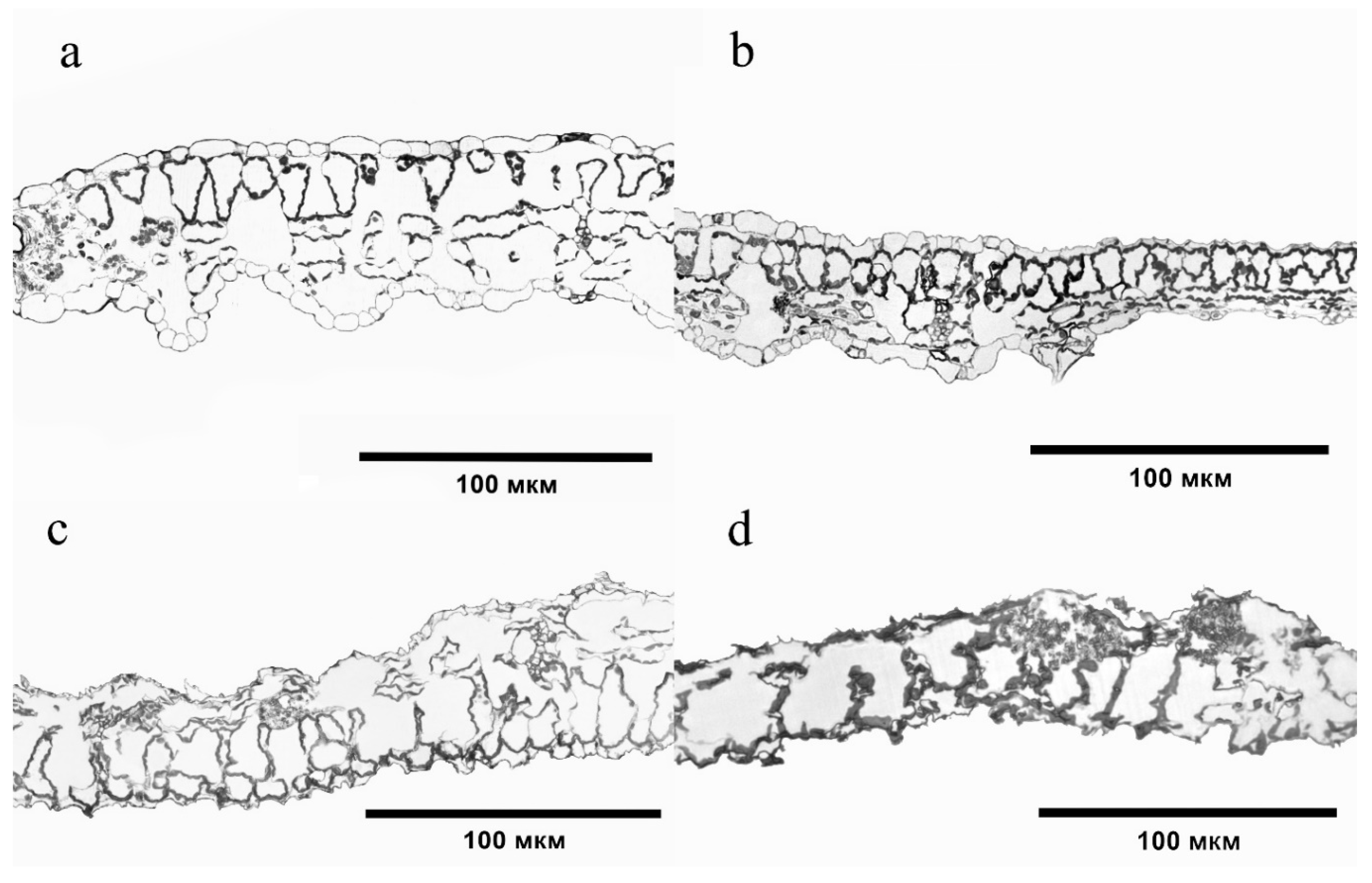
2.5. Microscopic Examination
3. Discussion
4. Materials and Methods
4.1. Experimental Setup and Plant Growth Conditions
4.2. Measurement of Morphometric Parameters
4.3. BaP Content in Plants Tissues
4.4. Determination of Chlorophyll a,b, Total Chlorophyll, and Carotenoids
4.5. Localization of Superoxide in Situ
4.6. Determination of Lipid Peroxidation
4.7. Plant Sample Preparation to Determine the Activity of Antioxidant Enzymes and Total Protein
4.7.1. Protein Definition
4.7.2. Determination of Proline (L: 210-189-3)
4.7.3. Determination of Superoxide Dismutase Activity (EC 1.15.1.1)
4.7.4. Determination of Catalase Activity (EC 1.11.1.6)
4.7.5. Determination of Glutathione Reductase Activity (EC 1.8.1.7)
4.7.6. Determination of Glutathione Peroxidase Activity (EC 1.11.1.9)
4.7.7. Determination of Ascorbate Peroxidase Activity (EC 1.11.1.11)
4.7.8. Determination of Total Peroxidase Activity (EC 1.11.1)
4.7.9. Glutathione-S-Transferase (EC2.5.1.18)
4.7.10. Definition of Glutathione
4.8. Light Microscopy
4.9. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| BaP | Benzo[a]pyrene |
| MPC | Maximum permissible concentration |
| CAT | Catalase |
| SOD | Superoxide dismutase |
| MDA | Malonic dialdehyde |
| PRX | Peroxidase |
| APOX | Ascorbate peroxidase |
| GP | Glutathione peroxidase |
| PAHs | Polycyclic aromatic hydrocarbons |
| GSH | Gluthatione |
| AS | Antioxidant system |
| HPLC | High performance liquid chromatography |
| HEPES | 4-(2-Hydroxyethyl)-1-piperazineethanesulfonic acid |
| NBT | Nitro blue tetrazolium |
| TBA | Thiobarbituric acid |
| DW | Dry weight |
| PVP | Polyvinylpyrrolidone |
| PMSF | Phenylmethylsulfonyl fluoride |
| NADPH | Nicotinamide adenine dinucleotide phosphate |
| Tris | Tris (hydroxymethyl) aminomethane |
| CDNB | 1-Chloro-2,4-dinitrobenzene |
| GST | Glutathione-S-transferase |
References
- Selvi, S.; Polat, R.; Çakilcioğlu, U.; Celep, F.; Dirmenci, T.; Ertuğ, Z.F. An ethnobotanical review on medicinal plants of the Lamiaceae family in Turkey. Turk. J. Bot. 2022, 46, 283–332. [Google Scholar] [CrossRef]
- Sushkova, S.N.; Vasilyeva, G.K.; Minkina, T.M.; Mandzhieva, S.S.; Tjurina, I.G.; Kolesnikov, S.I.; Askin, T. New method for benzo [a] pyrene analysis in plant material using subcritical water extraction. J. Geochem. Explor. 2014, 144, 267–272. [Google Scholar] [CrossRef]
- GN 2.1.5.689-98; Maximum Allowable Concentrations (MPC) of Chemicals in the Water of Drinking, Cultural and Social Waters. State Committee for Sanitary and Epidemiological Supervision and Ministry of Health of Russian Federation: Moscow, Russia, 1998; 35p.
- Sushkova, S.; Minkina, T.; Mandzhieva, S.; Zamulina, I.; Bauer, T.; Vasilyeva, G.; Antonenko, E.; Rajput, V.; Kızılkaya, R. Features of accumulation, migration, and transformation of benzo [a] pyrene in soil-plant system in a model condition of soil contamination. J. Soils Sediments 2018, 18, 2361–2367. [Google Scholar] [CrossRef]
- Abdel-Shafy, H.I.; Mansour, M.S. A review on polycyclic aromatic hydrocarbons: Source, environmental impact, effect on human health and remediation. Egypt. J. Pet. 2016, 25, 107–123. [Google Scholar] [CrossRef]
- Fedorenko, G.M.; Fedorenko, A.G.; Minkina, T.M.; Mandzhieva, S.S.; Rajput, V.D.; Usatov, A.V.; Sushkova, S.N. Method for hydrophytic plant sample preparation for light and electron microscopy (studies on Phragmites australis Cav.). MethodsX 2018, 5, 1213–1220. [Google Scholar] [CrossRef]
- González, A.; Espinoza, D.; Vidal, C.; Moenne, A. Benzopyrene induces oxidative stress and increases expression and activities of antioxidant enzymes, and CYP450 and GST metabolizing enzymes in Ulva lactuca (Chlorophyta). Planta 2020, 6, 1–13. [Google Scholar] [CrossRef]
- Belz, R.G.; Sinkkonen, A. Low toxin doses change plant size distribution in dense populations–Glyphosate exposed Hordeum vulgare as a greenhouse case study. Environ. Int. 2019, 132, 105072. [Google Scholar] [CrossRef]
- Lai, R.W.; Yeung, K.W.Y.; Yung, M.M.N.; Djurišić, A.B.; Giesy, J.P.; Leung, K.M.Y. Regulation of engineered nanomaterials: Current challenges insights and future directions. Environ. Sci. Pollut. Res. 2018, 25, 3060–3077. [Google Scholar] [CrossRef]
- Sam, S.M. Biochemical and physiological response of Abelmoschus esculentus (L.) moench to food industry processing effluent. J. Appl. Sci. Environ. Manag. 2022, 26, 517–527. [Google Scholar] [CrossRef]
- Amin, M.A.; Ahmad, U.B.; Aliyu, A.M.; Adam, I.A.; Aliyu, R.W. Growth and yield response of tomato to different soil amendment techniques under different water Stress. South Asian Res. J. Agri. Fish 2023, 5, 1–9. [Google Scholar] [CrossRef]
- Gerszberg, A.; Hnatuszko-Konka, K.; Kowalczyk, T.; Kononowicz, A.K. Tomato (Solanum lycopersicum L.) in the service of biotechnology. Plant Cell Tissue Organ Cult. 2015, 120, 881–902. [Google Scholar] [CrossRef]
- Paduchuri, P.; Gohokar, S.; Thamke, B.; Subhas, M. Transgenic tomatoes-a review. Int. J. Adv. Biotechnol. Res. 2010, 1, 69–72. [Google Scholar]
- Tarigholizadeh, S.; Motafakkerazad, R.; Salehi-Lisar, S.Y.; Kazemi, E.M. High resistance of Panicum miliaceum L. to phenanthrene toxicity based on growth response and antioxidant system assessment. Acta Agric. Slov. 2021, 117, 1–13. [Google Scholar] [CrossRef]
- Fedorenko, A.G.; Chernikova, N.P.; Minkina, T.M.; Sushkova, S.N.; Dudnikova, T.S.; Antonenko, E.M.; Fedorenko, G.M.; Bauer, T.V.; Mandzhieva, S.S. Effects of benzo[a]pyrene toxicity on morphology and ultrastructure of Hordeum sativum. Environ. Geochem. Health 2021, 43, 1551–1562. [Google Scholar] [CrossRef]
- Daresta, B.E.; Italiano, F.; de Gennaro, G.; Trotta, M.; Tutino, M.; Veronico, P. Atmospheric particulate matter (PM) effect on the growth of Solanum lycopersicum cv. Roma plants. Chemosphere 2015, 119, 37–42. [Google Scholar] [CrossRef]
- Kummerová, M.; Barták, M.; Dubová, J.; Tříska, J.; Zubrová, E.; Zezulka, Š. Inhibitory effect of fluoranthene on photosynthetic processes in lichens detected by chlorophyll fluorescence. Ecotoxicology 2006, 15, 121–131. [Google Scholar] [CrossRef]
- Sharma, P.; Jha, A.B.; Dubey, R.S.; Pessarakli, M. Reactive oxygen species, oxidative damage, and antioxidative defense mechanism in plants under stressful conditions. J. Bot. 2012, 2012, 217037. [Google Scholar] [CrossRef]
- Abbas, S.K.; Hassan, H.A.; Asif, J.; Ahmed, B.; Haider, S.S. Integration of TTF, UTAUT, and ITM for mobile Banking Adoption. Int. J. Adv. Eng. Manag. Sci. 2018, 4, 5. [Google Scholar] [CrossRef]
- Lushchak, V.I. Contaminant-induced oxidative stress in fish: A mechanistic approach. Fish Physiol. Biochem. 2016, 42, 711–747. [Google Scholar] [CrossRef]
- Naveed, M.; Bukhari, S.S.; Mustafa, A.; Ditta, A.; Alamri, S.; El-Esawi, M.A.; Rafique, M.; Ashraf, S.; Siddiqui, M.H. Mitigation of nickel toxicity and growth promotion in sesame through the application of a bacterial endophyte and zeolite in nickel contaminated soil. Int. J. Environ. Res. Public Health 2020, 17, 8859. [Google Scholar] [CrossRef]
- Naveed, M.; Mustafa, A.; Azhar, S.Q.; Kamran, M.; Zahir, Z.A.; Unez-Delgado, A.N. Burkholderia phytofirmans PsJN and tree twigs derived biochar together retrieved Pb-induced growth, physiological and biochemical disturbances by minimizing its uptake and translocation in mung bean (Vigna radiata L.). J. Environ. Manag. 2020, 257, 109974. [Google Scholar] [CrossRef] [PubMed]
- Ahammed, G.J.; Yuan, H.L.; Ogweno, J.O.; Zhou, Y.H.; Xia, X.J.; Mao, W.H.; Shi, K.; Yu, J.Q. Brassinosteroid alleviates phenanthrene and pyrene phytotoxicity by increasing detoxification activity and photosynthesis in tomato. Chemosphere 2012, 86, 546–555. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Zhu, L.; Wang, J.; Xie, H.; Wang, J.; Han, Y.; Yang, J. Oxidative stress and lipid peroxidation in the earthworm Eisenia fetida induced by low doses of fomesafen. Environ. Sci. Pollut. Res. 2013, 20, 201–208. [Google Scholar] [CrossRef] [PubMed]
- Wang, R.; Paul, V.J.; Luesch, H. Seaweed extracts and unsaturated fatty acid constituents from the green alga Ulva lactuca as activators of the cytoprotective Nrf2–ARE pathway. Free Radic. Biol. Med. 2013, 57, 141–153. [Google Scholar] [CrossRef] [PubMed]
- Wang, W. The effect of storage condition and duration on the deterioration of primed rice seeds. Front. Plant Sci. 2018, 9, 172. [Google Scholar] [CrossRef]
- Shen, Q.; Yu, J.; Fu, L.; Wu, L.; Dai, F.; Jiang, L.; Wu, D.; Zhang, G. Ionomic, metabolomic and proteomic analyses reveal molecular mechanisms of root adaption to salt stress in Tibetan wild barley. Plant Physiol. Biochem. 2018, 123, 319–330. [Google Scholar] [CrossRef]
- Liu, H.; Weisman, D.; Ye, Y.B.; Cui, B.; Huang, Y.H.; Colón-Carmona, A.; Wang, Z.H. An oxidative stress response to polycyclic aromatic hydrocarbon exposure is rapid and complex in Arabidopsis thaliana. Plant Sci. 2009, 176, 375–382. [Google Scholar] [CrossRef]
- Cevher-Keskin, B.; Selçukcan-Erol, Ç.; Yüksel, B.; Ertekin, O.; Yıldızhan, Y.; Onarıcı, S.; Kulen, O.; Memon, A.R. Comparative transcriptome analysis of Zea mays in response to petroleum hydrocarbon stress. Environ. Sci. Pollut. Res. 2018, 25, 32660–32674. [Google Scholar] [CrossRef]
- Kummerová, M.; Zezulka, S.; Babula, P.; Váňová, L. Root response in Pisum sativum and Zea mays under fluoranthene stress: Morphological and anatomical traits. Chemosphere 2013, 90, 665–673. [Google Scholar] [CrossRef]
- Habig, W.H.; Pabst, M.J.; Jakoby, W.B. Glutathione S-transferases: The first enzymatic step in mercapturic acid formation. J. Biol. Chem. 1974, 249, 7130–7139. [Google Scholar] [CrossRef]
- Hernández-Vega, J.C.; Cady, B.; Kayanja, G.; Mauriello, A.; Cervantes, N.; Gillespie, A.; Lavia, L.; Trujillo, J.; Alkio, M.; Colón-Carmona, A. Detoxification of polycyclic aromatic hydrocarbons (PAHs) in Arabidopsis thaliana involves a putative flavonol synthase. J. Hazard. Mater. 2017, 321, 268–280. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Apel, K.; Hirt, H. Reactive oxygen species: Metabolism, oxidative stress, and signal transduction. Ann. Rev. Plant Biol. 2004, 55, 373–399. [Google Scholar] [CrossRef] [PubMed]
- Lichtenthaler, H.K.; Wellburn, A.R. Determinations of total carotenoids and chlorophylls a and b of leaf extracts in different solvents. Biochem. Soc. Trans. 1983, 11, 591–592. [Google Scholar] [CrossRef]
- IUSS Working Group WRB. World Reference Base for Soil Resources 2014, Update 2015. International Soil Classification System for Naming Soils and Creating Legends for Soil Maps; World soil resources reports no. 106; FAO: Rome, Italy, 2015. [Google Scholar]
- Sushkova, S.N.; Minkina, T.M.; Tarigholizadeh, S.; Rajput, V.; Fedorenko, A.G.; Antonenko, E.M.; Dudnikova, T.S.; Chernikova, N.P.; Yadav, B.K. Soil PAHs contamination effect on the cellular and subcellular organelle changes of Phragmites australis Cav. Environ. Geochem. Health 2021, 43, 2407–2421. [Google Scholar] [CrossRef] [PubMed]
- Tewari, H.B.; Bourne, G.H. Histochemical evidence of metabolic cycles in spinal ganglion cells of rat. J. Histochem. Cytochem. 1962, 10, 42–64. [Google Scholar] [CrossRef]
- Heath, R.L.; Packer, L. Photoperoxidation in isolated chloroplasts. I. Kinetics and stoichiometry of fatty acid peroxidation. Arch. Biochem. Biophys. 1968, 125, 180–198. [Google Scholar] [CrossRef]
- Hodges, D.M.; DeLong, J.M.; Forney, C.F.; Prange, R.K. Improving the thiobarbituric acid-reactive-substances assay for estimating lipid peroxidation in plant tissues containing anthocyanin and other interfering compounds. Planta 1999, 4, 604–611. [Google Scholar] [CrossRef]
- Bradford, M.M. Rapid and sensitive method for quantitation of microgram quantities of protein utilizing principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Demiral, T.; Türkan, I. Comparative lipid peroxidation, antioxidant defense systems and proline content in roots of two rice cultivars differing in salt tolerance. Environmen. Exp. Bot. 2005, 53, 247–257. [Google Scholar] [CrossRef]
- Sirota, T.V. Novel approach to the study od adrenaline auto-oxidation and its use for the measurements of superoxide dismutase activity. Vopr. Med. Khim. 1999, 45, 263–272. [Google Scholar]
- Nair, A.R.; Agbor, L.N.; Mukohda, M.; Liu, X.; Hu, C.; Wu, J.; Sigmund, C.D. Interference with endothelial PPAR (Peroxisome Proliferator–Activated Receptor)-γ causes accelerated cerebral vascular dysfunction in response to Endogenous Renin-Angiotensin System Activation. Hypertension 2018, 72, 1227–1235. [Google Scholar] [CrossRef] [PubMed]
- Arora, A.; Sairam, R.K.; Srivastava, G.C. Oxidative stress and antioxidative system in plants. Curr. Sci. 2002, 82, 1227–1238. [Google Scholar]
- Qayyum, S.; Nasir, A.; Mian, A.H.; Rehman, S.; Qayum, S.; Siddiqui, M.F.; Kalsoom, U. Extraction of peroxidase enzyme from different vegetables for biodetoxification of vat dyes. Appl. Nanosci. 2020, 10, 5191–5199. [Google Scholar] [CrossRef]
- Ellman, G.L. Tissue sulfhydryl groups. Arch. Biochem. Biophys. 1959, 82, 70–77. [Google Scholar] [CrossRef]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Voloshina, M.; Rajput, V.D.; Chernikova, N.; Minkina, T.; Vechkanov, E.; Mandzhieva, S.; Voloshin, M.; Krepakova, M.; Dudnikova, T.; Sushkova, S.; et al. Physiological and Biochemical Responses of Solanum lycopersicum L. to Benzo[a]pyrene Contaminated Soils. Int. J. Mol. Sci. 2023, 24, 3741. https://doi.org/10.3390/ijms24043741
Voloshina M, Rajput VD, Chernikova N, Minkina T, Vechkanov E, Mandzhieva S, Voloshin M, Krepakova M, Dudnikova T, Sushkova S, et al. Physiological and Biochemical Responses of Solanum lycopersicum L. to Benzo[a]pyrene Contaminated Soils. International Journal of Molecular Sciences. 2023; 24(4):3741. https://doi.org/10.3390/ijms24043741
Chicago/Turabian StyleVoloshina, Marina, Vishnu D. Rajput, Natalia Chernikova, Tatiana Minkina, Evgeniy Vechkanov, Saglara Mandzhieva, Mark Voloshin, Maria Krepakova, Tamara Dudnikova, Svetlana Sushkova, and et al. 2023. "Physiological and Biochemical Responses of Solanum lycopersicum L. to Benzo[a]pyrene Contaminated Soils" International Journal of Molecular Sciences 24, no. 4: 3741. https://doi.org/10.3390/ijms24043741