
Suppression of Rice Osa-miR444.2 Improves the Resistance to Sheath Blight in Rice Mediating through the Phytohormone Pathway
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
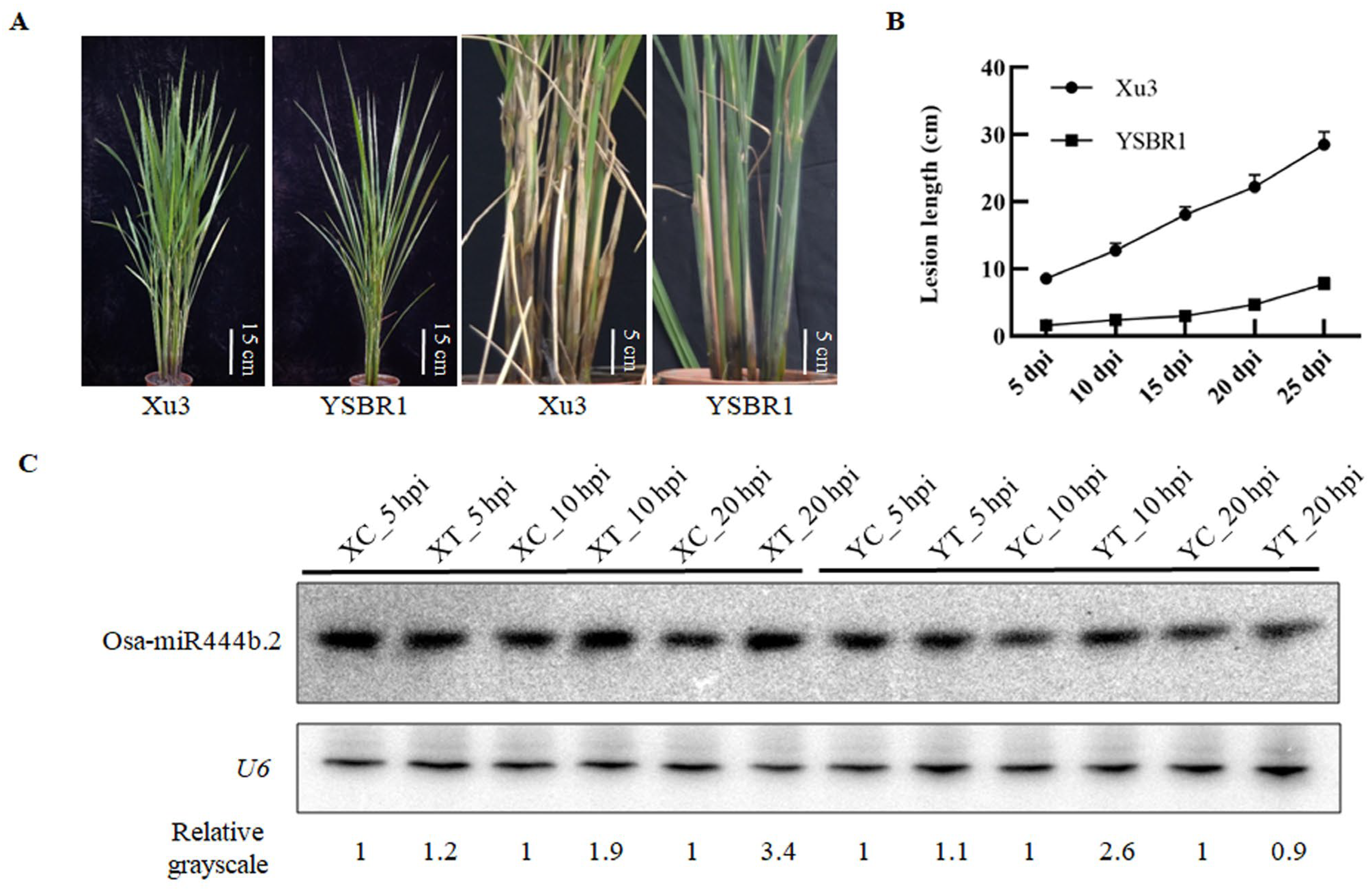
2.1. Characterization of Osa-miR444b.2 in Response to R. solani Infection
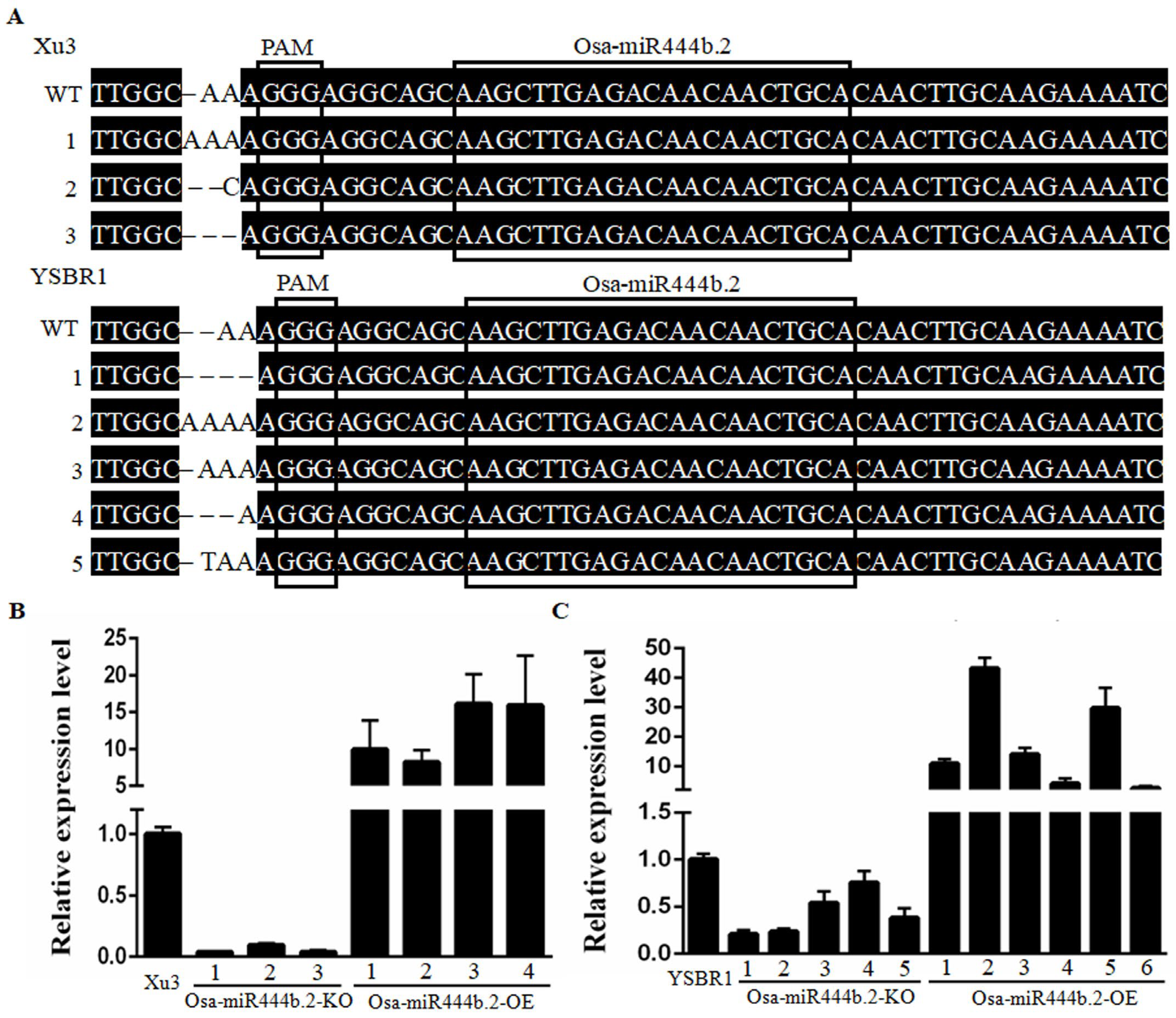
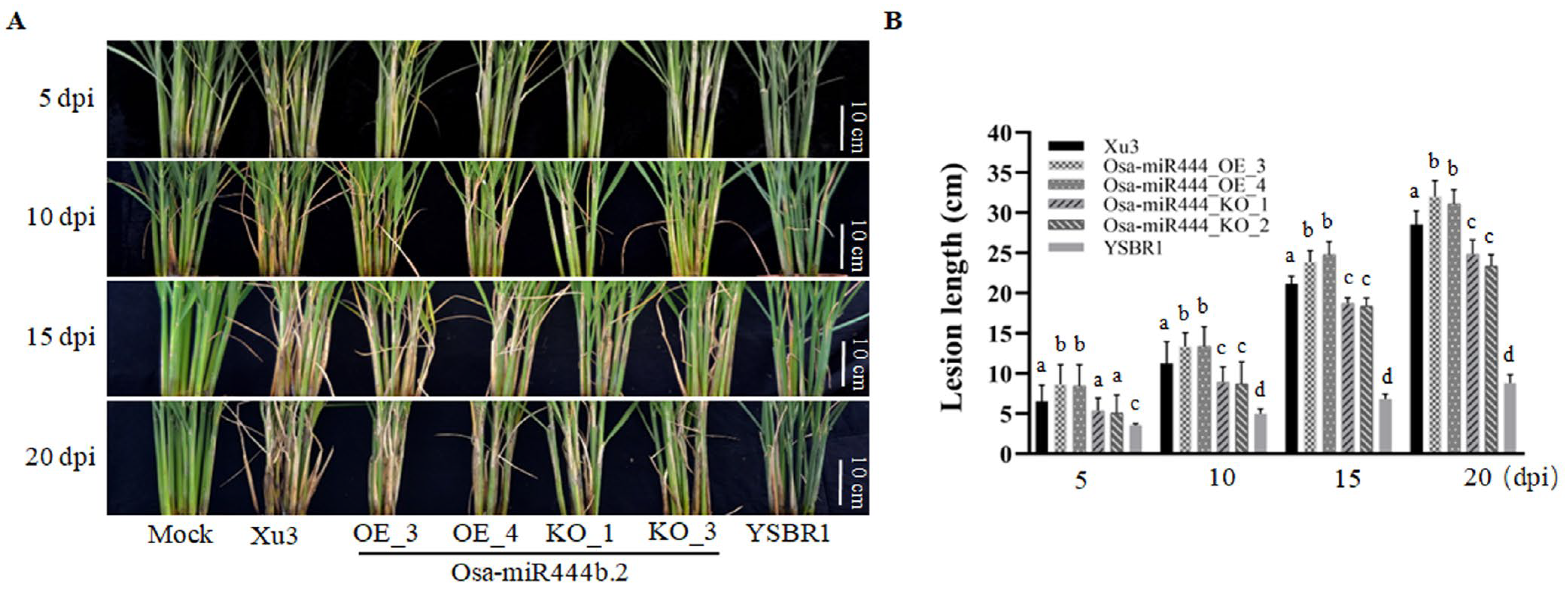
2.2. Overexpressing Osa-miR444b.2 Partially Compromises Sheath Blight Resistance in Rice
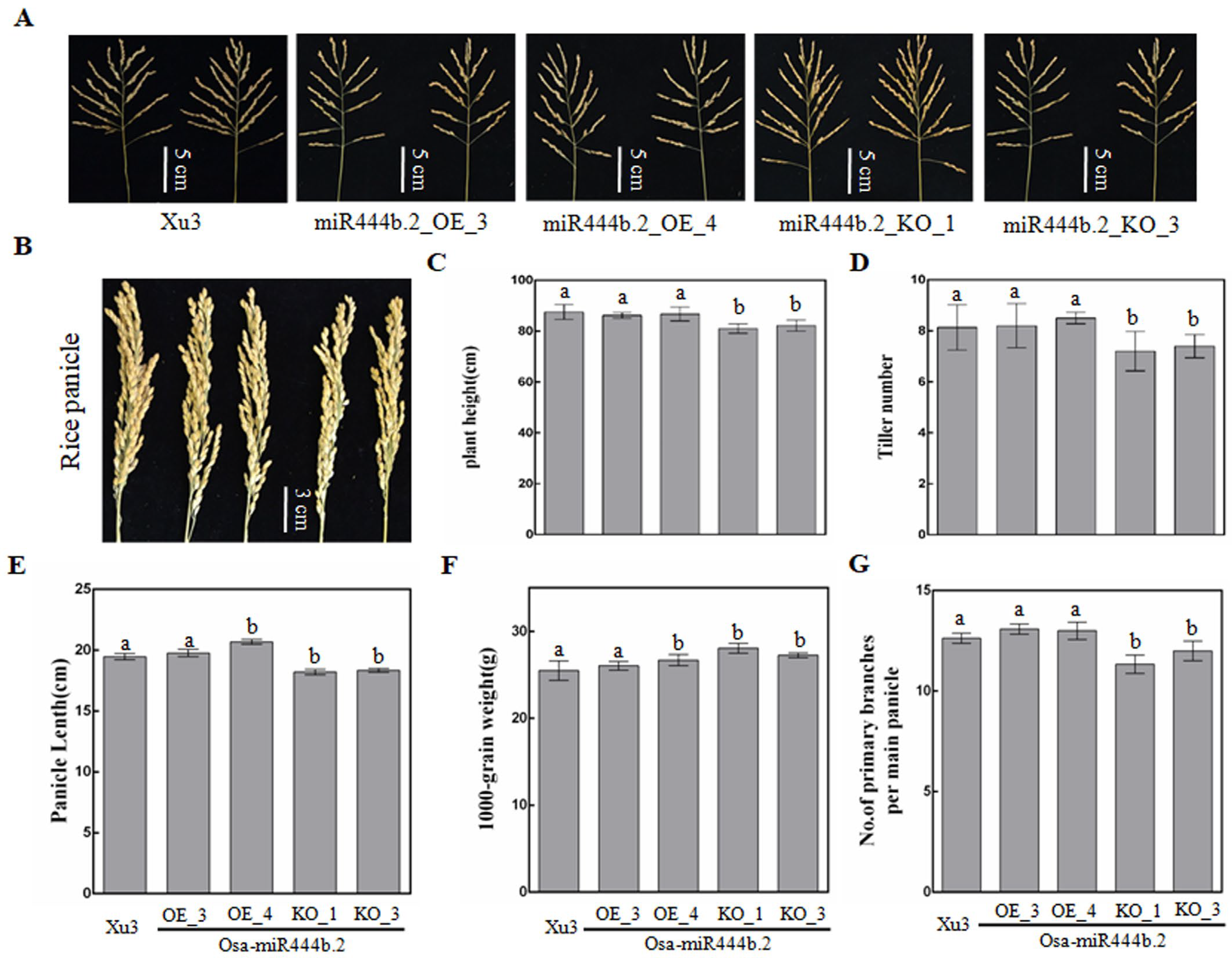
2.3. Osa-miR444b.2 Regulates the Agronomic Traits of Rice
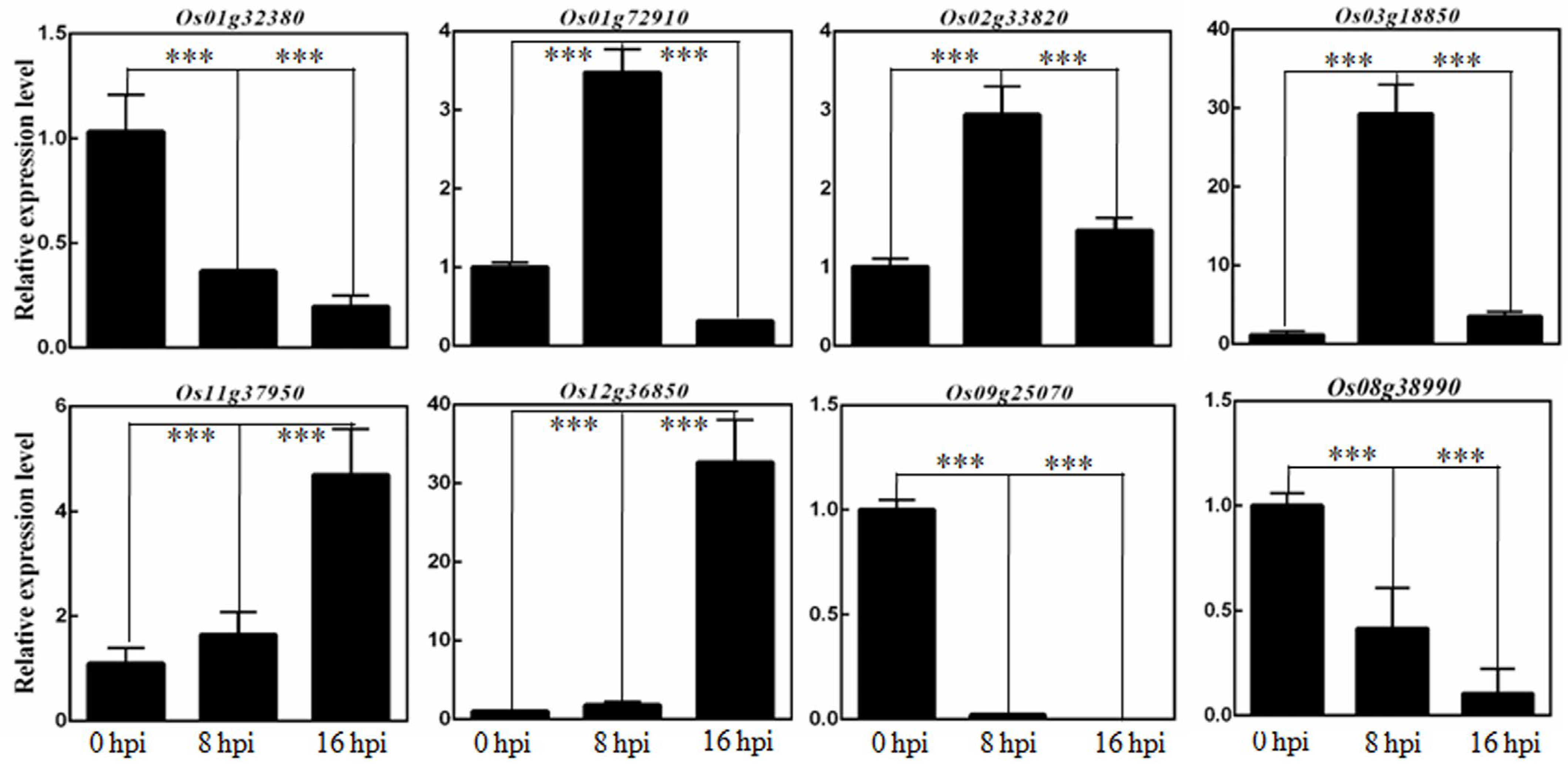
2.4. Osa-miR444b.2 Mediates the Response to R. solani Infection by Affecting Plant Hormone Signal Transduction Pathways and Transcription Factors
3. Discussion
4. Materials and Methods
4.1. Plant Materials and Growth Conditions
4.2. Pathogen Cultivation
4.3. Pathogen Inoculation and Plants Material Collection
4.4. RNA Extraction and Northern Blot Analysis
4.5. Gene Expression Assay
4.6. The CRISPR/Cas9 Plasmid Construct and Osa-miR444b.2 Transgenic Rice Acquisition
4.7. RNA-Seq and Data Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Yuan, M.; Jiang, Z.; Bi, G.; Nomura, K.; Liu, M.; Wang, Y.; Cai, B.; Zhou, J.M.; He, S.Y.; Xin, X.F. Pattern-recognition receptors are required for NLR-mediated plant immunity. Nature 2021, 592, 105–109. [Google Scholar] [CrossRef] [PubMed]
- Han, L.B.; Li, Y.B.; Wang, F.X.; Wang, W.Y.; Liu, J.; Wu, J.H.; Zhong, N.Q.; Wu, S.J.; Jiao, G.L.; Wang, H.Y.; et al. The cotton apoplastic protein CRR1 stabilizes chitinase 28 to facilitate defense against the fungal pathogen Verticillium dahlia. Plant Cell 2019, 31, 520–536. [Google Scholar] [CrossRef] [PubMed]
- Poonguzhali, P.; Chauhan, A.; Kar, A.; Lavale, S.; Nayak, S.N.; Prashanthi, S.K. New Sources of Resistance and Identification of DNA Marker Loci for Sheath Blight Disease Caused by Rhizoctonia solani Kuhn, in Rice. Plant Pathol. J. 2022, 38, 572–582. [Google Scholar] [CrossRef] [PubMed]
- Su’udi, M.; Park, J.M.; Kang, W.R.; Hwang, D.J.; Kim, S.; Ahn, I.P. Quantification of rice sheath blight progression caused by Rhizoctonia solani. J. Microbiol. 2013, 51, 380–388. [Google Scholar] [CrossRef]
- Yu, Y.Y.; Jiang, C.H.; Wang, C.; Chen, L.J.; Li, H.Y.; Xu, Q.; Guo, J.H. An improved strategy for stable biocontrol agents selecting to control rice sheath blight caused by Rhizoctonia solani. Microbiol. Res. 2017, 203, 1–9. [Google Scholar] [CrossRef]
- Iwakawa, H.O.; Tomari, Y. The functions of microRNAs: mRNA decay and translational repression. Trends Cell Biol. 2015, 25, 651–665. [Google Scholar] [CrossRef]
- Navarro, L.; Dunoyer, P.; Jay, F.; Arnold, B.; Dharmasiri, N.; Estelle, M.; Voinnet, O.; Jones, J.D. A plant miRNA contributes to antibacterial resistance by repressing auxin signaling. Science 2006, 312, 436–439. [Google Scholar] [CrossRef]
- Paul, P.; Chhapekar, S.S.; Rameneni, J.J.; Oh, S.H.; Dhandapani, V.; Subburaj, S.; Shin, S.Y.; Ramchiary, N.; Shin, C.; Choi, S.R.; et al. MiR1885 Regulates Disease Tolerance Genes in Brassica rapa during Early Infection with Plasmodiophora brassicae. Int. J. Mol Sci. 2021, 22, 9433. [Google Scholar] [CrossRef]
- Ji, H.M.; Zhao, M.; Gao, Y.; Cao, X.X.; Mao, H.Y.; Zhou, Y.; Fan, W.Y.; Borkovich, K.A.; Ouyang, S.Q.; Liu, P. FRG3, a Target of slmiR482e-3p, Provides Resistance against the Fungal Pathogen Fusarium oxysporum in Tomato. Front. Plant Sci. 2018, 9, 26. [Google Scholar] [CrossRef]
- Li, Y.; Li, T.T.; He, X.R.; Zhu, Y.; Feng, Q.; Yang, X.M.; Zhou, X.H.; Li, G.B.; Ji, Y.P.; Zhao, J.H.; et al. Blocking Osa-miR1871 enhances rice resistance against Magnapotheoryzae and yield. Plant Biotechnol. J. 2022, 20, 646–659. [Google Scholar] [CrossRef]
- Li, Y.; Lu, Y.G.; Shi, Y.; Wu, L.; Xu, Y.J.; Huang, F.; Guo, X.Y.; Zhang, Y.; Fan, J.; Zhao, J.Q.; et al. Multiple rice microRNAs are involved in immunity against the blast fungus Magnaportheoryzae. Plant Physiol. 2014, 164, 1077–1092. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.; Yang, R.; Yang, Z.; Yao, S.; Zhao, S.; Wang, Y.; Li, P.; Song, X.; Jin, L.; Zhou, T.; et al. ROS accumulation and antiviral defence control by microRNA528 in rice. Nat. Plants 2017, 3, 16203. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Cao, X.L.; Zhu, Y.; Yang, X.M.; Zhang, K.N.; Xiao, Z.Y.; Wang, H.; Zhao, J.H.; Zhang, L.L.; Li, G.B.; et al. Osa-miR398b boosts H2O2 production and rice blast disease-resistance via multiple superoxide dismutases. New Phytol. 2019, 222, 1507–1522. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.Y.; Xia, Y.Q.; Lin, S.Y.; Wang, Y.R.; Guo, B.H.; Song, X.N.; Ding, S.C.; Zheng, L.Y.; Feng, R.Y.; Chen, S.L.; et al. Osa-miR164a targets OsNAC60 and negatively regulates rice immunity against the blast fungus Magnaportheoryzae. Plant J. 2018, 95, 584–597. [Google Scholar] [CrossRef]
- Qiao, L.L.; Zheng, L.Y.; Sheng, C.; Zhao, H.W.; Jin, H.L.; Niu, D.D. Rice siR109944 suppresses plant immunity to sheath blight and impacts multiple agronomic traits by affecting auxin homeostasis. Plant J. 2020, 102, 948–964. [Google Scholar] [CrossRef]
- Cao, W.L.; Cao, X.X.; Zhao, J.H.; Zhang, Z.Y.; Feng, Z.M.; Ouyang, S.Q.; Zuo, S.M. Comprehensive characteristics of microRNA expression profile conferring to Rhizoctonia solani in rice. Rice Sci. 2020, 27, 101–112. [Google Scholar]
- Zhang, C.; Ding, Z.M.; Wu, K.C.; Yang, L.; Li, Y.; Yang, Z.; Shi, S.; Liu, X.J.; Zhao, S.S.; Yang, Z.R. Suppression of Jasmonic Acid-mediated defense by viral-inducible microRNA319 facilitates virus infection in rice. Mol. Plant 2016, 9, 1302–1314. [Google Scholar] [CrossRef]
- Yu, C.; Chen, Y.T.; Cao, Y.Q.; Chen, H.M.; Wang, J.C.; Bi, Y.M.; Tian, F.; Yang, F.H.; Rothstein, S.J.; Zhou, X.P. Overexpression of miR169o, an overlapping microRNA in response to both nitrogen limitation and bacterial infection, promotes nitrogen use efficiency and susceptibility to bacterial blight in rice. Plant Cell Physiol. 2018, 59, 1234–1247. [Google Scholar] [CrossRef]
- Jiao, X.; Wang, H.; Yan, J.; Kong, X.; Liu, Y.; Chu, J.; Chen, X.; Fang, R.; Yan, Y. Promotion of BR biosynthesis by miR444 is required for ammonium triggered inhibition of root growth. Plant Physiol. 2020, 182, 1454–1466. [Google Scholar] [CrossRef]
- Wu, L.; Zhang, Q.Q.; Zhou, H.Y.; Ni, F.R.; Wu, X.Y.; Qi, Y.J. Rice MicroRNA effector complexes and targets. Plant Cell 2009, 21, 3421–3435. [Google Scholar] [CrossRef]
- Zhou, R.; Sanz-Jimenez, P.; Zhu, X.T.; Feng, J.W.; Shao, L.; Song, J.M.; Chen, L.L. Analysis of rice transcriptome reveals the LncRNA/CircRNA regulation in tissue development. Rice 2021, 28, 14. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Weiberg, A.; Lin, F.M.; Thomma, B.P.; Huang, H.D.; Jin, H. Bidirectional cross-kingdom RNAi and fungal uptake of external RNAs confer plant protection. Nat. Plants 2016, 2, 16151. [Google Scholar] [CrossRef] [PubMed]
- Xiao, Z.Y.; Wang, Q.X.; Wang, H.; Li, J.L.; Zhao, S.L.; Fan, J.; Li, Y.; Wang, W.M. MiR444b.2 regulates resistance to Magnaportheoryzae and tillering in rice. Acta Phytopathol. Sin. 2017, 47, 511–522. [Google Scholar]
- Guo, S.Y.; Xu, Y.Y.; Liu, H.H.; Mao, Z.W.; Zhang, C.; Ma, Y.; Zhang, Q.R.; Meng, Z.; Chong, K. The interaction between OsMADS57 and OsTB1 modulates rice tillering via DWARF14. Nat. Commun. 2013, 4, 1566. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.S.; Lee, D.Y.; Cho, L.H.; An, G. Rice miR172 induces flowering by suppressing OsIDS1 and SNB, two AP2 genes that negatively regulate expression of Ehd1 and florigens. Rice 2014, 7, 31. [Google Scholar]
- Grant, M.R.; Jones, J.D.G. Hormone (dis)harmony moulds plant health and disease. Science 2009, 324, 750–752. [Google Scholar] [CrossRef]
- Zhu, Q.H.; Upadhyaya, N.M.; Gubler, F.; Helliwell, C.A. Over-expression of miR172 causes loss of spikelet determinacy and floral organ abnormalities in rice (Oryza sativa). BMC Plant Biol. 2019, 9, 149. [Google Scholar]
- Yang, D.; Yang, Y.; He, Z. Roles of plant hormones and their interplay in rice immunity. Mol. Plant 2013, 6, 675–685. [Google Scholar]
- Amorim, L.L.B.; da Fonseca Dos Santos, R.; Neto, J.P.B.; Guida-Santos, M.; Crovella, S.; Benko-Iseppon, A.M. Transcriptionfactors involved in plant resistance to pathogens. Curr. Protein Pept. Sci. 2017, 18, 335–351. [Google Scholar] [CrossRef]
- Xia, K.; Wang, R.; Ou, X.; Fang, Z.; Tian, C.; Duan, J.; Wang, Y.; Zhang, M. OsTIR1 and OsAFB2 downregulation via OsmiR393 overexpression leads to more tillers, early flowering and less tolerance to salt and drought in rice. PLoS ONE 2012, 7, e30039. [Google Scholar] [CrossRef]
- Ma, X.L.; Zhang, Q.Y.; Zhu, Q.L.; Liu, W.; Chen, Y.; Qiu, R.; Wang, B.; Yang, Z.F.; Li, H.Y.; Lin, Y.R. A robust CRISPR/Cas9 system for convenient, high-efficiency multiplex genome editing in monocot and dicot plants. Mol. Plant 2015, 8, 1274–1284. [Google Scholar] [CrossRef] [PubMed]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Feng, T.; Zhang, Z.-Y.; Gao, P.; Feng, Z.-M.; Zuo, S.-M.; Ouyang, S.-Q. Suppression of Rice Osa-miR444.2 Improves the Resistance to Sheath Blight in Rice Mediating through the Phytohormone Pathway. Int. J. Mol. Sci. 2023, 24, 3653. https://doi.org/10.3390/ijms24043653
Feng T, Zhang Z-Y, Gao P, Feng Z-M, Zuo S-M, Ouyang S-Q. Suppression of Rice Osa-miR444.2 Improves the Resistance to Sheath Blight in Rice Mediating through the Phytohormone Pathway. International Journal of Molecular Sciences. 2023; 24(4):3653. https://doi.org/10.3390/ijms24043653
Chicago/Turabian StyleFeng, Tao, Zhao-Yang Zhang, Peng Gao, Zhi-Ming Feng, Shi-Min Zuo, and Shou-Qiang Ouyang. 2023. "Suppression of Rice Osa-miR444.2 Improves the Resistance to Sheath Blight in Rice Mediating through the Phytohormone Pathway" International Journal of Molecular Sciences 24, no. 4: 3653. https://doi.org/10.3390/ijms24043653