
Anti-Atherogenic Actions of the Lab4b Consortium of Probiotics In Vitro

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Lab4b CM Has No Detrimental Effect on Cell Viability of Human Macrophages and VSMC
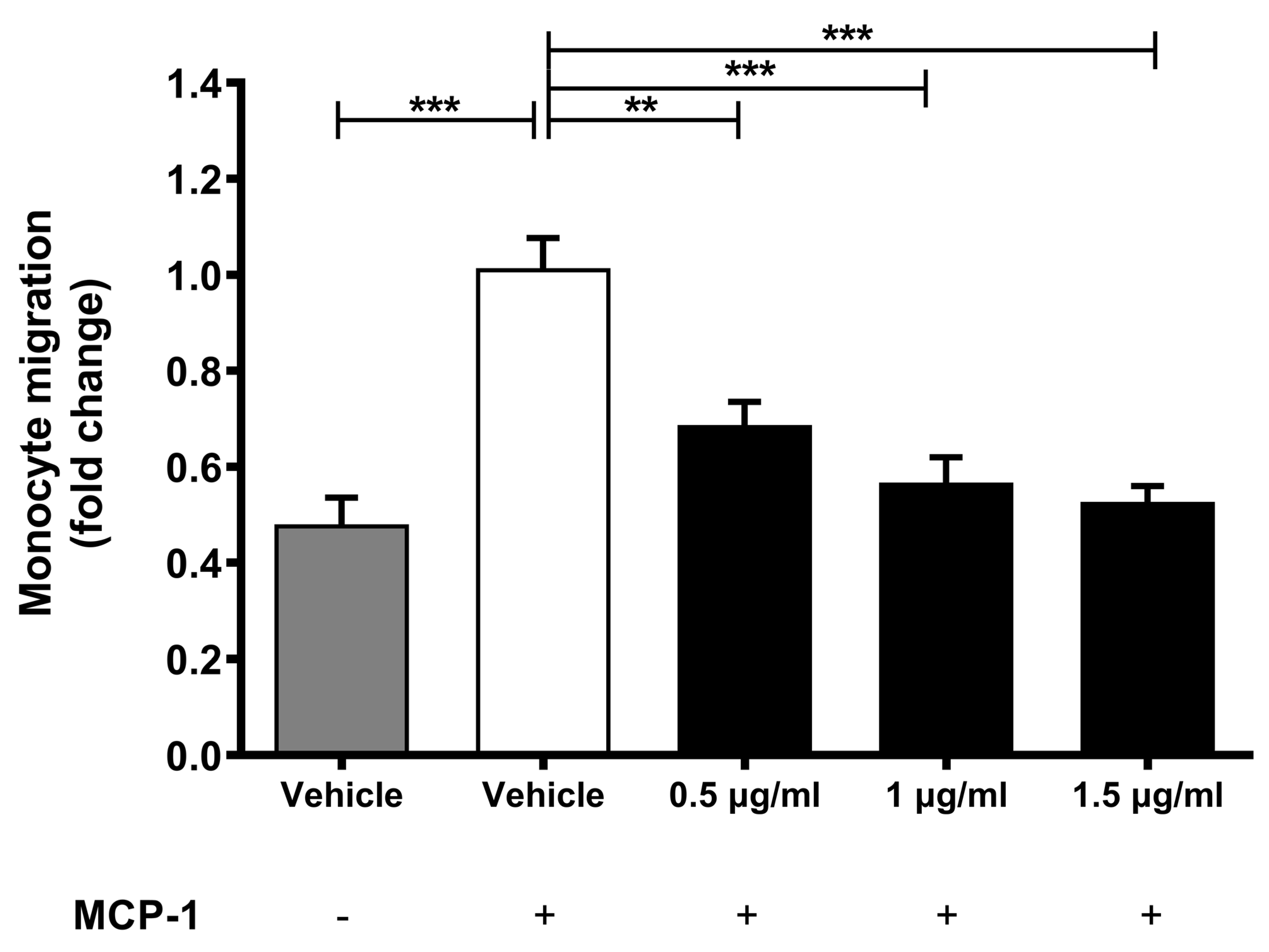
2.2. Lab4b Attenuates Chemokine-Driven Monocytic Migration
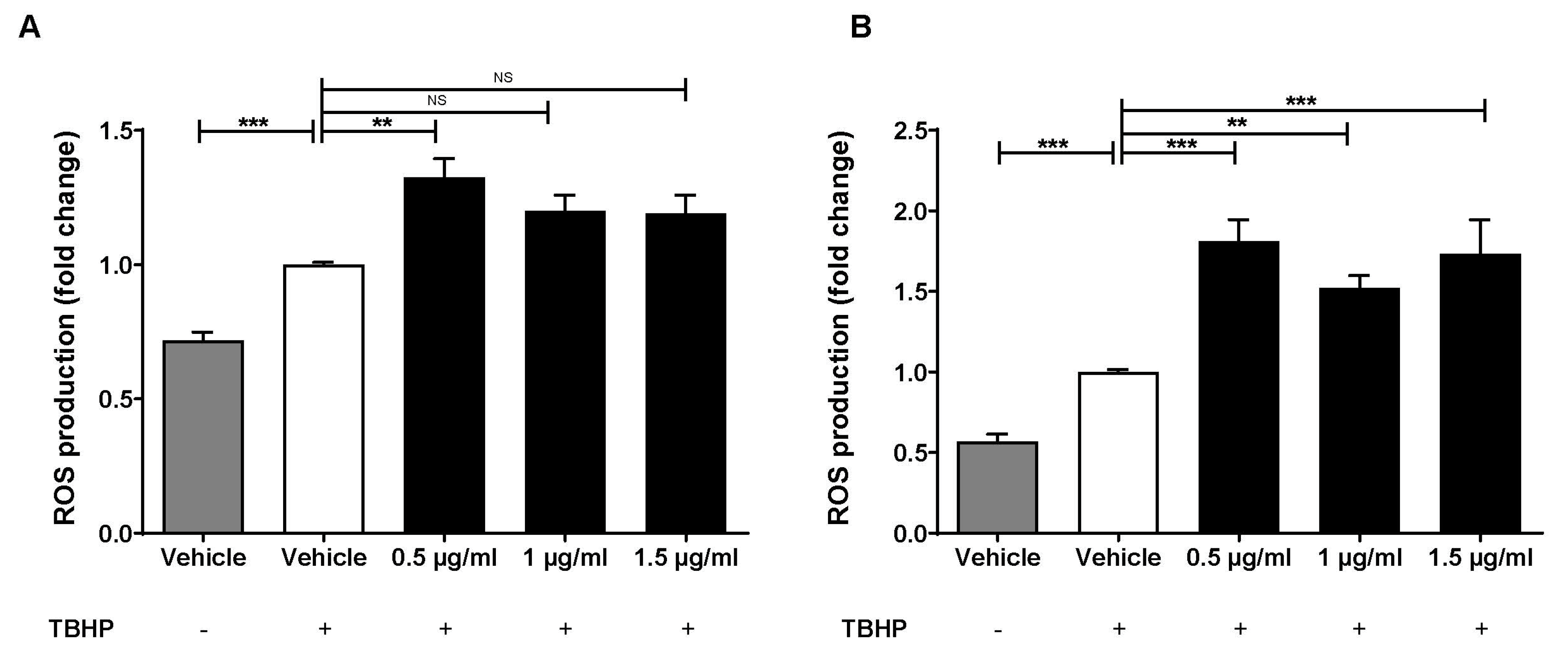
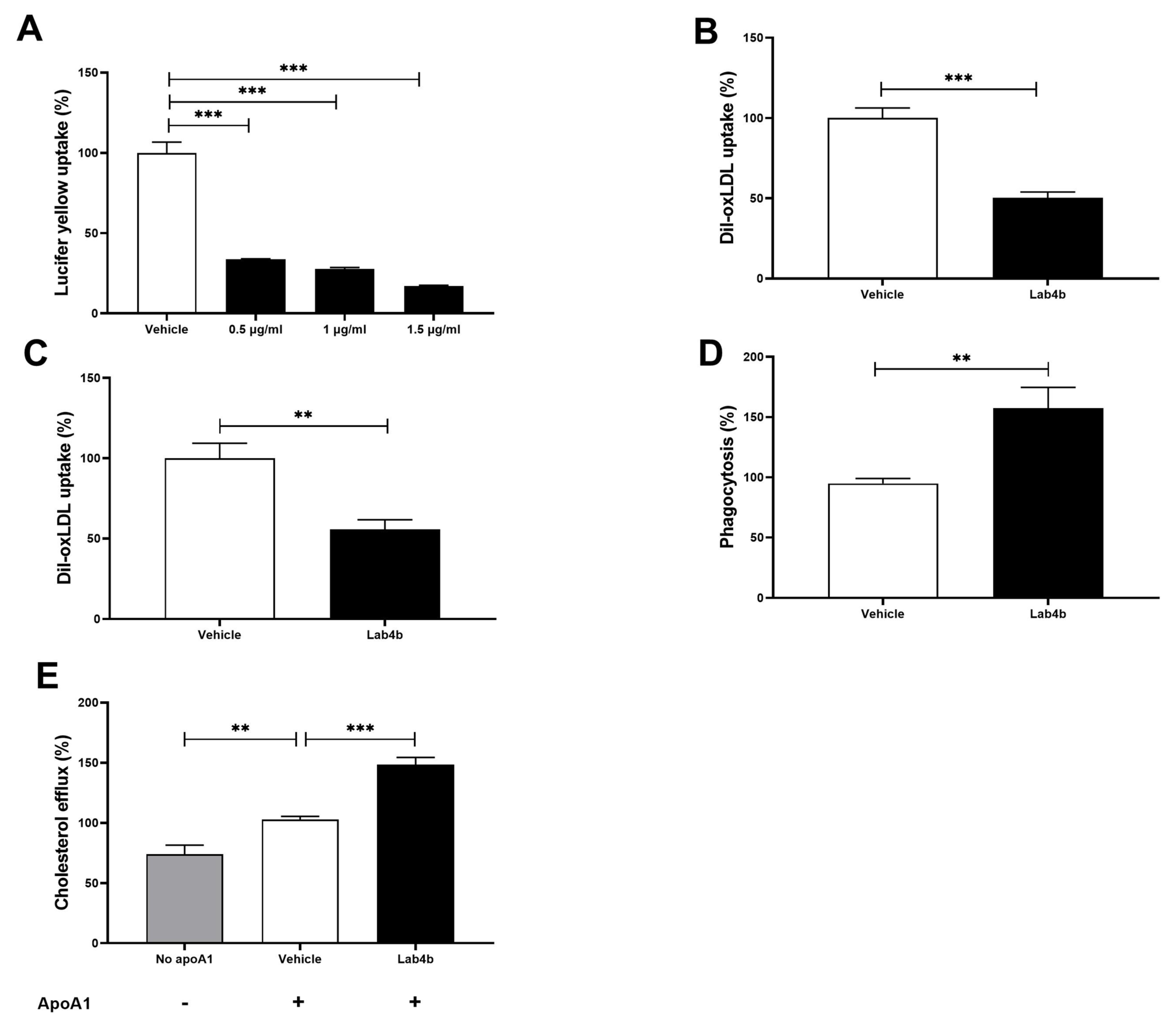
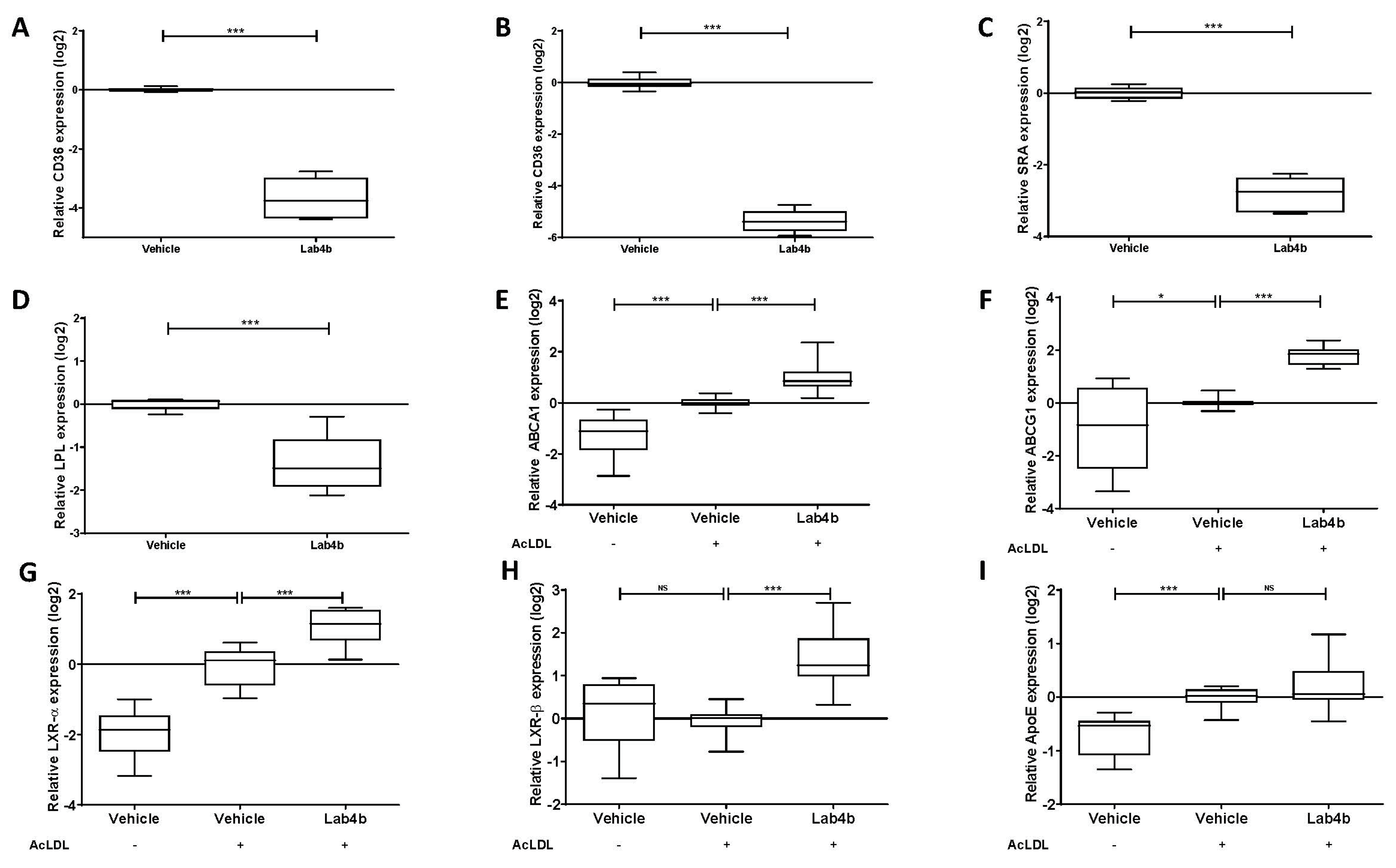
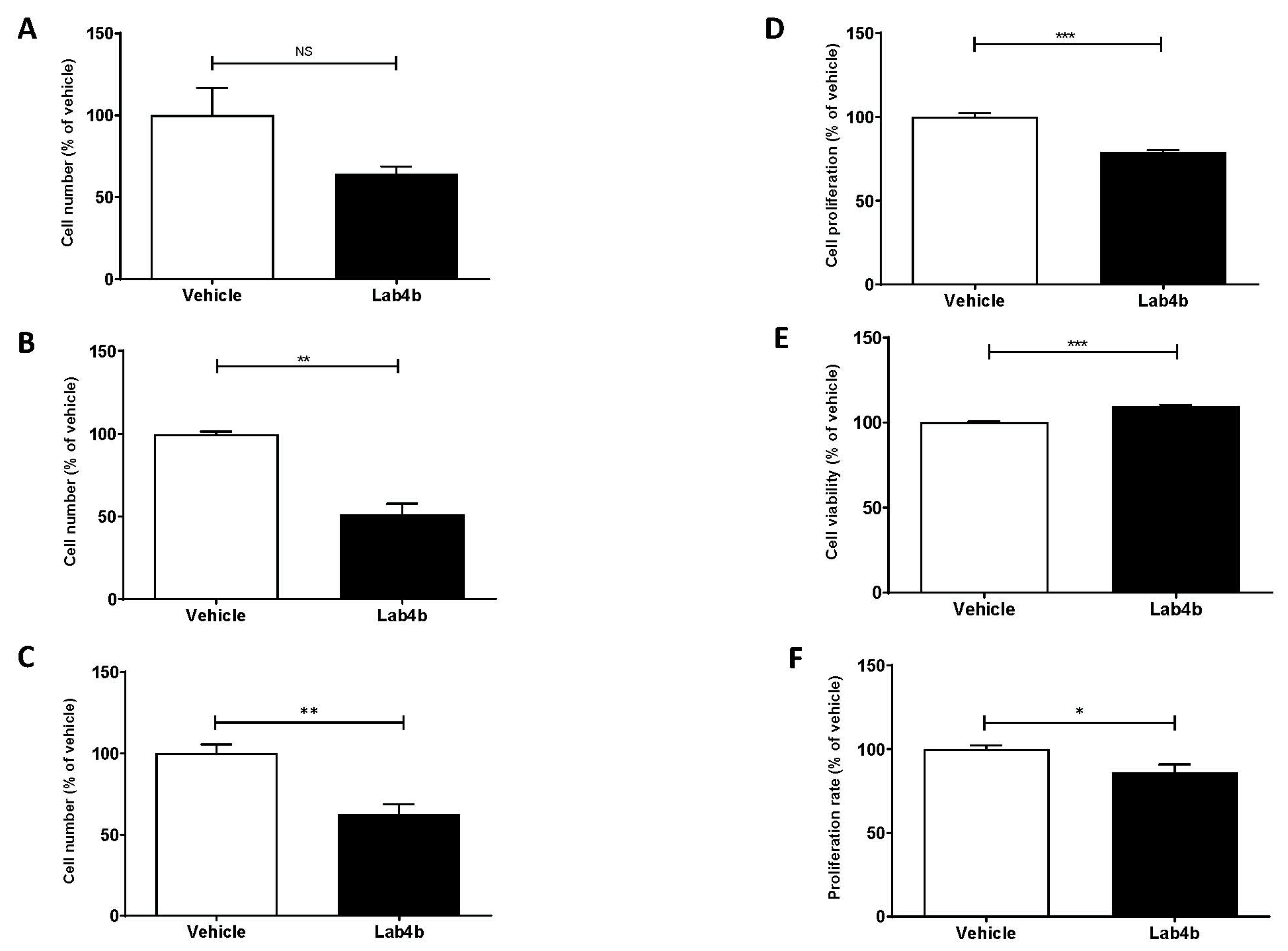
2.3. Lab4b CM Has Beneficial Actions on Several Parameters Associated with Macrophage Foam Cell Formation
2.4. The Effects of Lab4b CM on Monocyte/Macrophage Proliferation
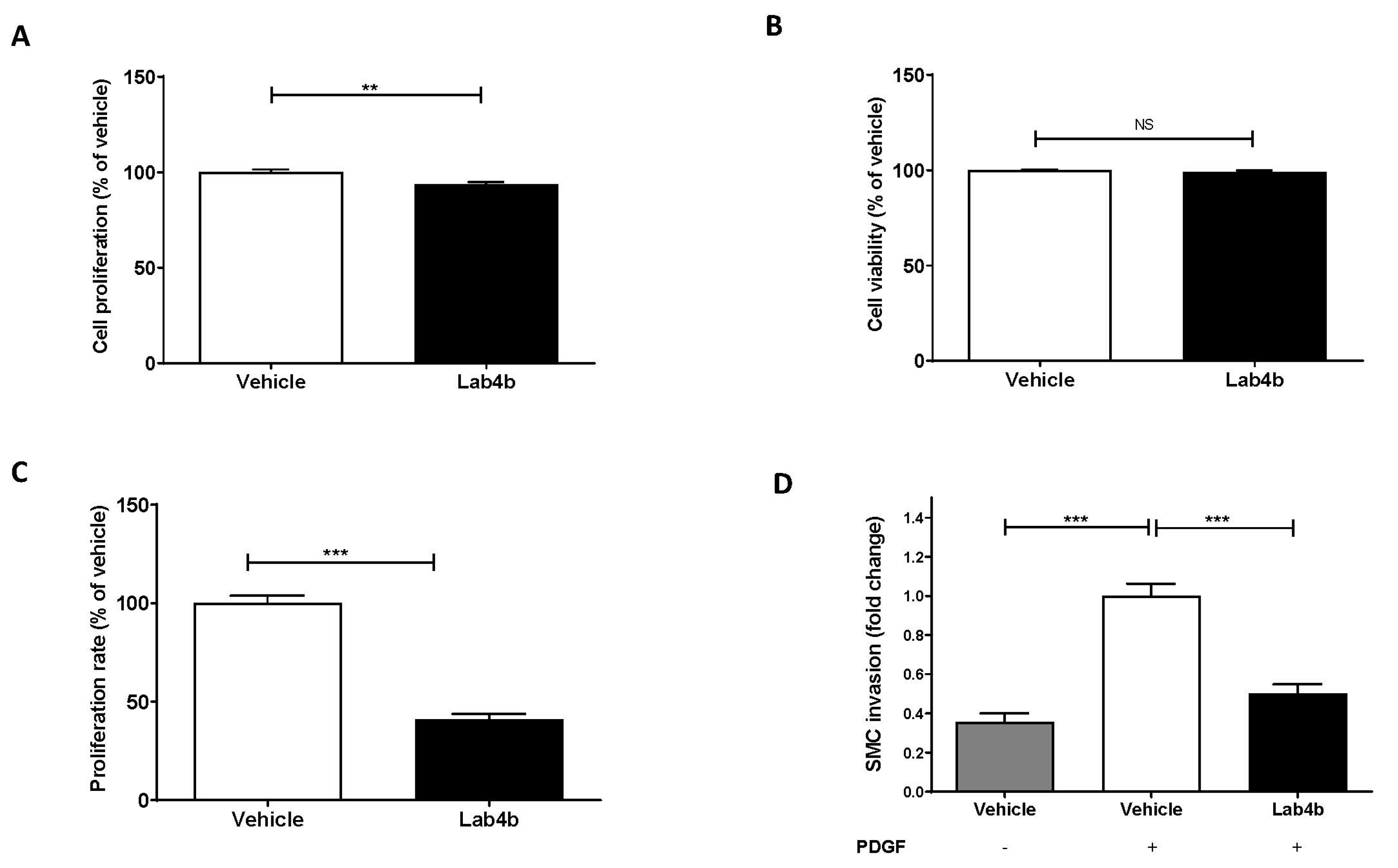
2.5. Lab4b CM Attenuates HASMC Proliferation and Platelet-Derived Growth Factor (PDGF)-Induced Migration
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Materials
5.2. Preparation of Lab4b CM
5.3. Cell Culture and In Vitro Assays
5.4. Gene Expression
5.5. Statistical Analysis of Data
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Moss, J.W.; Ramji, D.P. Nutraceutical therapies for atherosclerosis. Nat. Rev. Cardiol. 2016, 13, 513–532. [Google Scholar] [CrossRef]
- Chan, Y.H.; Ramji, D.P. A perspective on targeting inflammation and cytokine actions in atherosclerosis. Future Med. Chem. 2020, 12, 613–626. [Google Scholar] [CrossRef] [PubMed]
- O’Morain, V.L.; Ramji, D.P. The Potential of Probiotics in the Prevention and Treatment of Atherosclerosis. Mol. Nutr. Food Res. 2020, 64, e1900797. [Google Scholar] [CrossRef] [PubMed]
- Moss, J.W.E.; Williams, J.O.; Ramji, D.P. Nutraceuticals as therapeutic agents for atherosclerosis. Biochim. Biophys. Acta 2018, 1864, 1562–1572. [Google Scholar] [CrossRef]
- Michael, D.R.; Davies, T.S.; Moss, J.W.E.; Calvente, D.L.; Ramji, D.P.; Marchesi, J.R.; Pechlivanis, A.; Plummer, S.F.; Hughes, T.R. The anti-cholesterolaemic effect of a consortium of probiotics: An acute study in C57BL/6J mice. Sci. Rep. 2017, 7, 2883. [Google Scholar] [CrossRef]
- O’Morain, V.L.; Chan, Y.H.; Williams, J.O.; Alotibi, R.; Alahmadi, A.; Rodrigues, N.P.; Plummer, S.F.; Hughes, T.R.; Michael, D.R.; Ramji, D.P. The Lab4P consortium of probiotics attenuates atherosclerosis in LDL receptor deficient mice fed a high fat diet and causes plaque stabilization by inhibiting inflammation and several pro-atherogenic processes. Mol. Nutr. Food Res. 2021, 65, e2100214. [Google Scholar] [CrossRef] [PubMed]
- Allen, S.J.; Jordan, S.; Storey, M.; Thornton, C.A.; Gravenor, M.; Garaiova, I.; Plummer, S.F.; Wang, D.; Morgan, G. Dietary supplementation with lactobacilli and bifidobacteria is well tolerated and not associated with adverse events during late pregnancy and early infancy. J. Nutr. 2010, 140, 483–488. [Google Scholar] [CrossRef]
- Allen, S.J.; Jordan, S.; Storey, M.; Thornton, C.A.; Gravenor, M.B.; Garaiova, I.; Plummer, S.F.; Wang, D.; Morgan, G. Probiotics in the prevention of eczema: A randomised controlled trial. Arch. Dis. Child. 2014, 99, 1014–1019. [Google Scholar] [CrossRef] [PubMed]
- Michael, D.R.; Davies, T.S.; Loxley, K.E.; Allen, M.D.; Good, M.A.; Hughes, T.R.; Plummer, S.F. Neuroprotective activities of two distinct probiotic consortia. Benef. Microbes 2019, 10, 437–447. [Google Scholar] [CrossRef]
- Mclaren, J.E.; Michael, D.R.; Salter, R.C.; Ashlin, T.G.; Calder, C.J.; Miller, A.M.; Liew, F.Y.; Ramji, D.P. IL-33 Reduces Macrophage Foam Cell Formation. J. Immunol. 2010, 185, 1222–1229. [Google Scholar] [CrossRef]
- Gallagher, H.; Williams, J.O.; Ferekidis, N.; Ismail, A.; Chan, Y.H.; Michael, D.R.; Guschina, I.A.; Tyrrell, V.J.; O’Donnell, V.B.; Harwood, J.L.; et al. Dihomo-γ-linolenic acid inhibits several key cellular processes associated with atherosclerosis. Biochim. Biophys. Acta. 2019, 1865, 2538–2550. [Google Scholar] [CrossRef]
- McLaren, J.E.; Michael, D.R.; Ashlin, T.G.; Ramji, D.P. Cytokines, macrophage lipid metabolism and foam cells: Implications for cardiovascular disease therapy. Prog. Lipid Res. 2011, 50, 331–347. [Google Scholar] [CrossRef]
- Rosenfeld, M.E. Macrophage proliferation in atherosclerosis: An historical perspective. Arter. Thromb. Vasc. Biol. 2014, 34, e21–e22. [Google Scholar] [CrossRef]
- Vivarelli, S.; Falzone, L.; Basile, M.S.; Nicolosi, D.; Genovese, C.; Libra, M.; Salmeri, M. Benefits of using probiotics as adjuvants in anti cancer therapy. World Acad. Sci. J. 2019, 1, 125–135. [Google Scholar]
- Wu, H.; Chiou, J. Potential benefits of probiotics and prebiotics for coronary heart disease and stroke. Nutrients 2021, 13, 2878. [Google Scholar] [CrossRef]
- Xiang, S.; Ji, J.-L.; Li, S.; Cao, X.-P.; Xu, W.; Tan, L.; Tan, C.-C. Efficacy and safety of probiotics for the treatment of Azheimer’s disease, mild cognitive impairment, and Parkinson’s Disease: A systematic review and meta analysis. Front. Aging Neurosci. 2022, 14, 730036. [Google Scholar] [CrossRef]
- Thushara, R.M.; Gangadaran, S.; Solati, Z.; Moghadasian, G. Cardiovascular benefits of probiotics: A review of experimental and clinical studies. Food Funct. 2016, 7, 632–642. [Google Scholar] [CrossRef]
- Yu, E.; Mercer, J.; Bennett, M. Mitochondria in vascular disease. Cardiovasc. Res. 2012, 95, 173–182. [Google Scholar] [CrossRef]
- Zuo, J.; Zhang, Z.; Luo, M.; Zhou, L.; Nice, E.C.; Zhang, W.; Wang, C.; Huang, C. Redox signaling at the crossroads of human health and disease. MedComm 2022, 3, e127. [Google Scholar] [CrossRef]
- Noh, H.J.; Park, J.M.; Kwon, Y.J.; Kim, K.; Park, S.Y.; Kim, I.; Lim, J.H.; Kim, B.K.; Kim, B.Y. Immunostimulatory Effect of Heat-Killed Probiotics on RAW264.7 Macrophages. J. Microbiol. Biotechnol. 2022, 32, 638–644. [Google Scholar] [CrossRef]
- Jaffar, N.; Okinaga, T.; Nishihara, T.; Maeda, T. Enhanced phagocytosis of Aggregatibacter actinomycetemcomitans cells by macrophages activated by a probiotic Lactobacillus strain. J. Dairy Sci. 2018, 101, 5789–5798. [Google Scholar] [CrossRef] [PubMed]
- Deng, H.; Li, Z.; Tan, Y.; Guo, Z.; Liu, Y.; Wang, Y.; Yuan, Y.; Yang, R.; Bi, Y.; Bai, Y.; et al. A novel strain of Bacteroides fragilis enhances phagocytosis and polarises M1 macrophages. Sci. Rep. 2016, 6, 29401. [Google Scholar] [CrossRef] [PubMed]
- Laguna-Fernandez, A.; Checa, A.; Carracedo, M.; Artiach, G.; Petri, M.H.; Baumgartner, R.; Forteza, M.J.; Jiang, X.; Andonova, T.; Walker, M.E.; et al. ERV1/ChemR23 signaling protects against atherosclerosis by modifying oxidized low-density lipoprotein uptake and phagocytosis in macrophages. Circulation 2018, 138, 1693–1705. [Google Scholar] [CrossRef] [PubMed]
- Schrijvers, D.M.; De Meyer, G.R.; Herman, A.G.; Martinet, W. Phagocytosis in atherosclerosis: Molecular mechanisms and implications for plaque progression and stability. Cardiovasc. Res. 2007, 73, 470–480. [Google Scholar] [CrossRef] [PubMed]
- Flynn, M.C.; Pernes, G.; Lee, M.K.S.; Nagareddy, P.R.; Murphy, A.J. Monocytes, macrophages, and metabolic disease in atherosclerosis. Front. Pharmacol. 2019, 10, 666. [Google Scholar] [CrossRef]
- Robbins, C.S.; Hilgendorf, I.; Weber, G.F.; Theurl, I.; Iwamoto, Y.; Figueiredo, J.L.; Gorbatov, R.; Sukhova, G.K.; Gerhardt, L.M.; Smyth, D.; et al. Local proliferation dominates lesional macrophage accumulation in atherosclerosis. Nat. Med. 2013, 19, 1166–1172. [Google Scholar] [CrossRef]
- Lhoták, Š.; Gyulay, G.; Cutz, J.C.; Al-Hashimi, A.; Trigatti, B.L.; Richards, C.D.; Igdoura, S.A.; Steinberg, G.R.; Bramson, J.; Ask, K.; et al. Characterization of proliferating lesion-resident cells during all stages of atherosclerotic growth. J. Am. Heart Assoc. 2016, 5, e003945. [Google Scholar] [CrossRef]
- Kankaanpää, P.E.; Yang, B.; Kallio, H.P.; Isolauri, E.; Salminen, S.J. Influence of probiotic supplemented infant formula on composition of plasma lipids in atopic infants. J. Nutr. Biochem. 2002, 13, 364–369. [Google Scholar] [CrossRef]
- Bennett, M.R.; Sinha, S.; Owens, G.K. Vascular smooth muscle cells in atherosclerosis. Circ. Res. 2016, 118, 692–702. [Google Scholar] [CrossRef]
- Ricci, C.; Ferri, N. Naturally occurring PDGF receptor inhibitors with potential anti-atherosclerotic properties. Vascul. Pharmacol. 2015, 70, 1–7. [Google Scholar] [CrossRef]
- Sano, H.; Sudo, T.; Yokode, M.; Murayama, T.; Kataoka, H.; Takakura, N.; Nishikawa, S.; Nishikawa, S.I.; Kita, T. Functional blockade of platelet-derived growth factor receptor-beta but not of receptor-alpha prevents vascular smooth muscle cell accumulation in fibrous cap lesions in apolipoprotein E-deficient mice. Circulation 2001, 103, 2955–2960. [Google Scholar] [CrossRef]
- Wang, T.M.; Chen, K.C.; Hsu, P.Y.; Lin, H.F.; Wang, Y.S.; Chen, C.Y.; Liao, Y.C.; Juo, S.H. microRNA let-7g suppresses PDGF-induced conversion of vascular smooth muscle cell into the synthetic phenotype. J. Cell Mol. Med. 2017, 21, 3592–3601. [Google Scholar] [CrossRef]
- Lassila, M.; Allen, T.J.; Cao, Z.; Thallas, V.; Jandeleit-Dahm, K.A.; Candido, R.; Cooper, M.E. Imatinib attenuates diabetes-associated atherosclerosis. Arter. Thromb. Vasc. Biol. 2004, 24, 935–942. [Google Scholar] [CrossRef]
- Buckley, M.L.; Ramji, D.P. The influence of dysfunctional signaling and lipid homeostasis in mediating the inflammatory responses during atherosclerosis. Biochim. Biophys. Acta 2015, 1852, 1498–1510. [Google Scholar] [CrossRef]
- Mead, J.R.; Ramji, D.P. The pivotal role of lipoprotein lipase in atherosclerosis. Cardiovasc. Res. 2002, 55, 261–269. [Google Scholar] [CrossRef]
- Serri, O.; Li, L.; Maingrette, F.; Jaffry, N.; Renier, G. Enhanced lipoprotein lipase secretion and foam cell formation by macrophages of patients with growth hormone deficiency: Possible contribution to increased risk of atherogenesis? J. Clin. Endocrinol. Metab. 2004, 89, 979–985. [Google Scholar] [CrossRef]
- Michael, D.R.; Ashlin, T.G.; Buckley, M.L.; Ramji, D.P. Liver X receptors, atherosclerosis and inflammation. Curr. Atheroscler. Rep. 2012, 14, 284–293. [Google Scholar] [CrossRef]
- Jin, P.; Bian, Y.; Wang, K.; Cong, G.; Yan, R.; Sha, Y.; Ma, X.; Zhou, J.; Yuan, Z.; Jia, S. Homocysteine accelerates atherosclerosis via inhibiting LXRα-mediated ABCA1/ABCG1-dependent cholesterol efflux from macrophages. Life Sci. 2018, 214, 41–50. [Google Scholar] [CrossRef]
- Larrede, S.; Quinn, C.M.; Jessup, W.; Frisdal, E.; Olivier, M.; Hsieh, V.; Kim, M.J.; Van Eck, M.; Couvert, P.; Carrie, A.; et al. Stimulation of cholesterol efflux by LXR agonists in cholesterol-loaded human macrophages is ABCA1-dependent but ABCG1-independent. Arterioscler. Thromb. Vasc. Biol. 2009, 29, 1930–1936. [Google Scholar] [CrossRef]
- Buckley, M.L.; Williams, J.O.; Chan, Y.H.; Laubertová, L.; Gallagher, H.; Moss, J.W.E.; Ramji, D.P. The interleukin-33-mediated inhibition of expression of two key genes implicated in atherosclerosis in human macrophages requires MAP kinase, phosphoinositide 3-kinase and nuclear factor-κB signaling pathways. Sci. Rep. 2019, 9, 11317. [Google Scholar] [CrossRef]
- Moss, J.W.; Davies, T.S.; Garaiova, I.; Plummer, S.F.; Michael, D.R.; Ramji, D.P. A Unique Combination of Nutritionally Active Ingredients Can Prevent Several Key Processes Associated with Atherosclerosis In Vitro. PLoS ONE 2016, 11, e0151057. [Google Scholar] [CrossRef] [PubMed]
- Al-Ahmadi, W.; Webberley, T.S.; Joseph, A.; Harris, F.; Chan, Y.H.; Alotibi, R.; Williams, J.O.; Alahmadi, A.; Decker, T.; Hughes, T.R.; et al. Pro-atherogenic actions of signal transducer and activator of transcription 1 serine 727 phosphorylation in LDL receptor deficient mice via modulation of plaque inflammation. FASEB J. 2021, 35, e21892. [Google Scholar] [CrossRef] [PubMed]
- Takala, R.; Ramji, D.P.; Andrews, R.; Zhou, Y.; Burston, J.; Choy, E. Anti-inflammatory and immunoregulatory effects of pinolenic acid in rheumatoid arthritis. Rheumatology 2022, 61, 992–1004. [Google Scholar] [CrossRef] [PubMed]
- Alahmadi, A.; Ramji, D.P. Monitoring modified lipoprotein uptake and macropinocytosis associated with macrophage foam cell formation. Methods Mol. Biol. 2022, 2419, 247–255. [Google Scholar] [CrossRef]
- Moss, J.W.E.; Williams, J.O.; Al-Ahmadi, W.; O’Morain, V.; Chan, Y.H.; Hughes, T.R.; Menendez-Gonzalez, J.B.; Almotiri, A.; Plummer, S.F.; Rodrigues, N.P.; et al. Protective effects of a unique combination of nutritionally active ingredients on risk factors and gene expression associated with atherosclerosis in C57BL/6J mice fed a high fat diet. Food Funct. 2021, 12, 3657–3671. [Google Scholar] [CrossRef]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
O’Morain, V.L.; Chen, J.; Plummer, S.F.; Michael, D.R.; Ramji, D.P. Anti-Atherogenic Actions of the Lab4b Consortium of Probiotics In Vitro. Int. J. Mol. Sci. 2023, 24, 3639. https://doi.org/10.3390/ijms24043639
O’Morain VL, Chen J, Plummer SF, Michael DR, Ramji DP. Anti-Atherogenic Actions of the Lab4b Consortium of Probiotics In Vitro. International Journal of Molecular Sciences. 2023; 24(4):3639. https://doi.org/10.3390/ijms24043639
Chicago/Turabian StyleO’Morain, Victoria L., Jing Chen, Sue F. Plummer, Daryn R. Michael, and Dipak P. Ramji. 2023. "Anti-Atherogenic Actions of the Lab4b Consortium of Probiotics In Vitro" International Journal of Molecular Sciences 24, no. 4: 3639. https://doi.org/10.3390/ijms24043639