
Decreased Vitamin D Levels and Altered Placental Vitamin D Gene Expression at High Altitude: Role of Genetic Ancestry
 and
and
Abstract
:1. Introduction
2. Results
2.1. Demographic and Clinical Characteristics
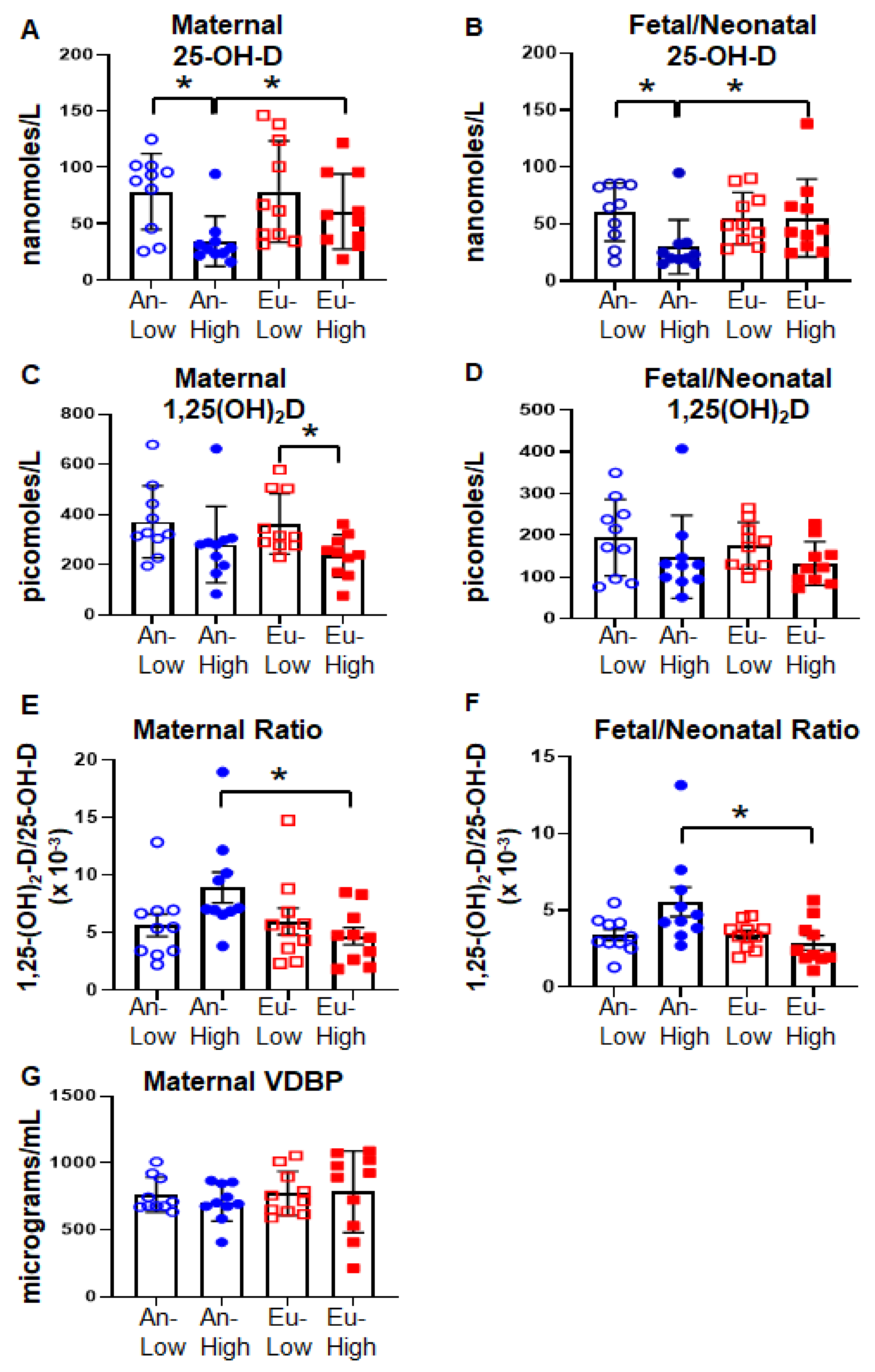
2.2. Vitamin D Status in Bolivian Pregnancies
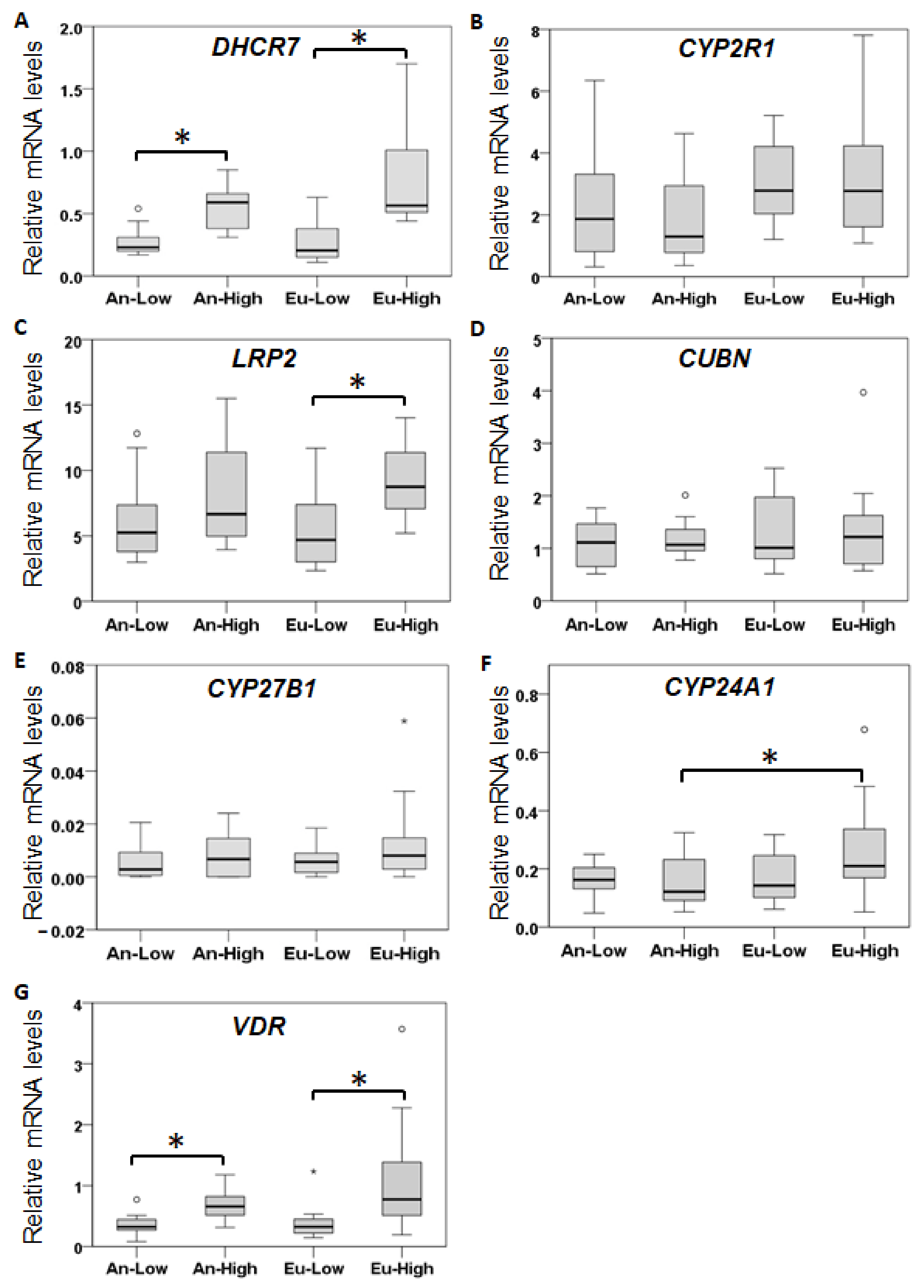
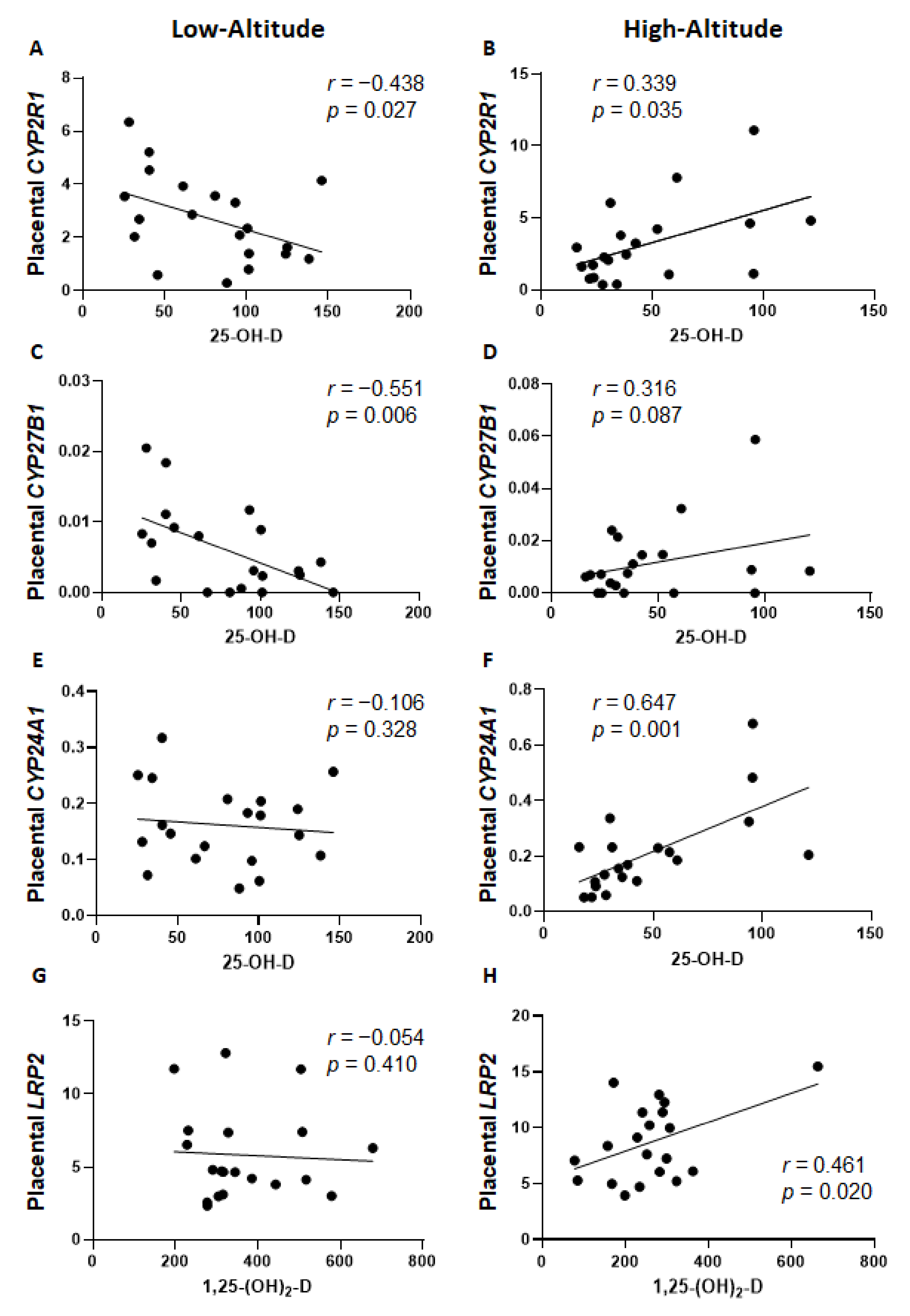
2.3. Placental Expression of Vitamin D-Related Genes
3. Discussion
4. Materials and Methods
4.1. Study Design
4.2. Vitamin D Status and Vitamin D Binding Protein ELISA Assays
4.3. Placental Vitamin D-Related Gene Expression Analysis
4.4. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Herrera, E.A.; Krause, B.; Ebensperger, G.; Reyes, R.V.; Casanello, P.; Parra-Cordero, M.; Llanos, A.J. The Placental Pursuit for an Adequate Oxidant Balance between the Mother and the Fetus. Front. Pharmacol. 2014, 5, 149. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez-Candia, A.; Herrera, E.A. High Altitude Pregnancies and Vascular Dysfunction: Observations from Latin American Studies. Front. Physiol. 2021, 12, 786038. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Helbich-Poschacher, V.; Cao, C.; Klebermass-Schrehof, K.; Waldhoer, T. Maternal Altitude and Risk of Low Birthweight: A Systematic Review and Meta-Analyses. Placenta 2020, 101, 124–131. [Google Scholar] [CrossRef] [PubMed]
- Julian, C.G. High Altitude during Pregnancy. Clin. Chest Med. 2011, 32, 21–31. [Google Scholar] [CrossRef]
- van Patot, M.C.T.; Ebensperger, G.; Gassmann, M.; Llanos, A.J. The Hypoxic Placenta. High Alt. Med. Biol. 2012, 13, 176–184. [Google Scholar] [CrossRef]
- Moore, L.G.; Niermeyer, S.; Zamudio, S. Human Adaptation to High Altitude: Regional and Life-Cycle Perspectives. Am. J. Phys. Anthropol. 1998, 107, 25–64. [Google Scholar] [CrossRef]
- Moore, L.G.; Shriver, M.; Bemis, L.; Hickler, B.; Wilson, M.; Brutsaert, T.; Parra, E.; Vargas, E. Maternal Adaptation to High-Altitude Pregnancy: An Experiment of Nature—A Review. Placenta 2004, 25, S60–S71. [Google Scholar] [CrossRef]
- Postigo, L.; Heredia, G.; Illsley, N.P.; Torricos, T.; Dolan, C.; Echalar, L.; Tellez, W.; Maldonado, I.; Brimacombe, M.; Balanza, E.; et al. Where the O 2 Goes to: Preservation of Human Fetal Oxygen Delivery and Consumption at High Altitude: Hypoxia and Fetal O 2 Consumption. J. Physiol. 2009, 587, 693–708. [Google Scholar] [CrossRef]
- Storz, J.F. High-Altitude Adaptation: Mechanistic Insights from Integrated Genomics and Physiology. Mol. Biol. Evol. 2021, 38, 2677–2691. [Google Scholar] [CrossRef]
- Arshad, R.; Sameen, A.; Murtaza, M.A.; Sharif, H.R.; Iahtisham-Ul-Haq, D.S.; Ahmed, Z.; Nemat, A.; Manzoor, M.F. Impact of Vitamin D on Maternal and Fetal Health: A Review. Food Sci. Nutr. 2022, 10, 3230–3240. [Google Scholar] [CrossRef]
- Ganguly, A.; Tamblyn, J.A.; Finn-Sell, S.; Chan, S.-Y.; Westwood, M.; Gupta, J.; Kilby, M.D.; Gross, S.R.; Hewison, M. Vitamin D, the Placenta and Early Pregnancy: Effects on Trophoblast Function. J. Endocrinol. 2018, 236, R93–R103. [Google Scholar] [CrossRef]
- Wei, S.-Q.; Qi, H.-P.; Luo, Z.-C.; Fraser, W.D. Maternal Vitamin D Status and Adverse Pregnancy Outcomes: A Systematic Review and Meta-Analysis. J. Matern.-Fetal Neonatal Med. 2013, 26, 889–899. [Google Scholar] [CrossRef]
- Weinert, L.S.; Silveiro, S.P. Maternal-Fetal Impact of Vitamin D Deficiency: A Critical Review. Matern. Child Health J. 2015, 19, 94–101. [Google Scholar] [CrossRef]
- van der Pligt, P.; Willcox, J.; Szymlek-Gay, E.; Murray, E.; Worsley, A.; Daly, R. Associations of Maternal Vitamin D Deficiency with Pregnancy and Neonatal Complications in Developing Countries: A Systematic Review. Nutrients 2018, 10, 640. [Google Scholar] [CrossRef]
- Mulligan, M.L.; Felton, S.K.; Riek, A.E.; Bernal-Mizrachi, C. Implications of Vitamin D Deficiency in Pregnancy and Lactation. Am. J. Obstet. Gynecol. 2010, 202, 429.e1–429.e9. [Google Scholar] [CrossRef]
- Lucas, R.M.; Ponsonby, A.-L.; Pasco, J.A.; Morley, R. Future Health Implications of Prenatal and Early-Life Vitamin D Status. Nutr. Rev. 2008, 66, 710–720. [Google Scholar] [CrossRef]
- Erkkola, M.; Nwaru, B.I.; Viljakainen, H.T. Maternal Vitamin D during Pregnancy and Its Relation to Immune-Mediated Diseases in the Offspring. Vitam. Horm. 2011, 86, 239–260. [Google Scholar] [CrossRef]
- Weiss, S.T.; Litonjua, A.A. The In Utero Effects of Maternal Vitamin D Deficiency: How It Results in Asthma and Other Chronic Diseases. Am. J. Respir. Crit. Care Med. 2011, 183, 1286–1287. [Google Scholar] [CrossRef]
- Rosen, C.J.; Abrams, S.A.; Aloia, J.F.; Brannon, P.M.; Clinton, S.K.; Durazo-Arvizu, R.A.; Gallagher, J.C.; Gallo, R.L.; Jones, G.; Kovacs, C.S.; et al. IOM Committee Members Respond to Endocrine Society Vitamin D Guideline. J. Clin. Endocrinol. Metab. 2012, 97, 1146–1152. [Google Scholar] [CrossRef]
- Li, H.; Ma, J.; Huang, R.; Wen, Y.; Liu, G.; Xuan, M.; Yang, L.; Yang, J.; Song, L. Prevalence of Vitamin D Deficiency in the Pregnant Women: An Observational Study in Shanghai, China. Arch. Public Health 2020, 78, 31. [Google Scholar] [CrossRef]
- Vandevijvere, S.; Amsalkhir, S.; Van Oyen, H.; Moreno-Reyes, R. High Prevalence of Vitamin D Deficiency in Pregnant Women: A National Cross-Sectional Survey. PLoS ONE 2012, 7, e43868. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.J.; Zhang, F.; Richards, J.B.; Kestenbaum, B.; Van Meurs, J.B.; Berry, D.; Kiel, D.P.; Streeten, E.A.; Ohlsson, C.; Koller, D.L.; et al. Common Genetic Determinants of Vitamin D Insufficiency: A Genome-Wide Association Study. Lancet 2010, 376, 180–188. [Google Scholar] [CrossRef] [PubMed]
- Neville, J.J.; Palmieri, T.; Young, A.R. Physical Determinants of Vitamin D Photosynthesis: A Review. JBMR Plus 2021, 5, e10460. [Google Scholar] [CrossRef] [PubMed]
- Snellman, G.; Melhus, H.; Gedeborg, R.; Olofsson, S.; Wolk, A.; Pedersen, N.L.; Michaëlsson, K. Seasonal Genetic Influence on Serum 25-Hydroxyvitamin D Levels: A Twin Study. PLoS ONE 2009, 4, e7747. [Google Scholar] [CrossRef]
- Alp, H.; Tekgündüz, K.Ş.; Akkar, M.K. Maternal and Cord Blood Vitamin D Status in High-Altitude Pregnancy. J. Matern.-Fetal Neonatal Med. 2016, 29, 571–575. [Google Scholar] [CrossRef]
- Gelsor, N.; Ma, L.; Duan, J.; Wangmu, T.; Gelsor, N. Human Vitamin D Deficiency in Tibet. Food Nutr. Sci. 2017, 08, 1127–1136. [Google Scholar] [CrossRef]
- Norsang, G.; Ma, L.; Dahlback, A.; Zhuoma, C.; Tsoja, W.; Porojnicu, A.; Lagunova, Z.; Moan, J. The Vitamin D Status among Tibetans. Photochem. Photobiol. 2009, 85, 1028–1031. [Google Scholar] [CrossRef]
- Orces, C.H. Vitamin D Status among Older Adults Residing in the Littoral and Andes Mountains in Ecuador. Sci. World J. 2015, 2015, 545297. [Google Scholar] [CrossRef]
- Hirschler, V.; Molinari, C.; Maccallini, G.; Intersimone, P.; Gonzalez, C.D. Vitamin D Levels and Cardiometabolic Markers in Indigenous Argentinean Children Living at Different Altitudes. Glob. Pediatr. Health 2019, 6, 1882194. [Google Scholar] [CrossRef]
- Rottenberg, M.E.; Nylén, S.; Rodriguez, C.; Terán, G.; Cuna, W.; Brañez, F.; Persson, K.E.M. Differences in Nutritional and Health Status in School Children from the Highlands and Lowlands of Bolivia. Am. J. Trop. Med. Hyg. 2018, 98, 326–333. [Google Scholar] [CrossRef] [Green Version]
- Gray, T.K.; Lowe, W.; Lester, G.E. Vitamin D and Pregnancy: The Maternal-Fetal Metabolism of Vitamin D. Endocr. Rev. 1981, 2, 264–274. [Google Scholar] [CrossRef]
- Paulson, S.K.; Deluca, H.F. Review Article: Vitamin D Metabolism during Pregnancy. Bone 1986, 7, 331–336. [Google Scholar] [CrossRef]
- Karras, S.N.; Wagner, C.L.; Castracane, V.D. Understanding Vitamin D Metabolism in Pregnancy: From Physiology to Pathophysiology and Clinical Outcomes. Metabolism 2018, 86, 112–123. [Google Scholar] [CrossRef]
- Dusso, A.S.; Brown, A.J.; Slatopolsky, E. Vitamin D. Am. J. Physiol. Renal Physiol. 2005, 289, F8–F28. [Google Scholar] [CrossRef]
- Norman, A.W. From Vitamin D to Hormone D: Fundamentals of the Vitamin D Endocrine System Essential for Good Health. Am. J. Clin. Nutr. 2008, 88, 491S–499S. [Google Scholar] [CrossRef]
- Haussler, M.R.; Haussler, C.A.; Bartik, L.; Whitfield, G.K.; Hsieh, J.-C.; Slater, S.; Jurutka, P.W. Vitamin D Receptor: Molecular Signaling and Actions of Nutritional Ligands in Disease Prevention. Nutr. Rev. 2008, 66, S98–S112. [Google Scholar] [CrossRef]
- Kasprzak, Z.; Śliwicka, E.; Hennig, K.; Pilaczyńska-Szcześniak, Ł.; Huta-Osiecka, A.; Nowak, A. Vitamin D, Iron Metabolism, and Diet in Alpinists During a 2-Week High-Altitude Climb. High Alt. Med. Biol. 2015, 16, 230–235. [Google Scholar] [CrossRef]
- Ashley, B.; Simner, C.; Manousopoulou, A.; Jenkinson, C.; Hey, F.; Frost, J.M.; Rezwan, F.I.; White, C.H.; Lofthouse, E.M.; Hyde, E.; et al. Placental Uptake and Metabolism of 25(OH)Vitamin D Determine Its Activity within the Fetoplacental Unit. eLife 2022, 11, e71094. [Google Scholar] [CrossRef]
- Zamudio, S.; Postigo, L.; Illsley, N.P.; Rodriguez, C.; Heredia, G.; Brimacombe, M.; Echalar, L.; Torricos, T.; Tellez, W.; Maldonado, I.; et al. Maternal Oxygen Delivery Is Not Related to Altitude- and Ancestry-Associated Differences in Human Fetal Growth: Maternal O 2 Transport and Fetal Growth. J. Physiol. 2007, 582, 883–895. [Google Scholar] [CrossRef]
- Park, H.; Wood, M.R.; Malysheva, O.V.; Jones, S.; Mehta, S.; Brannon, P.M.; Caudill, M.A. Placental Vitamin D Metabolism and Its Associations with Circulating Vitamin D Metabolites in Pregnant Women. Am. J. Clin. Nutr. 2017, 106, 1439–1448. [Google Scholar] [CrossRef] [Green Version]
- Martin, C.B.; Oshiro, B.T.; Sands, L.D.; Kabir, S.; Thorpe, D.; Clark, T.C.; Yao, R.; Mata-Greenwood, E. Vitamin-D Dysregulation in Early- and Late-Onset Preeclampsia: A Gestational-Age Matched Study. J. Steroid Biochem. Mol. Biol. 2020, 203, 105729. [Google Scholar] [CrossRef] [PubMed]
- Temte, J.L. Elevation of Serum Cholesterol at High Altitude and Its Relationship to Hematocrit. Wilderness Environ. Med. 1996, 7, 216–224. [Google Scholar] [CrossRef] [PubMed]
- Gonzales, G.F.; Tapia, V. Association of High Altitude-Induced Hypoxemia to Lipid Profile and Glycemia in Men and Women Living at 4100m in the Peruvian Central Andes. Endocrinol. Nutr. 2013, 60, 79–86. [Google Scholar] [CrossRef] [PubMed]
- Baardman, M.E.; Kerstjens-Frederikse, W.S.; Berger, R.M.F.; Bakker, M.K.; Hofstra, R.M.W.; Plösch, T. The Role of Maternal-Fetal Cholesterol Transport in Early Fetal Life: Current Insights1. Biol. Reprod. 2013, 88, 24. [Google Scholar] [CrossRef]
- Storm, T.; Christensen, E.I.; Christensen, J.N.; Kjaergaard, T.; Uldbjerg, N.; Larsen, A.; Honoré, B.; Madsen, M. Megalin Is Predominantly Observed in Vesicular Structures in First and Third Trimester Cytotrophoblasts of the Human Placenta. J. Histochem. Cytochem. 2016, 64, 769–784. [Google Scholar] [CrossRef]
- Zamudio, S.; Wu, Y.; Ietta, F.; Rolfo, A.; Cross, A.; Wheeler, T.; Post, M.; Illsley, N.P.; Caniggia, I. Human Placental Hypoxia-Inducible Factor-1α Expression Correlates with Clinical Outcomes in Chronic Hypoxia in Vivo. Am. J. Pathol. 2007, 170, 2171–2179. [Google Scholar] [CrossRef]
- Knabl, J.; Vattai, A.; Ye, Y.; Jueckstock, J.; Hutter, S.; Kainer, F.; Mahner, S.; Jeschke, U. Role of Placental VDR Expression and Function in Common Late Pregnancy Disorders. Int. J. Mol. Sci. 2017, 18, 2340. [Google Scholar] [CrossRef]
- Li, L.; Li, K.; Li, J.; Luo, Y.; Cheng, Y.; Jian, M.; Xie, C.; Ji, C.; Chuan, L.; Wang, Z.; et al. Ethnic, Geographic, and Seasonal Differences of Vitamin D Status among Adults in South-west China. J. Clin. Lab. Anal. 2020, 34, e23532. [Google Scholar] [CrossRef]
- Ahmed, L.H.M.; Butler, A.E.; Dargham, S.R.; Latif, A.; Chidiac, O.M.; Atkin, S.L.; Abi Khalil, C. Vitamin D3 Metabolite Ratio as an Indicator of Vitamin D Status and Its Association with Diabetes Complications. BMC Endocr. Disord. 2020, 20, 161. [Google Scholar] [CrossRef]
- Castillo-Peinado, L.d.L.S.; Calderón-Santiago, M.; Herrera-Martínez, A.D.; León-Idougourram, S.; Gálvez-Moreno, M.Á.; Sánchez-Cano, R.L.; Bouillon, R.; Quesada-Gómez, J.M.; Priego-Capote, F. Measuring Vitamin D3 Metabolic Status, Comparison between Vitamin D Deficient and Sufficient Individuals. Separations 2022, 9, 141. [Google Scholar] [CrossRef]
- Albahlol, I.A.; Almaeen, A.H.; Alduraywish, A.A.; Dar, U.F.; El-Metwally, T.H. Vitamin D Status and Pregnancy Complications: Serum 1,25-Di-Hydroxyl-Vitamin D and Its Ratio to 25-Hydroxy-Vitamin D Are Superior Biomarkers than 25-Hydroxy-Vitamin D. Int. J. Med. Sci. 2020, 17, 3039–3048. [Google Scholar] [CrossRef]
- Ma, R.; Gu, Y.; Zhao, S.; Sun, J.; Groome, L.J.; Wang, Y. Expressions of Vitamin D Metabolic Components VDBP, CYP2R1, CYP27B1, CYP24A1, and VDR in Placentas from Normal and Preeclamptic Pregnancies. Am. J. Physiol.-Endocrinol. Metab. 2012, 303, E928–E935. [Google Scholar] [CrossRef]
- Wang, Y.; Wang, T.; Huo, Y.; Liu, L.; Liu, S.; Yin, X.; Wang, R.; Gao, X. Placenta Expression of Vitamin D and Related Genes in Pregnant Women with Gestational Diabetes Mellitus. J. Steroid Biochem. Mol. Biol. 2020, 204, 105754. [Google Scholar] [CrossRef]
- O’Brien, K.O.; Li, S.; Cao, C.; Kent, T.; Young, B.V.; Queenan, R.A.; Pressman, E.K.; Cooper, E.M. Placental CYP27B1 and CYP24A1 Expression in Human Placental Tissue and Their Association with Maternal and Neonatal Calcitropic Hormones. J. Clin. Endocrinol. Metab. 2014, 99, 1348–1356. [Google Scholar] [CrossRef]
- Goyal, R.; Zhang, L.; Blood, A.B.; Baylink, D.J.; Longo, L.D.; Oshiro, B.; Mata-Greenwood, E. Characterization of an Animal Model of Pregnancy-Induced Vitamin D Deficiency Due to Metabolic Gene Dysregulation. Am. J. Physiol. Endocrinol. Metab. 2014, 306, E256–E266. [Google Scholar] [CrossRef]
- Goyal, R.; Billings, T.L.; Mansour, T.; Martin, C.; Baylink, D.J.; Longo, L.D.; Pearce, W.J.; Mata-Greenwood, E. Vitamin D Status and Metabolism in an Ovine Pregnancy Model: Effect of Long-Term, High-Altitude Hypoxia. Am. J. Physiol.-Endocrinol. Metab. 2016, 310, E1062–E1071. [Google Scholar] [CrossRef]
- Mata-Greenwood, E.; Huber, H.F.; Li, C.; Nathanielsz, P.W. Role of Pregnancy and Obesity on Vitamin D Status, Transport, and Metabolism in Baboons. Am. J. Physiol.-Endocrinol. Metab. 2019, 316, E63–E72. [Google Scholar] [CrossRef]
- Charles, S.M.; Julian, C.G.; Vargas, E.; Moore, L.G. Higher Estrogen Levels During Pregnancy in Andean Than European Residents of High Altitude Suggest Differences in Aromatase Activity. J. Clin. Endocrinol. Metab. 2014, 99, 2908–2916. [Google Scholar] [CrossRef]
- Harmon, Q.E.; Umbach, D.M.; Baird, D.D. Use of Estrogen-Containing Contraception Is Associated with Increased Concentrations of 25-Hydroxy Vitamin D. J. Clin. Endocrinol. Metab. 2016, 101, 3370–3377. [Google Scholar] [CrossRef]
- Zhao, D.; Ouyang, P.; de Boer, I.H.; Lutsey, P.L.; Farag, Y.M.K.; Guallar, E.; Siscovick, D.S.; Post, W.S.; Kalyani, R.R.; Billups, K.L.; et al. Serum Vitamin D and Sex Hormones Levels in Men and Women: The Multi-Ethnic Study of Atherosclerosis (MESA). Maturitas 2017, 96, 95–102. [Google Scholar] [CrossRef] [Green Version]
- Nashold, F.E.; Spach, K.M.; Spanier, J.A.; Hayes, C.E. Estrogen Controls Vitamin D3-Mediated Resistance to Experimental Autoimmune Encephalomyelitis by Controlling Vitamin D3 Metabolism and Receptor Expression. J. Immunol. 2009, 183, 3672–3681. [Google Scholar] [CrossRef] [PubMed]
- Illsley, N.P.; Caniggia, I.; Zamudio, S. Placental Metabolic Reprogramming: Do Changes in the Mix of Energy-Generating Substrates Modulate Fetal Growth? Int. J. Dev. Biol. 2010, 54, 409–419. [Google Scholar] [CrossRef] [PubMed]
- Krasniqi, E.; Boshnjaku, A.; Wagner, K.-H.; Wessner, B. Association between Polymorphisms in Vitamin D Pathway-Related Genes, Vitamin D Status, Muscle Mass and Function: A Systematic Review. Nutrients 2021, 13, 3109. [Google Scholar] [CrossRef] [PubMed]
- Ruiz-Ballesteros, A.I.; Meza-Meza, M.R.; Vizmanos-Lamotte, B.; Parra-Rojas, I.; de la Cruz-Mosso, U. Association of Vitamin D Metabolism Gene Polymorphisms with Autoimmunity: Evidence in Population Genetic Studies. Int. J. Mol. Sci. 2020, 21, 9626. [Google Scholar] [CrossRef]
- Galvão, A.A.; de Araújo Sena, F.; de Andrade Belitardo, E.M.M.; de Santana, M.B.R.; de Oliveira Costa, G.N.; Cruz, Á.A.; Barreto, M.L.; dos Santos Costa, R.; Alcantara-Neves, N.M.; Figueiredo, C.A. Genetic Polymorphisms in Vitamin D Pathway Influence 25(OH)D Levels and Are Associated with Atopy and Asthma. Allergy Asthma Clin. Immunol. 2020, 16, 62. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Characteristics a | Andean Ancestry at 400 m Altitude (n = 10) | European Ancestry at 400 m Altitude (n = 10) | Andean Ancestry at 3600 m Altitude (n = 10) | European Ancestry at 3600 m Altitude (n = 10) | p-Value |
|---|---|---|---|---|---|
| Maternal | |||||
| Age (yr) | 29.1 ± 1.7 | 25.6 ± 1.7 | 32.4 ± 1.6 | 30.0 ± 1.7 | 0.008 * |
| Pre-pregnancy BMI (kg/m2) | 23.1 ± 0.8 | 23.3 ± 1.0 | 26.4 ± 2.1 | 23.4 ± 0.9 | 0.277 * |
| Fetal/Neonatal | |||||
| Sex (% Male) | 60 | 70 | 50 | 50 | 0.771 † |
| Gestational age (wk) | 39.5 ± 0.4 | 38.4 ± 0.3 | 39.1 ± 0.4 | 37.9± 0.3 | 0.016 * |
| Birthweight-BW (g) | 3491 ± 106 | 3453 ± 94 | 3446 ± 119 | 2939 ± 95 | 0.001 * |
| BW Percentile (%) | 52.1 ± 7.0 | 63.6 ± 8.6 | 51.4 ± 7.6 | 31.0 ± 6.2 | 0.030 * |
| BW:PW ratio | 7.54 ± 0.41 | 7.76 ± 0.28 | 6.42 ± 0.38 | 6.23 ± 0.46 | 0.016 * |
| Ponderal Index | 2.65 ± 0.09 | 2.73 ± 0.05 | 2.84 ± 0.10 | 2.48 ± 0.06 | 0.022 * |
| Head Circumference | 35.1 ± 0.5 | 34.7 ± 0.3 | 34.6 ± 0.3 | 34.0 ± 0.5 | 0.387 * |
| Ab. Circumference | 34.7 ± 0.5 | 33.9 ± 0.3 | 34.9 ± 0.2 | 33.1 ± 0.4 | 0.005 * |
| Dependent Variable (n = 40) | Model * Adjusted R2 | Beta Coefficient | Confidence Interval 95% | p |
|---|---|---|---|---|
| Maternal 25-OH-D | 0.149 | 0.008 | ||
| 1. Altitude | 30.9 | 8.6–53.3 | 0.008 | |
| Fetal/neonatal 25-OH-D | 0.156 | 0.016 | ||
| 1. Altitude | 26.4 | 7.1–45.7 | 0.009 | |
| 2. BW: PW | −8.5 | −15.6–(−1.4) | 0.020 | |
| Maternal 1α,25-(OH)2-D | 0.282 | <0.001 | ||
| 1. Altitude | 168 | 83.8–252.5 | <0.001 | |
| 2. BW: PW | −44.0 | −75.1–(−12.8) | 0.007 | |
| Fetal/neonatal 1α,25-(OH)2-D | 0.256 | 0.002 | ||
| 1. Altitude | 84.9 | 35–134.7 | 0.001 | |
| 2. BW: PW | −30.3 | −48.7–(−11.9) | 0.002 |
| Model * Adjusted R2 | Beta Coefficient | Confidence Interval 95% | p | |
|---|---|---|---|---|
| Low Altitude Subjects (n = 20) | ||||
| Maternal 25-OH-D | 0.265 | 0.012 | ||
| 1. CYP27B1 | −3527 | −6173–(−881) | 0.012 | |
| Fetal/neonatal 25-OH-D | 0.221 | 0.021 | ||
| 1. CYP2R1 | −7.52 | −13.79–(−1.26) | 0.021 | |
| High-Altitude Subjects (n = 20) | ||||
| Maternal 25-OH-D | 0.505 | 0.001 | ||
| 1. CYP24A1 | 118.7 | 50.0–187.4 | 0.002 | |
| 2. CYP2R1 | 6.18 | 0.54–11.8 | 0.034 | |
| Fetal/Neonatal 25-OH-D | 0.351 | 0.010 | ||
| 1. CYP2R1 | 7.75 | 1.14–14.4 | 0.024 | |
| 2. CYP24A1 | 80.7 | 0.1–161.3 | 0.050 | |
| Maternal 1α,25-(OH)2-D | 0.297 | 0.019 | ||
| 1. LRP2 | 15.98 | 1.49–30.47 | 0.033 | |
| 2. CYP24A1 | 314.6 | −6.1–635.2 | 0.054 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mata-Greenwood, E.; Westenburg, H.C.A.; Zamudio, S.; Illsley, N.P.; Zhang, L. Decreased Vitamin D Levels and Altered Placental Vitamin D Gene Expression at High Altitude: Role of Genetic Ancestry. Int. J. Mol. Sci. 2023, 24, 3389. https://doi.org/10.3390/ijms24043389
Mata-Greenwood E, Westenburg HCA, Zamudio S, Illsley NP, Zhang L. Decreased Vitamin D Levels and Altered Placental Vitamin D Gene Expression at High Altitude: Role of Genetic Ancestry. International Journal of Molecular Sciences. 2023; 24(4):3389. https://doi.org/10.3390/ijms24043389
Chicago/Turabian StyleMata-Greenwood, Eugenia, Hans C. A. Westenburg, Stacy Zamudio, Nicholas P. Illsley, and Lubo Zhang. 2023. "Decreased Vitamin D Levels and Altered Placental Vitamin D Gene Expression at High Altitude: Role of Genetic Ancestry" International Journal of Molecular Sciences 24, no. 4: 3389. https://doi.org/10.3390/ijms24043389