
Integrative Proteomics and Phosphoproteomics Analysis of the Rat Adenohypophysis after GnRH Treatment
 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
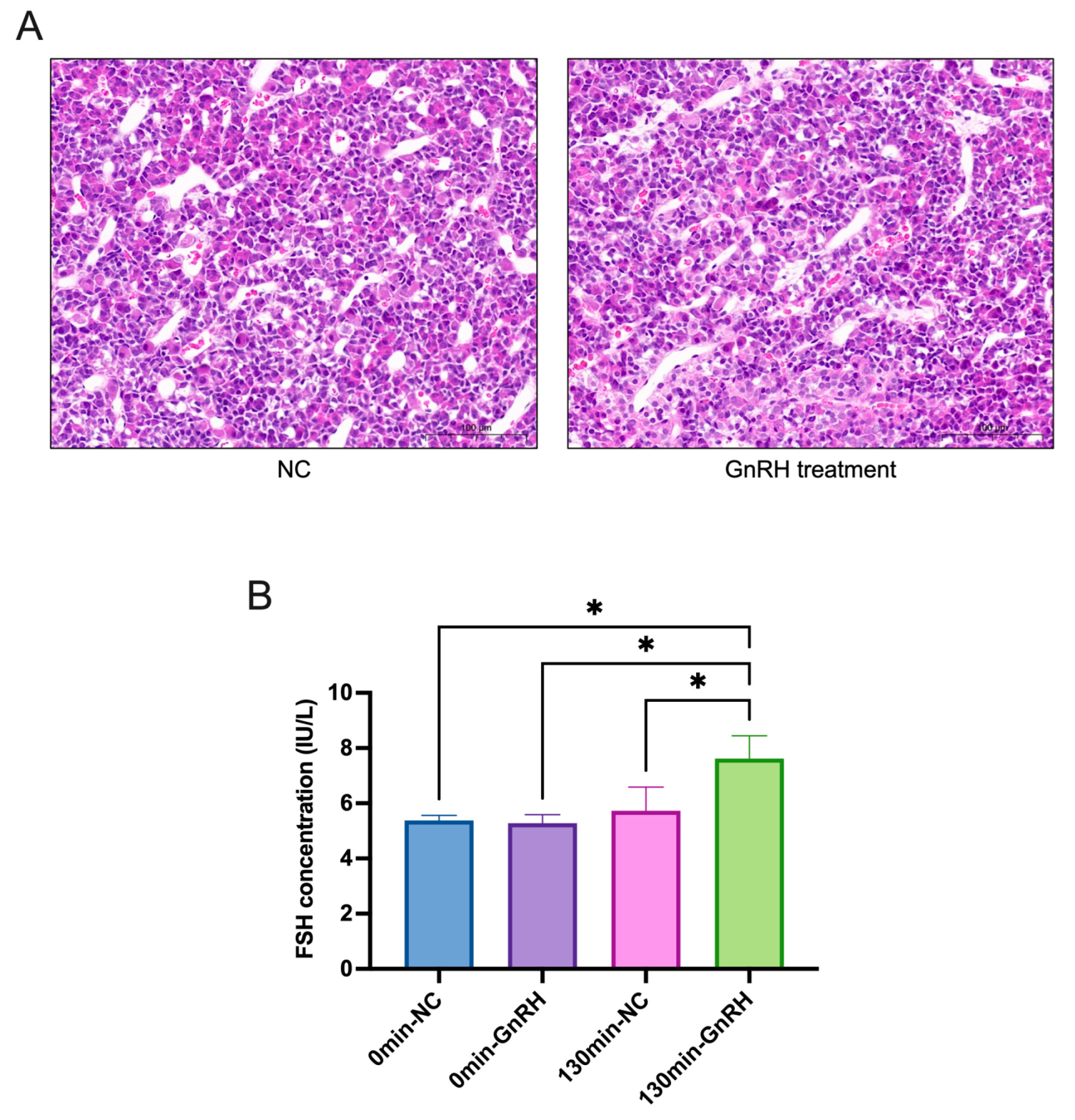
2.1. Changes in FSH after GnRH Treatment
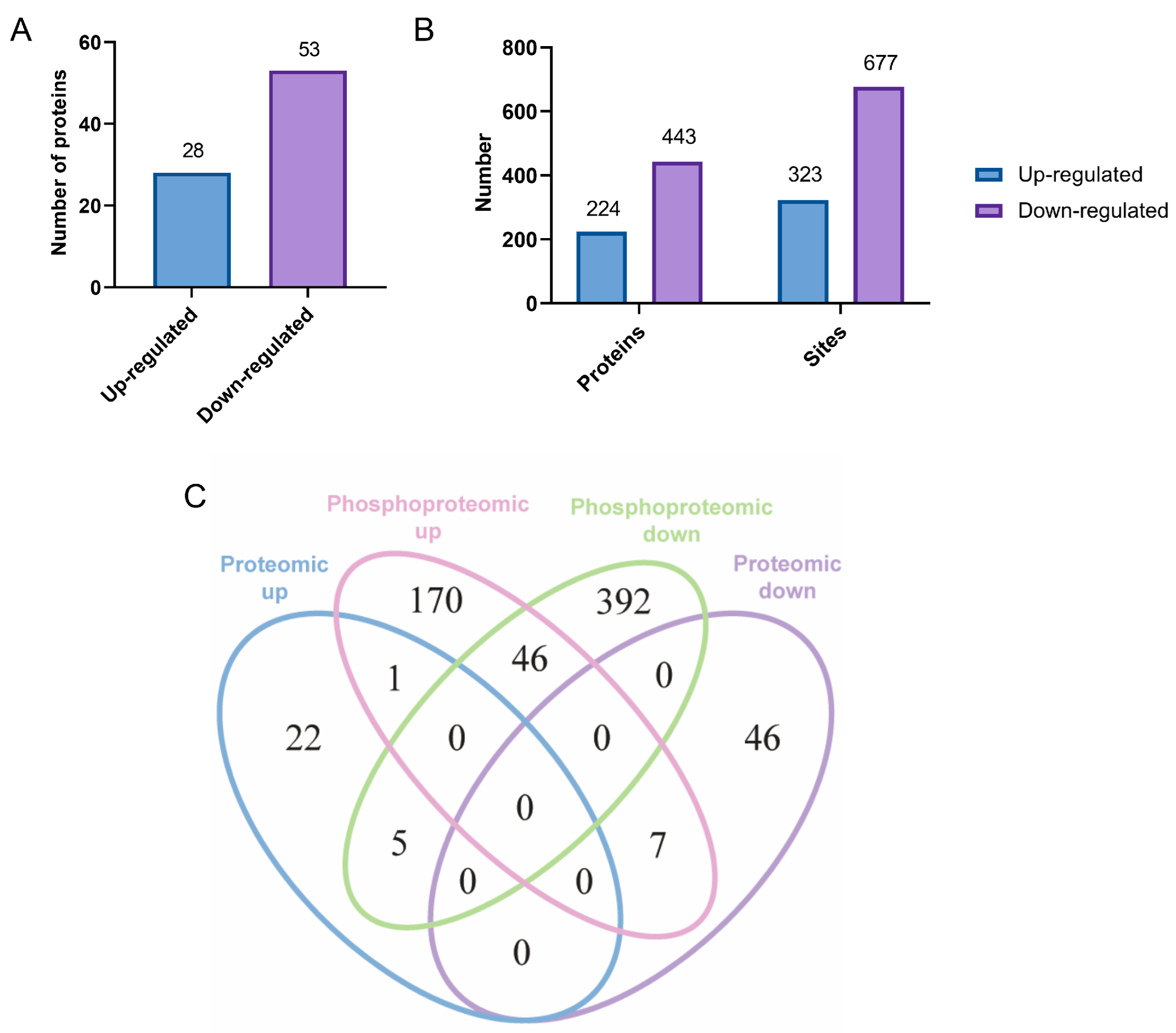
2.2. Identification and Analysis of Differential Proteins/Phosphoproteins and Phosphorylation Modification Sites
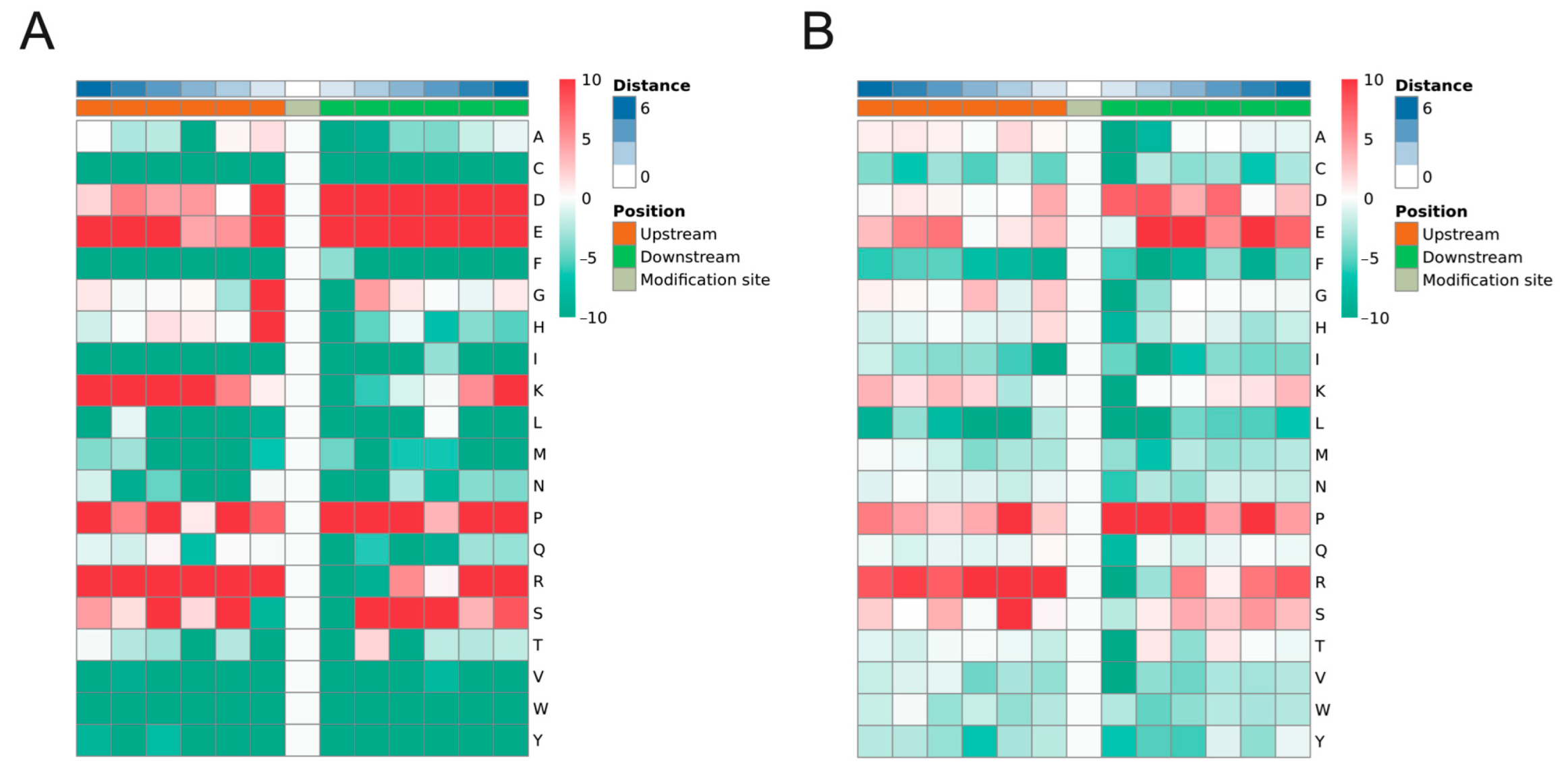
2.3. Motif Analysis of Phosphorylated Peptides
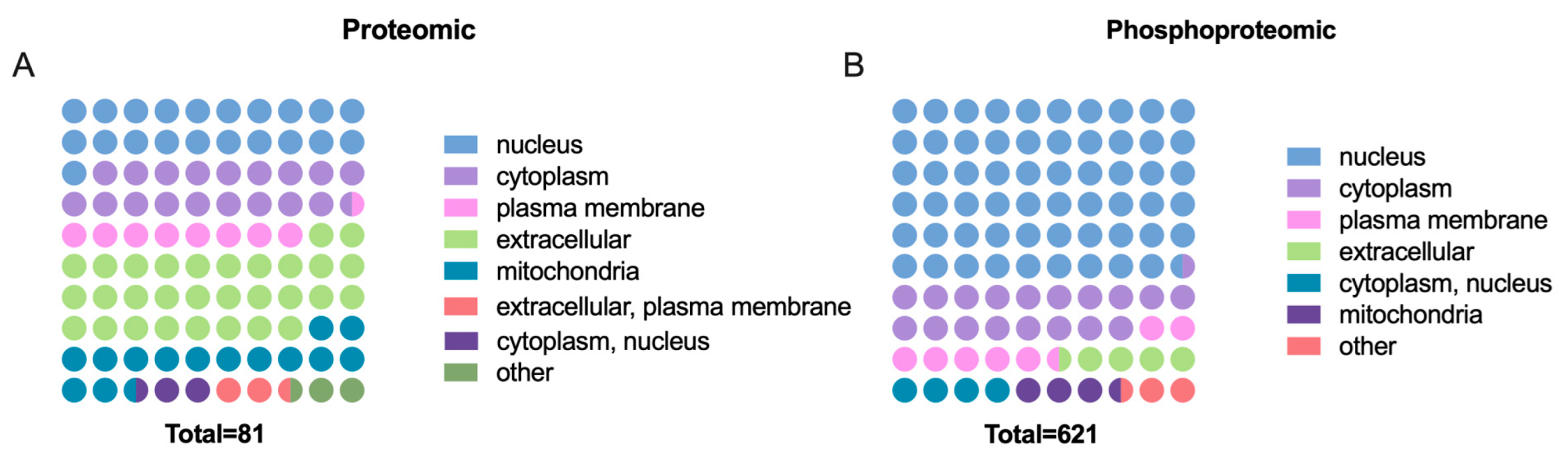
2.4. Characteristics of Cellular Localization
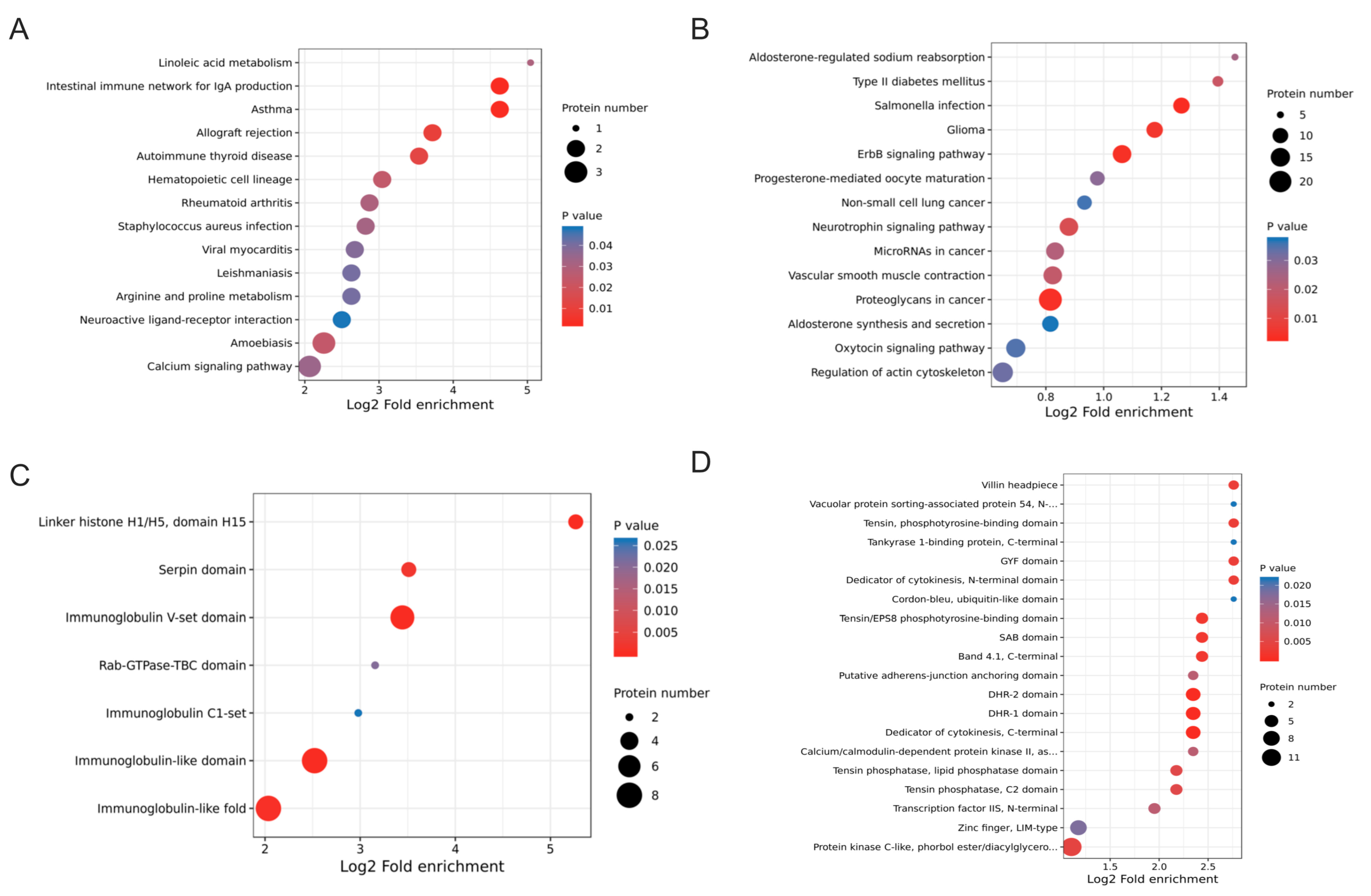
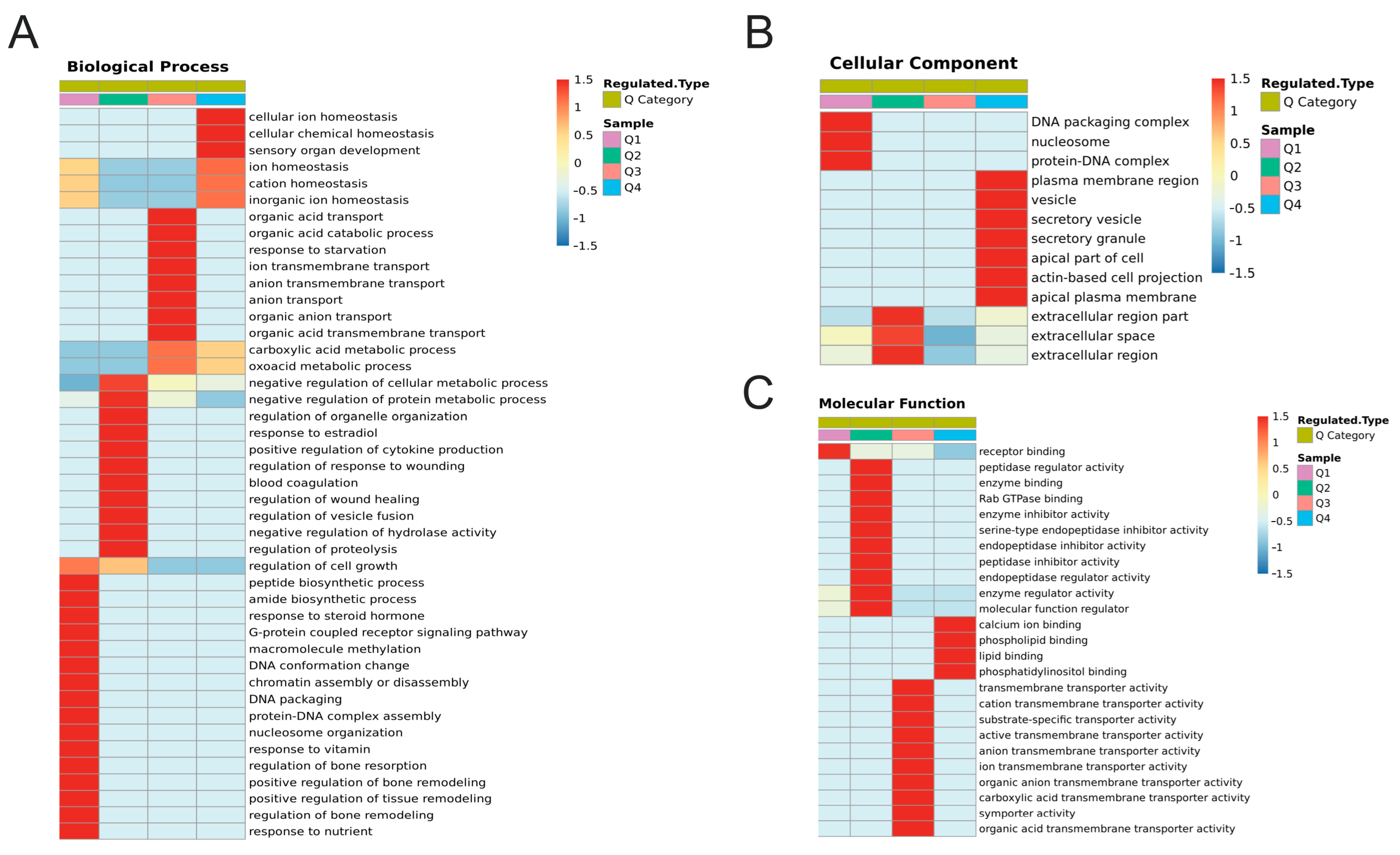
2.5. Functional Enrichment Analysis of DEPs and DEPPs
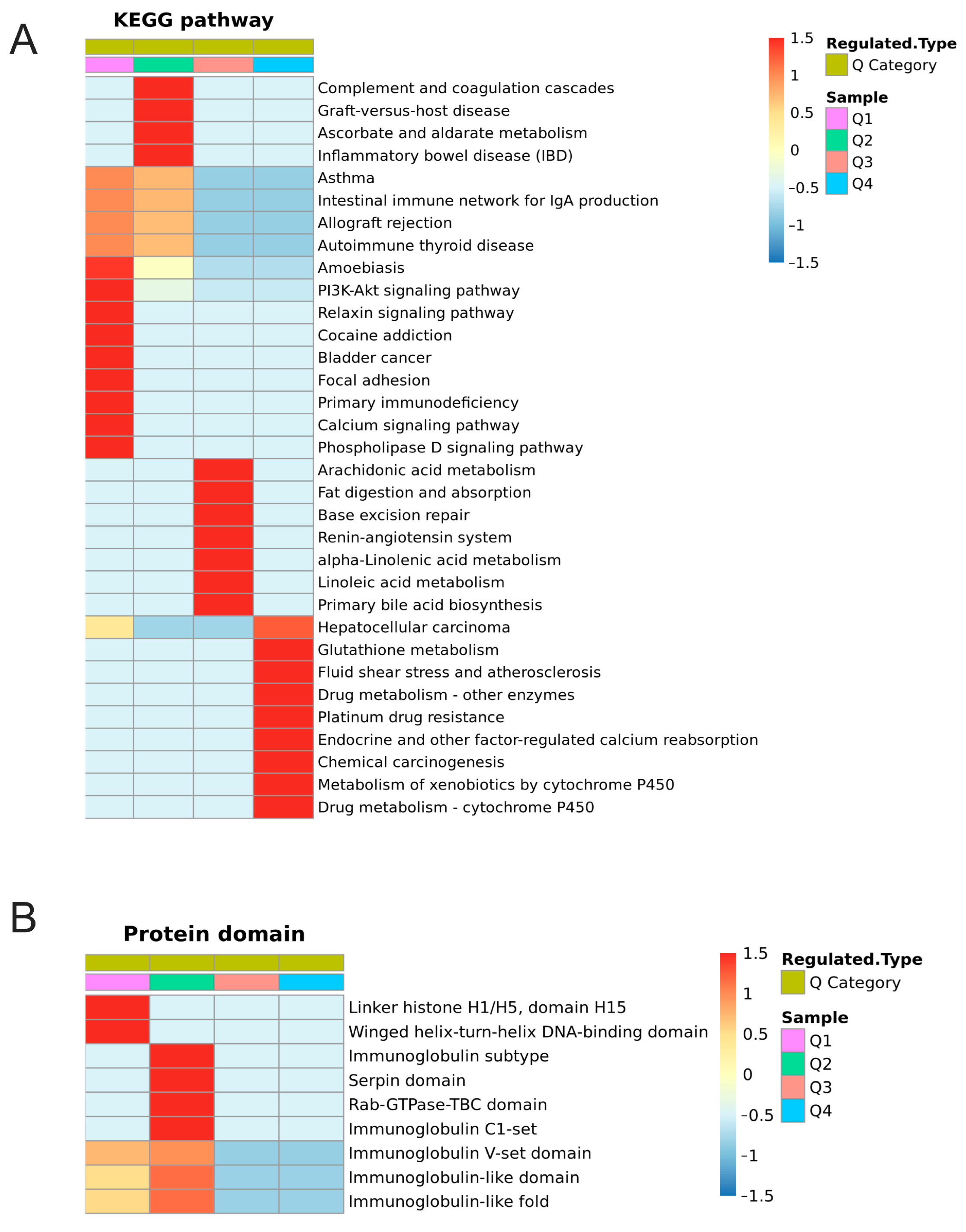
2.6. Cluster Analysis of DEPs and DEPPs
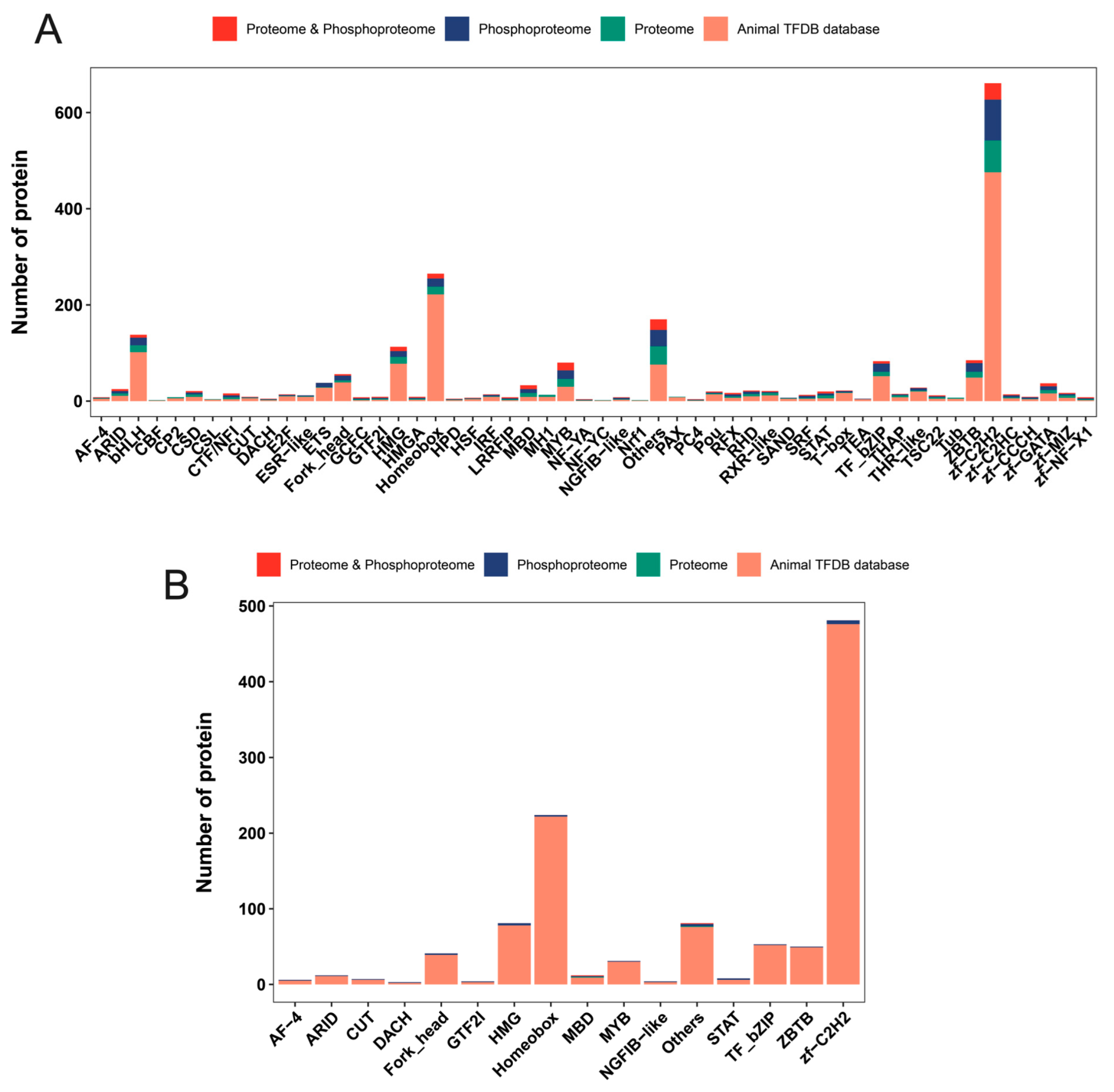
2.7. Mining for Differential Transcription Factors
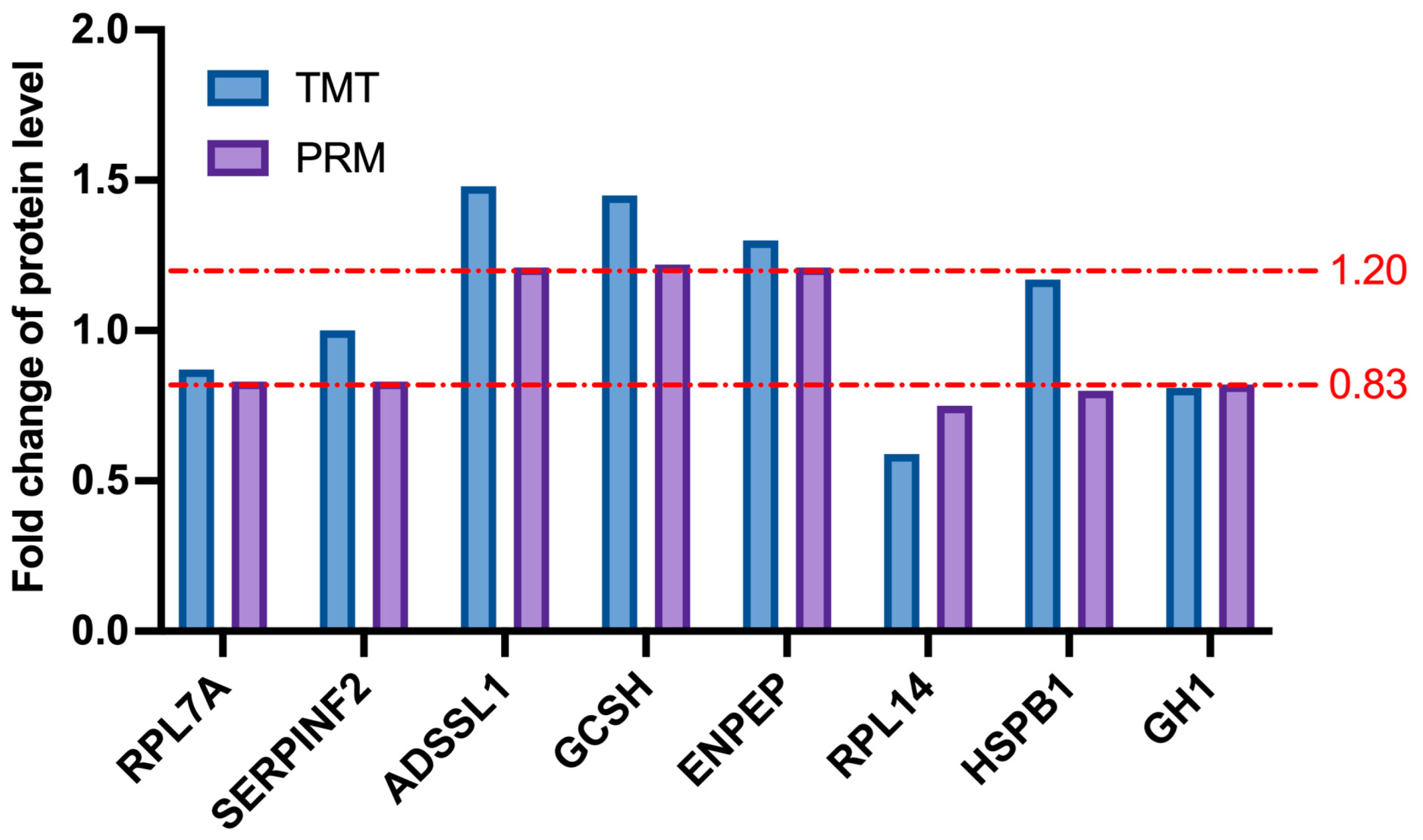
2.8. Confirmation of the Targets of Selected Differentially Abundant Proteins by PRM
3. Discussion
4. Materials and Methods
4.1. Ethics Statement
4.2. Animals & Tissue Collection
4.3. Protein Extraction and Trypsin Digestion
4.4. TMT Labeling
4.5. HPLC Fractionation and Enrichment with Phosphorylation Modifications
4.6. LC-MS/MS Analysis
4.7. Protein Annotation and Functional Enrichment Analysis
4.8. Phosphorylation Modification Site Motif Analysis
4.9. Protein Clustering Analysis
4.10. ELISA
4.11. H&E Staining
4.12. Parallel Reaction Monitoring (PRM)
4.13. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| ADSSL1 | adenylosuccinate synthetase like 1 |
| Akt | protein kinase B |
| ARID | AT-rich interaction domain |
| BCA | bicinchoninic Acid Assay |
| bHLH | basic/Helix-Loop-Helix |
| cAMP | cathelicidin antimicrobial peptide |
| DACH | dachshund family transcription factor |
| DEP | differentially expressed proteins |
| DEPP | differentially expressed phosphoproteins |
| ENPEP | glutamyl aminopeptidase |
| FHA domain | forkhead-associated domain |
| FSH | follicle-stimulating hormone |
| FSHR | follicle-stimulating hormone receptor |
| GCSH | glycine cleavage system protein H |
| GDP | guanosine diphosphate |
| GH1 | growth hormone 1 |
| GNA11 | G protein subunit alpha 11 |
| GNA15 | G protein subunit alpha 15 |
| GnRH | gonadotropin-releasing hormone |
| GnRHR | gonadotropin-releasing hormone receptor |
| GO | Gene Ontology |
| GTF2I | general transcription factor II I |
| GTP | guanosine triphosphate |
| HMG | high mobility group |
| HPG | hypothalamic-pituitary-gonadal |
| HPLC | high-performance liquid chromatography |
| HSPB1 | heat shock protein family B (small) member 1 |
| KEGG | Kyoto Encyclopedia of Genes and Genomes |
| LC-MS | liquid chromatography-tandem mass spectrometry |
| LH | luteinizing hormone |
| MAPK | mitogen-activated protein kinase |
| MBD | methyl-CpG-binding domain |
| MYB | v-myb avian myeloblastosis viral oncogene homolog |
| NGFIB | nerve growth factor IB |
| PI3K | phosphatidylinositol 3 kinase |
| PRKCA | protein kinase C, alpha |
| PRM | parallel reaction monitoring |
| PTB | phosphotyrosine binding |
| PTM | post-translational modification |
| RPL7A | ribosomal protein L7A |
| RPL10 | ribosomal protein L10 |
| RPL14 | ribosomal protein L14 |
| RPL27A | ribosomal protein L27A |
| RPL39L | ribosomal protein L39-like |
| SERPINF2 | serpin family F member 2 |
| SMAD | drosophila mothers against decapentaplegic protein |
| STAT | signal transducer and activator of transcription |
| TMT | tandem mass tags |
| rFSH | recombinant follicle stimulating hormone |
| uFSH | urinary follicle stimulating hormone |
| UPLC | ultra-performance liquid chromatography |
| ZBTB | zine finger and BTB domain protein |
References
- Wang, H.-Q.; Zhang, W.-D.; Yuan, B.; Zhang, J.-B. Advances in the Regulation of Mammalian Follicle-Stimulating Hormone Secretion. Animals 2021, 11, 1134. [Google Scholar] [CrossRef]
- Ulloa-Aguirre, A.; Zariñán, T. The Follitropin Receptor: Matching Structure and Function. Mol. Pharmacol. 2016, 90, 596–608. [Google Scholar] [CrossRef]
- Bhartiya, D.; Patel, H. An overview of FSH-FSHR biology and explaining the existing conundrums. J. Ovarian Res. 2021, 14, 144. [Google Scholar] [CrossRef]
- Zhu, L.-L.; Blair, H.; Cao, J.; Yuen, T.; Latif, R.; Guo, L.; Tourkova, I.L.; Li, J.; Davies, T.F.; Sun, L.; et al. Blocking antibody to the β-subunit of FSH prevents bone loss by inhibiting bone resorption and stimulating bone synthesis. Proc. Natl. Acad. Sci. USA 2012, 109, 14574–14579. [Google Scholar] [CrossRef]
- Wang, Y.; Zhang, M.; Huan, Z.; Shao, S.; Zhang, X.; Kong, D.; Xu, J. FSH directly regulates chondrocyte dedifferentiation and cartilage development. J. Endocrinol. 2021, 248, 193–206. [Google Scholar] [CrossRef]
- Liu, Z.; Sun, Y.; Jiang, Y.; Qian, Y.; Chen, S.; Gao, S.; Chen, L.; Li, C.; Zhou, X. Follicle-stimulating hormone (FSH) promotes retinol uptake and metabolism in the mouse ovary. Reprod. Biol. Endocrinol. 2018, 16, 52. [Google Scholar] [CrossRef]
- Liu, P.; Ji, Y.; Yuen, T.; Rendina-Ruedy, E.; DeMambro, V.E.; Dhawan, S.; Abu-Amer, W.; Izadmehr, S.; Zhou, B.; Shin, A.C.; et al. Blocking FSH induces thermogenic adipose tissue and reduces body fat. Nature 2017, 546, 107–112. [Google Scholar] [CrossRef]
- Guo, Y.; Zhao, M.; Bo, T.; Ma, S.; Yuan, Z.; Chen, W.; He, Z.; Hou, X.; Liu, J.; Zhang, Z.; et al. Blocking FSH inhibits hepatic cholesterol biosynthesis and reduces serum cholesterol. Cell Res. 2019, 29, 151–166. [Google Scholar] [CrossRef]
- Smitz, J.; Wolfenson, C.; Chappel, S.; Ruman, J. Follicle-Stimulating Hormone: A Review of Form and Function in the Treatment of Infertility. Reprod. Sci. 2016, 23, 706–716. [Google Scholar] [CrossRef]
- Stamatiades, G.A.; Carroll, R.S.; Kaiser, U.B. GnRH—A Key Regulator of FSH. Endocrinology 2019, 160, 57–67. [Google Scholar] [CrossRef] [Green Version]
- Liu, F.; Usui, I.; Evans, L.G.; Austin, D.A.; Mellon, P.L.; Olefsky, J.M.; Webster, N.J.G. Involvement of Both Gq/11 and Gs Proteins in Gonadotropin-Releasing Hormone Receptor-Mediated Signaling in Lβ T2 Cells. J. Biol. Chem. 2002, 277, 32099–32108. [Google Scholar] [CrossRef]
- Thompson, I.R.; Ciccone, N.A.; Xu, S.; Zaytseva, S.; Carroll, R.S.; Kaiser, U.B. GnRH Pulse Frequency-Dependent Stimulation of FSHβ Transcription Is Mediated via Activation of PKA and CREB. Mol. Endocrinol. 2013, 27, 606–618. [Google Scholar] [CrossRef]
- Thompson, I.R.; Ciccone, N.A.; Zhou, Q.; Xu, S.; Khogeer, A.; Carroll, R.S.; Kaiser, U.B. GnRH Pulse Frequency Control of Fshb Gene Expression Is Mediated via ERK1/2 Regulation of ICER. Mol. Endocrinol. 2016, 30, 348–360. [Google Scholar] [CrossRef]
- Sun, C.; He, M.; Ko, W.K.; Wong, A.O. Mechanisms for luteinizing hormone induction of growth hormone gene transcription in fish model: Crosstalk of the cAMP/PKA pathway with MAPK-and PI3K-dependent cascades. Mol. Cell. Endocrinol. 2014, 382, 835–850. [Google Scholar] [CrossRef]
- Haisenleder, D.J.; Dalkin, A.C.; Ortolano, G.A.; Marshall, J.C.; Shupnik, M.A. A Pulsatile Gonadotropin-Releasing Hormone Stimulus is Required to Increase Transcription of the Gonadotropin Subunit Genes: Evidence for Differential Regulation of Transcription by Pulse Frequency in Vivo. Endocrinology 1991, 128, 509–517. [Google Scholar] [CrossRef]
- Dalkin, A.C.; Burger, L.L.; Aylor, K.W.; Haisenleder, D.J.; Workman, L.J.; Cho, S.; Marshall, J.C. Regulation of Gonadotropin Subunit Gene Transcription by Gonadotropin-Releasing Hormone: Measurement of Primary Transcript Ribonucleic Acids by Quantitative Reverse Transcription-Polymerase Chain Reaction Assays. Endocrinology 2001, 142, 139–146. [Google Scholar] [CrossRef]
- Kaprara, A.; Huhtaniemi, I.T. The hypothalamus-pituitary-gonad axis: Tales of mice and men. Metabolism 2018, 86, 3–17. [Google Scholar] [CrossRef]
- Hess, G.P.; A Rupley, J. Structure and Function of Proteins. Annu. Rev. Biochem. 1971, 40, 1013–1044. [Google Scholar] [CrossRef]
- Xie, Q.; Tian, Y.; Hu, Z.; Zhang, L.; Tang, B.; Wang, Y.; Li, J.; Chen, G. Novel Translational and Phosphorylation Modification Regulation Mechanisms of Tomato (Solanum lycopersicum) Fruit Ripening Revealed by Integrative Proteomics and Phosphoproteomics. Int. J. Mol. Sci. 2021, 22, 11782. [Google Scholar] [CrossRef]
- Singh, V.; Ram, M.; Kumar, R.; Prasad, R.; Roy, B.K.; Singh, K.K. Phosphorylation: Implications in Cancer. Protein J. 2017, 36, 1–6. [Google Scholar] [CrossRef]
- Wu, M.; Feng, G.; Zhang, B.; Xu, K.; Wang, Z.; Cheng, S.; Chang, C.; Vyas, A.; Tang, Z.; Liu, X. Phosphoproteomics Reveals Novel Targets and Phosphoprotein Networks in Cell Cycle Mediated by Dsk1 Kinase. J. Proteome Res. 2020, 19, 1776–1787. [Google Scholar] [CrossRef]
- Bengoechea-Alonso, M.T.; Ericsson, J. The phosphorylation-dependent regulation of nuclear SREBP1 during mitosis links lipid metabolism and cell growth. Cell Cycle 2016, 15, 2753–2765. [Google Scholar] [CrossRef]
- Zhang, Y.; Yi, F.; Wang, L.; Wang, Z.; Zhang, N.; Wang, Z.; Li, Z.; Song, X.; Wei, S.; Cao, L. Phosphorylation of SMC1A promotes hepatocellular carcinoma cell proliferation and migration. Int. J. Biol. Sci. 2018, 14, 1081–1089. [Google Scholar] [CrossRef]
- De Groof, A.; Ducreux, J.; Aleva, F.; Long, A.J.; Ferster, A.; Van Der Ven, A.; van de Veerdonk, F.; A Houssiau, F.; Lauwerys, B.R. STAT3 phosphorylation mediates the stimulatory effects of interferon alpha on B cell differentiation and activation in SLE. Rheumatology 2020, 59, 668–677. [Google Scholar] [CrossRef]
- Liao, G.; Wang, R.; Tang, D.D. Plk1 Regulates Caspase-9 Phosphorylation at Ser-196 and Apoptosis of Human Airway Smooth Muscle Cells. Am. J. Respir. Cell Mol. Biol. 2022, 66, 223–234. [Google Scholar] [CrossRef]
- Humphrey, S.J.; James, D.E.; Mann, M. Protein Phosphorylation: A Major Switch Mechanism for Metabolic Regulation. Trends Endocrinol. Metab. 2015, 26, 676–687. [Google Scholar] [CrossRef]
- Graves, J.D.; Krebs, E.G. Protein Phosphorylation and Signal Transduction. Pharmacol. Ther. 1999, 82, 111–121. [Google Scholar] [CrossRef]
- Stamatiades, G.A.; Kaiser, U.B. Gonadotropin regulation by pulsatile GnRH: Signaling and gene expression. Mol. Cell. Endocrinol. 2018, 463, 131–141. [Google Scholar] [CrossRef]
- Ferris, H.A.; Shupnik, M.A. Mechanisms for Pulsatile Regulation of the Gonadotropin Subunit Genes by GNRH11. Biol. Reprod. 2006, 74, 993–998. [Google Scholar] [CrossRef]
- Knobil, E. The Neuroendocrine Control of the Menstrual Cycle. Recent Prog. Horm. Res. 1980, 36, 53–88. [Google Scholar] [CrossRef]
- Wildt, L.; Häusler, A.; Marshall, G.; Hutchison, J.S.; Plant, T.M.; Belchetz, P.E.; Knobil, E. Frequency and Amplitude of Gonadotropin-Releasing Hormone Stimulation and Gonadotropin Secretion in the Rhesus Monkey. Endocrinology 1981, 109, 376–385. [Google Scholar] [CrossRef]
- Wang, H.-Q.; Wang, W.-H.; Chen, C.-Z.; Guo, H.-X.; Jiang, H.; Yuan, B.; Zhang, J.-B. Regulation of FSH Synthesis by Differentially Expressed miR-488 in Anterior Adenohypophyseal Cells. Animals 2021, 11, 3262. [Google Scholar] [CrossRef]
- Lambert, N.A. Dissociation of Heterotrimeric G Proteins in Cells. Sci. Signal. 2008, 1, re5. [Google Scholar] [CrossRef]
- Oldham, W.M.; Hamm, H.E. Heterotrimeric G protein activation by G-protein-coupled receptors. Nat. Rev. Mol. Cell Biol. 2008, 9, 60–71. [Google Scholar] [CrossRef]
- Davignon, I.; Barnard, M.; Gavrilova, O.; Sweet, K.; Wilkie, T.M. Gene Structure of Murine Gna11 and Gna15: Tandemly Duplicated Gq Class G Protein α Subunit Genes. Genomics 1996, 31, 359–366. [Google Scholar] [CrossRef]
- Johnson, G.J.; Leis, L.A.; Dunlop, P.C. Specificity of Gαq and Gα11 gene expression in platelets and erythrocytes. Expressions of cellular differentiation and species differences. Biochem. J. 1996, 318, 1023–1031. [Google Scholar] [CrossRef]
- Coussens, L.; Parker, P.J.; Rhee, L.; Yang-Feng, T.L.; Chen, E.; Waterfield, M.D.; Francke, U.; Ullrich, A. Multiple, Distinct Forms of Bovine and Human Protein Kinase C Suggest Diversity in Cellular Signaling Pathways. Science 1986, 233, 859–866. [Google Scholar] [CrossRef]
- Dempsey, E.C.; Newton, A.C.; Mochly-Rosen, D.; Fields, A.P.; Reyland, M.E.; Insel, P.A.; Messing, R.O. Protein kinase C isozymes and the regulation of diverse cell responses. Am. J. Physiol. Lung Cell. Mol. Physiol. 2000, 279, L429–L438. [Google Scholar] [CrossRef]
- Ma, Y.; Usuwanthim, K.; Munawara, U.; Quach, A.; Gorgani, N.N.; Abbott, C.A.; Hii, C.S.; Ferrante, A. Protein Kinase Cα Regulates the Expression of Complement Receptor Ig in Human Monocyte—Derived Macrophages. J. Immunol. 2015, 194, 2855–2861. [Google Scholar] [CrossRef]
- Taylor, K.D.; Pikó, L. Expression of ribosomal protein genes in mouse oocytes and early embryos. Mol. Reprod. Dev. 1992, 31, 182–188. [Google Scholar] [CrossRef]
- Chakraborty, A.; Uechi, T.; Higa, S.; Torihara, H.; Kenmochi, N. Loss of Ribosomal Protein L11 Affects Zebrafish Embryonic Development through a p53-Dependent Apoptotic Response. PLoS ONE 2009, 4, e4152. [Google Scholar] [CrossRef]
- Kirn-Safran, C.B.; Oristian, D.S.; Focht, R.J.; Parker, S.G.; Vivian, J.L.; Carson, D.D. Global growth deficiencies in mice lacking the ribosomal protein HIP/RPL29. Dev. Dyn. 2007, 236, 447–460. [Google Scholar] [CrossRef]
- Tu, C.; Meng, L.; Nie, H.; Yuan, S.; Wang, W.; Du, J.; Lu, G.; Lin, G.; Tan, Y.-Q. A homozygous RPL10L missense mutation associated with male factor infertility and severe oligozoospermia. Fertil. Steril. 2020, 113, 561–568. [Google Scholar] [CrossRef]
- Sugihara, Y.; Sadohara, E.; Yonezawa, K.; Kugo, M.; Oshima, K.; Matsuda, T.; Nadano, D. Identification and expression of an autosomal paralogue of ribosomal protein S4, X-linked, in mice: Potential involvement of testis-specific ribosomal proteins in translation and spermatogenesis. Gene 2013, 521, 91–99. [Google Scholar] [CrossRef]
- Zou, Q.; Yang, L.; Shi, R.; Qi, Y.; Zhang, X.; Qi, H. Proteostasis regulated by testis-specific ribosomal protein RPL39L maintains mouse spermatogenesis. Iscience 2021, 24, 103396. [Google Scholar] [CrossRef]
- Li, H.; Zhang, H.; Huang, G.; Dou, Z.; Xie, Y.; Si, J.; Di, C. Heavy ion radiation-induced DNA damage mediates apoptosis via the Rpl27a-Rpl5-MDM2-p53/E2F1 signaling pathway in mouse spermatogonia. Ecotoxicol. Environ. Saf. 2020, 201, 110831. [Google Scholar] [CrossRef]
- Liu, Z.; Lv, J.; Liu, Y.; Wang, J.; Zhang, Z.; Chen, W.; Song, J.; Yang, B.; Tan, F.; Zou, X.; et al. Comprehensive Phosphoproteomic Analysis of Pepper Fruit Development Provides Insight into Plant Signaling Transduction. Int. J. Mol. Sci. 2020, 21, 1962. [Google Scholar] [CrossRef]
- Ma, Q.; Wu, M.; Pei, W.; Li, H.; Li, X.; Zhang, J.; Yu, J.; Yu, S. Quantitative phosphoproteomic profiling of fiber differentiation and initiation in a fiberless mutant of cotton. BMC Genom. 2014, 15, 466. [Google Scholar] [CrossRef]
- Bentem, S.D.L.F.V.; Hirt, H. Protein tyrosine phosphorylation in plants: More abundant than expected? Trends Plant Sci. 2009, 14, 71–76. [Google Scholar] [CrossRef]
- Wang, H.; Zhang, J.; Zheng, Y.; Zhang, W.; Guo, H.; Cong, S.; Ding, Y.; Yuan, B. Comprehensive analysis of differences in N6-methyladenosine RNA methylomes in the rat adenohypophysis after GnRH treatment. FASEB J. 2022, 36, e22204. [Google Scholar] [CrossRef]
- Perez-Riverol, Y.; Bai, J.; Bandla, C.; García-Seisdedos, D.; Hewapathirana, S.; Kamatchinathan, S.; Kundu, D.J.; Prakash, A.; Frericks-Zipper, A.; Eisenacher, M.; et al. The PRIDE database resources in 2022: A hub for mass spectrometry-based proteomics evidences. Nucleic Acids Res. 2022, 50, D543–D552. [Google Scholar] [CrossRef]










Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, T.; Wang, H.-Q.; Yuan, B.; Zhao, G.-K.; Ma, Y.-R.; Zhao, P.-S.; Xie, W.-Y.; Gao, F.; Gao, W.; Ren, W.-Z. Integrative Proteomics and Phosphoproteomics Analysis of the Rat Adenohypophysis after GnRH Treatment. Int. J. Mol. Sci. 2023, 24, 3339. https://doi.org/10.3390/ijms24043339
Wang T, Wang H-Q, Yuan B, Zhao G-K, Ma Y-R, Zhao P-S, Xie W-Y, Gao F, Gao W, Ren W-Z. Integrative Proteomics and Phosphoproteomics Analysis of the Rat Adenohypophysis after GnRH Treatment. International Journal of Molecular Sciences. 2023; 24(4):3339. https://doi.org/10.3390/ijms24043339
Chicago/Turabian StyleWang, Tian, Hao-Qi Wang, Bao Yuan, Guo-Kun Zhao, Yi-Ran Ma, Pei-Sen Zhao, Wen-Yin Xie, Fei Gao, Wei Gao, and Wen-Zhi Ren. 2023. "Integrative Proteomics and Phosphoproteomics Analysis of the Rat Adenohypophysis after GnRH Treatment" International Journal of Molecular Sciences 24, no. 4: 3339. https://doi.org/10.3390/ijms24043339