
Overexpression of the Liriodendron tulipifera BOP2 Gene (LtuBOP2) Affects Leaf Margin Development in Transgenic Arabidopsis thaliana


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
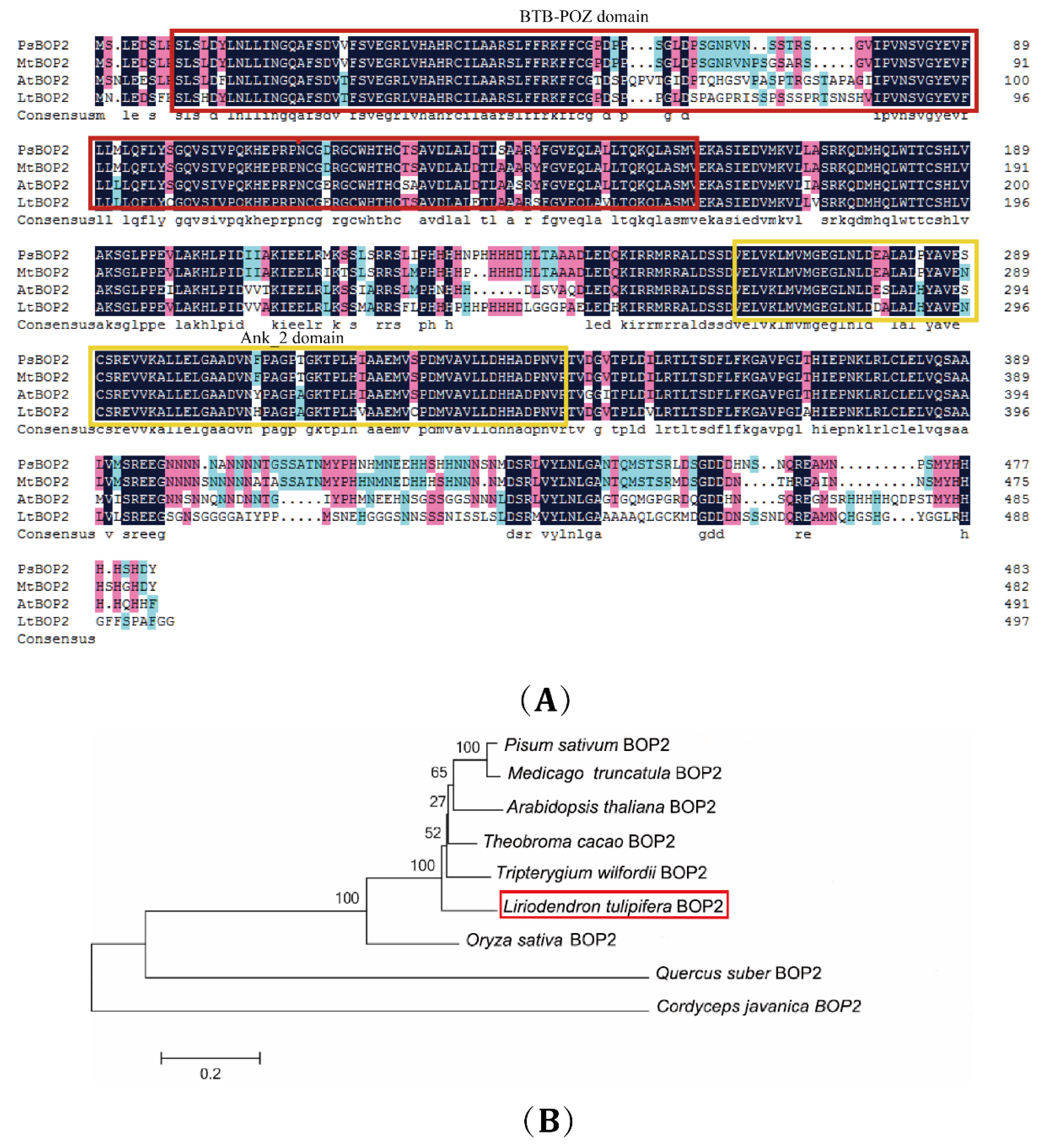
2.1. Cloning and Bioinformatics Analysis of LtuBOP2
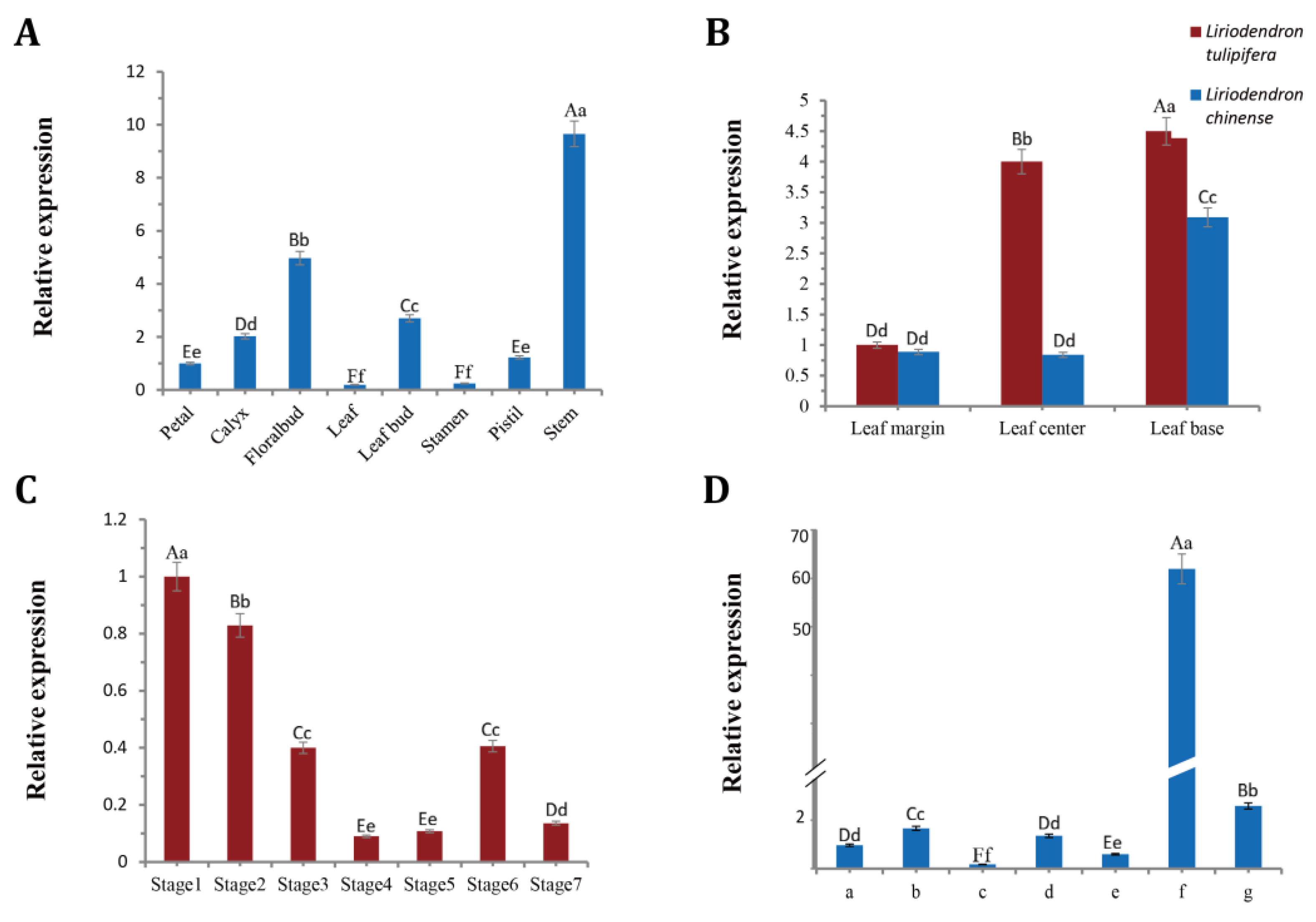
2.2. Spatiotemporal Expression Pattern of LtuBOP2
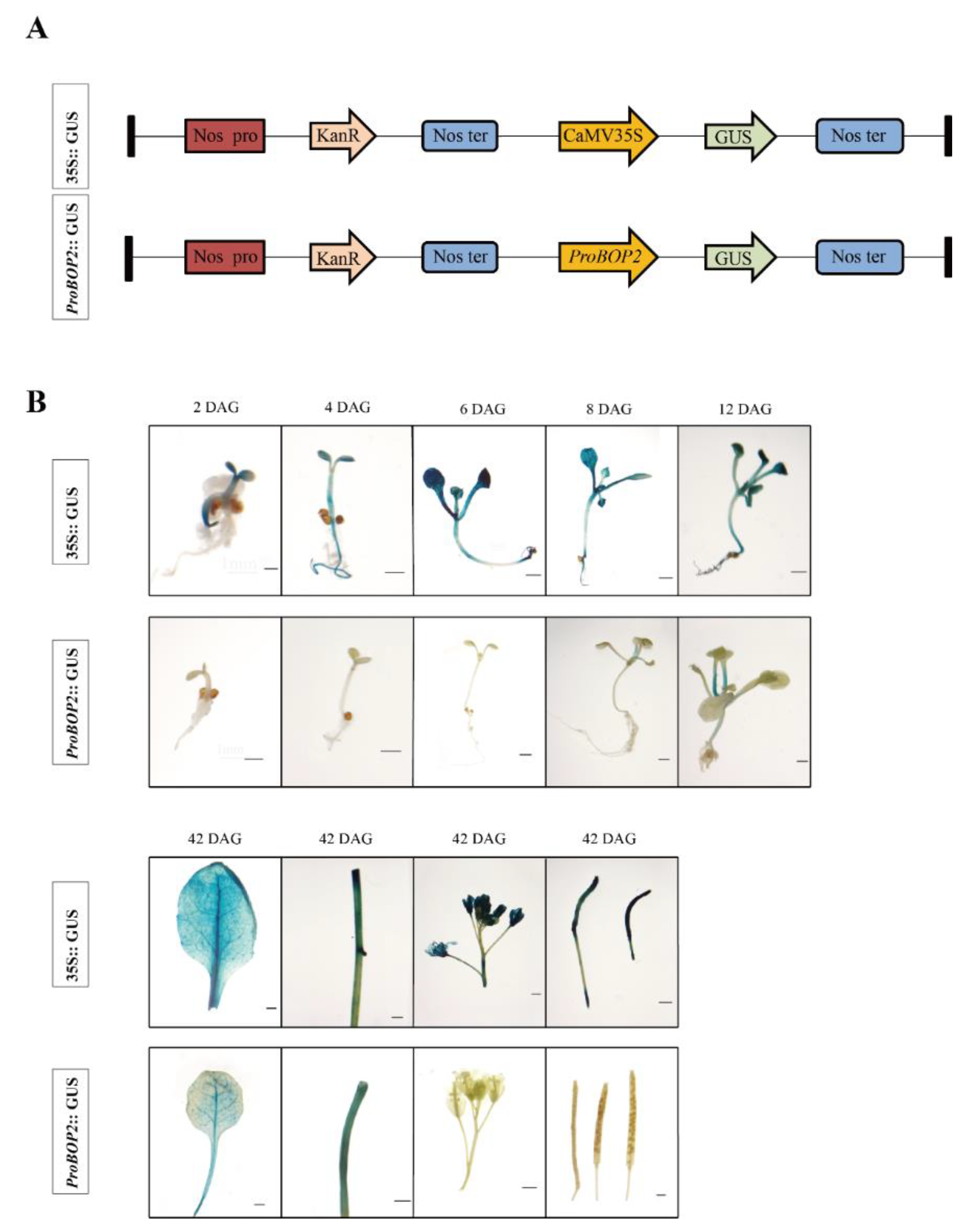
2.3. pLtuBOP2 Sequence Analysis
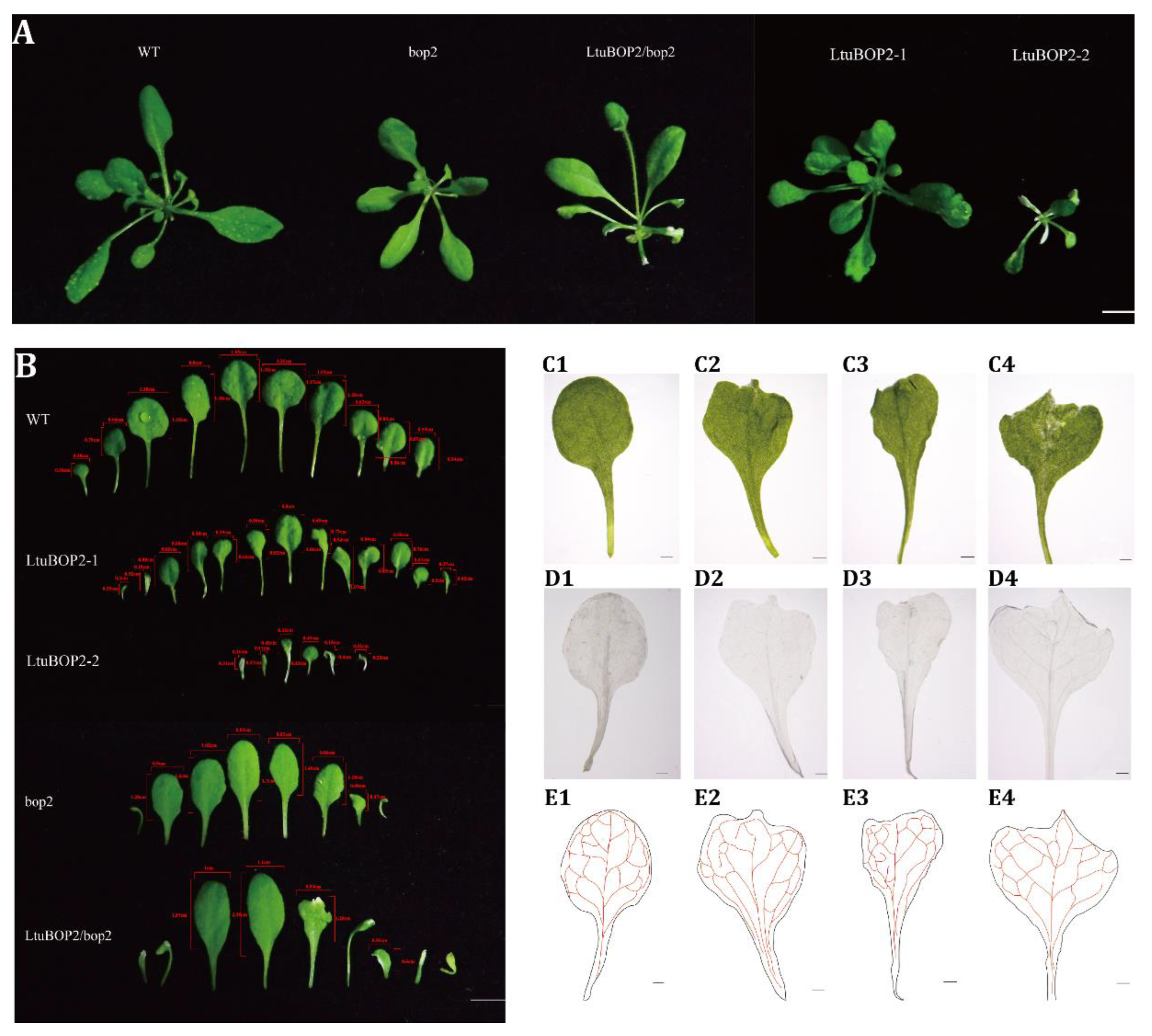
2.4. Phenotypic Analysis of LtuBOP2 Transgenic Plants
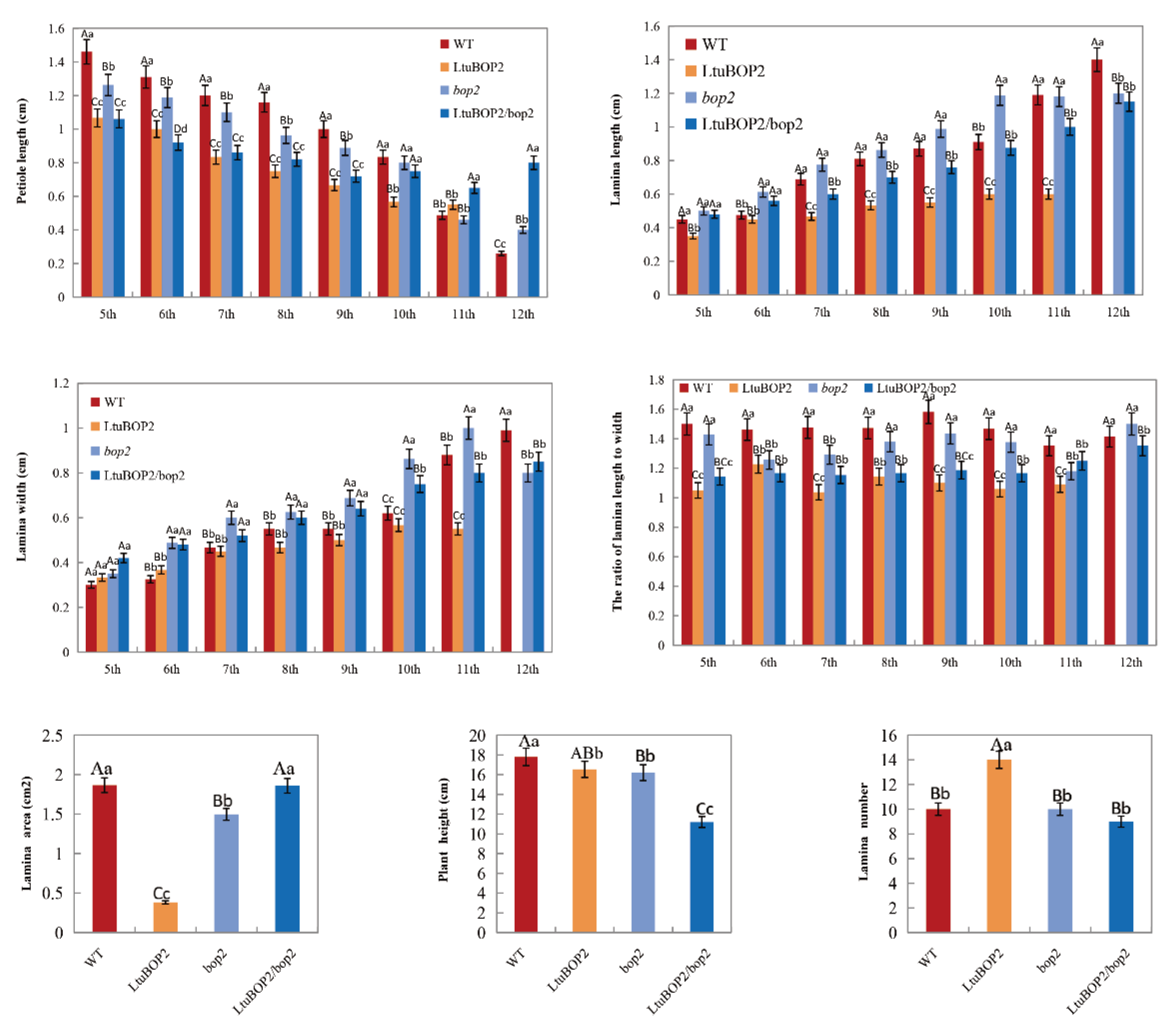
2.5. Leaf Morphology of Transgenic A. thaliana Lines
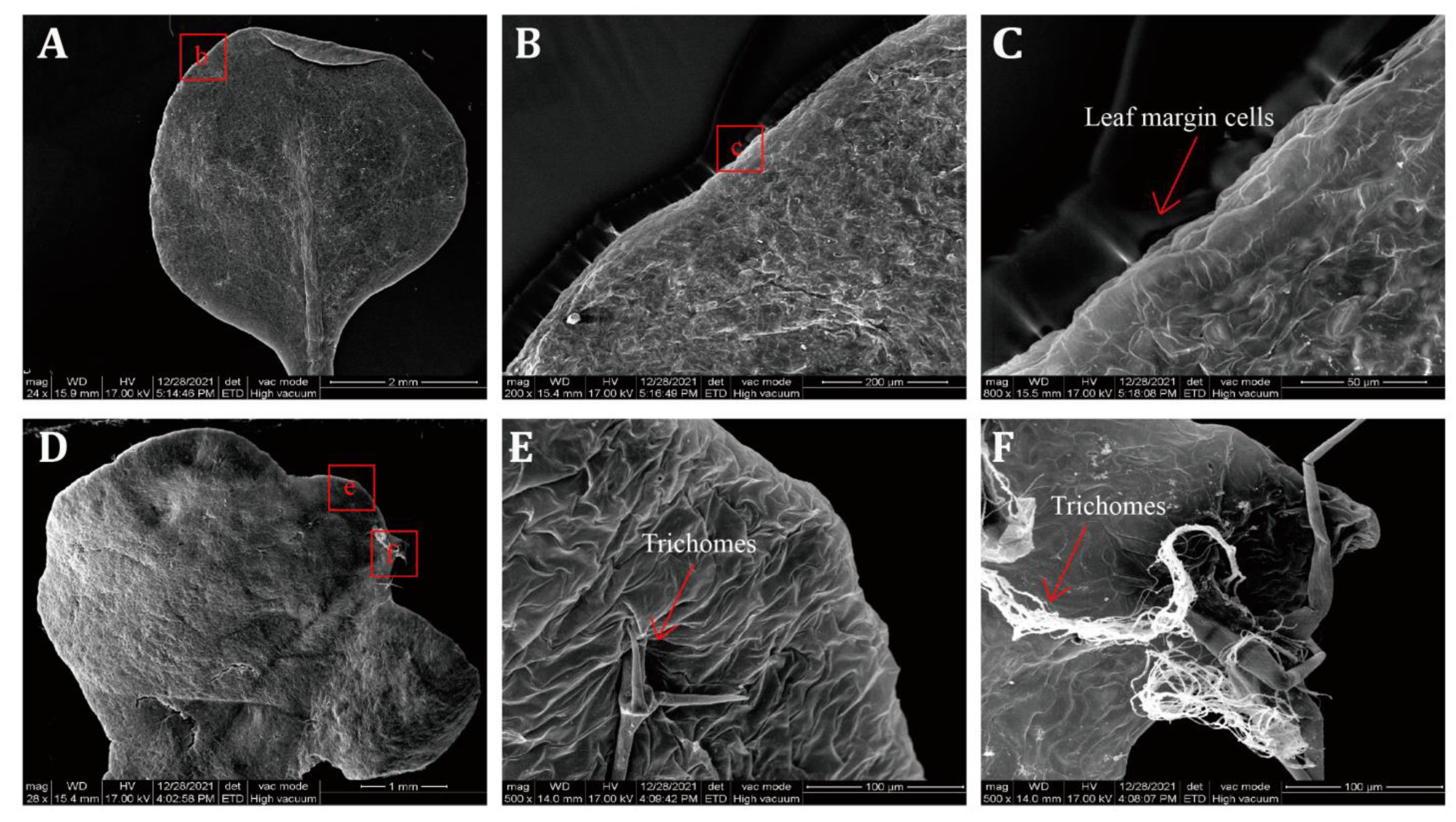
2.6. SEM Observation of Transgenic A. thaliana Lines
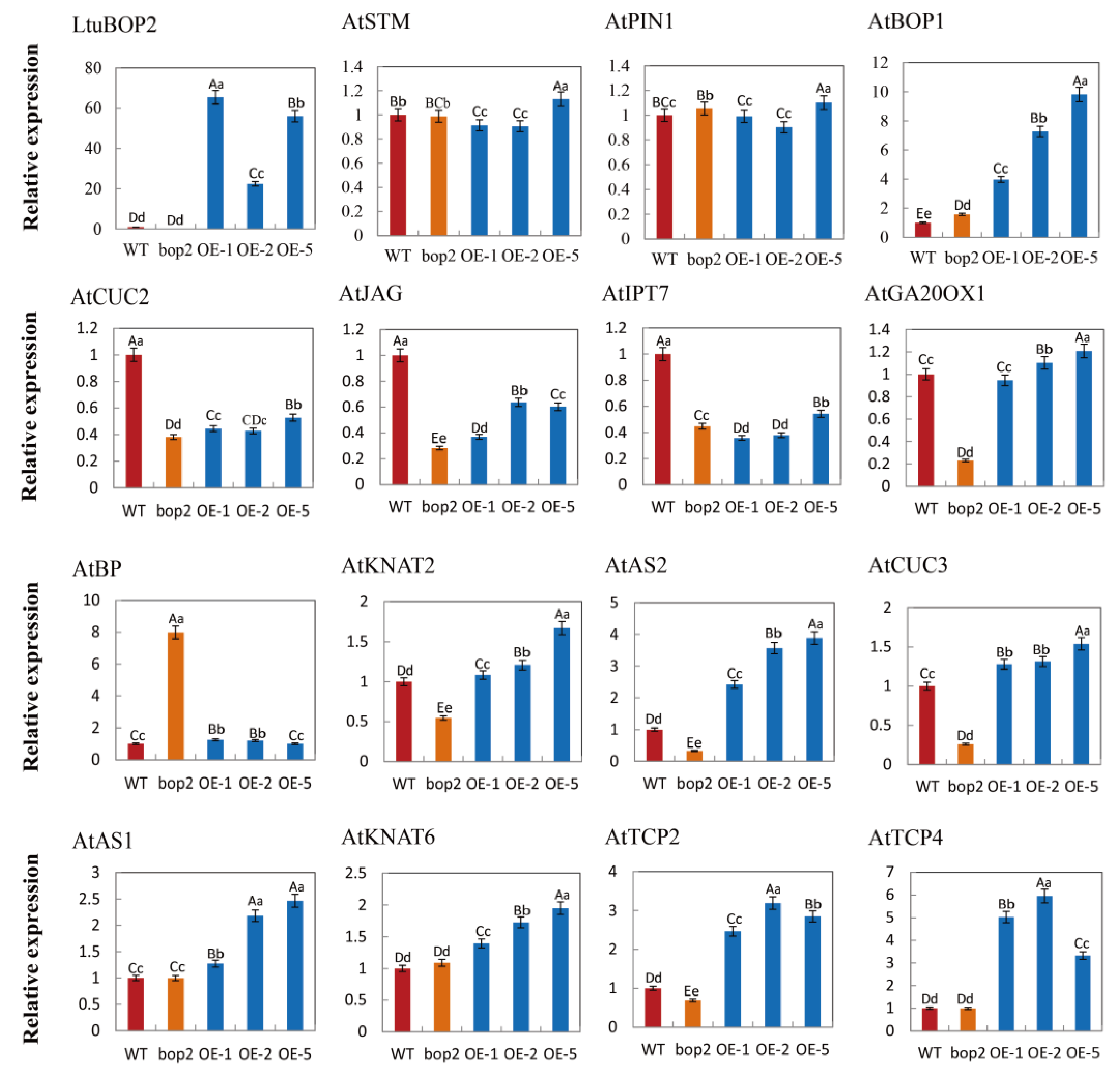
2.7. LtuBOP2 Affects the Expression of Genes Related to Leaf Shape Development
3. Discussion
3.1. Function of Domain in the LtuBOP2 Gene
3.2. Expression Pattern of LtuBOP2 during Growth and Development
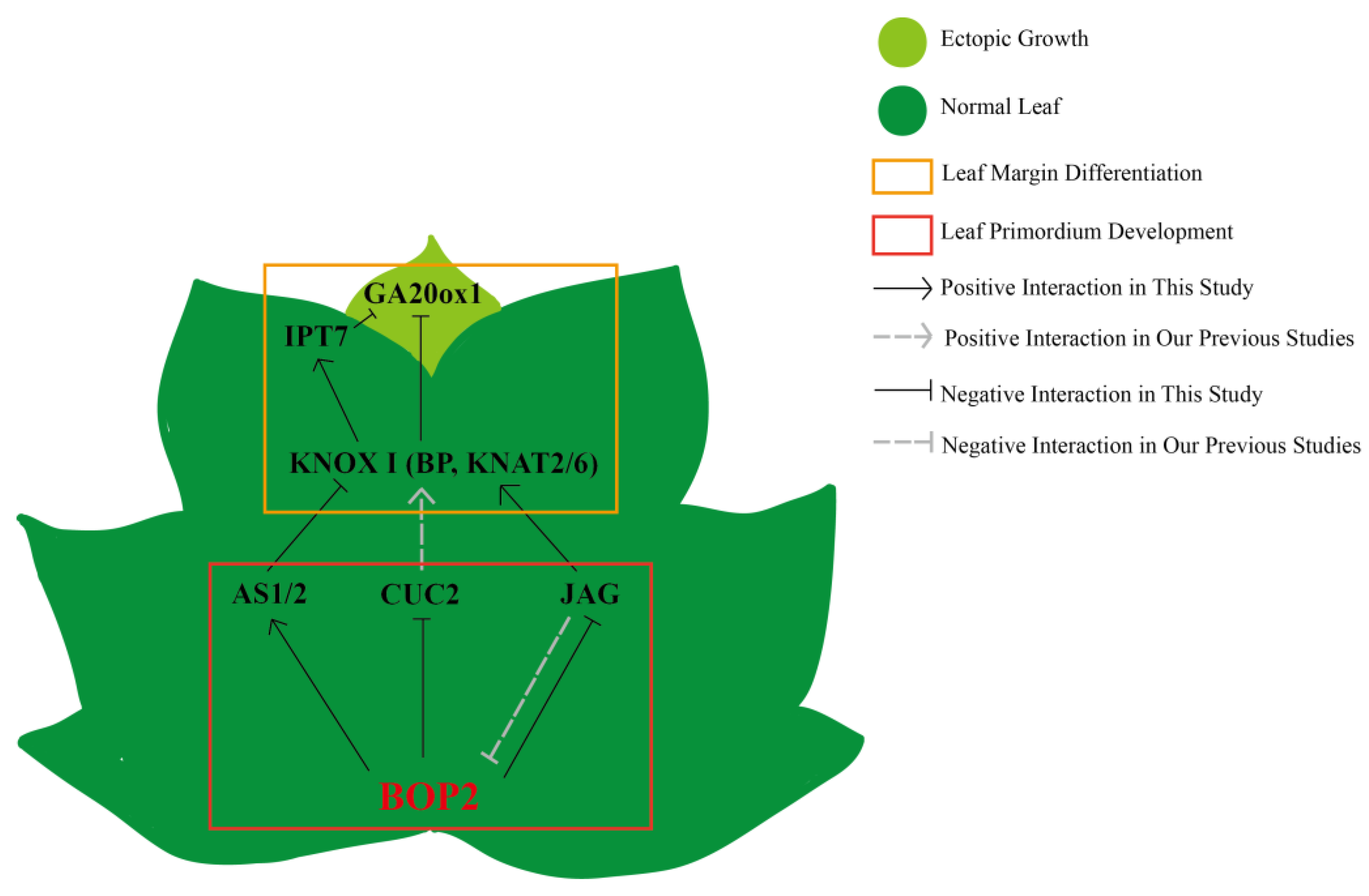
3.3. Roles of LtuBOP2 during Leaf Margin Formation
3.4. BOP2 Affects the Expression of Genes Related to Leaf Development
4. Materials and Methods
4.1. Plant Materials
4.2. cDNA Synthesis, Sequencing, and Sequence Analysis of LtuBOP2
4.3. Recombinant Plasmid Construction and Genetic Transformation
4.4. RT-qPCR
4.5. SEM Observation
4.6. Phenotypic Characterization and Statistical Analysis
4.7. GUS Histochemical Assay and Venation Pattern Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Tsukaya, H. Mechanism of leaf-shape determination. Annu. Rev. Plant Biol. 2006, 57, 477–496. [Google Scholar] [CrossRef] [PubMed]
- Li, L.C.; Kang, D.M.; Chen, Z.L.; Qu, L.J. Hormonal regulation of leaf morphogenesis in Arabidopsis. J. Integr. Plant Bior. 2007, 49, 75–80. [Google Scholar] [CrossRef]
- Nicotra, A.B.; Leigh, A.; Boyce, C.K.; Jones, C.S.; Niklas, K.J.; Royer, D.L.; Tsukaya, H. The evolution and functional significance of leaf shape in the angiosperms. Funct. Plant Biol. 2011, 38, 535–552. [Google Scholar] [CrossRef] [PubMed]
- Vogel, S. Leaves in the lowest and highest winds: Temperature, force and shape. New Phytol. 2009, 183, 13–26. [Google Scholar] [CrossRef] [PubMed]
- Semchenko, M.; Zobel, K. The role of leaf lobation in elongation responses to shade in the rosette-forming forb Serratula tinctoria (Asteraceae). Ann. Bot. 2007, 100, 83–90. [Google Scholar] [CrossRef]
- Dengler, N.G.; Tsukaya, H. Leaf morphogenesis in dicotyledons. Int. J. Plant Sci. 2001, 162, 459–464. [Google Scholar] [CrossRef]
- Truskina, J.; Vernoux, T. The growth of a stable stationary structure: Coordinating cell behavior and patterning at the shoot apical meristem. Curr. Opin. Plant Biol. 2018, 41, 83–88. [Google Scholar] [CrossRef]
- Ichihashi, Y.; Tsukaya, H. Behavior of leaf meristems and their modification. Front. Plant Sci. 2015, 6, 1060. [Google Scholar] [CrossRef]
- Shuai, B.; Reynaga-Peña, C.G.; Springer, P.S. The lateral organ boundaries gene defines a novel, plant-specific gene family. Plant Physiol. 2002, 129, 747–761. [Google Scholar] [CrossRef]
- Waites, R.; Hudson, A. Phantastica: A gene required for dorsoventrality of leaves in Antirrhinum Majus. Dev. 1995, 121, 2143–2154. [Google Scholar] [CrossRef]
- Barton, M.K. Twenty years on: The inner workings of the shoot apical meristem, a developmental dynamo. Develop. Biol. 2009, 341, 95–113. [Google Scholar] [CrossRef]
- Du, F.; Guan, C.; Jiao, Y.L. Molecular mechanisms of leaf morphogenesis. Mol. Plant 2018, 11, 1117–1134. [Google Scholar] [CrossRef]
- Idan, E.; Yuval, E.; Eliezer, L. Morphogenesis of simple and compound leaves: A critical review. Plant Cell 2010, 22, 1019–1032. [Google Scholar]
- Moon, J.; Hake, S. How a leaf gets its shape? Curr. Opin. Plant Biol. 2010, 14, 24–30. [Google Scholar] [CrossRef]
- Holtan, H.E.E.; Hake, S. Quantitative trait locus analysis of leaf dissection in tomato using Lycopersicon pennellii segmental introgression lines. Genetics 2003, 165, 1541–1550. [Google Scholar] [CrossRef]
- Sisó, S.; Camarero, J.; Gil-Pelegrín, E. Relationship between hydraulic resistance and leaf morphology in broad leaf Quercus species: A new interpretation of leaf lobation. Trees 2001, 15, 341–345. [Google Scholar] [CrossRef]
- Kidner, C.; Umbreen, S. Why is leaf shape so variable? Int. J. Plant Develop. Bior. 2010, 4, 64–75. [Google Scholar]
- Byrne, M.E.; Simorowski, J.; Martienssen, R.A. ASYMMETRIC LEAVES1 reveals knox gene redundancy in A.thaliana. Development 2002, 129, 1957–1965. [Google Scholar] [CrossRef]
- Wang, Q.; Hasson, A.; Rossmann, S.; Theres, K. Divide at impera: Boundaries shape the plantbody and initiate new meristems. New Phytol. 2016, 209, 485–498. [Google Scholar] [CrossRef]
- Furutani, M.; Vernoux, T.; Traas, J.; Kato, T.; Tasaka, M.; Aida, M. PIN-FORMED1 and PINOID regulate boundary formation and cotyledon development in A.thaliana embryogenesis. Development 2004, 131, 5021–5030. [Google Scholar] [CrossRef]
- Dinneny, J.R.; Yadegari, R.; Fischer, R.L.; Yanofsky, M.F.; Weigel, D. The role of JAGGED in shaping lateral organs. Development 2004, 131, 1101–1110. [Google Scholar] [CrossRef] [PubMed]
- Wei, L.; Wen, S.; Tu, Z.; Zhao, Y.; Li, H. Overexpression of Liriodendron tulipifera JAG Gene (LtuJAG) changes leaf shapes in transgenic A. thaliana. Int. J. Mol. Sci. 2022, 23, 1322. [Google Scholar] [CrossRef] [PubMed]
- Ha, C.M.; Jun, J.H.; Nam, H.G.; Fletcher, J.C. BLADE-ON-PETIOLE 1 and 2 control A.thaliana lateral organ fate through regulation of LOB domain and adaxial-abaxial polarity genes. Plant Cell 2007, 19, 1809–1825. [Google Scholar] [CrossRef] [PubMed]
- Jun, J.H.; Ha, C.M.; Fletcher, J.C. BLADE-ON-PETIOLE1 coordinates organ determinacy and axial polarity in arabidopsis by directly activating ASYMMETRIC LEAVES2. Plant Cell 2010, 22, 62–76. [Google Scholar] [CrossRef] [PubMed]
- Couzigou, J.M.; Zhukov, V.; Mondy, S.; Heba, G.A.; Cosson, V.; Noel, E.T.H.; Ambrose, M.; Wen, J.Q.; Tadege, M.; Tikhonovich, I.; et al. NODULE ROOT and COCHLEATA maintain nodule development and are legume orthologs of A.thaliana BLADE-ON-PETIOLE genes. Plant Cell 2012, 24, 4498–4510. [Google Scholar] [CrossRef]
- Yasunori, I.; José, A.A.; Moran, F.; Daniel, H.C.; Ravi, K.; Lee, V.M.; Jie, P.; Julin, N.M.; Neelima, R.S. Evolutionary developmental transcriptomics reveals a gene network module regulating interspecific diversity in plant leaf shape. Proc. Nat. Acad. Sci. USA 2014, 111, E2616–E2621. [Google Scholar]
- Chen, Y.N.; Wei, S.Y.; Qu, G.Z.; Hu, J.J.; Wang, J.H.; Yin, T.M.; Pan, H.X.; Lu, M.Z.; Kang, X.Y.; Li, L.G.; et al. The key and core technologies for accelerating the tree breeding process. J. Nanjing For. Univ. (Nat. Sci. Ed.) 2022, 46, 1–9. [Google Scholar]
- Yang, Y.; Li, H.G. Cloning of NAC gene from Liriodendron tulipifera and its expression analysis. J. Plant Resour. Environ. 2015, 24, 1–9. [Google Scholar]
- Ma, J.K.; Wei, L.M.; Li, H.G. The analysis of genes and phytohormone metabolic pathways associated with leaf shape development in Liriodendron chinense via De Novo transcriptome sequencing. Genes 2018, 9, 577. [Google Scholar] [CrossRef]
- Ma, J.K.; Mei, G.G.; Liu, H.H.; Li, H.G. Overexpression of a novel LcKNOX transcription factor from Liriodendron chinense induces lobed leaves in A.thaliana. Forests 2019, 11, 33. [Google Scholar] [CrossRef]
- Li, J.Y. Cloning and Functional Analysis of LcCUC Genes from Liriodendron chinense. Master’s Thesis, NanJing Forestry University, NanJing, China, 2019. [Google Scholar]
- Wen, S.Y.; Li, J.Y.; Hao, Z.Y.; Wei, L.M.; Ma, J.K.; Zong, Y.X.; Li, H.G. Overexpression of the LcCUC2-like gene in A.thaliana alters the cotyledon morphology and increases rosette leaf number. Peer J. 2022, 10, e12615. [Google Scholar] [CrossRef]
- Dey, N.; Sarkar, S.; Acharya, S.; Maiti, I.B. Synthetic promoters in planta. Planta 2015, 242, 1077–1094. [Google Scholar] [CrossRef]
- Xu, C.; Park, S.J.; Eck, J.V.; Lippman, Z.B. Control of inflorescence architecture in tomato by BTB/POZ transcriptional regulators. Gene. Develop. 2016, 30, 2048–2061. [Google Scholar] [CrossRef]
- Ha, C.M.; Jun, J.H.; Nam, H.G.; Fletcher, J.C. BLADE-ON-PETIOLE1 encodes a BTB/POZ domain protein required for leaf morphogenesis in Arabidopsis thaliana. Plant Cell Physiol. 2004, 45, 1361–1370. [Google Scholar] [CrossRef]
- Tian, C.H.; Zhang, X.N.; He, J.; Yu, H.P.; Wang, Y.; Shi, B.H.; Han, Y.Y.; Wang, G.X.; Feng, X.M.; Zhang, C.; et al. An organ boundary-enriched gene regulatory network uncovers regulatory hierarchies underlying axillary meristem initiation. Mol. Systems Bio. 2014, 755, 1744–4292. [Google Scholar] [CrossRef]
- Norberg, M.; Holmlund, M.; Nilsson, O. The BLADE-ON-PETIOLE genes act redundantly to control the growth and development of lateral organs. Development 2005, 132, 2203–2213. [Google Scholar] [CrossRef]
- Khan, M.; Xu, H.S.; Hepworth, S.R. BLADE-ON-PETIOLE genes: Setting boundaries in development and defense. Plant Sci. 2014, 216, 157–171. [Google Scholar] [CrossRef]
- Blein, T.; Pulido, A.; Vialette-Guiraud, A.; Nikovics, K.; Morin, H.; Hay, A.; Johansen, I.E.; Tsiantis, M.; Laufs, P. A conserved molecular framework for compound leaf development. Science 2008, 322, 1835–1839. [Google Scholar] [CrossRef]
- Rossmann, S.; Kohlen, W.; Hasson, A.; Theres, K. Lateral suppressor and Goblet act in hierarchical order to regulate ectopic meristem formation at the base of tomato leaflets. Plant J. 2015, 81, 837–848. [Google Scholar] [CrossRef]
- Ha, C.M.; Kim, G.T.; Kim, B.C.; Jun, J.H.; Soh, M.S.; Ueno, Y.; Machida, Y.; Tsukaya, H.; Nam, H.G. The BLADE-ON-PETIOLE 1 gene controls leaf pattern formation through the modulation of meristematic activity in Arabidopsis. Development 2003, 130, 161–172. [Google Scholar] [CrossRef]
- Toriba, T.; Tokunaga, H.; Shiga, T.; Nie, F.; Naramoto, S.; Honda, E.; Tanaka, K.; Taji, T.; Itoh, J.I.; Kyozuka, J. BLADE-ON-PETIOLE genes temporally and developmentally regulate the sheath to blade ratio of rice leaves. Nat. Commun. 2019, 10, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Kawamura, E.; Horiguchi, G.; Tsukaya, H. Mechanisms of leaf tooth formation in Arabidopsis. Plant J. 2010, 62, 429–441. [Google Scholar] [CrossRef] [PubMed]
- Scofield, S.; Murray, J. KNOX gene function in plant stem cell niches. Plant Mol. Biol. 2006, 60, 929–946. [Google Scholar] [CrossRef] [PubMed]
- Aida, M.; Ishida, T.; Fukaki, H.; Fujisawa, H.; Tasaka, M. Genes involved in organ separation in Arabidopsis: An analysis of the cup-shaped cotyledon mutant. Plant Cell 1997, 9, 841–857. [Google Scholar] [CrossRef] [PubMed]
- Yadav, R.K.; Tavakkoli, M.; Reddy, G.V. WUSCHEL mediates stem cell homeostasis by regulating stem cell number and patterns of cell division and differentiation of stem cell progenitors. Development 2010, 137, 3581–3589. [Google Scholar] [CrossRef] [Green Version]
- Byrne, M.E.; Barley, R.; Curtis, M.; Arroyo, J.M.; Dunham, M.; Hudson, A.; Martienssen, R.A. Asymmetric leaves1 mediates leaf patterning and stem cell function in Arabidopsis. Nature 2000, 408, 967–971. [Google Scholar] [CrossRef]
- Busch, B.L.; Schmitz, G.; Rossmann, S.; Piron, F.; Ding, J.; Bendahmane, A.; Theres, K. Shoot branching and leaf dissection in tomato are regulated by homologous gene modules. Plant Cell 2011, 23, 3595–3609. [Google Scholar] [CrossRef]
- Aida, M.; Tasaka, M. Genetic control of shoot organ boundaries. Curr. Opin. Plant Bio. 2006, 9, 72–77. [Google Scholar] [CrossRef]
- Xu, M.; Hu, T.; McKim, S.M.; Murmu, J.; Haughn, G.W.; Hepworth, S.R. Arabidopsis BLADE-ON-PETIOLE1 and 2 promote floral meristem fate and determinacy in a previously undefined pathway targeting APETALA1 and AGAMOUS-LIKE24. Plant J. 2010, 63, 974–989. [Google Scholar] [CrossRef]
- Jasinski, S.; Piazza, P.; Craft, J.; Hay, A.; Woolley, L.; Rieu, I.; Phillips, A.; Hedden, P.; Tsiantis, M. KNOX Action in Arabidopsis Is Mediated by Coordinate Regulation of Cytokinin and Gibberellin Activities. Curr. Biol. 2005, 15, 1560–1565. [Google Scholar] [CrossRef]
- Li, H.G.; Chen, L.; Liang, C.Y. A case study on provenance testing of tulip tree (Liriodendron spp.). China For. Sci. Technol. 2005, 5, 13–16. [Google Scholar]
- Jia, Z.H.; Jia, X.D.; Xu, M.Y.; Mo, Z.H.; Zhai, M.; Xuan, J.P.; Zhang, J.Y.; Wang, G.; Wang, T.; Guo, Z.R. Cloning and expression analyses of the key enzyme gene of procyanidins biosynthesis in pecan (Carya illinoinensis). J. Nanjing For. Univ. (Nat. Sci. Ed.) 2022, 46, 49–57. [Google Scholar]
- He, Q.Q.; Liu, C.Q.; Li, J.J.; Wang, J.J.; Yao, X.; Zhou, S.H.; Chen, Y.; Wang, H.R. Cloning of EoNLA gene in Eremochloa ophiuroides and the transgenic Arabidopsis phenotypic characterization under various phosphorus levels. J. Nanjing For. Univ. (Nat. Sci. Ed.) 2022, 46, 134–142. [Google Scholar]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular evolutionary genetics analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef]
- Clough, S.J.; Bent, A.F. Floral dip: A simplified method for Agrobacterium-mediated transformation of A.thaliana. Plant J. 1998, 16, 735–743. [Google Scholar] [CrossRef]
- Lin, L.L.; Hu, A.Q.; Chen, G.; Zhang, J.Y.; Cao, G.Q.; Cao, S.J. Cloning and expression characteristics of ClWRKY44 gene from Cunninghamia lanceolata. J. Nanjing For. Univ. (Nat. Sci. Ed.) 2022, 46, 203–209. [Google Scholar]
- Tu, Z.H.; Hao, Z.Y.; Zhong, W.; Li, H. Identification of suitable reference genes for RT-qPCR assays in Liriodendron chinense (Hemsl.) Sarg. Forests 2019, 10, 441–456. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Zhang, Q.; Qian, T.; Wang, H.; Zhu, M.W.; Li, S.X. Hardness breaking and mechanisms of water absorption in Cercis canadensis seeds. J. Nanjing For. Univ. (Nat. Sci. Ed.) 2021, 45, 137–142. [Google Scholar]
- Hepworth, S.R.; Zhang, Y.; McKim, S.; Li, X.; Haughn, G.W. BLADE-ONPETIOLE dependent signaling controls leaf and floral patterning in Arabidopsis. Plant Cell 2005, 17, 1434–1448. [Google Scholar] [CrossRef]
- Luo, M.; Yu, C.W.; Chen, F.F.; Zhao, L.; Tian, G.; Liu, X.; Cui, Y.; Yang, J.Y.; Wu, K. Histone deacetylase HDA6 is functionally associated with AS1 in repression of KNOX genes in A.thaliana. PLoS Genet. 2012, 8, e1003114. [Google Scholar] [CrossRef] [PubMed]
- Jefferson, R.A.; Kavanagh, T.A.; Bevan, M.W. GUS fusions: Beta-glucuronidase as a sensitive and versatile gene fusion marker in higher plants. EMBO J. 1987, 6, 3901–3907. [Google Scholar] [CrossRef] [PubMed]








Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhao, Y.; Wei, L.; Wen, S.; Li, H. Overexpression of the Liriodendron tulipifera BOP2 Gene (LtuBOP2) Affects Leaf Margin Development in Transgenic Arabidopsis thaliana. Int. J. Mol. Sci. 2023, 24, 3262. https://doi.org/10.3390/ijms24043262
Zhao Y, Wei L, Wen S, Li H. Overexpression of the Liriodendron tulipifera BOP2 Gene (LtuBOP2) Affects Leaf Margin Development in Transgenic Arabidopsis thaliana. International Journal of Molecular Sciences. 2023; 24(4):3262. https://doi.org/10.3390/ijms24043262
Chicago/Turabian StyleZhao, Yanqing, Lingmin Wei, Shaoying Wen, and Huogen Li. 2023. "Overexpression of the Liriodendron tulipifera BOP2 Gene (LtuBOP2) Affects Leaf Margin Development in Transgenic Arabidopsis thaliana" International Journal of Molecular Sciences 24, no. 4: 3262. https://doi.org/10.3390/ijms24043262