
Impact of Salinity on the Energy Transfer between Pigment–Protein Complexes in Photosynthetic Apparatus, Functions of the Oxygen-Evolving Complex and Photochemical Activities of Photosystem II and Photosystem I in Two Paulownia Lines
 ,
,  , and
, and 
Abstract
:1. Introduction
2. Results
2.1. Pigment Composition
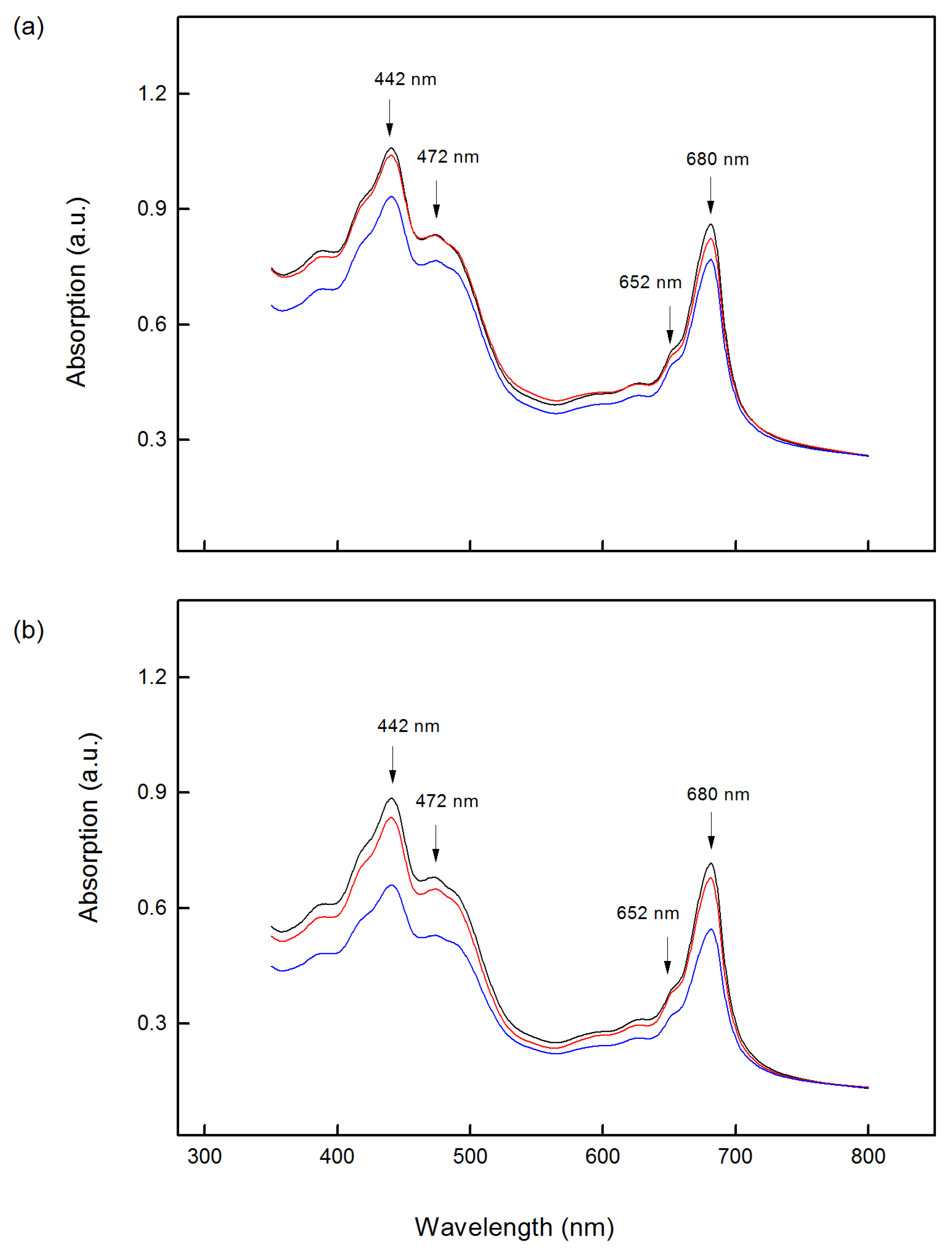
2.2. Absorption Spectra
2.3. Low-Temperature Chlorophyll Fluorescence
2.4. Photochemical Activity
2.5. Oxygen Evolution under Flash and Continuous Illumination
3. Discussion
4. Materials and Methods
4.1. Growth Conditions and Experimental Setup
4.2. Isolation of Thylakoid Membranes
4.3. Absorption Spectra
4.4. Pigment Analysis
4.5. Low-Temperature Fluorescence Measurements
4.6. Photochemical Activity of PSI and PSII
4.7. Measurement of Oxygen Evolution under Flash and Continuous Illumination
4.8. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Daliakopoulos, I.N.; Tsanis, I.K.; Koutroulis, A.; Kourgialas, N.N.; Varouchakis, A.E.; Karatzas, G.P.; Ritsema, C.J. The threat of soil salinity: A European scale review. Sci. Total Environ. 2016, 573, 727–739. [Google Scholar] [CrossRef]
- Hachicha, M.; Kahlaoui, B.; Khamassi, N.; Misle, E.; Jouzdan, O. Effect of electromagnetic treatment of saline water on soil and crops. J. Saudi Soc. Agric. Sci. 2018, 17, 154–162. [Google Scholar] [CrossRef]
- Shokri-Gharelo, R.; Noparvar, P.M. Molecular response of canola to salt stress: Insights on tolerance mechanisms. PeerJ 2018, 2018, e4822. [Google Scholar] [CrossRef] [PubMed]
- Shahid, M.A.; Sarkhosh, A.; Khan, N.; Balal, R.M.; Ali, S.; Rossi, L.; Gómez, C.; Mattson, N.; Nasim, W.; Garcia-Sanchez, F. Insights into the physiological and biochemical impacts of salt stress on plant growth and development. Agronomy 2020, 10, 938. [Google Scholar] [CrossRef]
- Ghosh, B.; Ali Md, N. Response of rice under salinity stress: A review update. Rice Res. Open Access 2016, 4, 167. [Google Scholar] [CrossRef]
- Stefanov, M.A.; Rashkov, G.D.; Yotsova, E.K.; Borisova, P.B.; Dobrikova, A.G.; Apostolova, E.L. Different sensitivity levels of the photosynthetic apparatus in Zea mays L. and Sorghum bicolor L. under salt stress. Plants 2021, 10, 1469. [Google Scholar] [CrossRef] [PubMed]
- Stefanov, M.A.; Rashkov, G.D.; Apostolova, E.L. Assessment of the photosynthetic apparatus functions by chlorophyll fluorescence and P700 absorbance in C3 and C4 plants under physiological conditions and under salt stress. Int. J. Mol. Sci. 2022, 23, 3768. [Google Scholar] [CrossRef] [PubMed]
- Stefanov, M.; Biswal, A.K.; Misra, M.; Misra, A.N.; Apostolova, E.L. Responses of photosynthetic apparatus to salt stress: Structure, function, and protection. In Handbook of Plant and Crop Stress, Fourth Edition; Pessarakli, M., Ed.; Taylor & Francis CRC Press: New York, NY, USA, 2019; pp. 233–250. ISBN 9781351104609. [Google Scholar]
- Ashraf, M.; Harris, P.J.C. Photosynthesis under stressful environments: An overview. Photosynthetica 2013, 51, 163–190. [Google Scholar] [CrossRef]
- Acosta-Motos, J.R.; Diaz-Vivancos, P.; Álvarez, S.; Fernández-García, N.; Sanchez-Blanco, M.J.; Hernández, J.A. Physiological and biochemical mechanisms of the ornamental Eugenia myrtifolia L. plants for coping with NaCl stress and recovery. Planta 2015, 242, 829–846. [Google Scholar] [CrossRef]
- Liang, W.; Ma, X.; Wan, P.; Liu, L. Plant salt-tolerance mechanism: A review. Biochem. Biophys. Res. Commun. 2018, 495, 286–291. [Google Scholar] [CrossRef]
- Shahzad, B.; Rehman, A.; Tanveer, M.; Wang, L.; Park, S.K.; Ali, A. Salt stress in Brassica: Effects, tolerance mechanisms, and management. J. Plant Growth Regul. 2022, 41, 781–795. [Google Scholar] [CrossRef]
- Mbarki, S.; Sytar, O.; Cerda, A.; Zivcak, M.; Rastogi, A.; He, X.; Zoghlami, A.; Abdelly, C.; Brestic, M. Strategies to mitigate the salt stress effects on photosynthetic apparatus and productivity of crop plants. In Salinity Responses and Tolerance in Plants, Volume 1: Targeting Sensory, Transport and Signaling Mechanisms; Springer: Cham, Switzerland, 2018; pp. 85–136. [Google Scholar]
- Arif, Y.; Singh, P.; Siddiqui, H.; Bajguz, A.; Hayat, S. Salinity induced physiological and biochemical changes in plants: An omic approach towards salt stress tolerance. Plant Physiol. Biochem. 2020, 156, 64–77. [Google Scholar] [CrossRef]
- Sachdev, S.; Ansari, S.A.; Ansari, M.I.; Fujita, M.; Hasanuzzaman, M. Abiotic stress and reactive oxygen species: Generation, signaling, and defense mechanisms. Antioxidants 2021, 10, 277. [Google Scholar] [CrossRef] [PubMed]
- Shang, J.X.; Li, X.; Li, C.; Zhao, L. The role of nitric oxide in plant responses to salt stress. Int. J. Mol. Sci. 2022, 23, 6167. [Google Scholar] [CrossRef] [PubMed]
- Dulai, S.; Tarnai, R.; Salamon, D.; Molnár, I.; Vojtkó, A.; Táborská, J.; Allem, A.; Molnár-Láng, M.; Szopkó, D. Effects of water deficit and salt stress on some photosynthesis parameters in wheat and Aegilops comosa lines. Acta Biol. Plant. Agriensis 2019, 7, 55–67. [Google Scholar] [CrossRef]
- Rossi, L.; Borghi, M.; Francini, A.; Lin, X.; Xie, D.-Y.; Sebastiani, L. Salt stress induces differential regulation of the phenylpropanoid pathway in Olea europaea cultivars Frantoio (salt-tolerant) and Leccino (salt-sensitive). J. Plant Physiol. 2016, 204, 8–15. [Google Scholar] [CrossRef]
- Acosta-Motos, J.R.; Ortuño, M.F.; Bernal-Vicente, A.; Diaz-Vivancos, P.; Sanchez-Blanco, M.J.; Hernandez, J.A. Plant responses to salt stress: Adaptive mechanisms. Agronomy 2017, 7, 18. [Google Scholar] [CrossRef]
- Jajoo, A. Changes in photosystem II in response to salt stress. In Ecophysiology and Responses of Plants under Salt Stress; Ahmad, P., Azooz, M.M., Prasad, M.N.V., Eds.; Springer: New York, NY, USA, 2013; pp. 149–168. [Google Scholar] [CrossRef]
- Kalaji, H.M.; Rastogi, A.; Živčák, M.; Brestic, M.; Daszkowska-Golec, A.; Sitko, K.; Alsharafa, K.Y.; Lotfi, R.; Stypiński, P.; Samborska, I.A.; et al. Prompt chlorophyll fluorescence as a tool for crop phenotyping: An example of barley landraces exposed to various abiotic stress factors. Photosynthetica 2018, 56, 953–961. [Google Scholar] [CrossRef]
- Yang, Z.; Li, J.L.; Liu, L.N.; Xie, Q.; Sui, N. Photosynthetic regulation under salt stress and salt-tolerance mechanism of sweet sorghum. Front. Plant Sci. 2020, 10, 1722. [Google Scholar] [CrossRef] [Green Version]
- Parihar, P.; Singh, S.; Singh, R.; Singh, V.P.; Prasad, S.M. Effect of salinity stress on plants and its tolerance strategies: A review. Environ. Sci. Pollut. Res. 2015, 22, 4056–4075. [Google Scholar] [CrossRef]
- Kawakami, K.; Umena, Y.; Kamiya, N.; Shen, J.R. Location of chloride and its possible functions in oxygen-evolving photosystem II revealed by X-ray crystallography. Proc. Natl. Acad. Sci. USA 2009, 106, 8567–8572. [Google Scholar] [CrossRef] [PubMed]
- Mehta, P.; Kreslavsky, V.; Bharti, S.; Allakhverdiev, S.I.; Jajoo, A. Analysis of salt stress induced changes in photosystem II heterogeneity by prompt fluorescence and delayed fluorescence in wheat (Triticum aestivum) leaves. J. Photochem. Photobiol. B Biol. 2011, 104, 308–313. [Google Scholar] [CrossRef]
- Zhang, L.; Xing, D. Rapid determination of the damage to photosynthesis caused by salt and osmotic stresses using delayed fluorescence of chloroplasts. Photochem. Photobiol. Sci. 2008, 7, 352–360. [Google Scholar] [CrossRef]
- Sun, Z.W.; Ren, L.K.; Fan, J.W.; Li, Q.; Wang, K.J.; Guo, M.M.; Wang, L.; Li, J.; Zhang, G.X.; Yang, Z.Y.; et al. Salt response of photosynthetic electron transport system in wheat cultivars with contrasting tolerance. Plant Soil Environ. 2016, 62, 515–521. [Google Scholar] [CrossRef]
- Athar, H.-R.; Zulfiqar, F.; Moosa, A.; Ashraf, M.; Zafar, Z.U.; Zhang, L.; Ahmed, N.; Kalaji, H.M.; Nafees, M.; Hossain, M.A.; et al. Salt stress proteins in plants: An overview. Front. Plant Sci. 2022, 13, 58. [Google Scholar] [CrossRef]
- Huang, L.; Li, Z.; Liu, Q.; Pu, G.; Zhang, Y.; Li, J. Research on the adaptive mechanism of photosynthetic apparatus under salt stress: New directions to increase crop yield in saline soils. Ann. Appl. Biol. 2019, 175, 1–17. [Google Scholar] [CrossRef]
- Pang, Q.; Chen, S.; Dai, S.; Chen, Y.; Wang, Y.; Yan, X. Comparative proteomics of salt tolerance in Arabidopsis thaliana and Thellungiella halophila. J. Proteome Res. 2010, 9, 2584–2599. [Google Scholar] [CrossRef]
- Liu, Z.; Zou, L.; Chen, C.; Zhao, H.; Yan, Y.; Wang, C.; Liu, X. ITRAQ-based quantitative proteomic analysis of salt stress in Spica Prunellae. Sci. Rep. 2019, 9, 9590. [Google Scholar] [CrossRef]
- Dąbrowski, P.; Baczewska, A.H.; Pawluśkiewicz, B.; Paunov, M.; Alexantrov, V.; Goltsev, V.; Kalaji, M.H. Prompt chlorophyll a fluorescence as a rapid tool for diagnostic changes in PSII structure inhibited by salt stress in Perennial ryegrass. J. Photochem. Photobiol. B 2016, 157, 22–31. [Google Scholar] [CrossRef]
- Caruso, G.; Cavaliere, C.; Guarino, C.; Gubbiotti, R.; Foglia, P.; Laganà, A. Identification of changes in Triticum durum L. leaf proteome in response to salt stress by two-dimensional electrophoresis and MALDI-TOF mass spectrometry. Anal. Bioanal. Chem. 2008, 391, 381–390. [Google Scholar] [CrossRef]
- Pandey, D.M.; Choi, I.; Yeo, U.-D. Photosystem 2-activity and thylakoid membrane polypeptides of in vitro cultured Chrysanthemum as affected by NaCl. Biol. Plant. 2009, 53, 329–333. [Google Scholar] [CrossRef]
- Mishra, S.K.; Subrahmanyam, D.; Singhal, G.S. Interrelationship between salt and light stress on primary processes of photosynthesis. J. Plant Physiol. 1991, 138, 92–96. [Google Scholar] [CrossRef]
- Jusovic, M.; Velitchkova, M.Y.; Misheva, S.P.; Börner, A.; Apostolova, E.L.; Dobrikova, A.G. Photosynthetic responses of a wheat mutant (Rht-B1c) with altered DELLA proteins to salt stress. J. Plant Growth Regul. 2018, 37, 645–656. [Google Scholar] [CrossRef]
- Asrar, H.; Hussain, T.; Hadi, S.M.S.; Gul, B.; Nielsen, B.L.; Khan, M.A. Salinity induced changes in light harvesting and carbon assimilating complexes of Desmostachya bipinnata (L.) Staph. Environ. Exp. Bot. 2017, 135, 86–95. [Google Scholar] [CrossRef]
- Zhang, H.; Han, B.; Wang, T.; Chen, S.; Li, H.; Zhang, Y.; Dai, S. Mechanisms of plant salt response: Insights from proteomics. J. Proteome Res. 2012, 11, 49–67. [Google Scholar] [CrossRef] [PubMed]
- Saleem, A.; Ashraf, M.; Akram, N.A. Salt (NaCl)-induced modulation in some key physio-biochemical attributes in Okra (Abelmoschus esculentus L.). J. Agron. Crop Sci. 2011, 197, 202–213. [Google Scholar] [CrossRef]
- Al-Taweel, K.; Iwaki, T.; Yabuta, Y.; Shigeoka, S.; Murata, N.; Wadano, A. A bacterial transgene for catalase protects translation of D1 protein during exposure of salt-stressed tobacco leaves to strong light. Plant Physiol. 2007, 145, 258–265. [Google Scholar] [CrossRef]
- Abdeshahian, M.; Nabipour, M.; Meskarbashee, M. Chlorophyll fluorescence as criterion for the diagnosis salt stress in wheat (Triticum aestivum) plants. Int. J. Chem. Biol. Eng. 2010, 4, 184–186. [Google Scholar]
- Masojidek, J.; Hall, D.O. Salinity and drought stresses are amplified by high irradiance in sorghum. Photosynthetica 1992, 27, 159–171. [Google Scholar]
- Rajesh, A.; Arumugam, R.; Venkatesalu, V. Growth and photosynthetic characteristics of Ceriops roxburghiana under NaCl stress. Photosynthetica 1998, 35, 285–287. [Google Scholar] [CrossRef]
- Kurban, H.; Saneoka, H.; Nehira, K.; Adilla, R.; Fujita, K. Effect of salinity on growth and accumulation of organic and inorganic solutes in the leguminous plants Alhagi pseudoalhagi and Vigna radiata. Soil Sci. Plant Nutr. 1998, 44, 589–597. [Google Scholar] [CrossRef]
- Hakala, M.; Tuominen, I.; Keränen, M.; Tyystjärvi, T.; Tyystjärvi, E. Evidence for the role of the oxygen-evolving manganese complex in photoinhibition of photosystem II. Biochim. Biophys. Acta-Bioenerg. 2005, 1706, 68–80. [Google Scholar] [CrossRef]
- Mehta, P.; Jajoo, A.; Mathur, S.; Bharti, S. Chlorophyll a fluorescence study revealing effects of high salt stress on photosystem II in wheat leaves. Plant Physiol. Biochem. 2010, 48, 16–20. [Google Scholar] [CrossRef] [PubMed]
- Vass, I. Molecular mechanisms of photodamage in the photosystem II complex. Biochim. Biophys. Acta 2012, 1817, 209–217. [Google Scholar] [CrossRef] [PubMed]
- Kosová, K.; Prášil, I.T.; Vítámvás, P. Protein contribution to plant salinity response and tolerance acquisition. Int. J. Mol. Sci. 2013, 14, 6757–6789. [Google Scholar] [CrossRef] [PubMed]
- Duarte, B.; Sleimi, N.; Cagador, I. Biophysical and biochemical constraints imposed by salt stress: Learning from halophytes. Front. Plant Sci. 2014, 5, 746. [Google Scholar] [CrossRef]
- Wungrampha, S.; Joshi, R.; Singla-Pareek, S.L.; Pareek, A. Photosynthesis and salinity: Are these mutually exclusive? Photosynthetica 2018, 56, 366–381. [Google Scholar] [CrossRef]
- Rahdari, P.; Hoseini, S.M. Salinity stress: A Review. Tech. J. Eng. Appl. Sci. 2011, 1, 63–66. [Google Scholar]
- Yang, J.Y.; Zheng, W.; Tian, Y.; Wu, Y.; Zhou, D.W. Effects of various mixed salt-alkaline stresses on growth, photosynthesis, and photosynthetic pigment concentrations of Medicago ruthenica seedlings. Photosynthetica 2011, 49, 275–284. [Google Scholar] [CrossRef]
- Stefanov, M.; Yotsova, E.; Rashkov, G.D.; Ivanova, K.; Markovska, Y.; Apostolova, E.L. Effects of salinity on the photosynthetic apparatus of two Paulownia lines. Plant Physiol. Biochem. 2016, 101, 54–59. [Google Scholar] [CrossRef]
- Khan, M.A.; Shirazi, M.U.; Khan, M.A.; Mujtaba, S.M.; Islam, E.; Mumtaz, S.; Shereen, A.; Ansari, R.U.; Yasin Ashraf, M. Role of proline, K/NA ratio and chlorophyll content in salt tolerance of wheat (Triticum aestivum L.). Pak. J. Bot. 2009, 41, 633–638. [Google Scholar]
- Akram, N.A.; Ashraf, M. Improvement in growth, chlorophyll pigments and photosynthetic performance in salt-stressed plants of sunflower (Helianthus animus L.) by foliar application of 5-aminolevulinic acid. Agrochimica 2011, 55, 94–104. [Google Scholar]
- Barbu, M.C.; Buresova, K.; Tudor, E.M.; Petutschnigg, A. Physical and mechanical properties of Paulownia tomentosa x elongata sawn wood from Spanish, Bulgarian and Serbian plantations. Forest 2022, 13, 1543. [Google Scholar] [CrossRef]
- San, H.P.; Long, L.K.; Zhang, C.Z.; Hui, T.C.; Seng, W.Y.; Lin, F.S.; Hun, A.T.; Fong, W.K. Anatomical features, fiber morphological, physical and mechanical properties of three years old new hybrid Paulownia: Green Paulownia. Res. J. For. 2016, 10, 30–35. [Google Scholar] [CrossRef] [Green Version]
- Kaymakci, A.; Bektas, İ.; Bal, B.C. Some mechanical properties of Paulownia (Paulownia elongata) wood. In Proceedings of the International Caucasion Forestry Symposium, Artvin, Turkey, 24–26 October 2013; pp. 917–919. [Google Scholar]
- Ivanova, K.; Dimitrova, V.; Georgieva, T.; Markovska, Y. Effect of soil salinity on growth, gas exchange and antioxidant defence of two Paulownia lines. Genet. Plant Physiol. 2015, 4, 163–173. [Google Scholar]
- Ivanova, K.; Tzvetkova, N.; Georgieva, T.; Markovska, Y. Photosynthesis and growth responses of five Paulownia lines to salt stress. Compt. Rend. Acad. Bulg. Sci. 2014, 67, 1101–1106. [Google Scholar]
- Cao, Y.; Sun, G.; Zhai, X.; Xu, P.; Ma, L.; Deng, M.; Zhao, Z.; Yang, H.; Dong, Y.; Shang, Z.; et al. Genomic insights into the fast growth of Paulownia and the formation of Paulownia witches’ broom. Mol. Plant 2021, 14, 1668–1682. [Google Scholar] [CrossRef] [PubMed]
- Stefanov, M.; Yotsova, E.; Markovska, Y.; Apostolova, E.L. Effect of high light intensity on the photosynthetic apparatus of two hybrid lines of Paulownia grown on soils with different salinity. Photosynthetica 2018, 56, 832–840. [Google Scholar] [CrossRef]
- Garstka, M.; Jagielski, A. Peroxidative reactions attenuate oxygen effect on spectroscopic properties of isolated chloroplasts. J. Photochem. Photobiol. B 2001, 64, 82–92. [Google Scholar] [CrossRef]
- Strasser, R.J.; Tsimilli-Michael, M.; Srivastava, A. Analysis of the chlorophyll a fluorescence transient. In Chlorophyll a Fluorescence. Advances in Photosynthesis and Respiration; Papageorgiou, G.C., Govindjee, Eds.; Springer: Dordrecht, The Netherlands, 2004; pp. 321–362. [Google Scholar]
- Andreeva, A.; Stoitchkova, K.; Busheva, M.; Apostolova, E. Changes in the energy distribution between chlorophyll–protein complexes of thylakoid membranes from pea mutants with modified pigment content. J. Photochem. Photobiol. B Biol. 2003, 70, 153–162. [Google Scholar] [CrossRef] [PubMed]
- Apostolova, E.L.; Dobrikova, A.G.; Ivanova, P.I.; Petkanchin, I.B.; Taneva, S.G. Relationship between the organization of the PSII supercomplex and the functions of the photosynthetic apparatus. J. Photochem. Photobiol. B Biol. 2006, 83, 114–122. [Google Scholar] [CrossRef]
- Ivanova, P.I.; Dobrikova, A.G.; Taneva, S.G.; Apostolova, E.L. Sensitivity of the photosynthetic apparatus to UV-A radiation: Role of light-harvesting complex II–photosystem II supercomplex organization. Radiat. Environ. Biophys. 2008, 47, 169–177. [Google Scholar] [CrossRef]
- Ashraf, M.; Munns, R. Evolution of approaches to increase the salt tolerance of crops. CRC Crit. Rev. Plant Sci. 2022, 41, 128–160. [Google Scholar] [CrossRef]
- Suo, J.; Zhao, Q.; David, L.; Chen, S.; Dai, S. Salinity response in chloroplasts: Insights from gene characterization. Int. J. Mol. Sci. 2017, 18, 1011. [Google Scholar] [CrossRef]
- Ünnep, R.; Zsiros, O.; Solymosi, K.; Kovács, L.; Lambrev, P.H.; Tóth, T.; Schweins, R.; Posselt, D.; Székely, N.K.; Rosta, L.; et al. The ultrastructure and flexibility of thylakoid membranes in leaves and isolated chloroplasts as revealed by small-angle neutron scattering. Biochim. Biophys. Acta-Bioenerg. 2014, 1837, 1572–1580. [Google Scholar] [CrossRef] [PubMed]
- Zeeshan, M.; Lu, M.; Sehar, S.; Holford, P.; Wu, F. Comparison of biochemical, anatomical, morphological, and physiological responses to salinity stress in wheat and barley genotypes deferring in salinity tolerance. Agronomy 2020, 10, 127. [Google Scholar] [CrossRef] [Green Version]
- Chaves, M.M.; Flexas, J.; Pinheiro, C. Photosynthesis under drought and salt stress: Regulation mechanisms from whole plant to cell. Ann. Bot. 2009, 103, 551–560. [Google Scholar] [CrossRef]
- Misra, A.N.; Sahu, S.M.; Misra, M.; Singh, P.; Meera, I.; Das, N.; Kar, M.; Sahu, P. Sodium chloride induced changes in leaf growth, and pigment and protein contents in two rice cultivars. Biol. Plant. 1997, 39, 257–262. [Google Scholar] [CrossRef]
- Shahbaz, M.; Ashraf, M.; Athar, H.U.R. Does exogenous application of 24-epibrassinolide ameliorate salt induced growth inhibition in wheat (Triticum aestivum L.)? Plant Growth Regul. 2008, 55, 51–64. [Google Scholar] [CrossRef]
- Edge, R.; McGarvey, D.J.; Truscott, T.G. The carotenoids as antioxidants-A review. J. Photochem. Photobiol. B Biol. 1997, 41, 189–200. [Google Scholar] [CrossRef] [PubMed]
- Brugnoli, E.; Lauteri, M. Effects of salinity on stomatal conductance, photosynthetic capacity, and carbon isotope discrimination of salt-tolerant (Gossypium hirsutum L.) and salt-sensitive (Phaseolus vulgaris L.) C3 Non-Halophytes. Plant Physiol. 1991, 95, 628–635. [Google Scholar] [CrossRef]
- Lakra, N.; Kaur, C.; Anwar, K.; Singla-Pareek, S.L.; Pareek, A. Proteomics of contrasting rice genotypes: Identification of potential targets for raising crops for saline environment. Plant Cell Environ. 2018, 41, 947–969. [Google Scholar] [CrossRef]
- Yu, Y.; Assmann, S.M. The effect of NaCl on stomatal opening in Arabidopsis wild type and agb1 heterotrimeric G-protein mutant plants. Plant Signal. Behav. 2016, 11, e1085275. [Google Scholar] [CrossRef] [PubMed]
- Shabala, S.; Cuin, T.A.; Prismall, L.; Nemchinov, L.G. Expression of animal CED-9 anti-apoptotic gene in tobacco modifies plasma membrane ion fluxes in response to salinity and oxidative stress. Planta 2007, 227, 189–197. [Google Scholar] [CrossRef]
- Liu, X.; Fan, Y.; Mak, M.; Babla, M.; Holford, P.; Wang, F.; Chen, G.; Scott, G.; Wang, G.; Shabala, S.; et al. QTLs for stomatal and photosynthetic traits related to salinity tolerance in barley. BMC Genom. 2017, 18, 9. [Google Scholar] [CrossRef]
- Gomathi, R.; Rakkiyapan, P. Comparative lipid peroxidation, leaf membrane thermostability, and antioxidant system in four Sugarcane genotypes differing in salt tolerance. Int. J. Plant Physiol. Biochem. 2011, 3, 67–74. [Google Scholar]
- Ziaf, K.; Amjad, M.; Pervez, M.A.; Iqbal, Q.; Rajwana, I.A.; Ayyub, M. Evaluation of different growth and physiological traits as indices of salt tolerance in hot pepper (Capsicum annuum L.). Pak. J. Bot. 2009, 41, 1797–1809. [Google Scholar]
- Khavari-Nejad, R.A.; Mostofi, Y. Effects of NaCl on photosynthetic pigments, saccharides, and chloroplast ultrastructure in leaves of tomato cultivars. Photosynthetica 1998, 35, 151–154. [Google Scholar] [CrossRef]
- Shu, S.; Guo, S.R.; Sun, J.; Yuan, L.Y. Effects of salt stress on the structure and function of the photosynthetic apparatus in Cucumis sativus and its protection by exogenous putrescine. Physiol. Plant. 2012, 146, 285–296. [Google Scholar] [CrossRef]
- Gao, H.J.; Yang, H.Y.; Bai, J.P.; Liang, X.Y.; Lou, Y.; Zhang, J.L.; Wang, D.; Zhang, J.L.; Niu, S.Q.; Chen, Y.L. Ultrastructural and physiological responses of potato (Solanum tuberosum L.) plantlets to gradient saline stress. Front. Plant Sci. 2015, 5, 787. [Google Scholar] [CrossRef]
- Yamamoto, Y.; Hori, H.; Kai, S.; Ishikawa, T.; Ohnishi, A.; Tsumura, N.; Morita, N. Quality control of photosystem II: Reversible and irreversible protein aggregation decides the fate of photosystem II under excessive illumination. Front. Plant Sci. 2013, 4, 433. [Google Scholar] [CrossRef] [PubMed]
- Tang, Y.; Wen, X.; Lu, Q.; Yang, Z.; Cheng, Z.; Lu, C. Heat stress induces an aggregation of the light-harvesting complex of photosystem II in spinach plants. Plant Physiol. 2007, 143, 629–638. [Google Scholar] [CrossRef]
- Stefanov, M.; Yotsova, E.; Gesheva, E.; Dimitrova, V.; Markovska, Y.; Doncheva, S.; Apostolova, E.L. Role of flavonoids and proline in the protection of photosynthetic apparatus in Paulownia under salt stress. S. Afr. J. Bot. 2021, 139, 246–253. [Google Scholar] [CrossRef]
- Allen, J.F.; Mullineaux, C.W. Probing the mechanism of state transitions in oxygenic photosynthesis by chlorophyll fluorescence spectroscopy, kinetics and imaging. In Chlorophyll a Fluorescence; Papageorgiou, G.C., Govindjee, Eds.; Kluwer Academic Publishers: Amsterdam, The Netherlands, 2007. [Google Scholar]
- Zahra, J.; Nazim, H.; Cai, S.; Han, Y.; Wu, D.; Zhang, B.; Haider, S.I.; Zhang, G. The influence of salinity on cell ultrastructures and photosynthetic apparatus of barley genotypes differing in salt stress tolerance. Acta Physiol. Plant. 2014, 36, 1261–1269. [Google Scholar] [CrossRef]
- Sudhir, P.R.; Pogoryelov, D.; Kovács, L.; Garab, G.; Murthy, S.D.S. The effects of salt stress on photosynthetic electron transport and thylakoid membrane proteins in the cyanobacterium Spirulina platensis. J. Biochem. Mol. Biol. 2005, 38, 481–485. [Google Scholar] [CrossRef]
- Alia; Pardha Saradhi, P.; Mohanty, P. Involvement of proline in protecting thylakoid membranes against free radical-induced photodamage. J. Photochem. Photobiol. B Biol. 1997, 38, 253–257. [Google Scholar] [CrossRef]
- Hoganson, C.W.; Babcock, G.T. A metalloradical mechanism for the generation of oxygen from water in photosynthesis. Science 1997, 277, 1953–1956. [Google Scholar] [CrossRef]
- Yotsova, E.K.; Stefanov, M.A.; Dobrikova, A.G.; Apostolova, E.L. Different sensitivities of photosystem II in green algae and cyanobacteria to phenylurea and phenol-type herbicides: Effect on electron donor side. Z. Naturforsch. C 2017, 72, 315–324. [Google Scholar] [CrossRef]
- Maslenkova, L.; Gambarova, N.; Zeinalov, Y. NaCl-induced changes in oxygen evolving activity and thylakoid membrane patterns of barley plants. Adaptation to salinity. Bulg. J. Plant Physiol. 1995, 21, 29–35. [Google Scholar]
- Dankov, K.G.; Dobrikova, A.G.; Ughy, B.; Bogos, B.; Gombos, Z.; Apostolova, E.L. LHCII organization and thylakoid lipids affect the sensitivity of the photosynthetic apparatus to high-light treatment. Plant Physiol. Biochem. 2011, 49, 629–635. [Google Scholar] [CrossRef]
- Dankov, K.; Busheva, M.; Stefanov, D.; Apostolova, E.L. Relationship between the degree of carotenoid depletion and function of the photosynthetic apparatus. J. Photochem. Photobiol. B Biol. 2009, 96, 49–56. [Google Scholar] [CrossRef] [PubMed]
- Harrison, M.A.; Melis, A. Organization and stability of polypeptides associated with the chlorophyll a-b light-harvesting complex of photosystem II. Plant Cell Physiol. 1992, 33, 627–637. [Google Scholar] [CrossRef]
- Lichtenthaler, H.K. Chlorophylls and carotenoids: Pigments of photosynthetic biomembranes. Methods Enzymol. 1987, 148, 350–382. [Google Scholar] [CrossRef]
- Zeinalov, Y. An equipment for investigations of photosynthetic oxygen production reactions. Bulg. J. Agric. Sci. 2002, 28, 57–67. [Google Scholar]
- Kok, B.; Forbush, B.; McGloin, M. Cooperation of charges in photosynthetic O2 evolution-I. A linear four step mechanism. Photochem. Photobiol. 1970, 11, 457–475. [Google Scholar] [CrossRef]
- Zeinalov, Y. On the minimum quantum requirement of photosynthesis. Z. Naturforsch.-Sect. C J. Biosci. 2009, 64, 673–679. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| NaCl | λexc = 436 nm | λexc = 472 nm | |||
|---|---|---|---|---|---|
| (mM) | F735/F685 | F695/F685 | F735/F685 | F695/F685 | |
| 10 days | |||||
| TF | 0 | 1.553 ± 0.031 d | 0.854 ± 0.017 c,d | 1.257 ± 0.026 f | 0.934 ± 0.014 b |
| 100 | 1.570 ± 0.035 d | 0.882 ± 0.013 c,d | 1.251 ± 0.030 f | 0.921 ± 0.007 b | |
| 150 | 1.784 ± 0.029 b,c | 0.933 ± 0.004 b | 1.332 ± 0.018 d,e | 0.951 ± 0.016 a | |
| EE | 0 | 1.715 ± 0.029 c | 0.924 ± 0.010 b | 1.442 ± 0.031 c | 0.909 ± 0.025 b |
| 100 | 1.715 ± 0.025 c | 0.923 ± 0.009 b | 1.410 ± 0.017 c | 0.898 ± 0.022 b | |
| 150 | 1.798 ± 0.019 b | 0.995 ± 0.016 a | 1.682 ± 0.044 a | 0.947 ± 0.023 a | |
| 25 days | |||||
| TF | 0 | 1.340 ± 0.019 e | 0.842 ± 0.016 d | 1.225± 0.025 f | 0.847 ± 0.014 c |
| 100 | 1.347 ± 0.011 e | 0.867 ± 0.017 c,d | 1.276 ± 0.021 e,f | 0.850 ± 0.017 c | |
| 150 | 1.427 ± 0.039 e | 0.869 ± 0.008 c,d | 1.292 ± 0.023 e,f | 0.880 ± 0.013 b,c | |
| EE | 0 | 1.909 ± 0.039 a | 0.884 ± 0.014 c,d | 1.484 ± 0.026 b,c | 0.944 ± 0.022 a |
| 100 | 1.954 ± 0.041 a | 0.886 ± 0.013 c | 1.538 ± 0.030 b | 0.957 ± 0.030 a | |
| 150 | * | * | * | * | |
| NaCl | Area (%) | |||||
|---|---|---|---|---|---|---|
| (mM) | LHCIIM+T | PSII RC | PSII Antenna | LHCIIA | PSI Core | PSI Antenna |
| TF | ||||||
| 0 | 9.161 ± 0.067 a | 21.642 ± 0.131 a | 20.676 ± 0.109 a | 8.995 ± 0.087 e | 12.128 ± 0.065 e | 27.399 ± 0.044 d |
| 100 | 9.184 ± 0.057 a | 18.925 ± 0.115 c | 17.945 ± 0.095 c | 10.697 ± 0.104 b,c | 14.267 ± 0.0786 c | 28.981 ± 0.046 c |
| 150 | 7.595 ± 0.056 b | 17.704 ± 0.129 d | 17.251 ± 0.105 d | 10.443 ± 0.055 b,c | 17.4362 ± 0.170 a | 29.571 ± 0.160 b |
| EE | ||||||
| 0 | 7.047 ± 0.085 c | 20.4834 ± 0.151 b | 19.083 ± 0.036 b | 10.735 ± 0.096 b | 13.612 ± 0.122 d | 29.039 ± 0.285 b,c |
| 100 | 7.078 ± 0.045 c | 18.978 ± 0.048 c | 19.075 ± 0.067 b | 12.389 ± 0.010 a | 13.666 ± 0.035 d | 28.812 ± 0.150 c |
| 150 | 6.054 ± 0.070 d | 16.319 ± 0.039 e | 16.319 ± 0.119 d | 14.455 ± 0.098 d | 14.984 ± 0.096 b | 34.518 ± 0.078 a |
| NaCl (mM) | S0 (%) | α (%) | β (%) | SB | AF/AS | |
|---|---|---|---|---|---|---|
| 10 days | ||||||
| TF | 0 | 24.70 ± 1.04 c | 22.96 ± 1.17 c | 5.78 ± 0.36 c | 0.98.± 0.11 d | 2.88 ± 0.17 a |
| 100 | 23.41 ± 0.85 c | 24.81 ± 1.01 b,c | 5 27 ± 0.30 c | 1.02 ± 0.05 d | 2.59 ± 0.14 a,b | |
| 150 | 30.73 ± 1.78 a | 27.42 ± 1.05 b | 7.64 ± 0.44 ab | 1.39 ± 0.08 b | 1.87 ± 0.18 c | |
| EE | 0 | 24.37 ± 0.55 c | 23.29 ± 1.14 b,c | 5.29 ± 0.33 c | 1.11 ± 0.10 cd | 2.43 ± 0.16 b |
| 100 | 26.58 ± 1.12 b,c | 24.40 ± 1.18 b | 5.54 ± 0.33 c | 1.34 ± 0.09 b | 2.32 ± 0.14 b | |
| 150 | 31.93 ± 1.49 a | 29.44 ± 1.02 a | 8.47 ± 0.52 a | 1.73 ± 0.11 a | 1.93 ± 0.15 c | |
| 25 days | ||||||
| TF | 0 | 23.80 ± 0.68 c | 24.32 ± 0.86 c | 4.53 ± 0.33 c | 1.15 ± 0.09 c | 2.67± 0.16 ab |
| 100 | 22.35 ± 1.12 c | 25.19 ± 0.82 b,c | 4.19 ± 0.21 c | 1.25 ± 0.11 c,b | 2.56 ± 0.07 b | |
| 150 | 26.41 ± 1.03 b | 27.58 ± 0.99 b | 6.08 ± 0.80 b | 1.33 ± 0.10 c,b | 2.42 ± 0.13 b | |
| EE | 0 | 23.72 ± 0.63 c | 26.40 ± 1.08 b | 5.92 ± 0.23 c | 1.19 ± 0.11 c | 2.39± 0.15 b |
| 100 | 25.51 ± 0.69 c | 27.53 ± 1.02 b | 5.97 ± 0.22 c | 1.38 ± 0.13 c,b | 2.34 ± 0.17 b | |
| 150 | * | * | * | * | * | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Stefanov, M.A.; Rashkov, G.D.; Yotsova, E.K.; Dobrikova, A.G.; Apostolova, E.L. Impact of Salinity on the Energy Transfer between Pigment–Protein Complexes in Photosynthetic Apparatus, Functions of the Oxygen-Evolving Complex and Photochemical Activities of Photosystem II and Photosystem I in Two Paulownia Lines. Int. J. Mol. Sci. 2023, 24, 3108. https://doi.org/10.3390/ijms24043108
Stefanov MA, Rashkov GD, Yotsova EK, Dobrikova AG, Apostolova EL. Impact of Salinity on the Energy Transfer between Pigment–Protein Complexes in Photosynthetic Apparatus, Functions of the Oxygen-Evolving Complex and Photochemical Activities of Photosystem II and Photosystem I in Two Paulownia Lines. International Journal of Molecular Sciences. 2023; 24(4):3108. https://doi.org/10.3390/ijms24043108
Chicago/Turabian StyleStefanov, Martin A., Georgi D. Rashkov, Ekaterina K. Yotsova, Anelia G. Dobrikova, and Emilia L. Apostolova. 2023. "Impact of Salinity on the Energy Transfer between Pigment–Protein Complexes in Photosynthetic Apparatus, Functions of the Oxygen-Evolving Complex and Photochemical Activities of Photosystem II and Photosystem I in Two Paulownia Lines" International Journal of Molecular Sciences 24, no. 4: 3108. https://doi.org/10.3390/ijms24043108