
The Antifungal Effect of Pyroligneous Acid on the Phytopathogenic Fungus Botrytis cinerea

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
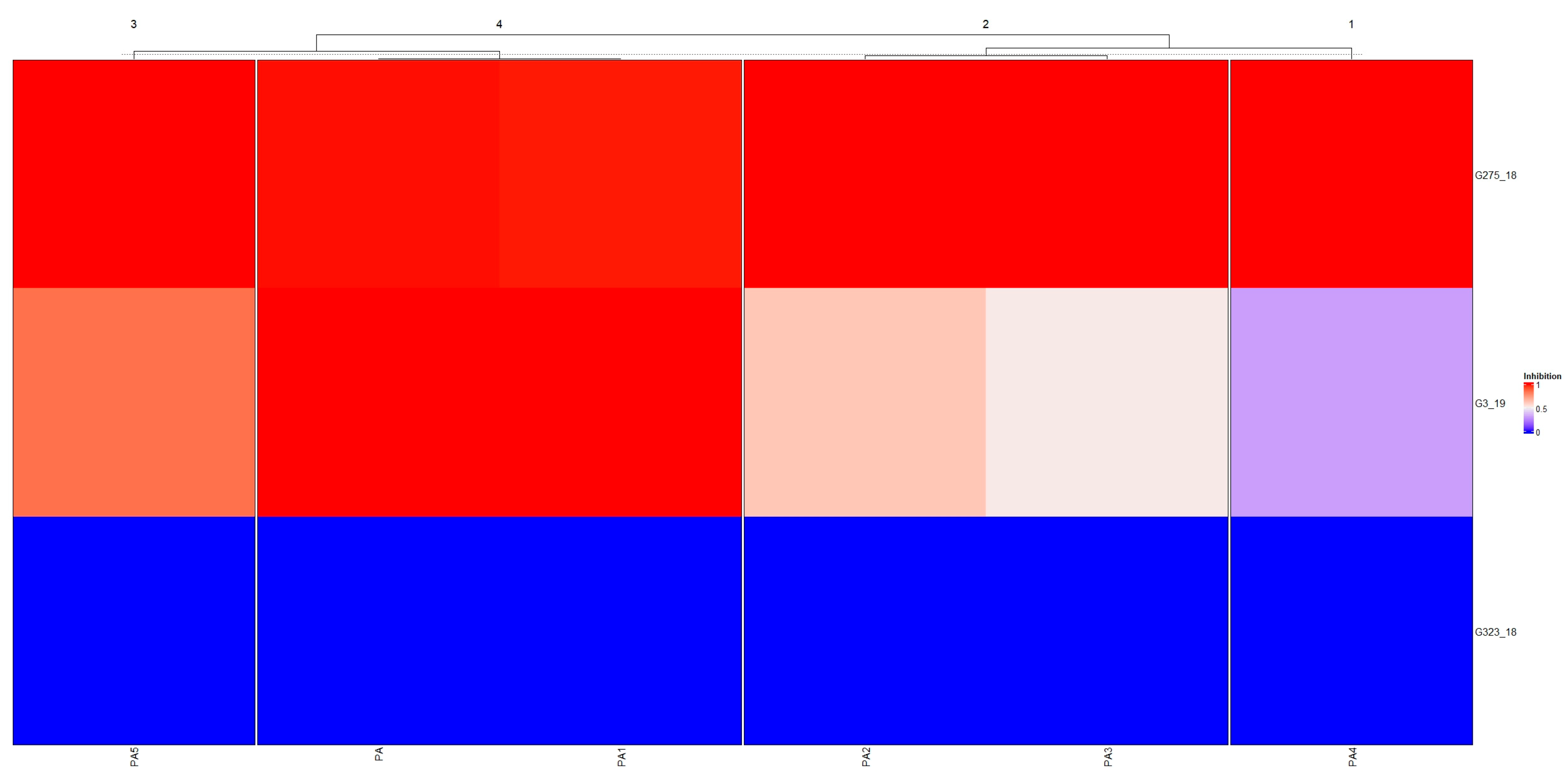
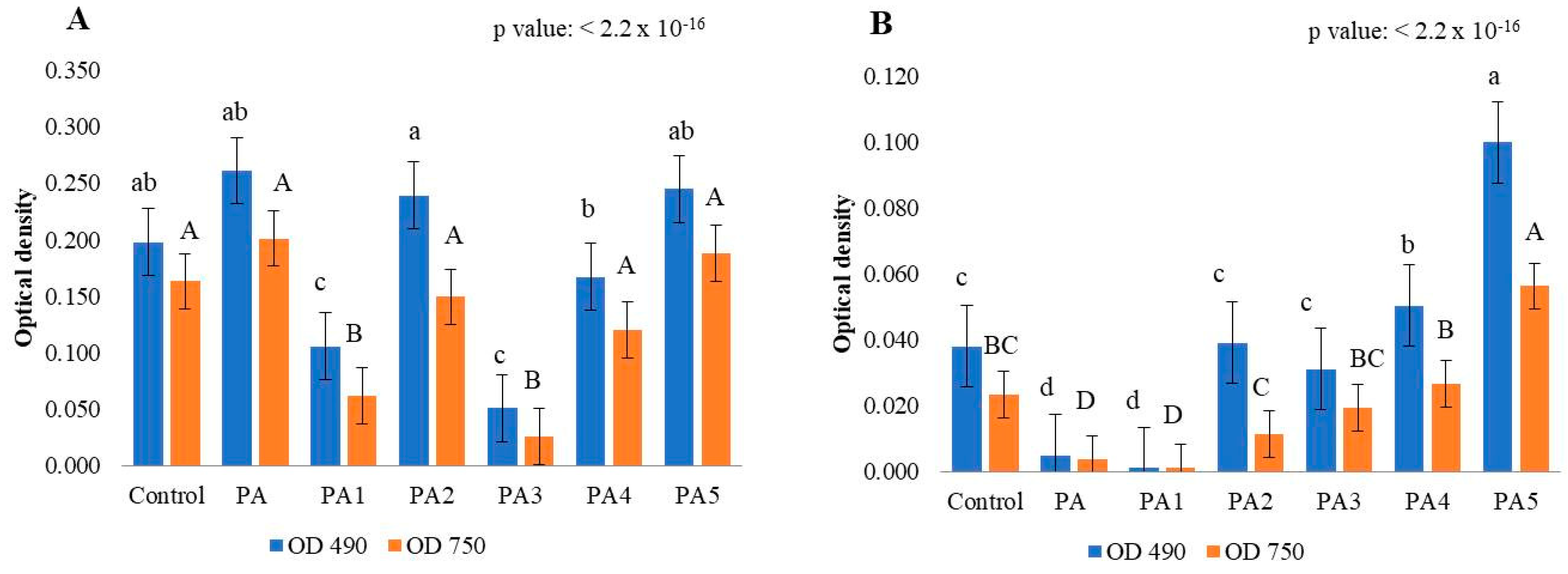
2.1. Effect of Pyroligneous Acid on Fungal Mycelium Growth on Petri Dishes
2.2. Effect of Pyroligneous Acid on Growth of Botrytis cinerea in MT2 Plates
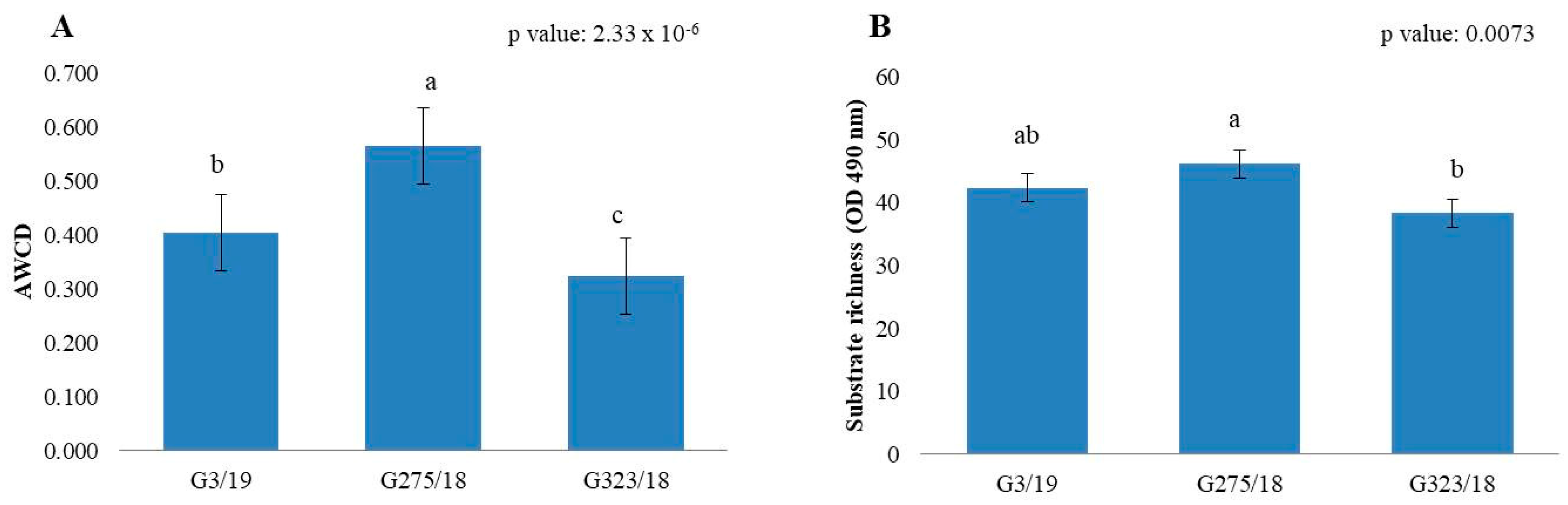
2.3. Metabolic Profile of Botrytis cinerea Pre-Cultivated on Pyroligneous Acid
3. Discussion
4. Materials and Methods
4.1. Fungal Strains
4.2. Pyroligneous Acid
4.3. Inhibition Test
4.4. MT2 Microplates Analysis
4.5. FF Microplates Analysis
4.6. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Khanal, R.C. Climate change and organic agriculture. J. Agric. Environ. 2009, 10, 116–127. [Google Scholar] [CrossRef]
- Peeters, A.; Lefebvre, O.; Balogh, L.; Barberi, P.; Batello, C.; Bellon, S.; Gaifami, T.; Gkisakis, V.; Lana, M.; Migliorini, P.; et al. A Green Deal for implementing agroecological systems: Reforming the Common Agricultural Policy of the European Union. J. Sustain. Org. Agric. Syst. 2020, 70, 83–93. [Google Scholar] [CrossRef]
- Prandecki, K.; Wrzaszcz, W.; Zieliński, M. Environmental and climate challenges to agriculture in Poland in the context of objectives adopted in the European Green Deal strategy. Sustainability 2021, 13, 10318. [Google Scholar] [CrossRef]
- FAO. Faostat. Available online: https://www.fao.org/faostat/en/#data (accessed on 4 April 2022).
- Malarczyk, D.G.; Panek, J.; Frąc, M. Triplex Real-Time PCR approach for the detection of crucial fungal berry pathogens—Botrytis spp., Colletotrichum spp. and Verticillium spp. Int. J. Mol. Sci. 2020, 21, 8469. [Google Scholar] [CrossRef]
- Velásquez, A.C.; Castroverde, C.D.M.; He, S.Y. Plant–pathogen warfare under changing climate conditions. Curr. Biol. 2018, 28, R619–R634. [Google Scholar] [CrossRef] [PubMed]
- Singh, J.; Bhatnagar, S.K.; Tomar, A. study on fungicidal effect of plant extracts on plant pathogenic fungi and the economy of extract preparation and efficacy in comparison to synthetic/chemical fungicides. J. Appl. Nat. Sci. 2019, 11, 333–337. [Google Scholar] [CrossRef]
- Jiménez-Reyes, M.F.; Carrasco, H.; Olea, A.F.; Silva-Moreno, E. Natural compounds: A sustainable alternative to the phytopathogens control. J. Chil. Chem. Soc. 2019, 64, 4459–4465. [Google Scholar] [CrossRef]
- Holz, G.; Coertze, S.; Williamson, B. The ecology of Botrytis on plant surfaces. In Botrytis: Biology, Pathology and Control; Elad, Y., Williamson, B., Tudzynski, P., Delen, N., Eds.; Springer: Berlin/Heidelberg, Germany, 2007; pp. 9–27. ISBN 9781119130536. [Google Scholar]
- Cheung, N.; Tian, L.; Liu, X.; Li, X. The destructive fungal pathogen Botrytis cinerea—Insights from genes studied with mutant analysis. Pathogens 2020, 9, 923. [Google Scholar] [CrossRef]
- Weber, R.W.S.; Hahn, M. Grey mould disease of strawberry in Northern Germany: Causal agents, fungicide resistance and management Strategies. Appl. Microbiol. Biotechnol. 2019, 103, 1589–1597. [Google Scholar] [CrossRef]
- Rupp, S.; Plesken, C.; Rumsey, S.; Dowling, M.; Schnabel, G.; Weber, R.W.S.; Hahn, M. Botrytis fragariae, a new species causing gray mold on strawberries, shows high fungicide resistance. Appl. Environ. Microbiol. 2017, 83, e00269-17. [Google Scholar] [CrossRef] [Green Version]
- Oliveira, M.S.; Amiri, A.; Zuniga, A.I.; Peres, N.A. Sources of primary inoculum of Botrytis cinerea and their impact on fungicide resistance development in commercial strawberry fields. Plant Dis. 2017, 101, 1761–1768. [Google Scholar] [CrossRef] [PubMed]
- Hahn, M. The rising threat of fungicide resistance in plant pathogenic fungi: Botrytis as a case study. J. Chem. Biol. 2014, 7, 133–141. [Google Scholar] [CrossRef] [PubMed]
- Rupp, S.; Weber, R.W.S.; Rieger, D.; Detzel, P.; Hahn, M. Spread of Botrytis cinerea strains with multiple fungicide resistance in German horticulture. Front. Microbiol. 2017, 7, 1200. [Google Scholar] [CrossRef] [PubMed]
- Malarczyk, D.; Panek, J.; Frąc, M. Alternative molecular-based diagnostic methods of plant pathogenic fungi affecting berry crops—A review. Molecules 2019, 24, 1200. [Google Scholar] [CrossRef] [PubMed]
- Sanzani, S.M.; Schena, L.; De Cicco, V.; Ippolito, A. Early detection of Botrytis cinerea latent infections as a tool to improve postharvest quality of table grapes. Postharvest Biol. Technol. 2012, 68, 64–71. [Google Scholar] [CrossRef]
- Van Kan, J.A.L. Infection strategies of Botrytis cinerea. Acta Hortic. 2005, 669, 77–90. [Google Scholar] [CrossRef]
- Grewal, A.; Abbey, L.; Gunupuru, L.R. Production, prospects and potential application of pyroligneous acid in agriculture. J. Anal. Appl. Pyrolysis 2018, 135, 152–159. [Google Scholar] [CrossRef]
- Becagli, M.; Arduini, I.; Cantini, V.; Cardelli, R. Soil and foliar applications of wood distillate differently affect soil properties and field bean traits in preliminary field tests. Plants 2023, 12, 121. [Google Scholar] [CrossRef]
- Mungkunkamchao, T.; Kesmala, T.; Pimratch, S.; Toomsan, B.; Jothityangkoon, D. Wood vinegar and fermented bioextracts: Natural products to enhance growth and yield of tomato (Solanum lycopersicum L.). Sci. Hortic. 2013, 154, 66–72. [Google Scholar] [CrossRef]
- Wei, Q.; Ma, X.; Dong, J. Preparation, chemical constituents and antimicrobial activity of pyroligneous acids from walnut tree branches. J. Anal. Appl. Pyrolysis 2010, 87, 24–28. [Google Scholar] [CrossRef]
- Simma, B.; Polthanee, A.; Goggi, A.S. Wood vinegar seed priming improves yield and suppresses weeds in dryland direct-seeding rice under rainfed production. Agron. Sustain. Dev. 2017, 37, 56. [Google Scholar] [CrossRef]
- Mao, K.; Li, S.; Li, B.; Wu, W.; Wei, C.; Yuan, S.; Niu, Y.; Du, H.; Zhang, L. Effect of wood vinegar on growth, yield and quality of upper leaves of flue-cured tobacco of Nanzheng. Southwest China J. Agric. Sci. 2019, 32, 645–652. [Google Scholar]
- Wang, Y.; Qiu, L.; Song, Q.; Wang, S.; Wang, Y.; Ge, Y. Root proteomics reveals the effects of wood vinegar on wheat growth and subsequent tolerance to drought stress. Int. J. Mol. Sci. 2019, 20, 669. [Google Scholar] [CrossRef] [PubMed]
- Fanfarillo, E.; Fedeli, R.; Fiaschi, T.; De Simone, L.; Vannini, A.; Angiolini, C.; Loppi, S.; Maccherini, S. Effects of wood distillate on seedling emergence and first-stage growth in five threatened arable plants. Diversity 2022, 14, 669. [Google Scholar] [CrossRef]
- Luo, X.; Wang, Z.; Meki, K.; Wang, X.; Liu, B.; Zheng, H.; You, X.; Li, F. Effect of co-application of wood vinegar and biochar on seed germination and seedling growth. J. Soils Sediments 2019, 19, 3934–3944. [Google Scholar] [CrossRef]
- Zhu, K.; Gu, S.; Liu, J.; Luo, T.; Khan, Z.; Zhang, K.; Hu, L. Wood vinegar as a complex growth regulator promotes the growth, yield, and quality of rapeseed. Agronomy 2021, 11, 510. [Google Scholar] [CrossRef]
- Mourant, D.; Yang, D.-Q.; Lu, X.; Roy, C. Anti-fungal properties of the pyroligneous liquors from the pyrolysis of softwood bark. Wood Fiber Sci. 2007, 37, 542–548. [Google Scholar]
- Vannini, A.; Fedeli, R.; Guarnieri, M.; Loppi, S. Foliar application of wood distillate alleviates ozone-induced damage in lettuce (Lactuca sativa L.). Toxics 2022, 10, 178. [Google Scholar] [CrossRef]
- Sun, H.; Feng, Y.; Xue, L.; Mandal, S.; Wang, H.; Shi, W.; Yang, L. Responses of ammonia volatilization from rice paddy soil to application of wood vinegar alone or combined with biochar. Chemosphere 2020, 242, 125247. [Google Scholar] [CrossRef]
- Chalermsan, Y.; Peerapan, S. Wood vinegar: By-product from rural charcoal kiln and its role in plant protection. Asian J. Food Agro-Industry 2009, 2, 189–195. [Google Scholar]
- Quan, S. Application of wood vinegar to control diseases. J. Agric. Sci. Yanbian Univ. 1994, 2, 113–116. [Google Scholar]
- Mao, Q.; Ding, F.; Zhao, Z.; Ma, X. Preparation of apricot tree vinegar and its antibacterial activity and chemical composition analysis. J. Northwest A F Univ. 2009, 37, 91–96. [Google Scholar]
- Zeng, J.; Hai, M.; Wang, X. The effect of dabuxilatu wood vinegar on the quality and disease of organic tobacco. Agric. Sci. Bull 2014, 7, 91–96. [Google Scholar]
- Suresh, G.; Pakdel, H.; Rouissi, T.; Brar, S.K.; Fliss, I.; Roy, C. In vitro evaluation of antimicrobial efficacy of pyroligneous acid from softwood mixture. Biotechnol. Res. Innov. 2019, 3, 47–53. [Google Scholar] [CrossRef]
- Jung, K.-H. Growth inhibition effect of pyroligneous acid on pathogenic fungus, Alternaria mali, the agent of Alternaria blotch of apple. Biotechnol. Bioprocess Eng. 2007, 12, 318–322. [Google Scholar] [CrossRef]
- Pinzari, F.; Ceci, A.; Abu-Samra, N.; Canfora, L.; Maggi, O.; Persiani, A. Phenotype microArray TM system in the study of fungal functional diversity and catabolic versatility. Res. Microbiol. 2016, 167, 710–722. [Google Scholar] [CrossRef]
- Chen, Y.H.; Li, Y.F.; Wei, H.; Li, X.X.; Zheng, H.T.; Dong, X.Y.; Xu, T.F.; Meng, J.F. Inhibition efficiency of wood vinegar on grey mould of table grapes. Food Biosci. 2020, 38, 100755. [Google Scholar] [CrossRef]
- Mathew, S.; Zakaria, Z.A. Pyroligneous acid—The smoky acidic liquid from plant biomass. Appl. Microbiol. Biotechnol. 2015, 99, 611–622. [Google Scholar] [CrossRef]
- Oszust, K.; Panek, J.; Pertile, G.; Siczek, A.; Oleszek, M.; Frąc, M. Metabolic and genetic properties of Petriella setifera precultured on waste. Front. Microbiol. 2018, 9, 115. [Google Scholar] [CrossRef]
- Pertile, G.; Panek, J.; Oszust, K.; Siczek, A.; Oleszek, M.; Gryta, A.; Frąc, M. Effect of different organic waste on cellulose-degrading enzymes secreted by Petriella setifera in the presence of cellobiose and glucose. Cellulose 2019, 4, 7905–7922. [Google Scholar] [CrossRef]
- Frąc, M.; Gryta, A.; Oszust, K.; Kotowicz, N. Fast and accurate microplate method (Biolog MT2) for detection of Fusarium fungicides resistance/sensitivity. Front. Microbiol. 2016, 7, 489. [Google Scholar] [CrossRef] [PubMed]
- Pylak, M.; Oszust, K.; Frąc, M. Searching for new beneficial bacterial isolates of wild raspberries for biocontrol of phytopathogens-antagonistic properties and functional characterization. Int. J. Mol. Sci. 2020, 21, 9361. [Google Scholar] [CrossRef] [PubMed]
- Frąc, M. Mycological Evaluation of Dairy Sewage Sludge and Its Influence on Functional Diversity of Soil Microorganisms; Institute of Agrophysics Polish Academy of Sciences: Lublin, Poland, 2012; ISBN 978-83-89969-96-5. [Google Scholar]
- Chou, H.-Y.; Chiang, M.W.-L.; Lin, W.-R.; Hsieh, S.-Y.; Jones, E.B.G.; Guo, S.-Y.; Pang, K.-L. Metabolic activity on Biolog FF microPlate suggests organic substrate decomposition by Aspergillus terreus NTOU4989 isolated from Kueishan Island Hydrothermal Vent Field, Taiwan. Fungal Ecol. 2022, 60, 101157. [Google Scholar] [CrossRef]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pertile, G.; Frąc, M. The Antifungal Effect of Pyroligneous Acid on the Phytopathogenic Fungus Botrytis cinerea. Int. J. Mol. Sci. 2023, 24, 3080. https://doi.org/10.3390/ijms24043080
Pertile G, Frąc M. The Antifungal Effect of Pyroligneous Acid on the Phytopathogenic Fungus Botrytis cinerea. International Journal of Molecular Sciences. 2023; 24(4):3080. https://doi.org/10.3390/ijms24043080
Chicago/Turabian StylePertile, Giorgia, and Magdalena Frąc. 2023. "The Antifungal Effect of Pyroligneous Acid on the Phytopathogenic Fungus Botrytis cinerea" International Journal of Molecular Sciences 24, no. 4: 3080. https://doi.org/10.3390/ijms24043080