
Poly-L-Lysine to Fight Antibiotic Resistances of Pseudomonas aeruginosa

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
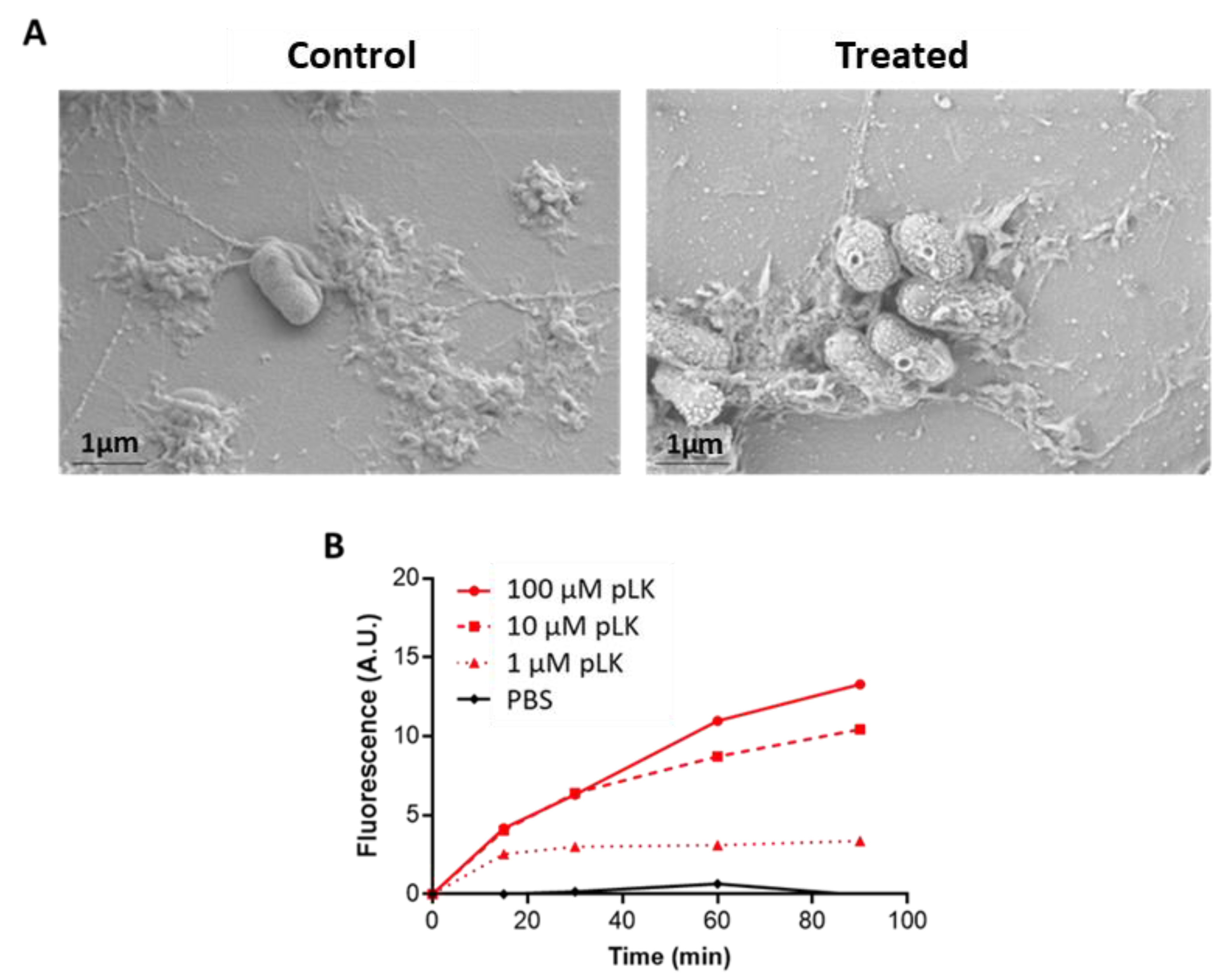
2.1. Permeabilization of P. aeruginosa Membrane by pLK
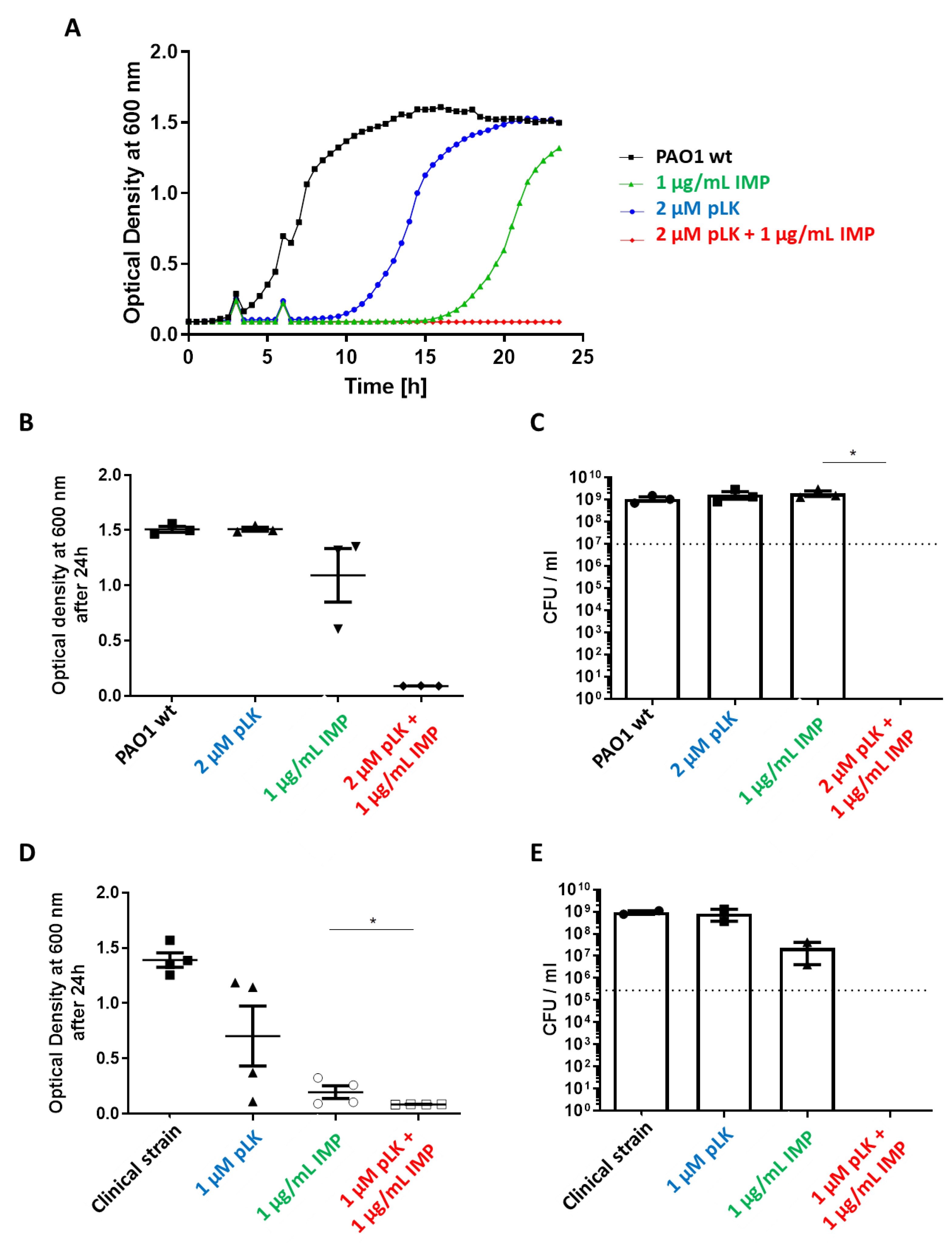
2.2. In Vitro Synergistic Effect of pLK and Imipenem Association against Reference (PAO1) and Imipenem-Sensitive P. aeruginosa Strains
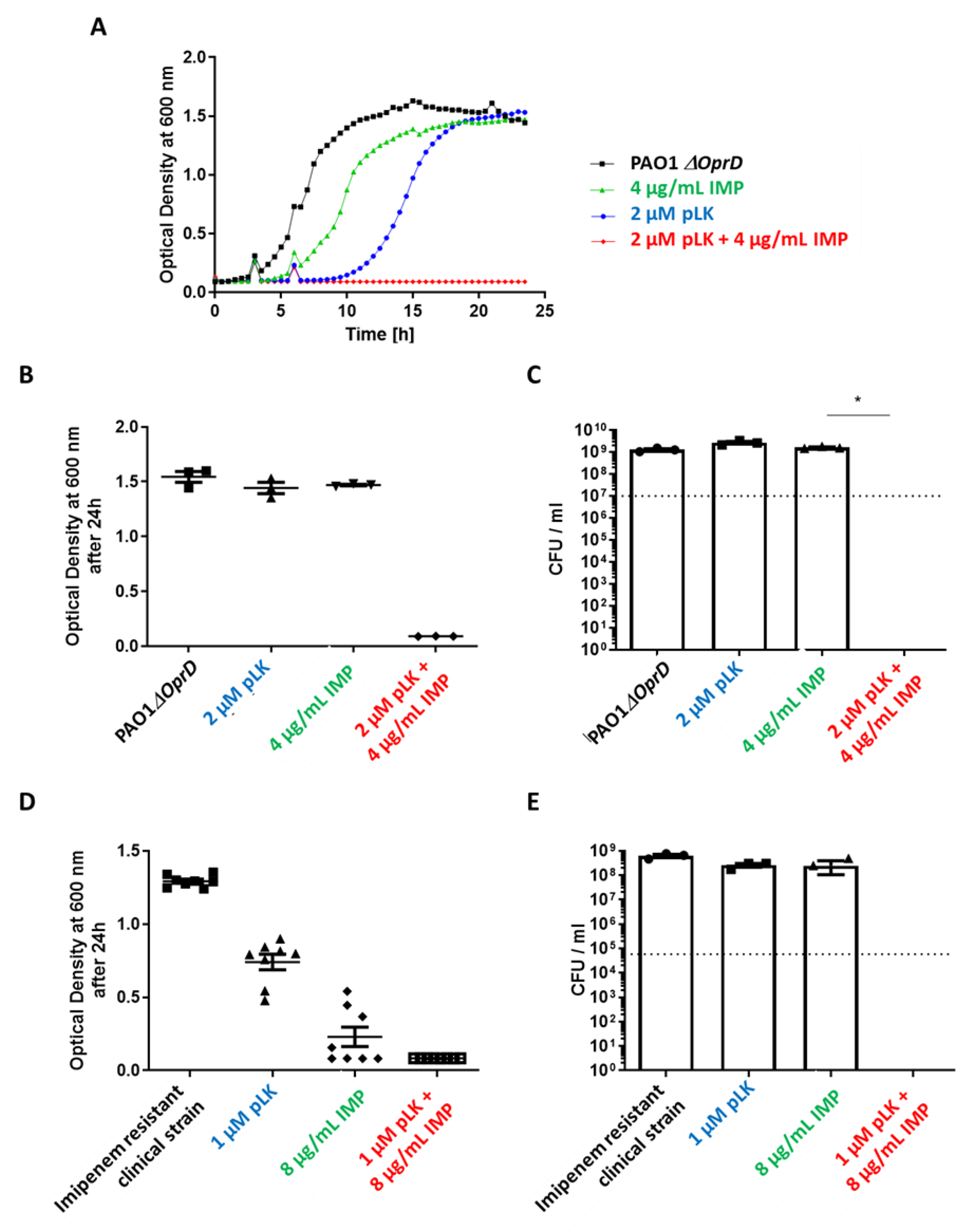
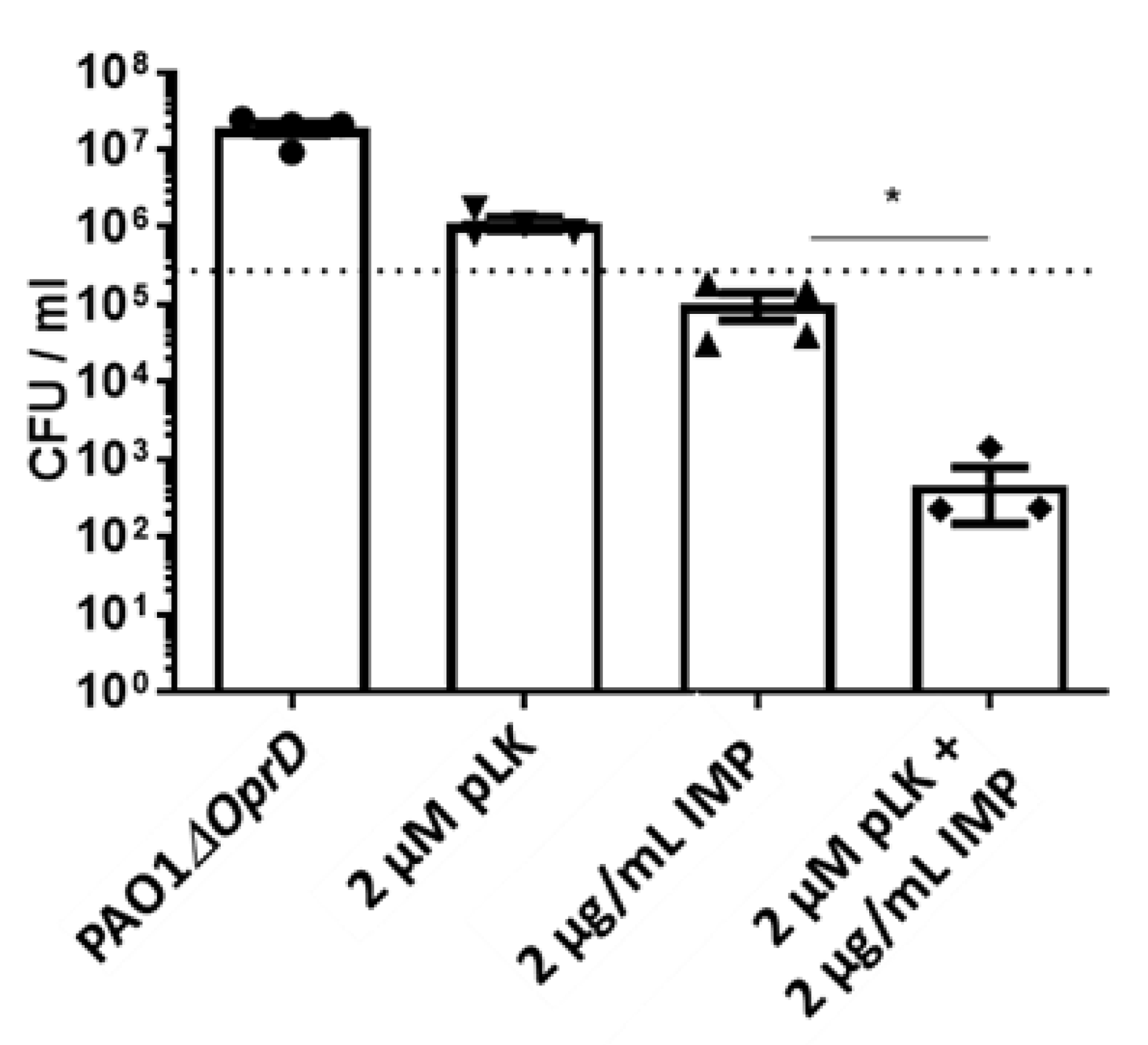
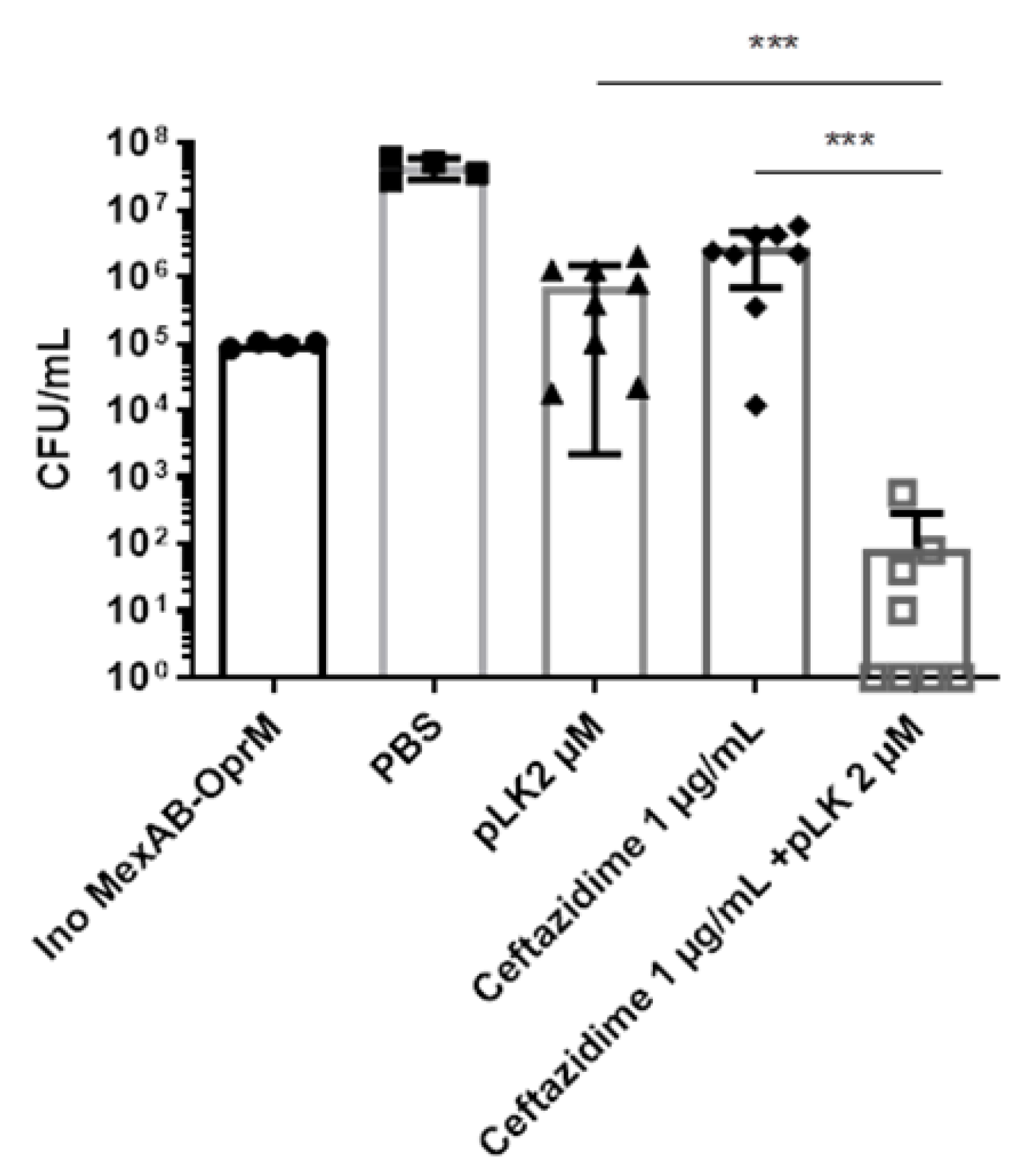
2.3. In Vitro and Ex Vivo Synergistic Effect of the Association of pLK and Imipenem against PAO1ΔoprD and Imipenem-Resistant Clinical P. aeruginosa Strains
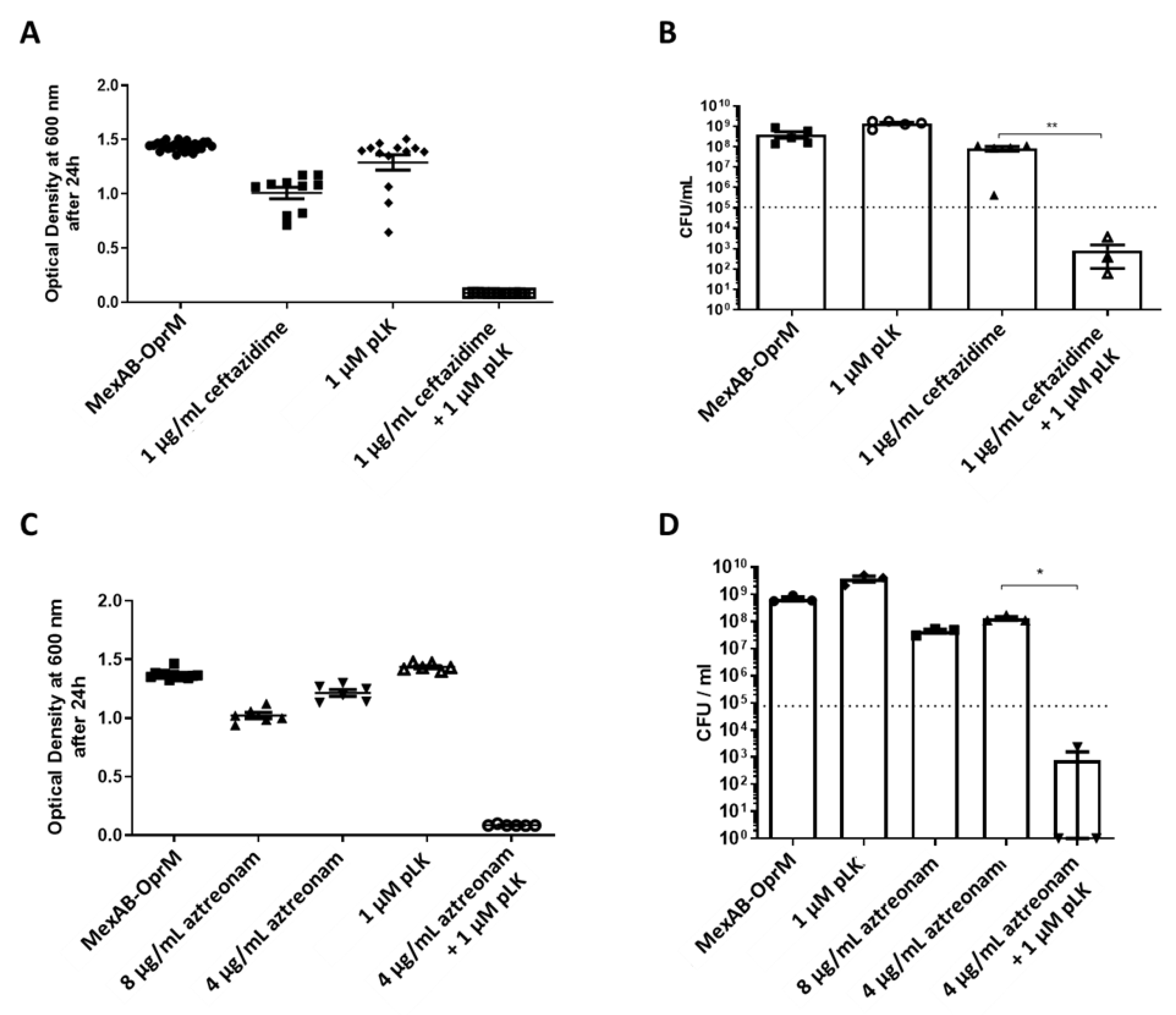
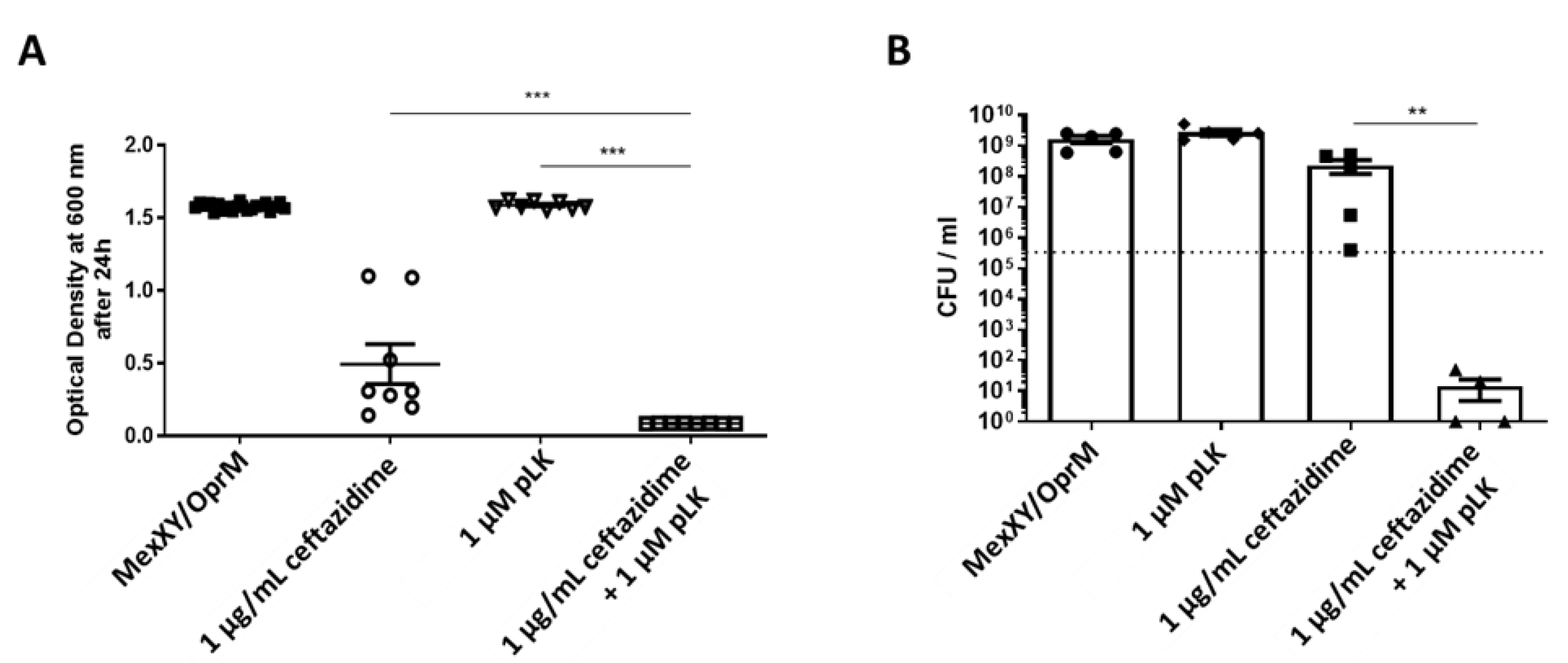
2.4. In Vitro Synergistic Effect of the Association of pLK and Ceftazidime or Aztreonam on MexAB-OprM and MexXY/OprM Clinical P. aeruginosa Isolates
3. Discussion
4. Materials and Methods
4.1. Chemical Reagents
4.2. Bacterial Strains and Culture Conditions
4.3. Scanning Electron Microscopy
4.4. Membrane Permeabilization Assay
4.5. Susceptibility Testing
4.6. Synergy Determination
4.7. Air–Liquid Interface (ALI) PBEC
4.8. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- O’Neill, J. Tackling Drug-Resistant Infections Globally: Final Report and Recommendations. Review on Antimicrobial Resistance; The Welcome Trust: London, UK, 2016. [Google Scholar]
- De Oliveira, D.M.P.; Forde, B.M.; Kidd, T.J.; Harris, P.N.A.; Schembri, M.A.; Beatson, S.A.; Paterson, D.L.; Walker, M.J. Antimicrobial Resistance in ESKAPE Pathogens. Clin. Microbiol. Rev. 2020, 33, e00181-19. [Google Scholar] [CrossRef] [PubMed]
- Pendleton, J.N.; Gorman, S.P.; Gilmore, B.F. Clinical Relevance of the ESKAPE Pathogens. Expert Rev. Anti-Infect. Ther. 2013, 11, 297–308. [Google Scholar] [CrossRef]
- Streeter, K.; Neuman, C.; Thompson, J.; Hatje, E.; Katouli, M. The Characteristics of Genetically Related Pseudomonas Aeruginosa from Diverse Sources and Their Interaction with Human Cell Lines. Can. J. Microbiol. 2016, 62, 233–240. [Google Scholar] [CrossRef] [PubMed]
- Barbier, F.; Andremont, A.; Wolff, M.; Bouadma, L. Hospital-Acquired Pneumonia and Ventilator-Associated Pneumonia: Recent Advances in Epidemiology and Management. Curr. Opin. Pulm. Med. 2013, 19, 216–228. [Google Scholar] [CrossRef] [PubMed]
- Flume, P.A.; VanDevanter, D.R. Clinical Applications of Pulmonary Delivery of Antibiotics. Adv. Drug Deliv. Rev. 2015, 85, 1–6. [Google Scholar] [CrossRef]
- Taccetti, G.; Francalanci, M.; Pizzamiglio, G.; Messore, B.; Carnovale, V.; Cimino, G.; Cipolli, M. Cystic Fibrosis: Recent Insights into Inhaled Antibiotic Treatment and Future Perspectives. Antibiotics 2021, 10, 338. [Google Scholar] [CrossRef]
- Hirsch, E.B.; Tam, V.H. Impact of Multidrug-Resistant Pseudomonas Aeruginosa Infection on Patient Outcomes. Expert Rev. Pharm. Outcomes Res. 2010, 10, 441–451. [Google Scholar] [CrossRef]
- Ikuta, K.S.; Swetschinski, L.R.; Robles Aguilar, G.; Sharara, F.; Mestrovic, T.; Gray, A.P.; Davis Weaver, N.; Wool, E.E.; Han, C.; Gershberg Hayoon, A.; et al. Global Mortality Associated with 33 Bacterial Pathogens in 2019: A Systematic Analysis for the Global Burden of Disease Study 2019. Lancet 2022, 400, 2221–2248. [Google Scholar] [CrossRef]
- Pang, Z.; Raudonis, R.; Glick, B.R.; Lin, T.-J.; Cheng, Z. Antibiotic Resistance in Pseudomonas Aeruginosa: Mechanisms and Alternative Therapeutic Strategies. Biotechnol. Adv. 2019, 37, 177–192. [Google Scholar] [CrossRef]
- Li, X.Z.; Nikaido, H.; Poole, K. Role of MexA-MexB-OprM in Antibiotic Efflux in Pseudomonas Aeruginosa. Antimicrob. Agents Chemother. 1995, 39, 1948–1953. [Google Scholar] [CrossRef] [Green Version]
- Jeannot, K.; Sobel, M.L.; El Garch, F.; Poole, K.; Plésiat, P. Induction of the MexXY Efflux Pump in Pseudomonas Aeruginosa Is Dependent on Drug-Ribosome Interaction. J. Bacteriol. 2005, 187, 5341–5346. [Google Scholar] [CrossRef]
- Aires, J.R.; Köhler, T.; Nikaido, H.; Plésiat, P. Involvement of an Active Efflux System in the Natural Resistance of Pseudomonas Aeruginosa to Aminoglycosides. Antimicrob. Agents Chemother. 1999, 43, 2624–2628. [Google Scholar] [CrossRef]
- Lyczak, J.B.; Cannon, C.L.; Pier, G.B. Lung Infections Associated with Cystic Fibrosis. Clin. Microbiol. Rev. 2002, 15, 194–222. [Google Scholar] [CrossRef]
- Flemming, H.-C.; Wingender, J. The Biofilm Matrix. Nat. Rev. Microbiol. 2010, 8, 623–633. [Google Scholar] [CrossRef]
- Shortridge, D.; Gales, A.C.; Streit, J.M.; Huband, M.D.; Tsakris, A.; Jones, R.N. Geographic and Temporal Patterns of Antimicrobial Resistance in Pseudomonas Aeruginosa Over 20 Years from the SENTRY Antimicrobial Surveillance Program, 1997–2016. Open Forum Infect. Dis. 2019, 6, S63–S68. [Google Scholar] [CrossRef]
- Appaneal, H.J.; Caffrey, A.R.; Lopes, V.; Piehl, E.C.; Puzniak, L.A.; LaPlante, K.L. Assessing Rates of Co-Resistance and Patient Outcomes in Multidrug-Resistant Pseudomonas Aeruginosa. Microbiol. Spectr. 2022, 10, e02336-22. [Google Scholar] [CrossRef]
- Barbier, F.; Wolff, M. Multirésistance Chez Pseudomonas Aeruginosa: Vers l’impasse Thérapeutique? Med. Sci. 2010, 26, 960–968. [Google Scholar] [CrossRef]
- Smith, W.D.; Bardin, E.; Cameron, L.; Edmondson, C.L.; Farrant, K.V.; Martin, I.; Murphy, R.A.; Soren, O.; Turnbull, A.R.; Wierre-Gore, N.; et al. Current and Future Therapies for Pseudomonas Aeruginosa Infection in Patients with Cystic Fibrosis. FEMS Microbiol. Lett. 2017, 364, fnx121. [Google Scholar] [CrossRef]
- Kipnis, E.; Sawa, T.; Wiener-Kronish, J. Targeting Mechanisms of Pseudomonas Aeruginosa Pathogenesis. Médecine Mal. Infect. 2006, 36, 78–91. [Google Scholar] [CrossRef]
- Bals, R.; Hubert, D.; Tümmler, B. Antibiotic Treatment of CF Lung Disease: From Bench to Bedside. J. Cyst. Fibros. 2011, 10, S146–S151. [Google Scholar] [CrossRef] [Green Version]
- Bellemare, A.; Vernoux, N.; Morin, S.; Gagné, S.M.; Bourbonnais, Y. Structural and Antimicrobial Properties of Human Pre-Elafin/Trappin-2 and Derived Peptides against Pseudomonas Aeruginosa. BMC Microbiol. 2010, 10, 253. [Google Scholar] [CrossRef]
- Dubois, A.V.; Midoux, P.; Gras, D.; Si-Tahar, M.; Bréa, D.; Attucci, S.; Khelloufi, M.-K.; Ramphal, R.; Diot, P.; Gauthier, F.; et al. Poly-l-Lysine Compacts DNA, Kills Bacteria, and Improves Protease Inhibition in Cystic Fibrosis Sputum. Am. J. Respir. Crit. Care Med. 2013, 188, 703–709. [Google Scholar] [CrossRef]
- Guillon, A.; Fouquenet, D.; Morello, E.; Henry, C.; Georgeault, S.; Si-Tahar, M.; Hervé, V. Treatment of Pseudomonas Aeruginosa Biofilm Present in Endotracheal Tubes by Poly-l-Lysine. Antimicrob. Agents Chemother. 2018, 62, e00564-18. [Google Scholar] [CrossRef]
- Pai, M.P.; Cottrell, M.L.; Kashuba, A.D.M.; Bertino, J.S. Pharmacokinetics and Pharmacodynamics of Anti-Infective Agents. In Mandell, Douglas, and Bennett’s Principles and Practice of Infectious Diseases; Elsevier: Amsterdam, The Netherlands, 2015; pp. 252–262.e2. ISBN 978-1-4557-4801-3. [Google Scholar]
- Hall, M.J.; Middleton, R.F.; Westmacott, D. The Fractional Inhibitory Concentration (FIC) Index as a Measure of Synergy. J. Antimicrob. Chemother. 1983, 11, 427–433. [Google Scholar] [CrossRef]
- Botelho, J.; Grosso, F.; Peixe, L. Antibiotic Resistance in Pseudomonas Aeruginosa—Mechanisms, Epidemiology and Evolution. Drug Resist. Updates 2019, 44, 100640. [Google Scholar] [CrossRef]
- Plackett, B. Why Big Pharma Has Abandoned Antibiotics. Nature 2020, 586, S50–S52. [Google Scholar] [CrossRef]
- Davido, B.; Fellous, L.; Lawrence, C.; Maxime, V.; Rottman, M.; Dinh, A. Ceftazidime-Avibactam and Aztreonam, an Interesting Strategy to Overcome β-Lactam Resistance Conferred by Metallo-β-Lactamases in Enterobacteriaceae and Pseudomonas Aeruginosa. Antimicrob. Agents Chemother. 2017, 61, e01008-17. [Google Scholar] [CrossRef]
- Chatterjee, M.; Anju, C.P.; Biswas, L.; Anil Kumar, V.; Gopi Mohan, C.; Biswas, R. Antibiotic Resistance in Pseudomonas Aeruginosa and Alternative Therapeutic Options. Int. J. Med. Microbiol. 2016, 306, 48–58. [Google Scholar] [CrossRef]
- Wang, N.; Chen, X.; Luo, J.; Deng, F.; Shi, F.; Wu, Q.; Huang, Y.; Ouyang, Q.; Qin, R.; Zhou, H. Artemisinin Derivative DHA27 Enhances the Antibacterial Effect of Aminoglycosides against Pseudomonas Aeruginosa by Inhibiting MRNA Expression of Aminoglycoside-Modifying Enzymes. Front. Pharm. 2022, 13, 970400. [Google Scholar] [CrossRef]
- Tuersuntuoheti, T.; Wang, Z.; Wang, Z.; Liang, S.; Li, X.; Zhang, M. Review of the Application of Ε-poly-L-lysine in Improving Food Quality and Preservation. J. Food Process. Preserv. 2019, 43, e14153. [Google Scholar] [CrossRef]
- Shima, S.; Matsuoka, H.; Iwamoto, T.; Sakai, H. Antimicrobial Action of ε-Poly-L-Lysine. J. Antibiot. 1984, 37, 1449–1455. [Google Scholar] [CrossRef]
- Sundaran, S.; Kok, L.-C.; Chang, H.-Y. Combination Effect of Epsilon-Poly-L-Lysine and Antibiotics against Common Bacterial Pathogens. J. Antibiot. 2022, 75, 354–359. [Google Scholar] [CrossRef] [PubMed]
- Wei, L.; Wu, R.; Wang, C.; Wu, Z. Effects of ε-Polylysine on Pseudomonas Aeruginosa and Aspergillus Fumigatus Biofilm In Vitro. Med. Sci. Monit. 2017, 23, 4225–4229. [Google Scholar] [CrossRef] [PubMed]
- Hamano, Y. Occurrence, Biosynthesis, Biodegradation, and Industrial and Medical Applications of a Naturally Occurring ε-Poly-L-Lysine. Biosci. Biotechnol. Biochem. 2011, 75, 1226–1233. [Google Scholar] [CrossRef] [PubMed]
- May, K.L.; Grabowicz, M. The Bacterial Outer Membrane Is an Evolving Antibiotic Barrier. Proc. Natl. Acad. Sci. USA 2018, 115, 8852–8854. [Google Scholar] [CrossRef] [PubMed]
- Brogden, K.A. Antimicrobial Peptides: Pore Formers or Metabolic Inhibitors in Bacteria? Nat. Rev. Microbiol. 2005, 3, 238–250. [Google Scholar] [CrossRef] [PubMed]
- Yoshida, T.; Nagasawa, T. ??-Poly-l-Lysine: Microbial Production, Biodegradation and Application Potential. Appl. Microbiol. Biotechnol. 2003, 62, 21–26. [Google Scholar] [CrossRef]
- Falagas, M.E.; Kasiakou, S.K.; Saravolatz, L.D. Colistin: The Revival of Polymyxins for the Management of Multidrug-Resistant Gram-Negative Bacterial Infections. Clin. Infect. Dis. 2005, 40, 1333–1341. [Google Scholar] [CrossRef]
- Wang, G.; Brunel, J.-M.; Preusse, M.; Mozaheb, N.; Willger, S.D.; Larrouy-Maumus, G.; Baatsen, P.; Häussler, S.; Bolla, J.-M.; Van Bambeke, F. The Membrane-Active Polyaminoisoprenyl Compound NV716 Re-Sensitizes Pseudomonas Aeruginosa to Antibiotics and Reduces Bacterial Virulence. Commun. Biol. 2022, 5, 871. [Google Scholar] [CrossRef]
- Rázquin-Olazarán, I.; Shahrour, H.; Martínez-de-Tejada, G. A Synthetic Peptide Sensitizes Multi-Drug Resistant Pseudomonas Aeruginosa to Antibiotics for More than Two Hours and Permeabilizes Its Envelope for Twenty Hours. J. Biomed. Sci. 2020, 27, 85. [Google Scholar] [CrossRef]
- Egorov, A.M.; Ulyashova, M.M.; Rubtsova, M.Y. Bacterial Enzymes and Antibiotic Resistance. Acta Nat. 2018, 10, 33–48. [Google Scholar] [CrossRef]
- Sims, P.J.; Waggoner, A.S.; Wang, C.-H.; Hoffman, J.F. Mechanism by Which Cyanine Dyes Measure Membrane Potential in Red Blood Cells and Phosphatidylcholine Vesicles. Biochemistry 1974, 13, 3315–3330. [Google Scholar] [CrossRef] [PubMed]
- M07-A11; Methods for Dilution Antimicrobial Susceptibility Tests for Bacteria That Grow Aerobically. 10th ed. Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2018.
- Acar, J.F. Antibiotic Synergy and Antagonism. Med. Clin. N. Am. 2000, 84, 1391–1406. [Google Scholar] [CrossRef] [PubMed]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cezard, A.; Fouquenet, D.; Vasseur, V.; Jeannot, K.; Launay, F.; Si-Tahar, M.; Hervé, V. Poly-L-Lysine to Fight Antibiotic Resistances of Pseudomonas aeruginosa. Int. J. Mol. Sci. 2023, 24, 2851. https://doi.org/10.3390/ijms24032851
Cezard A, Fouquenet D, Vasseur V, Jeannot K, Launay F, Si-Tahar M, Hervé V. Poly-L-Lysine to Fight Antibiotic Resistances of Pseudomonas aeruginosa. International Journal of Molecular Sciences. 2023; 24(3):2851. https://doi.org/10.3390/ijms24032851
Chicago/Turabian StyleCezard, Adeline, Delphine Fouquenet, Virginie Vasseur, Katy Jeannot, Fabien Launay, Mustapha Si-Tahar, and Virginie Hervé. 2023. "Poly-L-Lysine to Fight Antibiotic Resistances of Pseudomonas aeruginosa" International Journal of Molecular Sciences 24, no. 3: 2851. https://doi.org/10.3390/ijms24032851