
Insights into the Peritumoural Brain Zone of Glioblastoma: CDK4 and EXT2 May Be Potential Drivers of Malignancy
 , , , , , , ,
, , , , , , , 
Abstract
:1. Introduction
2. Results
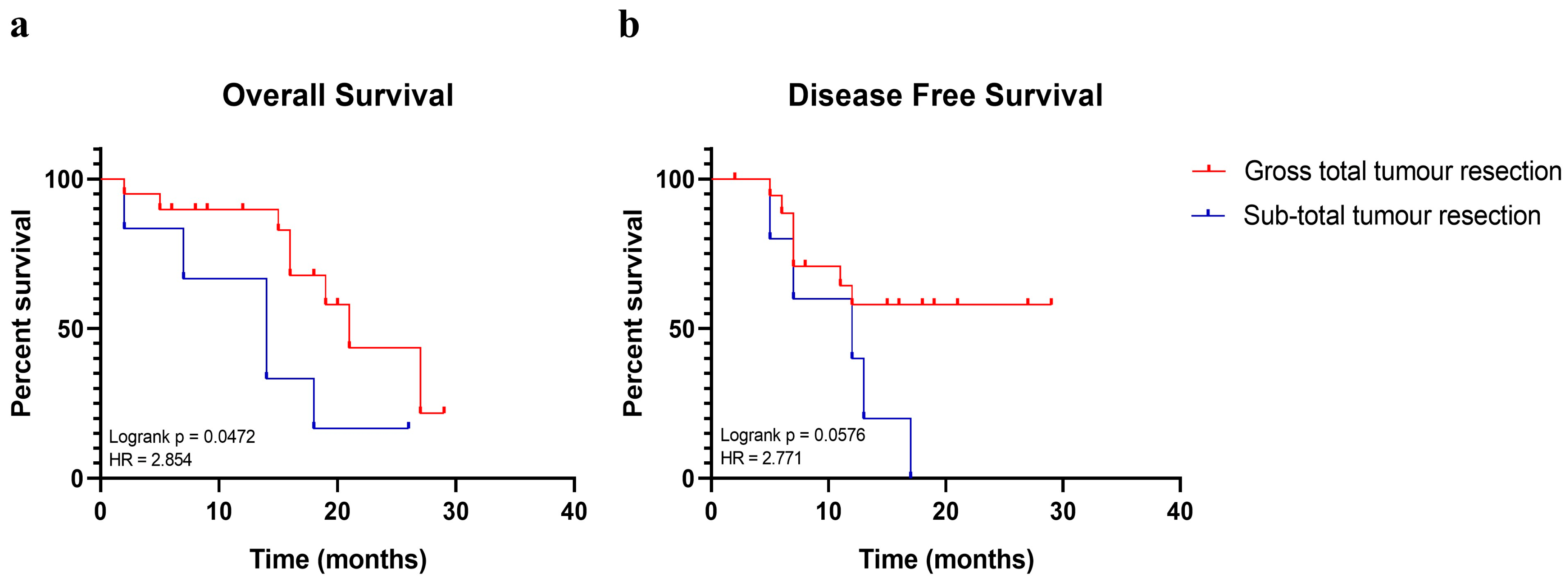
2.1. Gross Total Tumour Resection Affects Patients’ Overall Survival
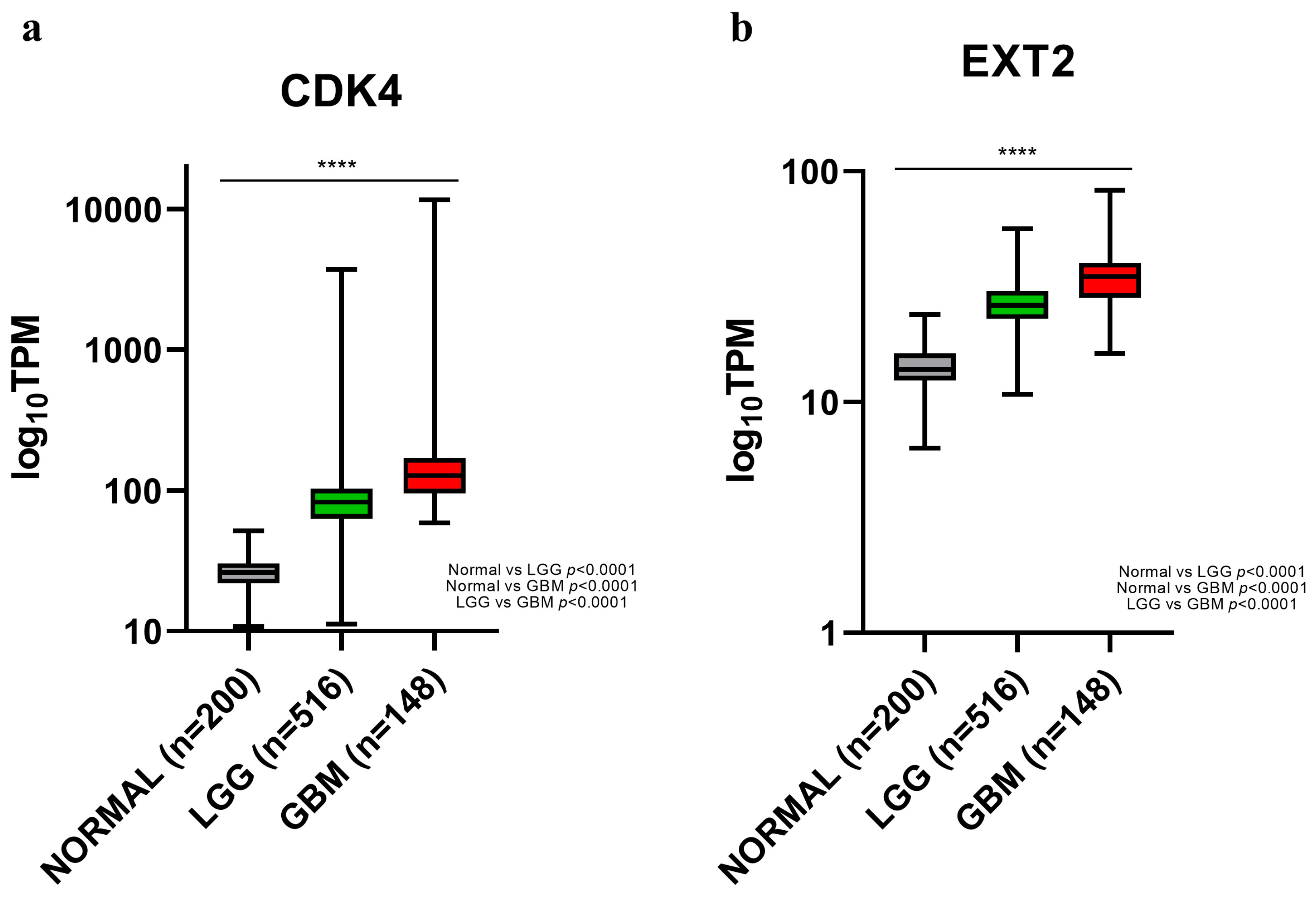
2.2. Genes Involved in Copy Number Gains Were Also Transcriptionally Over-Expressed in PBZs
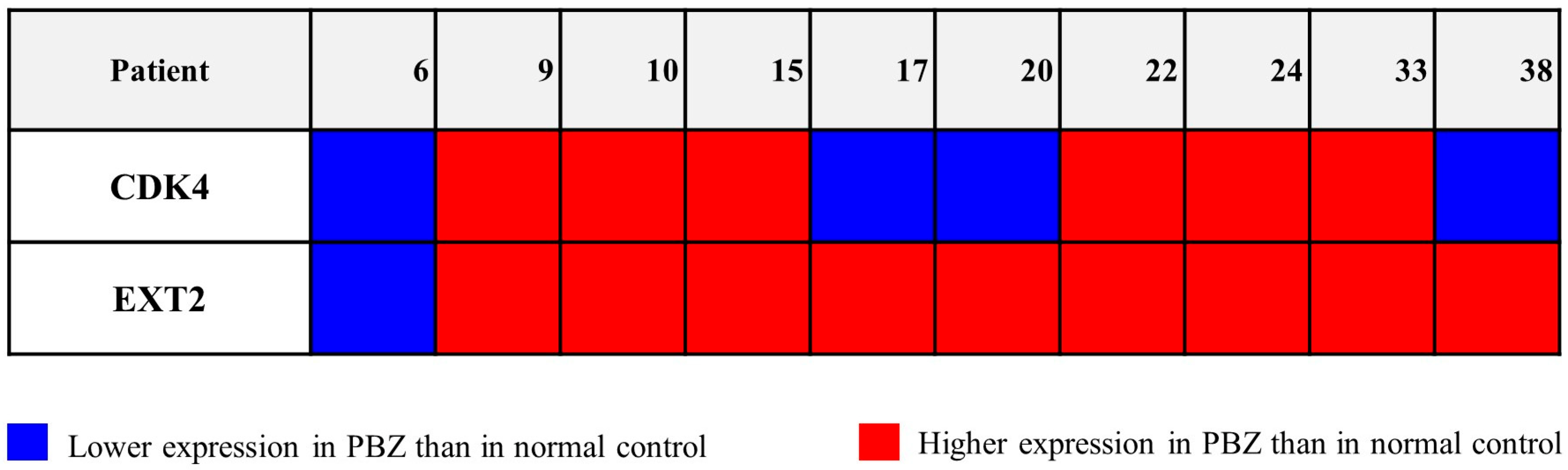
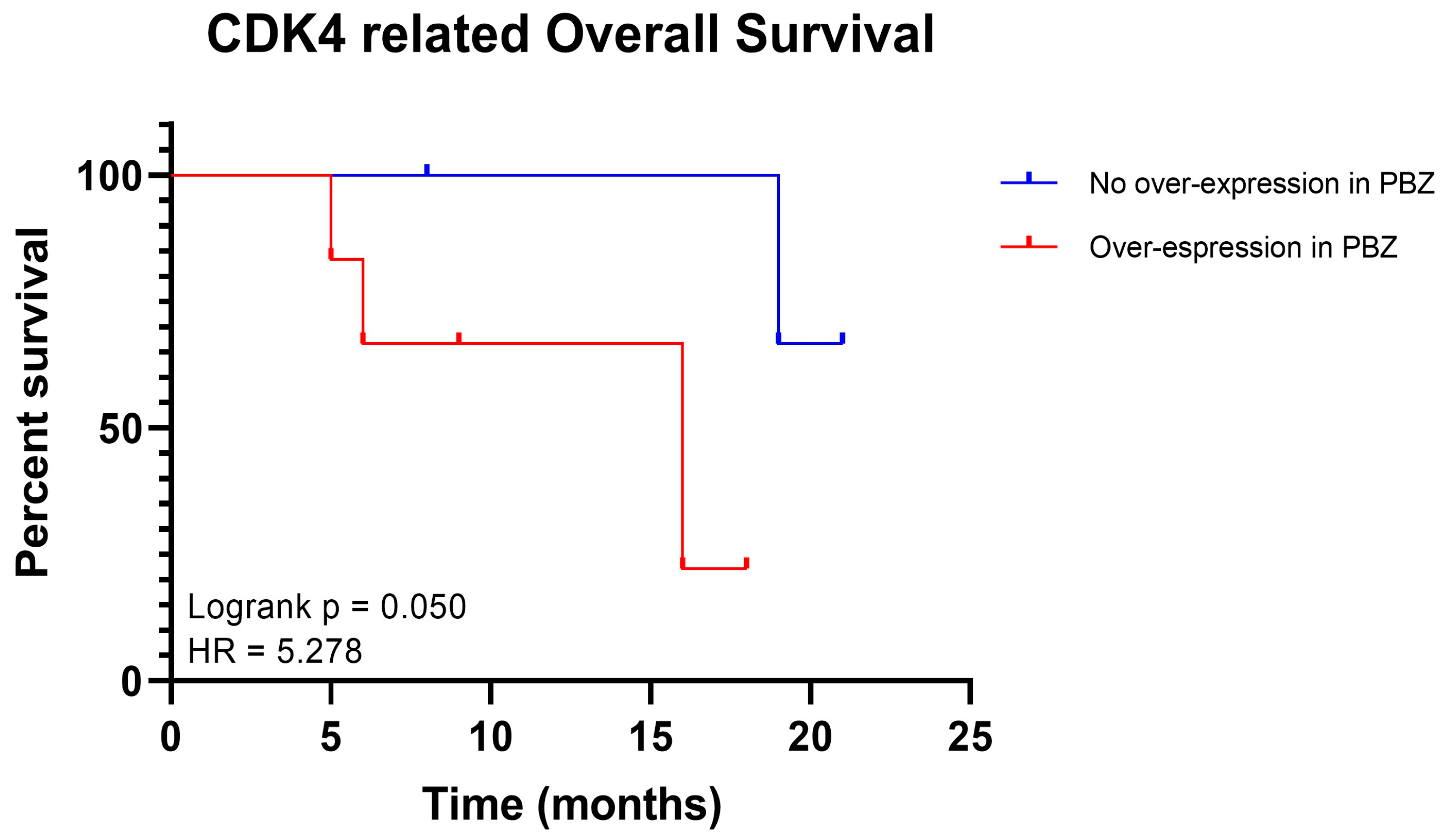
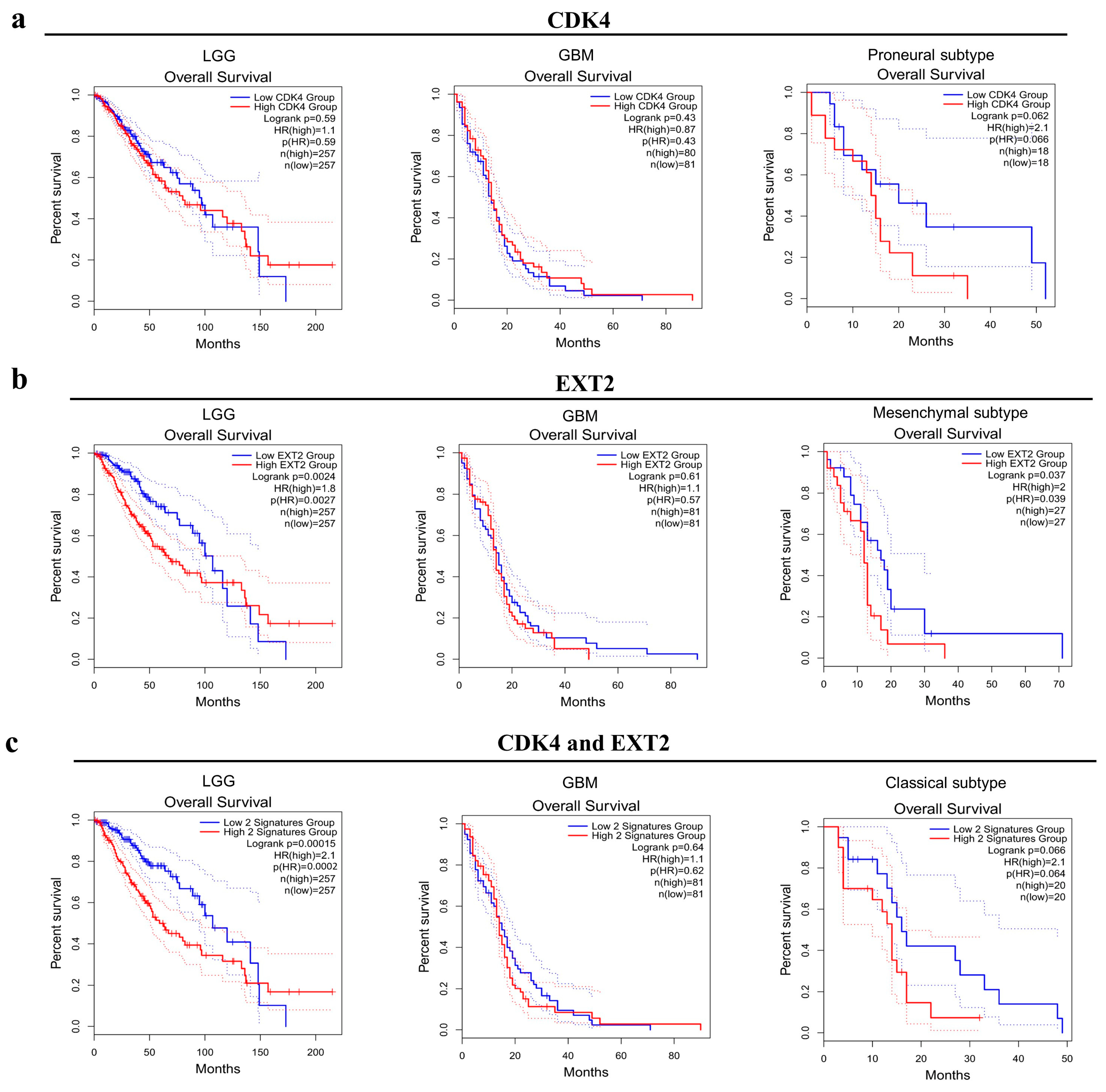
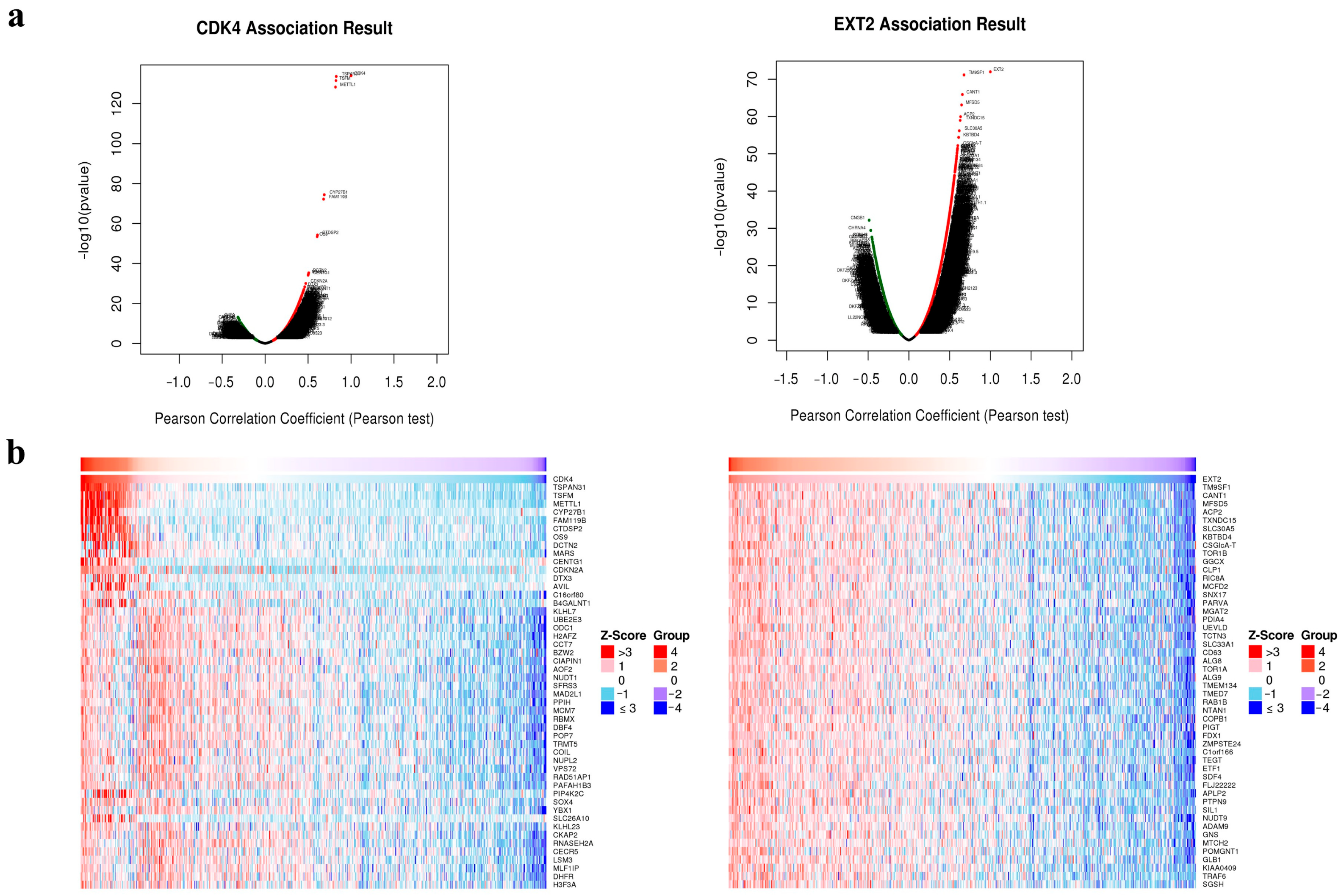
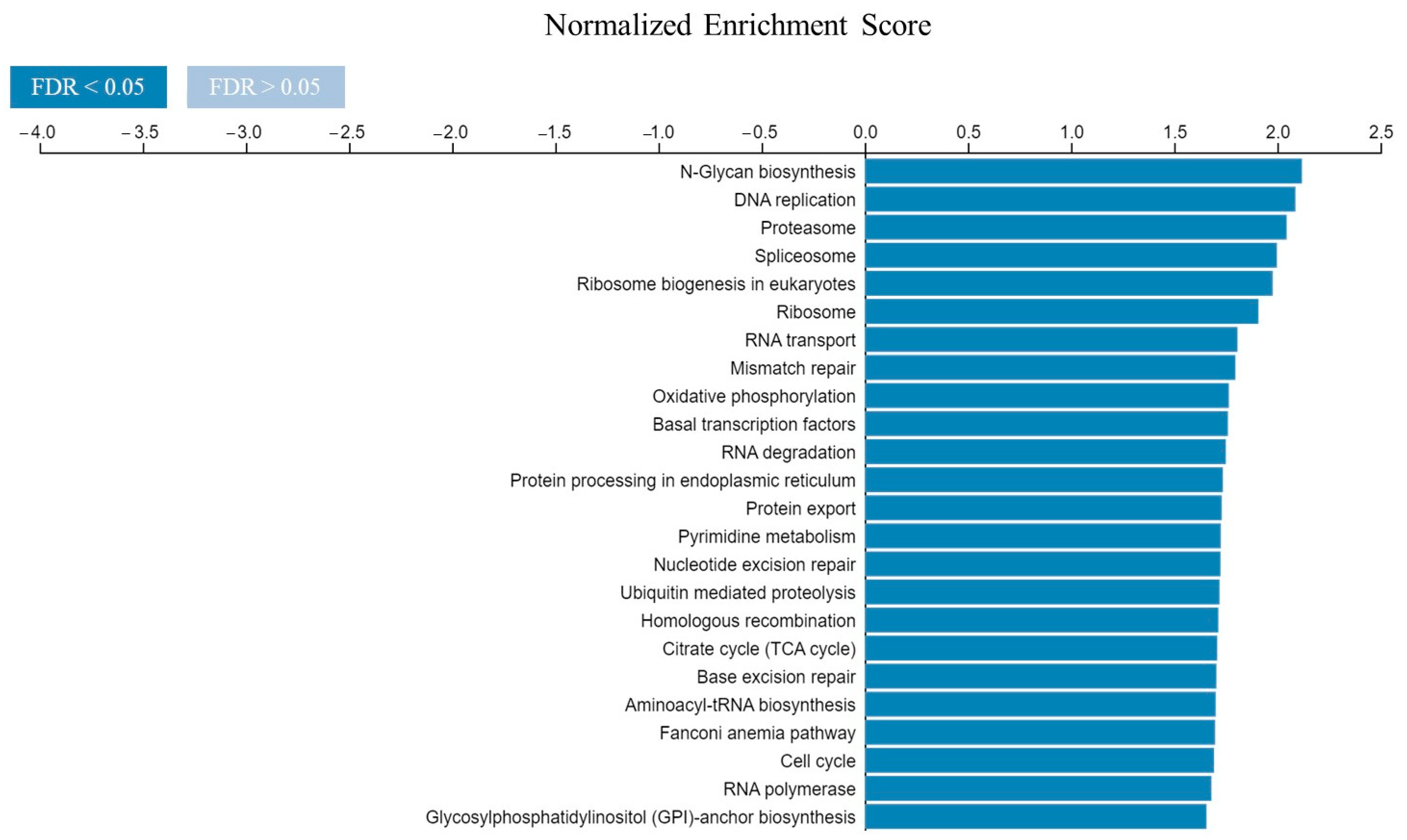
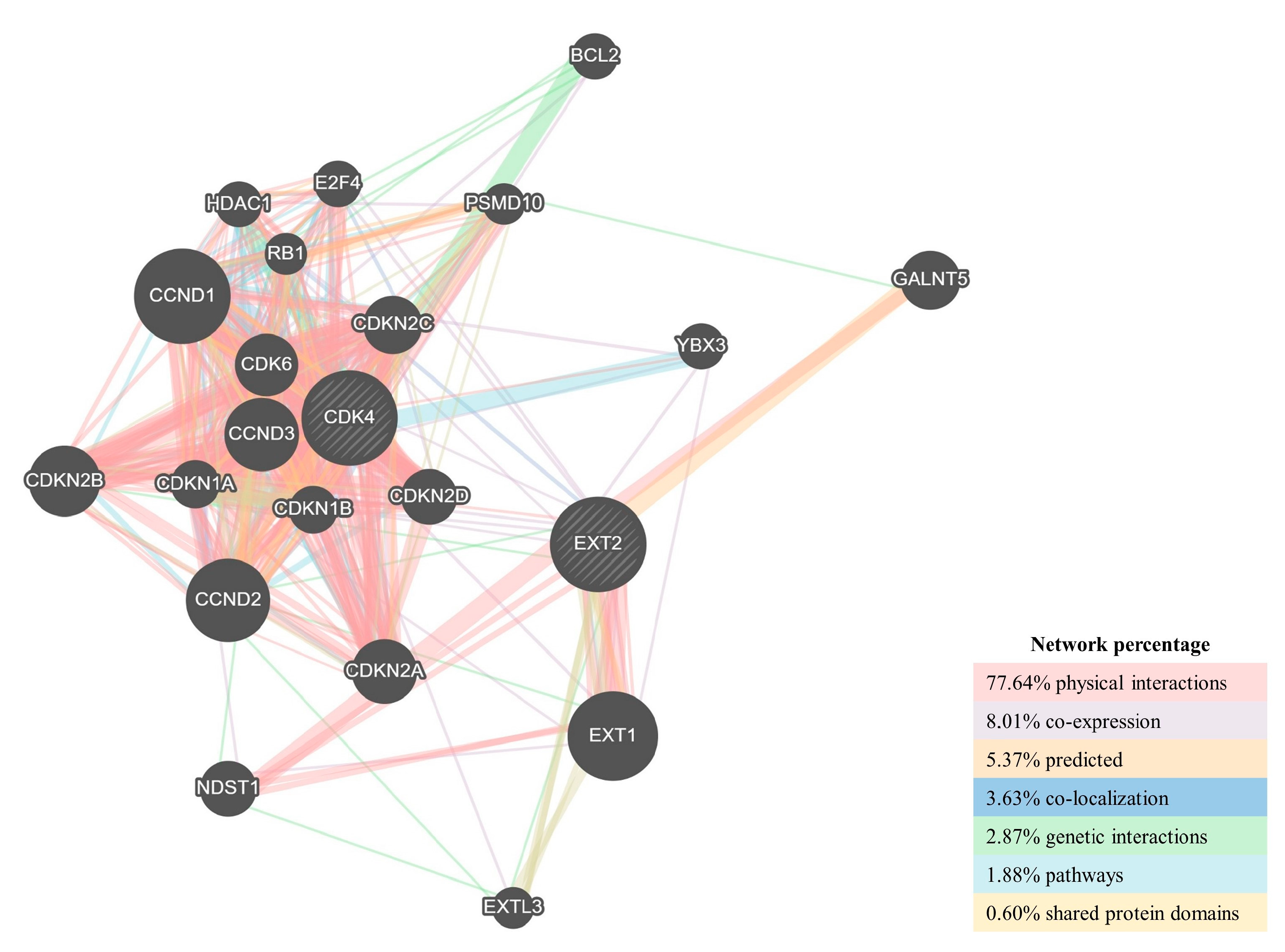
2.3. Potential Role of CDK4 and EXT2 in the Malignant Transformation of the PBZ
3. Discussion
4. Materials and Methods
4.1. Study Population
4.2. Immuno-Molecular Analysis
4.3. Genomic Analysis by Array-Based Genomic Comparative Hybridization
4.4. Strategies Adopted to Select Interesting Genes of the PBZ Involved in Copy Number Alterations
4.4.1. Selection of CNAs
4.4.2. Selection of Genes
4.5. TaqMan Gene Expression Assay
4.6. Databases
4.6.1. Selection of CDK4 and EXT2
4.6.2. Analysis of CDK4 and EXT2 Expression in Low-Grade Glioma and Glioblastoma
4.6.3. Survival Analysis and Relationship with GBM Molecular Subtypes
4.6.4. CDK4 and EXT2 Co-Expression Networks
4.6.5. Protein–Protein Interaction Network Construction
4.7. Statistics
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ostrom, Q.T.; Cote, D.J.; Ascha, M.; Kruchko, C.; Barnholtz-Sloan, J.S. Adult Glioma Incidence and Survival by Race or Ethnicity in the United States from 2000 to 2014. JAMA Oncol. 2018, 4, 1254–1262. [Google Scholar] [CrossRef]
- Stupp, R.; Hegi, M.E.; Mason, W.P.; van den Bent, M.J.; Taphoorn, M.J.; Janzer, R.C.; Ludwin, S.K.; Allgeier, A.; Fisher, B.; Belanger, K.; et al. Effects of radiotherapy with concomitant and adjuvant temozolomide versus radiotherapy alone on survival in glioblastoma in a randomised phase III study: 5-year analysis of the EORTC-NCIC trial. Lancet Oncol. 2009, 10, 459–466. [Google Scholar] [CrossRef] [PubMed]
- Giambra, M.; Di Cristofori, A.; Valtorta, S.; Manfrellotti, R.; Bigiogera, V.; Basso, G.; Moresco, R.M.; Giussani, C.; Bentivegna, A. The peritumoral brain zone in glioblastoma: Where we are and where we are going. J. Neurosci. Res. 2022, 101, 199–216. [Google Scholar] [CrossRef] [PubMed]
- Ricard, D.; Idbaih, A.; Ducray, F.; Lahutte, M.; Hoang-Xuan, K.; Delattre, J.Y. Primary brain tumours in adults. Lancet 2012, 379, 1984–1996. [Google Scholar] [CrossRef] [PubMed]
- Petrecca, K.; Guiot, M.C.; Panet-Raymond, V.; Souhami, L. Failure pattern following complete resection plus radiotherapy and temozolomide is at the resection margin in patients with glioblastoma. J. Neurooncol. 2013, 111, 19–23. [Google Scholar] [CrossRef] [PubMed]
- Seker-Polat, F.; Pinarbasi Degirmenci, N.; Solaroglu, I.; Bagci-Onder, T. Tumor Cell Infiltration into the Brain in Glioblastoma: From Mechanisms to Clinical Perspectives. Cancers 2022, 14, 443. [Google Scholar] [CrossRef]
- D’Alessandro, G.; Catalano, M.; Sciaccaluga, M.; Chece, G.; Cipriani, R.; Rosito, M.; Grimaldi, A.; Lauro, C.; Cantore, G.; Santoro, A.; et al. KCa3.1 channels are involved in the infiltrative behavior of glioblastoma in vivo. Cell Death Dis. 2013, 4, e773. [Google Scholar] [CrossRef] [PubMed]
- Alieva, M.; Leidgens, V.; Riemenschneider, M.J.; Klein, C.A.; Hau, P.; van Rheenen, J. Intravital imaging of glioma border morphology reveals distinctive cellular dynamics and contribution to tumor cell invasion. Sci. Rep. 2019, 9, 2054. [Google Scholar] [CrossRef]
- Bello, L.; Gambini, A.; Castellano, A.; Carrabba, G.; Acerbi, F.; Fava, E.; Giussani, C.; Cadioli, M.; Blasi, V.; Casarotti, A.; et al. Motor and language DTI Fiber Tracking combined with intraoperative subcortical mapping for surgical removal of gliomas. Neuroimage 2008, 39, 369–382. [Google Scholar] [CrossRef]
- Certo, F.; Stummer, W.; Farah, J.O.; Freyschlag, C.; Visocchi, M.; Morrone, A.; Altieri, R.; Toccaceli, G.; Peschillo, S.; Thomè, C.; et al. Supramarginal resection of glioblastoma: 5-ALA fluorescence, combined intraoperative strategies and correlation with survival. J. Neurosurg. Sci. 2019, 63, 625–632. [Google Scholar] [CrossRef]
- Giambra, M.; Messuti, E.; Di Cristofori, A.; Cavandoli, C.; Bruno, R.; Buonanno, R.; Marzorati, M.; Zambuto, M.; Rodriguez-Menendez, V.; Redaelli, S.; et al. Characterizing the Genomic Profile in High-Grade Gliomas: From Tumor Core to Peritumoral Brain Zone, Passing through Glioma-Derived Tumorspheres. Biology 2021, 10, 1157. [Google Scholar] [CrossRef]
- Bertolini, I.; Terrasi, A.; Martelli, C.; Gaudioso, G.; Di Cristofori, A.; Storaci, A.M.; Formica, M.; Braidotti, P.; Todoerti, K.; Ferrero, S.; et al. A GBM-like V-ATPase signature directs cell-cell tumor signaling and reprogramming via large oncosomes. EBioMedicine 2019, 41, 225–235. [Google Scholar] [CrossRef]
- Bertolini, I.; Storaci, A.M.; Terrasi, A.; Cristofori, A.D.; Locatelli, M.; Caroli, M.; Ferrero, S.; Altieri, D.C.; Vaira, V. Interplay Between V-ATPase G1 and Small EV-miRNAs Modulates ERK1/2 Activation in GBM Stem Cells and Nonneoplastic Milieu. Mol. Cancer Res. 2020, 18, 1744–1754. [Google Scholar] [CrossRef] [PubMed]
- Molinaro, A.M.; Hervey-Jumper, S.; Morshed, R.A.; Young, J.; Han, S.J.; Chunduru, P.; Zhang, Y.; Phillips, J.J.; Shai, A.; Lafontaine, M.; et al. Association of Maximal Extent of Resection of Contrast-Enhanced and Non-Contrast-Enhanced Tumor with Survival within Molecular Subgroups of Patients with Newly Diagnosed Glioblastoma. JAMA Oncol. 2020, 6, 495–503. [Google Scholar] [CrossRef]
- Altieri, R.; Barbagallo, D.; Certo, F.; Broggi, G.; Ragusa, M.; Di Pietro, C.; Caltabiano, R.; Magro, G.; Peschillo, S.; Purrello, M.; et al. Peritumoral Microenvironment in High-Grade Gliomas: From FLAIRectomy to Microglia-Glioma Cross-Talk. Brain Sci. 2021, 11, 200. [Google Scholar] [CrossRef] [PubMed]
- Otto, T.; Sicinski, P. Cell cycle proteins as promising targets in cancer therapy. Nat. Rev. Cancer 2017, 17, 93–115. [Google Scholar] [CrossRef]
- Wu, D.; Huo, C.; Jiang, S.; Huang, Y.; Fang, X.; Liu, J.; Yang, M.; Ren, J.; Xu, B.; Liu, Y. Exostosin1 as a novel prognostic and predictive biomarker for squamous cell lung carcinoma: A study based on bioinformatics analysis. Cancer Med. 2021, 10, 2787–2801. [Google Scholar] [CrossRef]
- Wu, H.; Zhao, X.; Zhu, T.; Rong, D.; Wang, Y.; Leng, D.; Wu, D. A Glycosyltransferase-Related Signature for Predicting Overall Survival in Head and Neck Squamous Cell Carcinoma. Front. Genet. 2022, 13, 856671. [Google Scholar] [CrossRef] [PubMed]
- Verhaak, R.G.; Hoadley, K.A.; Purdom, E.; Wang, V.; Qi, Y.; Wilkerson, M.D.; Miller, C.R.; Ding, L.; Golub, T.; Mesirov, J.P.; et al. Integrated genomic analysis identifies clinically relevant subtypes of glioblastoma characterized by abnormalities in PDGFRA, IDH1, EGFR, and NF1. Cancer Cell 2010, 17, 98–110. [Google Scholar] [CrossRef]
- Ma, S.; Rudra, S.; Campian, J.L.; Dahiya, S.; Dunn, G.P.; Johanns, T.; Goldstein, M.; Kim, A.H.; Huang, J. Prognostic impact of CDKN2A/B deletion, TERT mutation, and EGFR amplification on histological and molecular IDH-wildtype glioblastoma. Neurooncol. Adv. 2020, 2, vdaa126. [Google Scholar] [CrossRef]
- Kruthika, B.S.; Sugur, H.; Nandaki, K.; Arimappamagan, A.; Paturu, K.; Santosh, V. Expression pattern and prognostic significance of myosin light chain 9 (MYL9): A novel biomarker in glioblastoma. J. Clin. Pathol. 2019, 72, 677–681. [Google Scholar] [CrossRef]
- Kruthika, B.S.; Jain, R.; Arivazhagan, A.; Bharath, R.D.; Yasha, T.C.; Kondaiah, P.; Santosh, V. Transcriptome profiling reveals PDZ binding kinase as a novel biomarker in peritumoral brain zone of glioblastoma. J. Neurooncol. 2019, 141, 315–325. [Google Scholar] [CrossRef] [PubMed]
- Nimbalkar, V.P.; Kruthika, B.S.; Sravya, P.; Rao, S.; Sugur, H.S.; Chickabasaviah, Y.T.; Somanna, S.; Arivazhagan, A.; Kondaiah, P.; Santosh, V. Chitinase 3-Like 2. Am. J. Clin. Pathol. 2022, 158, 521–529. [Google Scholar] [CrossRef] [PubMed]
- Nimbalkar, V.P.; Kruthika, B.S.; Sravya, P.; Rao, S.; Sugur, H.S.; Verma, B.K.; Chickabasaviah, Y.T.; Arivazhagan, A.; Kondaiah, P.; Santosh, V. Differential gene expression in peritumoral brain zone of glioblastoma: Role of SERPINA3 in promoting invasion, stemness and radioresistance of gli.ioma cells and association with poor patient prognosis and recurrence. J. Neurooncol. 2021, 152, 55–65. [Google Scholar] [CrossRef] [PubMed]
- Cai, H.; Kumar, N.; Baudis, M. arrayMap: A reference resource for genomic copy number imbalances in human malignancies. PLoS ONE 2012, 7, e36944. [Google Scholar] [CrossRef] [PubMed]
- Baronchelli, S.; Bentivegna, A.; Redaelli, S.; Riva, G.; Butta, V.; Paoletta, L.; Isimbaldi, G.; Miozzo, M.; Tabano, S.; Daga, A.; et al. Delineating the cytogenomic and epigenomic landscapes of glioma stem cell lines. PLoS ONE 2013, 8, e57462. [Google Scholar] [CrossRef]
- Conconi, D.; Jemma, A.; Giambra, M.; Redaelli, S.; Croci, G.A.; Dalprà, L.; Lavitrano, M.; Bentivegna, A. Analysis of copy number alterations in bladder cancer stem cells revealed a prognostic role of LRP1B. World J. Urol. 2022, 40, 2267–2273. [Google Scholar] [CrossRef]
- Conconi, D.; Redaelli, S.; Lissoni, A.A.; Cilibrasi, C.; Perego, P.; Gautiero, E.; Sala, E.; Paderno, M.; Dalprà, L.; Landoni, F.; et al. Genomic and Epigenomic Profile of Uterine Smooth Muscle Tumors of Uncertain Malignant Potential (STUMPs) Revealed Similarities and Differences with Leiomyomas and Leiomyosarcomas. Int. J. Mol. Sci. 2021, 22, 1580. [Google Scholar] [CrossRef]
- Bowers, R.R.; Jones, C.M.; Paz, E.A.; Barrows, J.K.; Armeson, K.E.; Long, D.T.; Delaney, J.R. SWAN pathway-network identification of common aneuploidy-based oncogenic drivers. Nucleic Acids Res. 2022, 50, 3673–3692. [Google Scholar] [CrossRef]
- Labani, M.; Afrasiabi, A.; Beheshti, A.; Lovell, N.H.; Alinejad-Rokny, H. PeakCNV: A multi-feature ranking algorithm-based tool for genome-wide copy number variation-association study. Comput. Struct. Biotechnol. J. 2022, 20, 4975–4983. [Google Scholar] [CrossRef]
- Pesenti, C.; Navone, S.E.; Guarnaccia, L.; Terrasi, A.; Costanza, J.; Silipigni, R.; Guarneri, S.; Fusco, N.; Fontana, L.; Locatelli, M.; et al. The Genetic Landscape of Human Glioblastoma and Matched Primary Cancer Stem Cells Reveals Intratumour Similarity and Intertumour Heterogeneity. Stem Cells Int. 2019, 2019, 2617030. [Google Scholar] [CrossRef] [PubMed]
- Lemée, J.M.; Clavreul, A.; Aubry, M.; Com, E.; de Tayrac, M.; Eliat, P.A.; Henry, C.; Rousseau, A.; Mosser, J.; Menei, P. Characterizing the peritumoral brain zone in glioblastoma: A multidisciplinary analysis. J. Neurooncol. 2015, 122, 53–61. [Google Scholar] [CrossRef]
- Shao, Y.; Shen, Y.Q.; Li, Y.L.; Liang, C.; Zhang, B.J.; Lu, S.D.; He, Y.Y.; Wang, P.; Sun, Q.L.; Jin, Y.X.; et al. Direct repression of the oncogene CDK4 by the tumor suppressor miR-486-5p in non-small cell lung cancer. Oncotarget 2016, 7, 34011–34021. [Google Scholar] [CrossRef]
- Anders, L.; Ke, N.; Hydbring, P.; Choi, Y.J.; Widlund, H.R.; Chick, J.M.; Zhai, H.; Vidal, M.; Gygi, S.P.; Braun, P.; et al. A systematic screen for CDK4/6 substrates links FOXM1 phosphorylation to senescence suppression in cancer cells. Cancer Cell 2011, 20, 620–634. [Google Scholar] [CrossRef] [PubMed]
- Heilmann, A.M.; Perera, R.M.; Ecker, V.; Nicolay, B.N.; Bardeesy, N.; Benes, C.H.; Dyson, N.J. CDK4/6 and IGF1 receptor inhibitors synergize to suppress the growth of p16INK4A-deficient pancreatic cancers. Cancer Res. 2014, 74, 3947–3958. [Google Scholar] [CrossRef] [PubMed]
- Ortega, S.; Malumbres, M.; Barbacid, M. Cyclin D-dependent kinases, INK4 inhibitors and cancer. Biochim. Biophys. Acta 2002, 1602, 73–87. [Google Scholar] [CrossRef]
- Cadoo, K.A.; Gucalp, A.; Traina, T.A. Palbociclib: An evidence-based review of its potential in the treatment of breast cancer. Breast Cancer Targets Ther. 2014, 6, 123–133. [Google Scholar] [CrossRef]
- Royce, M.; Osgood, C.; Mulkey, F.; Bloomquist, E.; Pierce, W.F.; Roy, A.; Kalavar, S.; Ghosh, S.; Philip, R.; Rizvi, F.; et al. FDA Approval Summary: Abemaciclib with Endocrine Therapy for High-Risk Early Breast Cancer. J. Clin. Oncol. 2022, 40, 1155–1162. [Google Scholar] [CrossRef] [PubMed]
- Chong, Q.Y.; Kok, Z.H.; Bui, N.L.; Xiang, X.; Wong, A.L.; Yong, W.P.; Sethi, G.; Lobie, P.E.; Wang, L.; Goh, B.C. A unique CDK4/6 inhibitor: Current and future therapeutic strategies of abemaciclib. Pharmacol. Res. 2020, 156, 104686. [Google Scholar] [CrossRef]
- Cen, L.; Carlson, B.L.; Schroeder, M.A.; Ostrem, J.L.; Kitange, G.J.; Mladek, A.C.; Fink, S.R.; Decker, P.A.; Wu, W.; Kim, J.S.; et al. p16-Cdk4-Rb axis controls sensitivity to a cyclin-dependent kinase inhibitor PD0332991 in glioblastoma xenograft cells. Neuro Oncol. 2012, 14, 870–881. [Google Scholar] [CrossRef] [Green Version]
- Wiedemeyer, W.R.; Dunn, I.F.; Quayle, S.N.; Zhang, J.; Chheda, M.G.; Dunn, G.P.; Zhuang, L.; Rosenbluh, J.; Chen, S.; Xiao, Y.; et al. Pattern of retinoblastoma pathway inactivation dictates response to CDK4/6 inhibition in GBM. Proc. Natl. Acad. Sci. USA 2010, 107, 11501–11506. [Google Scholar] [CrossRef] [PubMed]
- Cemeli, T.; Guasch-Vallés, M.; Nàger, M.; Felip, I.; Cambray, S.; Santacana, M.; Gatius, S.; Pedraza, N.; Dolcet, X.; Ferrezuelo, F.; et al. Cytoplasmic cyclin D1 regulates glioblastoma dissemination. J. Pathol. 2019, 248, 501–513. [Google Scholar] [CrossRef] [PubMed]
- Wishart, D.S.; Feunang, Y.D.; Guo, A.C.; Lo, E.J.; Marcu, A.; Grant, J.R.; Sajed, T.; Johnson, D.; Li, C.; Sayeeda, Z.; et al. DrugBank 5.0: A major update to the DrugBank database for 2018. Nucleic Acids Res. 2018, 46, D1074–D1082. [Google Scholar] [CrossRef] [PubMed]
- Busse, M.; Feta, A.; Presto, J.; Wilén, M.; Grønning, M.; Kjellén, L.; Kusche-Gullberg, M. Contribution of EXT1, EXT2, and EXTL3 to heparan sulfate chain elongation. J. Biol. Chem. 2007, 282, 32802–32810. [Google Scholar] [CrossRef] [PubMed]
- Knelson, E.H.; Nee, J.C.; Blobe, G.C. Heparan sulfate signaling in cancer. Trends Biochem. Sci. 2014, 39, 277–288. [Google Scholar] [CrossRef]
- Epis, M.R.; Giles, K.M.; Candy, P.A.; Webster, R.J.; Leedman, P.J. miR-331-3p regulates expression of neuropilin-2 in glioblastoma. J. Neurooncol. 2014, 116, 67–75. [Google Scholar] [CrossRef]
- Iozzo, R.V.; Sanderson, R.D. Proteoglycans in cancer biology, tumour microenvironment and angiogenesis. J. Cell. Mol. Med. 2011, 15, 1013–1031. [Google Scholar] [CrossRef]
- Ushakov, V.S.; Tsidulko, A.Y.; de La Bourdonnaye, G.; Kazanskaya, G.M.; Volkov, A.M.; Kiselev, R.S.; Kobozev, V.V.; Kostromskaya, D.V.; Gaytan, A.S.; Krivoshapkin, A.L.; et al. Heparan Sulfate Biosynthetic System Is Inhibited in Human Glioma Due to EXT1/2 and HS6ST1/2 Down-Regulation. Int. J. Mol. Sci. 2017, 18, 2301. [Google Scholar] [CrossRef]
- Cheng, L.; Wang, P.; Yang, S.; Yang, Y.; Zhang, Q.; Zhang, W.; Xiao, H.; Gao, H. Identification of genes with a correlation between copy number and expression in gastric cancer. BMC Med. Genom. 2012, 5, 14. [Google Scholar] [CrossRef]
- Shao, X.; Lv, N.; Liao, J.; Long, J.; Xue, R.; Ai, N.; Xu, D.; Fan, X. Copy number variation is highly correlated with differential gene expression: A pan-cancer study. BMC Med. Genet. 2019, 20, 175. [Google Scholar] [CrossRef]
- Mischel, P.S.; Shai, R.; Shi, T.; Horvath, S.; Lu, K.V.; Choe, G.; Seligson, D.; Kremen, T.J.; Palotie, A.; Liau, L.M.; et al. Identification of molecular subtypes of glioblastoma by gene expression profiling. Oncogene 2003, 22, 2361–2373. [Google Scholar] [CrossRef]
- Lee, E.; Yong, R.L.; Paddison, P.; Zhu, J. Comparison of glioblastoma (GBM) molecular classification methods. Semin Cancer Biol. 2018, 53, 201–211. [Google Scholar] [CrossRef] [PubMed]
- Brennan, C.W.; Verhaak, R.G.; McKenna, A.; Campos, B.; Noushmehr, H.; Salama, S.R.; Zheng, S.; Chakravarty, D.; Sanborn, J.Z.; Berman, S.H.; et al. The somatic genomic landscape of glioblastoma. Cell 2013, 155, 462–477. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Wang, X.; Xu, L.; Zhang, J.; Cao, H. Analysis of the EGFR Amplification and CDKN2A Deletion Regulated Transcriptomic Signatures Reveals the Prognostic Significance of. Front. Oncol. 2021, 11, 551160. [Google Scholar] [CrossRef] [PubMed]
- Funakoshi, Y.; Hata, N.; Takigawa, K.; Arita, H.; Kuga, D.; Hatae, R.; Sangatsuda, Y.; Fujioka, Y.; Sako, A.; Umehara, T.; et al. Clinical significance of CDKN2A homozygous deletion in combination with methylated MGMT status for IDH-wildtype glioblastoma. Cancer Med. 2021, 10, 3177–3187. [Google Scholar] [CrossRef]
- Lu, V.M.; O’Connor, K.P.; Shah, A.H.; Eichberg, D.G.; Luther, E.M.; Komotar, R.J.; Ivan, M.E. The prognostic significance of CDKN2A homozygous deletion in IDH-mutant lower-grade glioma and glioblastoma: A systematic review of the contemporary literature. J. Neurooncol. 2020, 148, 221–229. [Google Scholar] [CrossRef]
- Louis, D.N.; Perry, A.; Wesseling, P.; Brat, D.J.; Cree, I.A.; Figarella-Branger, D.; Hawkins, C.; Ng, H.K.; Pfister, S.M.; Reifenberger, G.; et al. The 2021 WHO Classification of Tumors of the Central Nervous System: A summary. Neuro Oncol. 2021, 23, 1231–1251. [Google Scholar] [CrossRef]
- Al-Holou, W.N.; Hodges, T.R.; Everson, R.G.; Freeman, J.; Zhou, S.; Suki, D.; Rao, G.; Ferguson, S.D.; Heimberger, A.B.; McCutcheon, I.E.; et al. Perilesional Resection of Glioblastoma Is Independently Associated with Improved Outcomes. Neurosurgery 2020, 86, 112–121. [Google Scholar] [CrossRef]
- Sanai, N.; Polley, M.Y.; McDermott, M.W.; Parsa, A.T.; Berger, M.S. An extent of resection threshold for newly diagnosed glioblastomas. J. Neurosurg. 2011, 115, 3–8. [Google Scholar] [CrossRef]
- Stupp, R.; Hegi, M.E.; Gilbert, M.R.; Chakravarti, A. Chemoradiotherapy in malignant glioma: Standard of care and future directions. J. Clin. Oncol. 2007, 25, 4127–4136. [Google Scholar] [CrossRef]
- Łysiak, M.; Das, J.; Malmström, A.; Söderkvist, P. Methylation associated with long- or short-term survival in glioblastoma patients from the Nordic phase 3 trial. Front. Genet. 2022, 13, 934519. [Google Scholar] [CrossRef]
- Wen, P.Y.; Macdonald, D.R.; Reardon, D.A.; Cloughesy, T.F.; Sorensen, A.G.; Galanis, E.; Degroot, J.; Wick, W.; Gilbert, M.R.; Lassman, A.B.; et al. Updated response assessment criteria for high-grade gliomas: Response assessment in neuro-oncology working group. J. Clin. Oncol. 2010, 28, 1963–1972. [Google Scholar] [CrossRef]
- Marchi, F.; Sahnane, N.; Cerutti, R.; Cipriani, D.; Barizzi, J.; Stefanini, F.M.; Epistolio, S.; Cerati, M.; Balbi, S.; Mazzucchelli, L.; et al. The Impact of Surgery in IDH 1 Wild Type Glioblastoma in Relation with the MGMT Deregulation. Front. Oncol. 2019, 9, 1569. [Google Scholar] [CrossRef]
- Tang, G.; Cho, M.; Wang, X. OncoDB: An interactive online database for analysis of gene expression and viral infection in cancer. Nucleic Acids Res. 2022, 50, D1334–D1339. [Google Scholar] [CrossRef] [PubMed]
- Tang, Z.; Kang, B.; Li, C.; Chen, T.; Zhang, Z. GEPIA2: An enhanced web server for large-scale expression profiling and interactive analysis. Nucleic Acids Res. 2019, 47, W556–W560. [Google Scholar] [CrossRef]
- Vasaikar, S.V.; Straub, P.; Wang, J.; Zhang, B. LinkedOmics: Analyzing multi-omics data within and across 32 cancer types. Nucleic Acids Res. 2018, 46, D956–D963. [Google Scholar] [CrossRef] [PubMed]
- Warde-Farley, D.; Donaldson, S.L.; Comes, O.; Zuberi, K.; Badrawi, R.; Chao, P.; Franz, M.; Grouios, C.; Kazi, F.; Lopes, C.T.; et al. The GeneMANIA prediction server: Biological network integration for gene prioritization and predicting gene function. Nucleic Acids Res. 2010, 38, W214–W220. [Google Scholar] [CrossRef]
- Gamboa, N.T.; Crabb, B.; Henson, J.C.; Cole, K.L.; Weaver, B.D.; Karsy, M.; Jensen, R.L. High-grade glioma imaging volumes and survival: A single-institution analysis of 101 patients after resection using intraoperative MRI. J. Neurooncol. 2022, 160, 555–565. [Google Scholar] [CrossRef]
- Chaichana, K.L.; Jusue-Torres, I.; Navarro-Ramirez, R.; Raza, S.M.; Pascual-Gallego, M.; Ibrahim, A.; Hernandez-Hermann, M.; Gomez, L.; Ye, X.; Weingart, J.D.; et al. Establishing percent resection and residual volume thresholds affecting survival and recurrence for patients with newly diagnosed intracranial glioblastoma. Neuro Oncol. 2014, 16, 113–122. [Google Scholar] [CrossRef] [PubMed]
- Muscas, G.; Orlandini, S.; Bonaudo, C.; Dardo, M.; Esposito, A.; Campagnaro, L.; Carrai, R.; Fainardi, E.; Ciccarino, P.; Della Puppa, A. Functional outcomes, extent of resection, and bright/vague fluorescence interface in resection of glioblastomas involving the motor pathways assisted by 5-ALA. Acta Neurochir. 2022, 164, 3267–3274. [Google Scholar] [CrossRef]
- De Witt Hamer, P.C.; Robles, S.G.; Zwinderman, A.H.; Duffau, H.; Berger, M.S. Impact of intraoperative stimulation brain mapping on glioma surgery outcome: A meta-analysis. J. Clin. Oncol. 2012, 30, 2559–2565. [Google Scholar] [CrossRef]
- Weller, M.; van den Bent, M.; Preusser, M.; Le Rhun, E.; Tonn, J.C.; Minniti, G.; Bendszus, M.; Balana, C.; Chinot, O.; Dirven, L.; et al. EANO guidelines on the diagnosis and treatment of diffuse gliomas of adulthood. Nat. Rev. Clin. Oncol. 2021, 18, 170–186. [Google Scholar] [CrossRef]
- Lee, S.Y. Temozolomide resistance in glioblastoma multiforme. Genes Dis. 2016, 3, 198–210. [Google Scholar] [CrossRef]
- Wu, W.; Klockow, J.L.; Zhang, M.; Lafortune, F.; Chang, E.; Jin, L.; Wu, Y.; Daldrup-Link, H.E. Glioblastoma multiforme (GBM): An overview of current therapies and mechanisms of resistance. Pharmacol. Res. 2021, 171, 105780. [Google Scholar] [CrossRef]
- Malacrida, A.; Di Domizio, A.; Bentivegna, A.; Cislaghi, G.; Messuti, E.; Tabano, S.M.; Giussani, C.; Zuliani, V.; Rivara, M.; Nicolini, G. MV1035 Overcomes Temozolomide Resistance in Patient-Derived Glioblastoma Stem Cell Lines. Biology 2022, 11, 70. [Google Scholar] [CrossRef]
- Wijaya, J.; Fukuda, Y.; Schuetz, J.D. Obstacles to Brain Tumor Therapy: Key ABC Transporters. Int. J. Mol. Sci. 2017, 18, 2544. [Google Scholar] [CrossRef]
- Di Cristofori, A.; Ferrero, S.; Bertolini, I.; Gaudioso, G.; Russo, M.V.; Berno, V.; Vanini, M.; Locatelli, M.; Zavanone, M.; Rampini, P.; et al. The vacuolar H+ ATPase is a novel therapeutic target for glioblastoma. Oncotarget 2015, 6, 17514–17531. [Google Scholar] [CrossRef] [Green Version]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Patient | Sex | Age | Tumour Location | Immuno-phenotype | GTR | DFS (Months) | OS (Months) | PBZ Biopsy |
|---|---|---|---|---|---|---|---|---|
| 1 | M | 71 | Right parietal lobe | ATRX+, p53−, MGMT met | No | 12 | 14 | No |
| 2 | M | 54 | Right frontal lobe | ATRX−, p53+, MGMT met | Yes | No progression | 27 | No |
| 3 | F | 61 | Left temporal lobe | ATRX+, p53−, MGMT met | Yes | No progression | Alive (29) | No |
| 5 | M | 57 | Left temporal lobe | ATRX+, p53−, MGMT wt | Yes | No progression | 21 | No |
| 6 | M | 44 | Right temporal lobe | ATRX+, p53−, MGMT wt | Yes | 11 | 19 | Yes |
| 7 | M | 54 | Right parietal lobe | ATRX+, p53−, MGMT wt | Yes | 6 | 15 | No |
| 8 | M | 57 | Right temporal lobe | ATRX+, p53−, MGMT met | No | 17 | Alive (26) | No |
| 9 | M | 71 | Left occipital lobe | ATRX+, p53−, MGMT wt | Yes | 12 | 16 | Yes |
| 10 | M | 74 | Right temporo- occipital lobe | ATRX+, p53−, MGMT met | Yes | No progression | 5 | Yes |
| 11 | M | 76 | Right temporal lobe | ATRX+, p53−, MGMT met | No | 13 | 18 | Yes |
| 12 | F | 70 | Multifocal | ATRX+, p53−, MGMT wt | Yes | <1 | 2 | No |
| 13 | M | 58 | Left temporal lobe | ATRX+, p53−, MGMT met | No | 7 | 14 | No |
| 14 | M | 73 | Left temporal lobe | ATRX+, p53−, MGMT met | - | No progression | <1 (died for PE) | No |
| 15 | M | 78 | Left frontal lobe | ATRX+, p53−, MGMT met | Yes | No progression | 16 | Yes |
| 17 | M | 81 | Right frontal lobe | ATRX+, p53−, MGMT wt | Yes | No progression | Alive (21) | Yes |
| 18 | F | 49 | Right occipital lobe | ATRX+, p53−, MGMT wt | Yes | 7 | Alive (20) | No |
| 20 | F | 60 | Right occipital lobe | ATRX+, p53−, MGMT wt | Yes | No progression | Alive (19) | Yes |
| 21 | M | 39 | Right frontal lobe | ATRX−, p53+, MGMT met | Yes | No progression | Alive (18) | No |
| 22 | M | 53 | Left temporal lobe | ATRX+, p53−, MGMT met | Yes | No progression | Alive (18) | Yes |
| 23 | M | 71 | Left frontal lobe | ATRX+, p53−, MGMT met | No | 5 | 7 | No |
| 24 | M | 69 | Left parieto- occipital lobe | ATRX+, p53−, MGMT wt | Yes | 5 | 6 | Yes |
| 26 | F | 69 | Left parietal lobe | ATRX+, p53−, MGMT met | Yes | No progression | Alive (15) | No |
| 27 | M | 57 | Right temporal lobe | ATRX+, p53−, MGMT wt | No | <1 | 2 | Yes |
| 28 | M | 59 | Right frontal lobe | ATRX+, p53−, MGMT wt | Yes | No progression | Alive (2) | Yes |
| 30 | F | 73 | Right frontal lobe | ATRX+, p53−, MGMT met | - | No progression | <1 (died for ventriculitis) | Yes |
| 31 | M | 63 | Left temporo- occipital lobe | ATRX+, p53−, MGMT wt | Yes | 7 | Alive (12) | Yes |
| 33 | M | 54 | Right temporo-occipital lobe | ATRX−, p53−, MGMT met | Yes | 7 | Alive (9) | Yes |
| 38 | M | 49 | Right parieto-occipital lobe | ATRX+, p53−, MGMT wt | Yes | No progression | Alive (8) | Yes |
| Patient | R | N | DF | Critical Value of R | Correlation |
|---|---|---|---|---|---|
| 10 # | 0.62 | 71 | 69 | 0.232 * | Moderately strong |
| 17 # | 0.42 | 24 | 22 | 0.404 * | Moderately strong |
| 15 # | −0.87 | 24 | 22 | 0.404 * | Low |
| 22 # | −0.52 | 23 | 21 | 0.413 * | Low |
| 24 # | 0.37 | 36 | 34 | 0.325 * | Low |
| 28 | −0.74094 | 13 | 11 | 0.553 * | Low |
| 30 | 0.226147 | 23 | 21 | 0.413 | Low |
| 31 | 0.061161 | 37 | 35 | 0.325 | Low |
| 38 | 0.671874 | 19 | 17 | 0.456 * | Moderately strong |
| CNAs Locus | Patients | List of Genes Affected | |
|---|---|---|---|
| Strategy I | 11p11.2 Gain | 9 22 28 | EXT2 |
| 16p13.3 Gain | 15 22 | SSTR5-AS1, SSTR5, C1QTNF8, CACNA1H, TPSG1, TPSB2,TPSAB1,TPSD1 | |
| Strategy II | 7q21.12 Gain | 11 20 22 27 38 | GRM3, KIAA1324L |
| 10q21.3 Loss | 11 20 22 27 | DNA2, SLC25A16, TET1,CCAR1, SNORD98, STOX1, DDX50,DDX21, KIF1BP, SRGN, VPS26A, SUPV3L1, LOC101928994, HKDC1 | |
| 12q14.1 Gain | 10 22 24 27 | AGAP2, AGAP2-AS1, TSPAN31, CDK4, MIR6759, MARCHF9, CYP27B1, METTL1, EEF1AKMT3, TSFM, AVIL, CTDSP2 | |
| 9p21.3 Loss | 10 11 17 20 22 | CDKN2A-DT, CDKN2A, CDKN2B-AS1, CDKN2B |
| Gene | Cancer-Related Pathway | Associated Tumours | Prognostic Marker | Targeting Compound |
|---|---|---|---|---|
| CDK4 | Cell cycle, p53 signalling pathway, PI3K-Akt signalling pathway, cellular senescence (KEGG database) | Metastatic breast cancer, bladder, head and neck squamous cell carcinoma, oesophageal [16] | Unfavourable in renal and liver cancers (HPA) | Alvocidib, purvalanol, palbociclib, ribociclib, abemaciclib, fostamatinib (DrugBank) |
| EXT2 | Glycosaminoglycan biosynthesis–heparan sulphate/heparin, metabolic pathways (KEGG database) | Squamous cell lung carcinoma [17], head and neck squamous cell carcinoma [18] | Unfavourable in renal and head and neck cancers (HPA) | Uridine-diphosphate-N-acetylgalactosamine, uridine-5′-diphosphate (DrugBank) |
| Type of Sample | CDK4 | EXT2 | ||
|---|---|---|---|---|
| Median Expression | Log2 (Fold Change) | Median Expression | Log2 (Fold Change) | |
| Normal brain tissue | 26.0 | 13.9 | ||
| LGG | 82.5 | 1.67 | 26.3 | 0.92 |
| GBM | 127.1 | 2.29 | 35.0 | 1.32 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Giambra, M.; Di Cristofori, A.; Conconi, D.; Marzorati, M.; Redaelli, S.; Zambuto, M.; Rocca, A.; Roumy, L.; Carrabba, G.; Lavitrano, M.; et al. Insights into the Peritumoural Brain Zone of Glioblastoma: CDK4 and EXT2 May Be Potential Drivers of Malignancy. Int. J. Mol. Sci. 2023, 24, 2835. https://doi.org/10.3390/ijms24032835
Giambra M, Di Cristofori A, Conconi D, Marzorati M, Redaelli S, Zambuto M, Rocca A, Roumy L, Carrabba G, Lavitrano M, et al. Insights into the Peritumoural Brain Zone of Glioblastoma: CDK4 and EXT2 May Be Potential Drivers of Malignancy. International Journal of Molecular Sciences. 2023; 24(3):2835. https://doi.org/10.3390/ijms24032835
Chicago/Turabian StyleGiambra, Martina, Andrea Di Cristofori, Donatella Conconi, Matilde Marzorati, Serena Redaelli, Melissa Zambuto, Alessandra Rocca, Louis Roumy, Giorgio Carrabba, Marialuisa Lavitrano, and et al. 2023. "Insights into the Peritumoural Brain Zone of Glioblastoma: CDK4 and EXT2 May Be Potential Drivers of Malignancy" International Journal of Molecular Sciences 24, no. 3: 2835. https://doi.org/10.3390/ijms24032835