

Transient Changes in the Plasma of Astrocytic and Neuronal Injury Biomarkers in COVID-19 Patients without Neurological Syndromes
 ,
,  , , , ,
, , , ,
Abstract
:1. Introduction
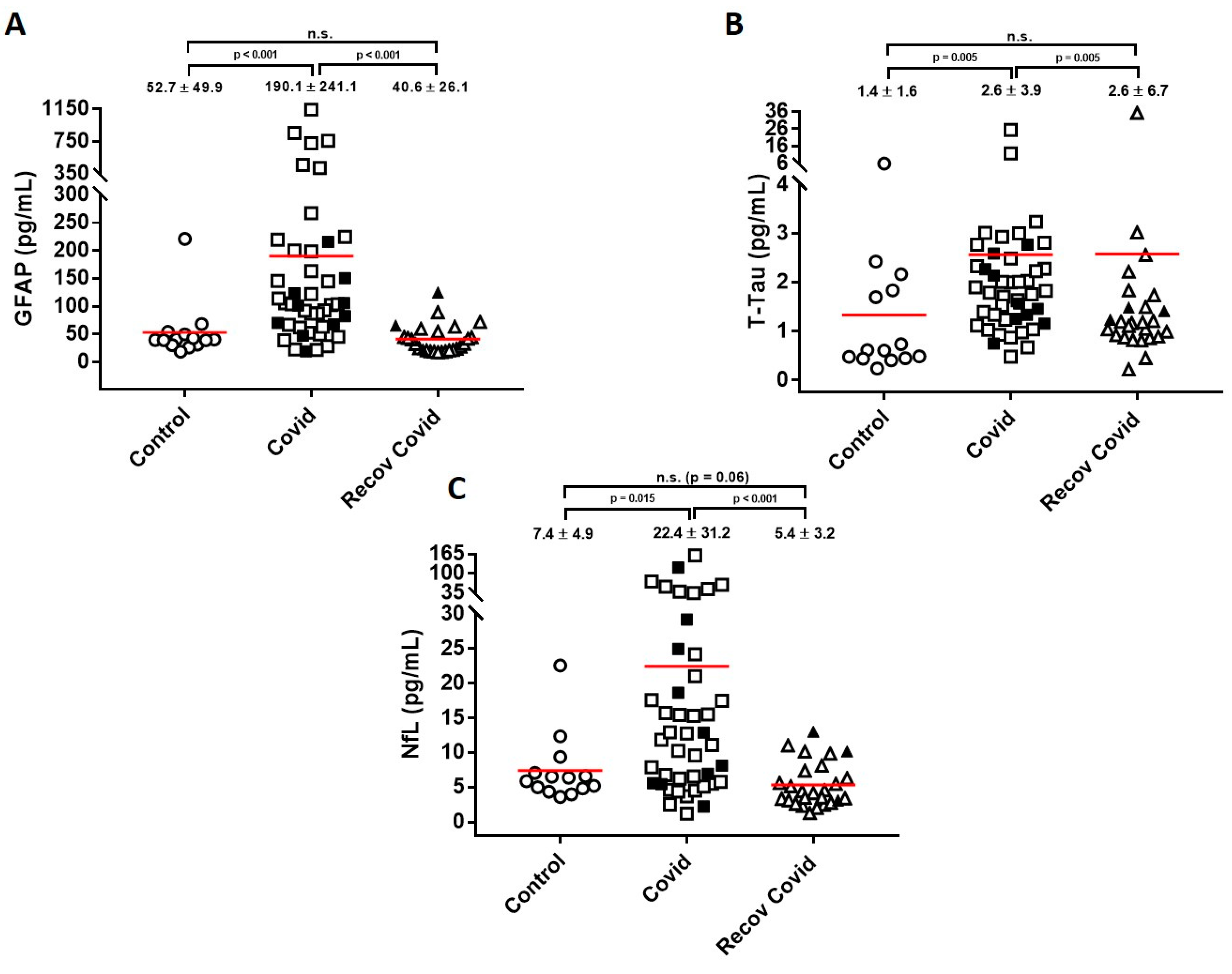
2. Results
3. Discussion
4. Methods
4.1. Plasma Samples
4.2. Biomarker Measurements
4.3. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Ellul, M.A.; Benjamin, L.; Singh, B.; Lant, S.; Michael, B.D.; Easton, A.; Kneen, R.; Defres, S.; Sejvar, J.; Solomon, T. Neurological associations of COVID-19. Lancet Neurol. 2020, 19, 767–783. [Google Scholar] [CrossRef] [PubMed]
- Kanberg, N.; Ashton, N.J.; Andersson, L.-M.; Yilmaz, A.; Lindh, M.; Nilsson, S.; Price, R.W.; Blennow, K.; Zetterberg, H.; Gisslén, M. Neurochemical evidence of astrocytic and neuronal injury commonly found in COVID-19. Neurology 2020, 95, e1754–e1759. [Google Scholar] [CrossRef] [PubMed]
- Sutter, R.; Hert, L.; De Marchis, G.M.; Twerenbold, R.; Kappos, L.; Naegelin, Y.; Kuster, G.M.; Benkert, P.; Jost, J.; Msc, A.M.M.; et al. Serum Neurofilament Light Chain Levels in the Intensive Care Unit: Comparison between Severely Ill Patients with and without Coronavirus Disease 2019. Ann. Neurol. 2020, 89, 610–616. [Google Scholar] [CrossRef] [PubMed]
- Aamodt, A.H.; Høgestøl, E.A.; Popperud, T.H.; Holter, J.C.; Dyrhol-Riise, A.M.; Tonby, K. Blood neurofilament light concentration at admittance: A potential prognostic marker in COVID-19. J. Neurol. 2021, 268, 3574–3583. [Google Scholar] [CrossRef]
- De Lorenzo, R.; Loré, N.I.; Finardi, A.; Mandelli, A.; Cirillo, D.M.; Tresoldi, C.; Benedetti, F.; Ciceri, F.; Rovere-Querini, P.; Comi, G.; et al. Blood neurofilament light chain and total tau levels at admission predict death in COVID-19 patients. J Neurol. 2021, 268, 4436–4442. [Google Scholar] [CrossRef]
- Frontera, J.A.; Boutajangout, A.; Masurkar, A.; van Betensky, R.A.; Ge, Y.; Vedvyas, A. Comparison of serum neurodegenerative biomarkers among hospitalized COVID-19 patients versus non-COVID subjects with normal cognition, mild cognitive impair-ment, or Alzheimer’s dementia. Alzheimers Dement. 2022, 18, 899–910. [Google Scholar] [CrossRef]
- Patel, H.; Ashton, N.J.; Dobson, R.J.B.; Andersson, L.-M.; Yilmaz, A.; Blennow, K.; Gisslen, M.; Zetterberg, H. Proteomic blood profiling in mild, severe and critical COVID-19 patients. Sci. Rep. 2021, 11, 6357. [Google Scholar] [CrossRef]
- Sahin, B.; Celikbilek, A.; Kocak, Y.; Saltoglu, G.; Konar, N.; Hizmali, L. Plasma biomarkers of brain injury in COVID-19 patients with neurological symptoms. J. Neurol. Sci. 2022, 439, 120324. [Google Scholar] [CrossRef]
- Hanson, B.A.; Visvabharathy, L.; Ali, S.T.; Kang, A.K.; Patel, T.R.; Clark, J.R. Plasma Biomarkers of Neuropathogenesis in Hospitalized Patients With COVID-19 and Those with Postacute Sequelae of SARS-CoV-2 Infection. Neurol. (R) Neuroimmunol. Neuroinflamm. 2022, 9. [Google Scholar] [CrossRef]
- Verde, F.; Milone, I.; Bulgarelli, I.; Peverelli, S.; Colombrita, C.; Maranzano, A.; Calcagno, N.; Ticozzi, N.; Perego, G.B.; Parati, G.; et al. Serum neurofilament light chain levels in Covid-19 patients without major neurological manifestations. J. Neurol. 2022, 269, 5691–5701. [Google Scholar] [CrossRef]
- Geis, T.; Brandstetter, S.; Toncheva, A.A.; Laub, O.; Leipold, G.; Wagner, R.; Kabesch, M.; Kasser, S.; Kuhle, J.; Wellmann, S.; et al. Serum neurofilament light chain (sNfL) values in a large cross-sectional population of children with asymptomatic to moderate COVID-19. J. Neurol. 2021, 268, 3969–3974. [Google Scholar] [CrossRef] [PubMed]
- Valenzuela, R.; Pedrosa, M.A.; Garrido-Gil, P.; Labandeira, C.M.; Navarro, G.; Franco, R.; Rodriguez-Perez, A.I.; Labandeira-Garcia, J.L. Interactions between ibuprofen, ACE2, renin-angiotensin system, and spike protein in the lung. Implications for COVID-19. Clin. Transl. Med. 2021, 11, e371. [Google Scholar] [CrossRef] [PubMed]
- García-Ayllón, M.; Moreno-Pérez, O.; García-Arriaza, J.; Ramos-Rincón, J.; Cortés-Gómez, M.; Brinkmalm, G.; Andrés, M.; León-Ramírez, J.; Boix, V.; Gil, J.; et al. Plasma ACE2 species are differentially altered in COVID-19 patients. FASEB J. 2021, 35, e21745. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Ren, Z.; Ma, L.; Han, Y.; Wei, W.; Jiang, E. Progress in Research on SARS-CoV-2 Infection Causing Neurological Diseases and Its Infection Mechanism. Front. Neurol. 2020, 11, 592888. [Google Scholar] [CrossRef] [PubMed]
- Guadarrama-Ortiz, P.; Choreño-Parra, J.A.; Sánchez-Martínez, C.M.; Pacheco-Sánchez, F.J.; Rodríguez-Nava, A.I.; García-Quintero, G. Neurological Aspects of SARS-CoV-2 Infection: Mechanisms and Manifestations. Front Neurol. 2020, 11, 1039. [Google Scholar] [CrossRef]
- Han, Y.; Yuan, K.; Wang, Z.; Liu, W.-J.; Lu, Z.-A.; Liu, L.; Le Shi, L.; Yan, W.; Yuan, J.-L.; Li, J.-L.; et al. Neuropsychiatric manifestations of COVID-19, potential neurotropic mechanisms, and therapeutic interventions. Transl. Psychiatry 2021, 11, 1–20. [Google Scholar] [CrossRef]
- Song, E.; Zhang, C.; Israelow, B.; Lu-Culligan, A.; Prado, A.V.; Skriabine, S. Neuroinvasion of SARS-CoV-2 in human and mouse brain. J. Exp. Med. 2021, 218, e20202135. [Google Scholar] [CrossRef]
- Serrano, G.E.; Walker, J.E.; Arce, R.; Glass, M.J.; Vargas, D.; Sue, L.I. Mapping of SARS-CoV-2 Brain Invasion and Histopathology in COVID-19 Disease. medRxiv 2021. [Google Scholar] [CrossRef]
- Pilotto, A.; Masciocchi, S.; Volonghi, I.; de Giuli, V.; Caprioli, F.; Mariotto, S. Severe Acute Respiratory Syndrome Coronavirus 2 (SARS-CoV-2) Encephalitis Is a Cytokine Release Syndrome: Evidences From Cerebrospinal Fluid Analyses. Clin. Infect Dis. 2021, 73, e3019-26. [Google Scholar] [CrossRef]
- McQuaid, C.; Brady, M.; Deane, R. SARS-CoV-2: Is there neuroinvasion? Fluids Barriers CNS 2021, 18, 32. [Google Scholar] [CrossRef]
- Needham, E.J.; Ren, A.L.; Digby, R.J.; Norton, E.J.; Ebrahimi, S.; Outtrim, J.G. Brain injury in COVID-19 is associated with dysreg-ulated innate and adaptive immune responses. Brain 2022, 145, 4097–4107. [Google Scholar] [CrossRef] [PubMed]
- Frithiof, R.; Rostami, E.; Kumlien, E.; Virhammar, J.; Fällmar, D.; Hultström, M. Critical illness polyneuropathy, myopathy and neuronal biomarkers in COVID-19 patients: A prospective study. Clin. Neurophysiol. 2021, 132, 1733–1740. [Google Scholar] [CrossRef]
- Passos, F.R.S.; Heimfarth, L.; Monteiro, B.S.; Corrêa, C.B.; de Moura, T.R.; Araújo, A.A.D.S.; Martins-Filho, P.R.; Quintans-Júnior, L.J.; Quintans, J.D.S.S. Oxidative stress and inflammatory markers in patients with COVID-19: Potential role of RAGE, HMGB1, GFAP and COX-2 in disease severity. Int. Immunopharmacol. 2022, 104, 108502. [Google Scholar] [CrossRef]
- Masvekar, R.R.; Kosa, P.; Jin, K.; Dobbs, K.; Stack, M.A.; Castagnoli, R.; Quaresima, V.; Su, H.C.; Imberti, L.; Notarangelo, L.D.; et al. Prognostic value of serum/plasma neurofilament light chain for COVID -19-associated mortality. Ann. Clin. Transl. Neurol. 2022, 9, 622–632. [Google Scholar] [CrossRef] [PubMed]
- Hay, M.; Ryan, L.; Huentelman, M.; Konhilas, J.; Hoyer-Kimura, C.; Beach, T.G.; Serrano, G.E.; Reiman, E.M.; Blennow, K.; Parthasarathy, S. Serum Neurofilament Light is elevated in COVID-19 Positive Adults in the ICU and is associated with Co-Morbid Cardiovascular Disease, Neurological Complications, and Acuity of Illness. Cardiol. Cardiovasc. Med. 2021, 5, 551–565. [Google Scholar] [CrossRef]
- Prudencio, M.; Erben, Y.; Marquez, C.P.; Jansen-West, K.R.; Franco-Mesa, C.; Heckman, M.G.; White, L.J.; Dunmore, J.A.; Cook, C.N.; Lilley, M.T.; et al. Serum neurofilament light protein correlates with unfavorable clinical outcomes in hospitalized patients with COVID-19. Sci. Transl. Med. 2021, 13, 7643. [Google Scholar] [CrossRef] [PubMed]
- Paterson, R.W.; Benjamin, L.A.; Mehta, P.R.; Brown, R.L.; Athauda, D.; Ashton, N.J. Serum and cerebrospinal fluid biomarker profiles in acute SARS-CoV-2-associated neurological syndromes. Brain Commun. 2021, 3, fcab099. [Google Scholar] [CrossRef] [PubMed]
- Mehta, P.; McAuley, D.F.; Brown, M.; Sanchez, E.; Tattersall, R.S.; Manson, J.J. COVID-19: Consider cytokine storm syndromes and immunosuppression. Lancet 2020, 395, 1033–1034. [Google Scholar] [CrossRef] [PubMed]
- Tsagkaris, C.; Bilal, M.; Aktar, I.; Aboufandi, Y.; Tas, A.; Aborode, A.T.; Suvvari, T.K.; Ahmad, S.; Shkodina, A.; Phadke, R.; et al. Cytokine storm and neuropathological alterations in patients with neurological manifestations of COVID-19. Curr. Alzheimer Res. 2022, 19, 641–657. [Google Scholar] [CrossRef]
- Guasp, M.; Muñoz-Sánchez, G.; Martínez-Hernández, E.; Santana, D.; Carbayo, A.; Naranjo, L.; Bolós, U.; Framil, M.; Saiz, A.; Balasa, M.; et al. CSF Biomarkers in COVID-19 Associated Encephalopathy and Encephalitis Predict Long-Term Outcome. Front. Immunol. 2022, 13, 1600. [Google Scholar] [CrossRef]
- Kanberg, N.; Simrén, J.; Edén, A.; Andersson, L.-M.; Nilsson, S.; Ashton, N.J.; Sundvall, P.-D.; Nellgård, B.; Blennow, K.; Zetterberg, H.; et al. Neurochemical signs of astrocytic and neuronal injury in acute COVID-19 normalizes during long-term follow-up. Ebiomedicine 2021, 70, 103512. [Google Scholar] [CrossRef] [PubMed]
- Lennol, M.P.; García-Ayllón, M.-S.; Esteban, M.; García-Arriaza, J.; Sáez-Valero, J. Serum angiotensin-converting enzyme 2 as a potential biomarker for SARS-CoV-2 infection and vaccine efficacy. Front Immunol. 2022, 13, 1001951. [Google Scholar] [CrossRef] [PubMed]
- Qiao, J.; Li, W.; Bao, J.; Peng, Q.; Wen, D.; Wang, J.; Sun, B. The expression of SARS-CoV-2 receptor ACE2 and CD147, and protease TMPRSS2 in human and mouse brain cells and mouse brain tissues. Biochem. Biophys. Res. Commun. 2020, 533, 867–871. [Google Scholar] [CrossRef]
- Dong, M.; Zhang, J.; Ma, X.; Tan, J.; Chen, L.; Liu, S.; Xin, Y.; Zhuang, L. ACE2, TMPRSS2 distribution and extrapulmonary organ injury in patients with COVID-19. Biomed. Pharmacother. 2020, 131, 110678. [Google Scholar] [CrossRef] [PubMed]
- Li, T.; Huang, H.-Y.; Wang, H.-D.; Gao, C.-C.; Liang, H.; Deng, C.-L.; Zhao, X.; Han, Y.-L.; Zhou, M.-L. Restoration of Brain Angiotensin-Converting Enzyme 2 Alleviates Neurological Deficits after Severe Traumatic Brain Injury via Mitigation of Pyroptosis and Apoptosis. J. Neurotrauma 2022, 39, 423–434. [Google Scholar] [CrossRef] [PubMed]
- Ramani, A.; Müller, L.; Ostermann, P.N.; Gabriel, E.; Abida-Islam, P.; Müller-Schiffmann, A. SARS-CoV-2 targets neurons of 3D human brain organoids. EMBO J. 2020, 39, e106230. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| COVID-19 Patients Characteristics | |
|---|---|
| Demographic and clinical data | |
| Presentation (moderate/severe) | 35/10 |
| Hospitalization (days) | 10.5 |
| Diabetes | 6.2 (%) |
| Hypertension | 32.3 (%) |
| Obesity | 34.2 (%) |
| Neurological symptoms | |
| Headache | 26.9 (%) |
| Memory loss | 23.1 (%) |
| Asthenia | 50.0 (%) |
| Myalgia/arthralgia | 26.9 (%) |
| Anosmia/ageusia | 26.9 (%) |
| Vision impairment | 3.8 (%) |
| Hemaetological parameters | |
| Lymphocytes (per mm3) | 1130 [790–1410] |
| Troponin T (ng/L) | 12 [5–21] |
| CRP (mg/dL) | 5.0 [2.4–7.9] |
| BNP (pg/mL) | 109 [37–475] |
| LDH (U/L) | 255 [201–304] |
| D-dimer (mg/mL) | 0.75 [0.40–1.23] |
| IL-6 (pg/mL) | 28 [19–59] |
| Ferritin (mg/L) | 720 [423–1104] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lennol, M.P.; Ashton, N.J.; Moreno-Pérez, O.; García-Ayllón, M.-S.; Ramos-Rincon, J.-M.; Andrés, M.; León-Ramírez, J.-M.; Boix, V.; Gil, J.; Blennow, K.; et al. Transient Changes in the Plasma of Astrocytic and Neuronal Injury Biomarkers in COVID-19 Patients without Neurological Syndromes. Int. J. Mol. Sci. 2023, 24, 2715. https://doi.org/10.3390/ijms24032715
Lennol MP, Ashton NJ, Moreno-Pérez O, García-Ayllón M-S, Ramos-Rincon J-M, Andrés M, León-Ramírez J-M, Boix V, Gil J, Blennow K, et al. Transient Changes in the Plasma of Astrocytic and Neuronal Injury Biomarkers in COVID-19 Patients without Neurological Syndromes. International Journal of Molecular Sciences. 2023; 24(3):2715. https://doi.org/10.3390/ijms24032715
Chicago/Turabian StyleLennol, Matthew P., Nicholas J. Ashton, Oscar Moreno-Pérez, María-Salud García-Ayllón, Jose-Manuel Ramos-Rincon, Mariano Andrés, José-Manuel León-Ramírez, Vicente Boix, Joan Gil, Kaj Blennow, and et al. 2023. "Transient Changes in the Plasma of Astrocytic and Neuronal Injury Biomarkers in COVID-19 Patients without Neurological Syndromes" International Journal of Molecular Sciences 24, no. 3: 2715. https://doi.org/10.3390/ijms24032715