
Phytochemicals as Immunomodulatory Agents in Melanoma
 ,
,  ,
,  , , , ,
, , , ,  ,
,  ,
,
Abstract
:1. Introduction
2. Immunomodulation and Immunotherapy of Melanoma
2.1. Melanoma and Interaction with Tumor Microenvironment
2.2. Immune Checkpoints
2.3. Cytokines
3. Classification and Extraction Methods of Plant Secondary Metabolites
4. Alkaloids
4.1. Methylxanthines
4.2. Other Alkaloids
5. Phenolic Compounds
5.1. Flavonoids
5.2. Phenolic Acids
5.3. Stilbenes
5.4. Other Phenolic Compounds
6. Terpenes and Terpenoids
7. Sulfur-Containing Compounds
8. Discussion and Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Tímár, J.; Ladányi, A. Molecular Pathology of Skin Melanoma: Epidemiology, Differential Diagnostics, Prognosis and Therapy Prediction. Int. J. Mol. Sci. 2022, 23, 5384. [Google Scholar] [CrossRef]
- Zhong, J.; Yan, W.; Wang, C.; Liu, W.; Lin, X.; Zou, Z.; Sun, W.; Chen, Y. BRAF Inhibitor Resistance in Melanoma: Mechanisms and Alternative Therapeutic Strategies. Curr. Treat. Options Oncol. 2022, 23, 1503–1521. [Google Scholar] [CrossRef]
- Pitcovski, J.; Shahar, E.; Aizenshtein, E.; Gorodetsky, R. Melanoma antigens and related immunological markers. Crit. Rev. Oncol. Hematol. 2017, 115, 36–49. [Google Scholar] [CrossRef] [PubMed]
- Lazaroff, J.; Bolotin, D. Targeted Therapy and Immunotherapy in Melanoma. Dermatol. Clin. 2023, 41, 65–77. [Google Scholar] [CrossRef] [PubMed]
- Lázár-Molnár, E.; Hegyesi, H.; Tóth, S.; Falus, A. Autocrine and paracrine regulation by cytokines and growth factors in melanoma. Cytokine 2000, 12, 547–554. [Google Scholar] [CrossRef]
- Cuevas, A.; Saavedra, N.; Salazar, L.A.; Abdalla, D.S. Modulation of immune function by polyphenols: Possible contribution of epigenetic factors. Nutrients 2013, 5, 2314–2332. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chang, Z.; Wang, Y.; Liu, C.; Smith, W.; Kong, L. Natural Products for Regulating Macrophages M2 Polarization. Curr. Stem Cell Res. Ther. 2020, 5, 559–569. [Google Scholar] [CrossRef]
- Renda, G.; Gökkaya, İ.; Şöhretoğlu, D. Immunomodulatory properties of triterpenes. Phytochem. Rev. 2022, 21, 537–563. [Google Scholar] [CrossRef]
- Sun, K.; Wu, L.; Wang, S.; Deng, W. Antitumor effects of Chinese herbal medicine compounds and their nano-formulations on regulating the immune system microenvironment. Front. Oncol. 2022, 12, 949332. [Google Scholar] [CrossRef]
- Amalinei, C.; Grigoraș, A.; Lozneanu, L.; Căruntu, I.-D.; Giușcă, S.-E.; Balan, R.A. The Interplay between Tumour Microenvironment Components in Malignant Melanoma. Medicina 2022, 58, 365. [Google Scholar] [CrossRef]
- Pardoll, D.M. The blockade of immune checkpoints in cancer immunotherapy. Nat. Rev. Cancer 2012, 12, 252–264. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karin, M. Nuclear factor-kappaB in cancer development and progression. Nature 2006, 441, 431–436. [Google Scholar] [CrossRef]
- Dantonio, P.M.; Klein, M.O.; Freire, M.R.V.B.; Araujo, C.N.; Chiacetti, A.C.; Correa, R.G. Exploring major signaling cascades in melanomagenesis: A rationale route for targetted skin cancer therapy. Biosci. Rep. 2018, 38, BSR20180511. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- D’Aguanno, S.; Mallone, F.; Marenco, M.; Del Bufalo, D.; Moramarco, A. Hypoxia-dependent drivers of melanoma progression. J. Exp. Clin. Cancer Res. 2021, 40, 159. [Google Scholar] [CrossRef] [PubMed]
- Koury, J.; Lucero, M.; Cato, C.; Chang, L.; Geiger, J.; Henry, D.; Hernandez, J.; Hung, F.; Kaur, P.; Teskey, G.; et al. Immunotherapies: Exploiting the immune system for cancer treatment. J. Immunol. Res. 2018, 2018, 9585614. [Google Scholar] [CrossRef]
- Lin, J.Y.; Fisher, D.E. Melanocyte Biology and Skin Pigmentation. Nature 2007, 445, 843–850. [Google Scholar] [CrossRef]
- D’Ischia, M.; Wakamatsu, K.; Napolitano, A.; Briganti, S.; Garcia-Borron, J.C.; Kovacs, D.; Meredith, P.; Pezzella, A.; Picardo, M.; Sarna, T.; et al. Melanins and Melanogenesis: Methods, Standards, Protocols. Pigment Cell Melanoma Res. 2013, 26, 616–633. [Google Scholar] [CrossRef]
- Del Bino, S.; Duval, C.; Bernerd, F. Clinical and Biological Characterization of Skin Pigmentation Diversity and Its Consequences on UV Impact. Int. J. Mol. Sci. 2018, 19, 2668. [Google Scholar] [CrossRef] [Green Version]
- Slominski, R.M.; Sarna, T.; Płonka, P.M.; Raman, C.; Brożyna, A.A.; Slominski, A.T. Melanoma, Melanin, and Melanogenesis: The Yin and Yang Relationship. Front. Oncol. 2022, 12, 842496. [Google Scholar] [CrossRef]
- Sarna, M.; Krzykawska-Serda, M.; Jakubowska, M.; Zadlo, A.; Urbanska, K. Melanin Presence Inhibits Melanoma Cell Spread in Mice in a Unique Mechanical Fashion. Sci. Rep. 2019, 9, 9280. [Google Scholar] [CrossRef]
- Cabaço, L.C.; Tomás, A.; Pojo, M.; Barral, D.C. The Dark Side of Melanin Secretion in Cutaneous Melanoma Aggressiveness. Front. Oncol. 2022, 12, 887366. [Google Scholar] [CrossRef] [PubMed]
- Wakamatsu, K.; Ito, A.; Tamura, Y.; Hida, T.; Kamiya, T.; Torigoe, T.; Honda, H.; Ito, S.; Jimbow, K. Molecular Events in the Melanogenesis Cascade as Novel Melanoma-Targeted Small Molecules: Principle and Development. Cancers 2022, 14, 5588. [Google Scholar] [CrossRef] [PubMed]
- Burkhart, C.G.; Burkhart, C.N. The mole theory: Primary function of melanocytes and melanin may be antimicrobial defense and immunomodulation (not solar protection). Int. J. Dermatol. 2005, 44, 340–342. [Google Scholar] [CrossRef] [PubMed]
- Koike, S.; Yamasaki, K. Melanogenesis Connection with Innate Immunity and Toll-Like Receptors. Int. J. Mol. Sci. 2020, 21, 9769. [Google Scholar] [CrossRef] [PubMed]
- Netanely, D.; Leibou, S.; Parikh, R.; Stern, N.; Vaknine, H.; Brenner, R.; Amar, S.; Factor, R.H.; Perluk, T.; Frand, J.; et al. Classification of node-positive melanomas into prognostic subgroups using keratin, immune, and melanogenesis expression patterns. Oncogene 2021, 40, 1792–1805. [Google Scholar] [CrossRef]
- Arciuli, M.; Fiocco, D.; Cicero, R.; Maida, I.; Zanna, P.T.; Guida, G.; Horsberg, T.E.; Koppang, E.O.; Gallone, A. Melanogenesis in visceral tissues of Salmo salar. A link between immunity and pigment production? Biochem. Cell Biol. 2012, 90, 769–778. [Google Scholar] [CrossRef]
- Nappi, A.J.; Christensen, B.M. Melanogenesis and associated cytotoxic reactions: Applications to insect innate immunity. Insect Biochem. Mol. Biol. 2005, 35, 443–459. [Google Scholar] [CrossRef]
- Manna, S.K.; Aggarwal, B.B. Alpha-Melanocyte-Stimulating Hormone Inhibits the Nuclear Transcription Factor NF-Kappa B Activation Induced by Various Inflammatory Agents. J. Immunol. 1998, 161, 2873–2880. [Google Scholar] [CrossRef]
- Pawlikowska, M.; Jędrzejewski, T.; Slominski, A.T.; Brożyna, A.A.; Wrotek, S. Pigmentation Levels Affect Melanoma Responses to Coriolus Versicolor Extract and Play a Crucial Role in Melanoma-Mononuclear Cell Crosstalk. Int. J. Mol. Sci. 2021, 22, 5735. [Google Scholar] [CrossRef]
- Buchbinder, E.I.; Desai, A. CTLA-4 and PD-1 Pathways: Similarities, Differences, and Implications of Their Inhibition. Am. J. Clin. Oncol. 2016, 39, 98–106. [Google Scholar] [CrossRef]
- Hodi, F.S.; O’Day, S.J.; McDermott, D.F.; Weber, R.W.; Sosman, J.A.; Haanen, J.B.; Gonzalez, R.; Robert, C.; Schadendorf, D.; Hassel, J.C.; et al. Improved survival with ipilimumab in patients with metastatic melanoma. N. Engl. J. Med. 2010, 363, 711–723. [Google Scholar] [CrossRef] [PubMed]
- Robert, C.; Ribas, A.; Schachter, J.; Arance, A.; Grob, J.J.; Mortier, L.; Daud, A.; Carlino, M.S.; McNeil, C.M.; Lotem, M.; et al. Pembrolizumab versus ipilimumab in advanced melanoma (KEYNOTE-006): Post-hoc 5-year results from an open-label, multicentre, randomised, controlled, phase 3 study. Lancet Oncol. 2019, 20, 1239–1251. [Google Scholar] [CrossRef] [PubMed]
- Michielin, O.; Atkins, M.B.; Koon, H.B.; Dummer, R.; Ascierto, P.A. Evolving impact of long-term survival results on metastatic melanoma treatment. J. Immunother. Cancer 2020, 8, e000948. [Google Scholar] [CrossRef] [PubMed]
- Larkin, J.; Chiarion-Sileni, V.; Gonzalez, R.; Grob, J.J.; Cowey, C.L.; Lao, C.D.; Schadendorf, D.; Dummer, R.; Smylie, M.; Rutkowski, P.; et al. Combined Nivolumab and Ipilimumab or Monotherapy in Untreated Melanoma. N. Engl. J. Med. 2015, 373, 23–34. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Larkin, J.; Chiarion-Sileni, V.; Gonzalez, R.; Grob, J.J.; Rutkowski, P.; Lao, C.D.; Cowey, C.L.; Schadendorf, D.; Wagstaff, J.; Dummer, R.; et al. Five-year survival with combined nivolumab and ipilimumab in advanced melanoma. N. Engl. J. Med. 2019, 381, 1535–1546. [Google Scholar] [CrossRef] [Green Version]
- Wolchok, J.D.; Chiarion-Sileni, V.; Gonzalez, R.; Grob, J.J.; Rutkowski, P.; Lao, C.D.; Cowey, C.L.; Schadendorf, D.; Wagstaff, J.; Dummer, R.; et al. Long-Term Outcomes With Nivolumab Plus Ipilimumab or Nivolumab Alone Versus Ipilimumab in Patients With Advanced Melanoma. J. Clin. Oncol. 2022, 40, 127–137. [Google Scholar] [CrossRef] [PubMed]
- Tawbi, H.A.; Schadendorf, D.; Lipson, E.J.; Ascierto, P.A.; Matamala, L.; Castillo Gutiérrez, E.; Rutkowski, P.; Gogas, H.J.; Lao, C.D.; De Menezes, J.J.; et al. Relatlimab and nivolumab versus nivolumab in untreated advanced melanoma. N. Engl. J. Med. 2022, 386, 24–34. [Google Scholar] [CrossRef] [PubMed]
- Giampietri, C.; Scatozza, F.; Crecca, E.; Vigiano Benedetti, V.; Natali, P.G.; Facchiano, A. Analysis of gene expression levels and their impact on survival in 31 cancer-types patients identifies novel prognostic markers and suggests unexplored immunotherapy treatment options in a wide range of malignancies. J. Transl. Med. 2022, 20, 467. [Google Scholar] [CrossRef]
- Schadendorf, D.; van Akkooi, A.C.J.; Berking, C.; Griewank, K.G.; Gutzmer, R.; Hauschild, A.; Stang, A.; Roesch, A.; Ugurel, S. Melanoma. Lancet 2018, 392, 971–984. [Google Scholar] [CrossRef]
- Andtbacka, R.H.; Kaufman, H.L.; Collichio, F.; Amatruda, T.; Senzer, N.; Chesney, J.; Delman, K.A.; Spitler, L.E.; Puzanov, I.; Agarwala, S.S.; et al. Talimogene Laherparepvec Improves Durable Response Rate in Patients With Advanced Melanoma. J. Clin. Oncol. 2015, 33, 2780–2788. [Google Scholar] [CrossRef]
- Andtbacka, R.H.I.; Collichio, F.; Harrington, K.J.; Middleton, M.R.; Downey, G.; Öhrling, K.; Kaufman, H.L. Final analyses of OPTiM: A randomized phase III trial of talimogene laherparepvec versus granulocyte-macrophage colony-stimulating factor in unresectable stage III-IV melanoma. J. Immunother. Cancer 2019, 7, 145. [Google Scholar] [CrossRef] [Green Version]
- Sumimoto, H.; Imabayashi, F.; Iwata, T.; Kawakami, Y. The BRAF-MAPK signaling pathway is essential for cancer-immune evasion in human melanoma cells. J. Exp. Med. 2006, 203, 1651–1656. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Di Martile, M.; Farini, V.; Consonni, F.M.; Trisciuoglio, D.; Desideri, M.; Valentini, E.; D’Aguanno, S.; Tupone, M.G.; Buglioni, S.; Ercolani, C.; et al. Melanoma-specific bcl-2 promotes a protumoral M2-like phenotype by tumor-associated macrophages. J. Immunother. Cancer 2020, 8, e000489. [Google Scholar] [CrossRef] [PubMed]
- Lucianò, A.M.; Pérez-Oliva, A.B.; Mulero, V.; Del Bufalo, D. Bcl-xL: A Focus on Melanoma Pathobiology. Int. J. Mol. Sci. 2021, 22, 2777. [Google Scholar] [CrossRef]
- Barceló, C.; Sisó, P.; de la Rosa, I.; Megino-Luque, C.; Navaridas, R.; Maiques, O.; Urdanibia, I.; Eritja, N.; Soria, X.; Potrony, M.; et al. M-CSF as a therapeutic target in BRAFV600E melanoma resistant to BRAF inhibitors. Br. J. Cancer 2022, 127, 1142–1152. [Google Scholar] [CrossRef]
- Pedersen, J.G.; Madsen, A.T.; Gammelgaard, K.R.; Aggerholm-Pedersen, N.; Sørensen, B.S.; Øllegaard, T.H.; Jakobsen, M.R. Inflammatory Cytokines and ctDNA Are Biomarkers for Progression in Advanced-Stage Melanoma Patients Receiving Checkpoint Inhibitors. Cancers 2020, 12, 1414. [Google Scholar] [CrossRef] [PubMed]
- Tsukamoto, H.; Fujieda, K.; Miyashita, A.; Fukushima, S.; Ikeda, T.; Kubo, Y.; Senju, S.; Ihn, H.; Nishimura, Y.; Oshiumi, H. Combined Blockade of IL6 and PD-1/PD-L1 Signaling Abrogates Mutual Regulation of Their Immunosuppressive Effects in the Tumor Microenvironment. Cancer Res. 2018, 78, 5011–5022. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vanpouille-Box, C.; Formenti, S.C. Dual Transforming Growth Factor-β and Programmed Death-1 Blockade: A Strategy for Immune-Excluded Tumors? Trends Immunol. 2018, 39, 435–437. [Google Scholar] [CrossRef] [PubMed]
- Mancuso, F.; Lage, S.; Rasero, J.; Díaz-Ramón, J.L.; Apraiz, A.; Pérez-Yarza, G.; Ezkurra, P.A.; Penas, C.; Sánchez-Diez, A.; García-Vazquez, M.D.; et al. Serum markers improve current prediction of metastasis development in early-stage melanoma patients: A machine learning-based study. Mol. Oncol. 2020, 14, 1705–1718. [Google Scholar] [CrossRef]
- Atkins, M.B.; Kunkel, L.; Sznol, M.; Rosenberg, S.A. High-dose recombinant interleukin-2 therapy in patients with metastatic melanoma: Long-term survival update. Cancer J. Sci. Am. 2000, 6, S11–S14. [Google Scholar]
- Dudley, M.E.; Wunderlich, J.R.; Yang, J.C.; Sherry, R.M.; Topalian, S.L.; Restifo, N.P.; Royal, R.E.; Kammula, U.; White, D.E.; Mavroukakis, S.A.; et al. Adoptive cell transfer therapy following non-myeloablative but lymphodepleting chemotherapy for the treatment of patients with refractory metastatic melanoma. J. Clin. Oncol. 2005, 23, 2346–2357. [Google Scholar] [CrossRef] [PubMed]
- Rosenberg, S.A.; Yang, J.C.; Sherry, R.M.; Kammula, U.S.; Hughes, M.S.; Phan, G.Q.; Citrin, D.E.; Restifo, N.P.; Robbins, P.F.; Wunderlich, J.R.; et al. Durable complete responses in heavily pretreated patients with metastatic melanoma using T-cell transfer immunotherapy. Clin. Cancer Res. 2011, 17, 4550–4557. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dudley, M.E.; Gross, C.A.; Somerville, R.P.; Hong, Y.; Schaub, N.P.; Rosati, S.F.; White, D.E.; Nathan, D.; Restifo, N.P.; Steinberg, S.M.; et al. Randomized selection design trial evaluating CD8+-enriched versus unselected tumor-infiltrating lymphocytes for adoptive cell therapy for patients with melanoma. J. Clin. Oncol. 2013, 31, 2152–2159. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dafni, U.; Michielin, O.; Lluesma, S.M.; Tsourti, Z.; Polydoropoulou, V.; Karlis, D.; Besser, M.J.; Haanen, J.; Svane, I.M.; Ohashi, P.S.; et al. Efficacy of adoptive therapy with tumor-infiltrating lymphocytes and recombinant interleukin-2 in advanced cutaneous melanoma: A systematic review and meta-analysis. Ann. Oncol. 2019, 30, 1902–1913. [Google Scholar] [CrossRef] [Green Version]
- Payne, R.; Glenn, L.; Hoen, H.; Richards, B.; Smith, J.W.; Lufkin, R.; Crocenzi, T.S.; Urba, W.J.; Curti, B.D. Durable responses and reversible toxicity of high-dose interleukin-2 treatment of melanoma and renal cancer in a community hospital biotherapy program. J. Immunother. Cancer 2014, 2, 13. [Google Scholar] [CrossRef] [Green Version]
- Bentebibel, S.-E.; Hurwitz, M.E.; Bernatchez, C.; Haymaker, C.; Hudgens, C.W.; Kluger, H.M.; Tetzlaff, M.T.; Tagliaferri, M.A.; Zalevsky, J.; Hoch, U.; et al. A First-in-Human Study and Biomarker Analysis of NKTR-214, a Novel IL2Rβγ-Biased Cytokine, in Patients with Advanced or Metastatic Solid Tumors. Cancer Discov. 2019, 9, 711–721. [Google Scholar] [CrossRef] [Green Version]
- Diab, A.; Tykodi, S.S.; Daniels, G.A.; Maio, M.; Curti, B.D.; Lewis, K.D.; Jang, S.; Kalinka, E.; Puzanov, I.; Spira, A.I.; et al. Bempegaldesleukin Plus Nivolumab in First-Line Metastatic Melanoma. J. Clin. Oncol. 2021, 39, 2914–2925. [Google Scholar] [CrossRef]
- Eggermont, A.M.; Ascierto, P.A.; Khushalani, N.I.; Schadendorf, D.; Boland, G.; Weber, J.; Lewis, K.D.; Johnson, D.; Rivalland, G.; Khattak, A.; et al. PIVOT-12: A phase III study of adjuvant bempegaldesleukin plus nivolumab in resected stage III/IV melanoma at high risk for recurrence. Future Oncol. 2022, 18, 903–913. [Google Scholar] [CrossRef]
- Naing, A.; Wong, D.J.; Infante, J.R.; Korn, W.M.; Aljumaily, R.; Papadopoulos, K.P.; Autio, K.A.; Pant, S.; Bauer, T.M.; Drakaki, A.; et al. Pegilodecakin combined with pembrolizumab or nivolumab for patients with advanced solid tumours (IVY): A multicentre, multicohort, open-label, phase 1b trial. Lancet Oncol. 2019, 20, 1544–1555. [Google Scholar] [CrossRef]
- Mortarini, R.; Borri, A.; Tragni, G.; Bersani, I.; Vegetti, C.; Bajetta, E.; Pilotti, S.; Cerundolo, V.; Anichini, A. Peripheral burst of tumor-specific cytotoxic T lymphocytes and infiltration of metastatic lesions by memory CD8+ T cells in melanoma patients receiving interleukin 12. Cancer Res. 2000, 60, 3559–3568. [Google Scholar]
- Rosenberg, S.A.; Sportès, C.; Ahmadzadeh, M.; Fry, T.J.; Ngo, L.T.; Schwarz, S.L.; Stetler-Stevenson, M.; Morton, K.E.; Mavroukakis, S.A.; Morre, M.; et al. IL-7 administration to humans leads to expansion of CD8+ and CD4+ cells but a relative decrease of CD4+ T-regulatory cells. J. Immunother. 2006, 29, 313–319. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sportès, C.; Babb, R.R.; Krumlauf, M.C.; Hakim, F.T.; Steinberg, S.M.; Chow, C.K.; Brown, M.R.; Fleisher, T.A.; Noel, P.; Maric, I.; et al. Phase I study of recombinant human interleukin-7 administration in subjects with refractory malignancy. Clin. Cancer Res. 2010, 16, 727–735. [Google Scholar] [CrossRef] [PubMed]
- Miller, J.S.; Morishima, C.; McNeel, D.G.; Patel, M.R.; Kohrt, H.E.K.; Thompson, J.A.; Sondel, P.M.; Wakelee, H.A.; Disis, M.L.; Kaiser, J.C.; et al. First-in-Human Phase I Study of Subcutaneous Outpatient Recombinant Human IL15 (rhIL15) in Adults with Advanced Solid Tumors. Clin. Cancer Res. 2018, 24, 1525–1535. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Margolin, K.; Morishima, C.; Velcheti, V.; Miller, J.S.; Lee, S.M.; Silk, A.W.; Holtan, S.G.; Lacroix, A.M.; Fling, S.P.; Kaiser, J.C.; et al. Phase I Trial of ALT-803, A Novel Recombinant IL15 Complex, in Patients with Advanced Solid Tumors. Clin. Cancer Res. 2018, 24, 5552–5561. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Davis, I.D.; Brady, B.; Kefford, R.F.; Millward, M.; Cebon, J.; Skrumsager, B.K.; Mouritzen, U.; Hansen, L.T.; Skak, K.; Lundsgaard, D.; et al. Clinical and biological efficacy of recombinant human interleukin-21 in patients with stage IV malignant melanoma without prior treatment: A phase IIa trial. Clin. Cancer Res. 2009, 15, 2123–2129. [Google Scholar] [CrossRef] [Green Version]
- Lawson, D.H.; Lee, S.; Zhao, F.; Tarhini, A.A.; Margolin, K.A.; Ernstoff, M.S.; Atkins, M.B.; Cohen, G.I.; Whiteside, T.L.; Butterfield, L.H.; et al. Randomized, Placebo-Controlled, Phase III Trial of Yeast-Derived Granulocyte-Macrophage Colony-Stimulating Factor (GM-CSF) Versus Peptide Vaccination Versus GM-CSF Plus Peptide Vaccination Versus Placebo in Patients With No Evidence of Disease After Complete Surgical Resection of Locally Advanced and/or Stage IV Melanoma: A Trial of the Eastern Cooperative Oncology Group-American College of Radiology Imaging Network Cancer Research Group (E4697). J. Clin. Oncol. 2015, 33, 4066–4076. [Google Scholar] [CrossRef]
- Kaneko, S.; Mastaglio, S.; Bondanza, A.; Ponzoni, M.; Sanvito, F.; Aldrighetti, L.; Radrizzani, M.; La Seta-Catamancio, S.; Provasi, E.; Mondino, A.; et al. IL-7 and IL-15 allow the generation of suicide gene-modified alloreactive self-renewing central memory human T lymphocytes. Blood 2009, 113, 1006–1015. [Google Scholar] [CrossRef] [Green Version]
- Kirkwood, J.M.; Strawderman, M.H.; Ernstoff, M.S.; Smith, T.J.; Borden, E.C.; Blum, R.H. Interferon alfa-2b adjuvant therapy of high-risk resected cutaneous melanoma: The Eastern Cooperative Oncology Group Trial EST 1684. J. Clin. Oncol. 1996, 14, 7–17. [Google Scholar] [CrossRef]
- Eggermont, A.M.; Suciu, S.; Testori, A.; Santinami, M.; Kruit, W.H.; Marsden, J.; Punt, C.J.; Salès, F.; Dummer, R.; Robert, C.; et al. Long-term results of the randomized phase III trial EORTC 18991 of adjuvant therapy with pegylated interferon alfa-2b versus observation in resected stage III melanoma. J. Clin. Oncol. 2012, 30, 3810–3818. [Google Scholar] [CrossRef]
- Ascierto, P.A.; Chiarion-Sileni, V.; Muggiano, A.; Mandalà, M.; Pimpinelli, N.; Del Vecchio, M.; Rinaldi, G.; Simeone, E.; Queirolo, P. Interferon alpha for the adjuvant treatment of melanoma: Review of international literature and practical recommendations from an expert panel on the use of interferon. J. Chemother. 2014, 26, 193–201. [Google Scholar] [CrossRef]
- Lens, M.B.; Dawes, M. Interferon alfa therapy for malignant melanoma: A systematic review of randomized controlled trials. J. Clin. Oncol. 2002, 20, 1818–1825. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Unger, J.M.; Darke, A.; Othus, M.; Truong, T.G.; Khushalani, N.; Kendra, K.; Lewis, K.D.; Faller, B.; Funchain, P.; Buchbinder, E.I.; et al. Effectiveness of Adjuvant Pembrolizumab vs High-Dose Interferon or Ipilimumab for Quality-of-Life Outcomes in Patients With Resected Melanoma: A Secondary Analysis of the SWOG S1404 Randomized Clinical Trial. JAMA Oncol. 2022, e225486. [Google Scholar] [CrossRef] [PubMed]
- Davar, D.; Wang, H.; Chauvin, J.M.; Pagliano, O.; Fourcade, J.J.; Ka, M.; Menna, C.; Rose, A.; Sander, C.; Borhani, A.A.; et al. Phase Ib/II Study of Pembrolizumab and Pegylated-Interferon Alfa-2b in Advanced Melanoma. J. Clin. Oncol. 2018, 36, JCO1800632. [Google Scholar] [CrossRef] [PubMed]
- Davar, D.; Simonelli, M.; Gutierrez, M.; Calvo, E.; Melear, J.; Piha-Paul, S.; Richards, D.; Dallos, M.; Parameswaran, J.; Kumar, V.; et al. Interleukin-8–neutralizing monoclonal antibody BMS-986253 plus nivolumab (NIVO) in biomarker-enriched, primarily anti–PD-(L)1–experienced patients with advanced cancer: Initial phase 1 results. J. Immunother. Cancer 2020, 8, A239–A240. [Google Scholar] [CrossRef]
- Morris, J.C.; Tan, A.R.; Olencki, T.E.; Shapiro, G.I.; Dezube, B.J.; Reiss, M.; Hsu, F.J.; Berzofsky, J.A.; Lawrence, D.P. Phase I study of GC1008 (fresolimumab): A human anti-transforming growth factor-beta (TGFbeta) monoclonal antibody in patients with advanced malignant melanoma or renal cell carcinoma. PLoS ONE 2014, 9, e90353. [Google Scholar] [CrossRef]
- Montfort, A.; Filleron, T.; Virazels, M.; Dufau, C.; Milhès, J.; Pagès, C.; Olivier, P.; Ayyoub, M.; Mounier, M.; Lusque, A.; et al. Combining Nivolumab and Ipilimumab with Infliximab or Certolizumab in Patients with Advanced Melanoma: First Results of a Phase Ib Clinical Trial. Clin. Cancer Res. 2021, 27, 1037–1047. [Google Scholar] [CrossRef]
- Heinrich, M.; Mah, J.; Amirkia, V. Alkaloids Used as Medicines: Structural Phytochemistry Meets Biodiversity—An Update and Forward Look. Molecules 2021, 26, 1836. [Google Scholar] [CrossRef]
- Khadem, S.; Marles, R.J. Chromone and flavonoid alkaloids: Occurrence and bioactivity. Molecules 2011, 17, 191–206. [Google Scholar] [CrossRef] [Green Version]
- Yang, L.; Stöckigt, J. Trends for diverse production strategies of plant medicinal alkaloids. Nat. Prod. Rep. 2010, 27, 1469–1479. [Google Scholar] [CrossRef]
- Dey, P.; Kundu, A.; Kumar, A.; Gupta, M.; Lee, B.M.; Bhakta, T.; Dash, S.; Kim, H.S. Analysis of alkaloids (indole alkaloids, isoquinoline alkaloids, tropane alkaloids). In Recent Advances in Natural Products Analysis; Silva, A.S., Navabi, S.F., Saeedi, M., Navabi, S.M., Eds.; Elsevier: Amsterdam, The Netherlands, 2020; pp. 505–567. [Google Scholar] [CrossRef]
- Lu, J.-J.; Bao, J.-L.; Chen, X.-P.; Huang, M.; Wang, Y.-T. Alkaloids Isolated from Natural Herbs as the Anticancer Agents. Evid. Based Complement. Alternat. Med. 2012, 2012, 485042. [Google Scholar] [CrossRef] [Green Version]
- Bhambhani, S.; Kondhare, K.R.; Giri, A.P. Diversity in Chemical Structures and Biological Properties of Plant Alkaloids. Molecules 2021, 26, 3374. [Google Scholar] [CrossRef]
- Harborne, J.B.; Simmonds, N.W. Biochemistry of Phenolic Compounds; Academic Press: London, UK, 1964; 101p. [Google Scholar]
- Zhang, Y.; Cai, P.; Cheng, G.; Zhang, Y. A brief review of phenolic compounds identified from plants: Their extraction, analysis, and biological activity. Nat. Prod. Commun. 2022, 17, 1–14. [Google Scholar] [CrossRef]
- Matos, M.J.; Santana, L.; Uriarte, E.; Abreu, O.A.; Molina, E.; Yordi, E.G. Coumarins—An Important Class of Phytochemicals. In Phytochemicals-Isolation, Characterisation and Role in Human Health; In Tech: London, UK, 2015; pp. 113–140. [Google Scholar] [CrossRef] [Green Version]
- Frezza, C.; Venditti, A.; Toniolo, C.; De Vita, D.; Franceschin, M.; Ventrone, A.; Tomassini, L.; Foddai, S.; Guiso, M.; Nicoletti, M.; et al. Nor-Lignans: Occurrence in Plants and Biological Activities-A Review. Molecules 2020, 25, 197. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cushnie, T.P.; Lamb, A.J. Antimicrobial activity of flavonoids. Int. J. Antimicrob. Agents 2005, 26, 343–356. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Cao, H.; Huang, Q.; Xiao, J.; Teng, H. Absorption, metabolism and bioavailability of flavonoids: A review. Crit. Rev. Food Sci. Nutr. 2022, 62, 7730–7742. [Google Scholar] [CrossRef]
- De la Rosa, L.A.; Moreno-Escamilla, J.O.; Rodrigo-García, J.; Alvarez-Parrilla, E. Chapter 12—Phenolic compounds. In Posthar-vest Physiology and Biochemistry of Fruits and Vegetables; Yahia, E.M., Ed.; Woodhead Publishing: Duxford, UK, 2019; pp. 253–271. [Google Scholar] [CrossRef]
- Yen, G.-C.; Duh, P.-D.; Chuang, D.-Y. Antioxidant activity of anthraquinones and anthrone. Food Chem. 2010, 70, 437–441. [Google Scholar] [CrossRef]
- Remali, J.; Sahidin, I.; Aizat, W.M. Xanthone Biosynthetic Pathway in Plants: A Review. Front. Plant Sci. 2022, 13, 809497. [Google Scholar] [CrossRef]
- Hook, I.; Mills, C.; Sheridan, H. Bioactive naphthoquinones from higher plants. Stud. Nat. Prod. Chem. 2014, 41, 119–160. [Google Scholar]
- Grasel Fdos, S.; Ferrão, M.F.; Wolf, C.R. Development of methodology for identification the nature of the polyphenolic extracts by FTIR associated with multivariate analysis. Spectrochim. Acta A Mol. Biomol. Spectrosc. 2016, 153, 94–101. [Google Scholar] [CrossRef]
- Chouhan, S.; Sharma, K.; Zha, J.; Guleria, S.; Koffas, M.A.G. Recent Advances in the Recombinant Biosynthesis of Polyphenols. Front. Microbiol. 2017, 16, 2259. [Google Scholar] [CrossRef]
- Wang, Q.; Quan, S.; Xiao, H. Towards efficient terpenoid biosynthesis: Manipulating IPP and DMAPP supply. Bioresour. Bioprocess 2019, 6, 6. [Google Scholar] [CrossRef] [Green Version]
- Palenzuela, M.; Sánchez-Roa, D.; Damián, J.; Sessini, V.; Mosquera, M.E. Polymerization of terpenes and terpenoids using metal catalysts. Adv. Organomet. Chem. 2021, 75, 55–93. [Google Scholar]
- Aldred, E.M.; Buck, C.; Vall, K. Chapter 22-terpenes. In Pharmacology; Aldred, E.M., Buck, C., Vall, K., Eds.; Churchill Livingstone: Edinburgh, UK, 2009; pp. 167–174. [Google Scholar] [CrossRef]
- Temelli, F.; Saldaña, M.D.A.; Comin, L. Application of Supercritical Fluid Extraction in Food Processing. In Comprehensive Sampling and Sample Preparation; Elsevier: Amsterdam, The Netherlands, 2012; Volume 4. [Google Scholar] [CrossRef]
- Blažević, I.; Montaut, S.; Burčul, F.; Olsen, C.E.; Burow, M.; Rollin, P.; Agerbirk, N. Glucosinolate structural diversity, identification, chemical synthesis and metabolism in plants. Phytochemistry 2020, 169, 112100. [Google Scholar] [CrossRef] [PubMed]
- El-Saber Batiha, G.; Magdy Beshbishy, A.; Wasef, L.G.; Elewa, Y.H.A.; A. Al-Sagan, A.; Abd El-Hack, M.E.; Taha, A.E.; Abd-Elhakim, Y.M.; Prasad Devkota, H. Chemical Constituents and Pharmacological Activities of Garlic (Allium sativum L.): A Review. Nutrients 2020, 12, 872. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jones, W.P.; Kinghorn, A.D. Extraction of Plant Secondary Metabolites. In Natural Products Isolation. Methods in Molecular Biology; Sarker, S., Nahar, L., Eds.; Humana Press: Totowa, NJ, USA, 2012; Volume 864. [Google Scholar] [CrossRef]
- Hu, C.; Gan, X.; Jia, Q.; Gao, P.; Du, T.; Zhang, F. Optimization of supercritical-CO2 extraction and pharmacokinetics in SD rats of alkaloids form Sophora moorcroftiana seed. Sci. Rep. 2022, 12, 3301. [Google Scholar] [CrossRef]
- Freire, M.G.; Neves, C.M.S.S.; Marrucho, I.M.; Canongia Lopes, J.N.; Rebelo, L.P.N.; Coutinho, J.a.P. High-performance extraction of alkaloids using aqueous two-phase systems with ionic liquids. Green Chem. 2010, 12, 1715. [Google Scholar] [CrossRef]
- Torres-Vega, J.; Gómez-Alonso, S.; Pérez-Navarro, J.; Pastene-Navarrete, E. Green Extraction of Alkaloids and Polyphenols from Peumus boldus Leaves with Natural Deep Eutectic Solvents and Profiling by HPLC-PDA-IT-MS/MS and HPLC-QTOF-MS/MS. Plants 2020, 9, 242. [Google Scholar] [CrossRef] [Green Version]
- Belwal, T.; Pandey, A.; Bhatt, I.D.; Rawal, R.S. Optimized microwave assisted extraction (MAE) of alkaloids and polyphenols from Berberis roots using multiple-component analysis. Sci. Rep. 2020, 10, 917. [Google Scholar] [CrossRef]
- Dai, J.; Mumper, R.J. Plant phenolics: Extraction, analysis and their antioxidant and anticancer properties. Molecules 2010, 15, 7313–7352. [Google Scholar] [CrossRef]
- Madia, V.N.; De Vita, D.; Ialongo, D.; Tudino, V.; De Leo, A.; Scipione, L.; Di Santo, R.; Costi, R.; Messore, A. Recent Advances in Recovery of Lycopene from Tomato Waste: A Potent Antioxidant with Endless Benefits. Molecules 2021, 26, 4495. [Google Scholar] [CrossRef]
- Madia, V.N.; De Angelis, M.; De Vita, D.; Messore, A.; De Leo, A.; Ialongo, D.; Tudino, V.; Saccoliti, F.; De Chiara, G.; Garzoli, S.; et al. Investigation of Commiphora myrrha (Nees) Engl. Oil and Its Main Components for Antiviral Activity. Pharmaceuticals 2021, 14, 243. [Google Scholar] [CrossRef] [PubMed]
- Machado, C.A.; Oliveira, F.O.; de Andrade, M.A.; Hodel, K.V.S.; Lepikson, H.; Machado, B.A.S. Steam Distillation for Essential Oil Extraction: An Evaluation of Technological Advances Based on an Analysis of Patent Documents. Sustainability 2022, 14, 7119. [Google Scholar] [CrossRef]
- Doheny-Adams, T.; Redeker, K.; Kittipol, V.; Bancroft, I.; Hartley, S.E. Development of an efficient glucosinolate extraction method. Plant Methods 2017, 13, 17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zaini, A.S.; Putra, N.R.; Idham, Z.; Md Norodin, N.S.; Mohd Rasidek, N.A.; Che Yunus, M.A. Mini Review: Extraction of Allicin from Allium sativum using Subcritical Water Extraction. IOP Conf. Ser. Mater. Sci. Eng. 2020, 932, 012023. [Google Scholar] [CrossRef]
- Luo, Y.; Yin, S.; Lu, J.; Zhou, S.; Shao, Y.; Bao, X.; Wang, T.; Qiu, Y.; Yu, H. Tumor microenvironment: A prospective target of natural alkaloids for cancer treatment. Cancer Cell. Int. 2021, 21, 386. [Google Scholar] [CrossRef]
- El Omari, N.; Bakrim, S.; Bakha, M.; Lorenzo, J.M.; Rebezov, M.; Shariati, M.A.; Aboulaghras, S.; Balahbib, A.; Khayrullin, M.; Bouyahya, A. Natural Bioactive Compounds Targeting Epigenetic Pathways in Cancer: A Review on Alkaloids, Terpenoids, Quinones, and Isothiocyanates. Nutrients 2021, 13, 3714. [Google Scholar] [CrossRef]
- Spiller, G.A. Overview of the methylxanthine beverages and foods and their effect on health. Prog. Clin. Biol. Res. 1984, 158, 1–7. [Google Scholar]
- Stavric, B. Methylxanthines: Toxicity to humans. 1. Theophylline. Food Chem. Toxicol. 1988, 26, 541–565. [Google Scholar] [CrossRef]
- Horrigan, L.A.; Kelly, J.P.; Connor, T.J. Immunomodulatory effects of caffeine: Friend or foe? Pharmacol. Ther. 2006, 111, 877–892. [Google Scholar] [CrossRef]
- Persson, C.G. Overview of effects of theophylline. J. Allergy Clin. Immunol. 1986, 78, 780–787. [Google Scholar] [CrossRef]
- Ohta, A.; Sitkovsky, M. Methylxanthines, inflammation, and cancer: Fundamental mechanisms. Handb. Exp. Pharmacol. 2011, 200, 469–481. [Google Scholar]
- Tabolacci, C.; De Martino, A.; Mischiati, C.; Feriotto, G.; Beninati, S. The Role of Tissue Transglutaminase in Cancer Cell Initiation, Survival and Progression. Med. Sci. 2019, 7, 19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lentini, A.; Tabolacci, C.; Provenzano, B.; Rossi, S.; Beninati, S. Phytochemicals and protein-polyamine conjugates by transglutaminase as chemopreventive and chemotherapeutic tools in cancer. Plant Physiol. Biochem. 2010, 48, 627–633. [Google Scholar] [CrossRef]
- White, R.; Hanson, G.C.; Hu, F. Tyrosinase maturation and pigment expression in B16 melanoma: Relation to theophylline treatment and intracellular cyclic AMP. J. Cell. Physiol. 1979, 99, 441–450. [Google Scholar] [CrossRef]
- Menon, L.G.; Ingle, A.D.; Gude, R.P. Tumor regression of B16F10 melanoma in vivo by prevention of neovascularization: Study on theophylline. Cancer Biother. Radiopharm. 2002, 17, 213–217. [Google Scholar] [CrossRef] [PubMed]
- Lentini, A.; Autuori, F.; Mattioli, P.; Caraglia, M.; Abbruzzese, A.; Beninati, S. Evaluation of the efficacy of potential antineoplastic drugs on tumour metastasis by a computer-assisted image analysis. Eur. J. Cancer 2000, 36, 1572–1577. [Google Scholar] [CrossRef]
- Lee, S.J.; Lee, K.B.; Son, Y.H.; Shin, J.; Lee, J.H.; Kim, H.J.; Hong, A.Y.; Bae, H.W.; Kwon, M.A.; Lee, W.J.; et al. Transglutaminase 2 mediates UV-induced skin inflammation by enhancing inflammatory cytokine production. Cell Death Dis. 2017, 8, e3148. [Google Scholar] [CrossRef] [Green Version]
- Tabolacci, C.; Cordella, M.; Rossi, S.; Bonaccio, M.; Eramo, A.; Mischiati, C.; Beninati, S.; Iacoviello, L.; Facchiano, A.; Facchiano, F. Targeting Melanoma-Initiating Cells by Caffeine: In Silico and In Vitro Approaches. Molecules 2021, 26, 3619. [Google Scholar] [CrossRef]
- Cordella, M.; Tabolacci, C.; Senatore, C.; Rossi, S.; Mueller, S.; Lintas, C.; Eramo, A.; D’Arcangelo, D.; Valitutti, S.; Facchiano, A.; et al. Theophylline induces differentiation and modulates cytoskeleton dynamics and cytokines secretion in human melanoma-initiating cells. Life Sci. 2019, 230, 121–131. [Google Scholar] [CrossRef]
- Kamo, H.; Kawahara, R.; Simizu, S. Tyrosinase suppresses vasculogenic mimicry in human melanoma cells. Oncol. Lett. 2022, 23, 169. [Google Scholar] [CrossRef]
- Gude, R.P.; Binda, M.M.; Presas, H.L.; Klein-Szanto, A.J.; Bonfil, R.D. Studies on the mechanisms responsible for inhibition of experimental metastasis of B16-F10 murine melanoma by pentoxifylline. J. Biomed. Sci. 1999, 6, 133–141. [Google Scholar] [CrossRef] [PubMed]
- Dhyani, P.; Quispe, C.; Sharma, E.; Bahukhandi, A.; Sati, P.; Attri, D.C.; Szopa, A.; Sharifi-Rad, J.; Docea, A.O.; Mardare, I.; et al. Anticancer potential of alkaloids: A key emphasis to colchicine, vinblastine, vincristine, vindesine, vinorelbine and vincamine. Cancer Cell Int. 2022, 22, 206. [Google Scholar] [CrossRef] [PubMed]
- Garbe, C.; Peris, K.; Hauschild, A.; Saiag, P.; Middleton, M.; Spatz, A.; Grob, J.J.; Malvehy, J.; Newton-Bishop, J.; Stratigos, A.; et al. Diagnosis and treatment of melanoma. European consensus-based interdisciplinary guideline–Update 2012. Eur. J. Cancer 2012, 48, 2375–2390. [Google Scholar] [CrossRef] [PubMed]
- Vincent, L.A.; Attaoua, C.; Bellis, M.; Rozkydalova, L.; Hadj-Kaddour, K.; Vian, L.; Cuq, P. Lysosomes and unfolded protein response, determinants of differential resistance of melanoma cells to vinca alkaloids. Fundam. Clin. Pharmacol. 2015, 29, 164–177. [Google Scholar] [CrossRef] [PubMed]
- Jeon, K.I.; Xu, X.; Aizawa, T.; Lim, J.H.; Jono, H.; Kwon, D.S.; Abe, J.; Berk, B.C.; Li, J.D.; Yan, C. Vinpocetine inhibits NF-kappaB-dependent inflammation via an IKK-dependent but PDE-independent mechanism. Proc. Natl. Acad. Sci. USA 2010, 107, 9795–9800. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shimizu, K.; Murata, T.; Watanabe, Y.; Sato, C.; Morita, H.; Tagawa, T. Characterization of phosphodiesterase 1 in human malignant melanoma cell lines. Anticancer Res. 2009, 29, 1119–1122. [Google Scholar]
- Zhang, F.; Yan, C.; Wei, C.; Yao, Y.; Ma, X.; Gong, Z.; Liu, S.; Zang, D.; Chen, J.; Shi, F.D.; et al. Vinpocetine Inhibits NF-κB-Dependent Inflammation in Acute Ischemic Stroke Patients. Transl. Stroke Res. 2018, 9, 174–184. [Google Scholar] [CrossRef]
- Weng, H.J.; Tsai, T.F. ABCB1 in dermatology: Roles in skin diseases and their treatment. J. Mol. Med. 2021, 99, 1527–1538. [Google Scholar] [CrossRef]
- Xiong, R.G.; Huang, S.Y.; Wu, S.X.; Zhou, D.D.; Yang, Z.J.; Saimaiti, A.; Zhao, C.N.; Shang, A.; Zhang, Y.J.; Gan, R.Y.; et al. Anticancer Effects and Mechanisms of Berberine from Medicinal Herbs: An Update Review. Molecules 2022, 27, 4523. [Google Scholar] [CrossRef]
- Liu, J.F.; Lai, K.C.; Peng, S.F.; Maraming, P.; Huang, Y.P.; Huang, A.C.; Chueh, F.S.; Huang, W.W.; Chung, J.G. Berberine Inhibits Human Melanoma A375.S2 Cell Migration and Invasion via Affecting the FAK, uPA, and NF-κB Signaling Pathways and Inhibits PLX4032 Resistant A375.S2 Cell Migration In Vitro. Molecules 2018, 23, 2019. [Google Scholar] [CrossRef] [Green Version]
- Palma, T.V.; Bianchin, N.B.; de Oliveira, J.S.; Assmann, C.E.; das Neves Oliveira, M.; Schetinger, M.R.C.; Morsch, V.M.; Ulrich, H.; Pillat, M.M.; de Andrade, C.M. Berberine increases the expression of cytokines and proteins linked to apoptosis in human melanoma cells. Mol. Biol. Rep. 2022, 49, 2037–2046. [Google Scholar] [CrossRef] [PubMed]
- Fofaria, N.M.; Kim, S.H.; Srivastava, S.K. Piperine causes G1 phase cell cycle arrest and apoptosis in melanoma cells through checkpoint kinase-1 activation. PLoS ONE 2014, 9, e94298. [Google Scholar] [CrossRef] [PubMed]
- Pradeep, C.R.; Kuttan, G. Piperine is a potent inhibitor of nuclear factor-kappaB (NF-kappaB), c-Fos, CREB, ATF-2 and proinflammatory cytokine gene expression in B16F-10 melanoma cells. Int. Immunopharmacol. 2004, 4, 1795–1803. [Google Scholar] [CrossRef]
- Yoo, E.S.; Choo, G.S.; Kim, S.H.; Woo, J.S.; Kim, H.J.; Park, Y.S.; Kim, B.S.; Kim, S.K.; Park, B.K.; Cho, S.D.; et al. Antitumor and Apoptosis-inducing Effects of Piperine on Human Melanoma Cells. Anticancer Res. 2019, 39, 1883–1892. [Google Scholar] [CrossRef] [PubMed]
- Chu, H.; Li, M.; Wang, X. Capsaicin induces apoptosis and autophagy in human melanoma cells. Oncol. Lett. 2019, 17, 4827–4834. [Google Scholar] [CrossRef] [Green Version]
- Hong, J.H.; Chen, H.J.; Xiang, S.J.; Cao, S.W.; An, B.C.; Ruan, S.F.; Zhang, B.; Weng, L.D.; Zhu, H.X.; Liu, Q. Capsaicin reverses the inhibitory effect of licochalcone A/β-Arbutin on tyrosinase expression in b16 mouse melanoma cells. Pharmacogn. Mag. 2018, 14, 110–115. [Google Scholar] [CrossRef]
- Wu, Q.; Bai, P.; Xia, Y.; Xia, Y.; Xu, B.; Dai, K.; Zheng, Z.; Guo, M.S.S.; Fung, K.W.C.; Dong, T.T.X.; et al. Capsaicin Inhibits the Expression of Melanogenic Proteins in Melanocyte via Activation of TRPV1 Channel: Identifying an Inhibitor of Skin Melanogenesis. J. Agric. Food Chem. 2020, 68, 14863–14873. [Google Scholar] [CrossRef]
- Zhu, Z.; Zhao, S.; Wang, C. Antibacterial, antifungal, antiviral, and antiparasitic activities of Peganum harmala and its ingredients: A review. Molecules 2022, 27, 4161. [Google Scholar] [CrossRef]
- Park, S.Y.; Kim, Y.H.; Kim, Y.H.; Park, G.; Lee, S.J. Beta-carboline alkaloids harmaline and harmalol induce melanogenesis through p38 mitogen-activated protein kinase in B16F10 mouse melanoma cells. BMB Rep. 2010, 43, 824–829. [Google Scholar] [CrossRef] [Green Version]
- Hamsa, T.; Kuttan, G. Ipobscurine, an indole alkaloid from Ipomoea obscura, inhibits tumor cell invasion and experimental metastasis by inducing apoptosis. J. Environ. Pathol. Toxicol. Oncol. 2011, 30, 163–178. [Google Scholar] [CrossRef]
- Hamsa, T.P.; Kuttan, G. Anti-angiogenic activity of Ipomoea obscura extract and Ipobscurine-A. Immunopharmacol. Immunotoxicol. 2011, 33, 488–497. [Google Scholar] [CrossRef] [PubMed]
- Shen, N.; Wang, T.; Gan, Q.; Liu, S.; Wang, L.; Jin, B. Plant flavonoids: Classification, distribution, biosynthesis, and antioxidant activity. Food Chem. 2022, 383, 132531. [Google Scholar] [CrossRef] [PubMed]
- Forni, C.; Facchiano, F.; Bartoli, M.; Pieretti, S.; Facchiano, A.; D’Arcangelo, D.; Norelli, S.; Valle, G.; Nisini, R.; Beninati, S.; et al. Beneficial Role of Phytochemicals on Oxidative Stress and Age-Related Diseases. Biomed Res. Int. 2019, 2019, 8748253. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Forni, C.; Rossi, M.; Borromeo, I.; Feriotto, G.; Platamone, G.; Tabolacci, C.; Mischiati, C.; Beninati, S. Flavonoids: A Myth or a Reality for Cancer Therapy? Molecules 2021, 26, 3583. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Pandey, A.K. Chemistry and biological activities of flavonoids: An overview. Sci. World J. 2013, 2013, 162750. [Google Scholar] [CrossRef] [Green Version]
- Leyva-López, N.; Gutierrez-Grijalva, E.P.; Ambriz-Perez, D.L.; Heredia, J.B. Flavonoids as Cytokine Modulators: A Possible Therapy for Inflammation-Related Diseases. Int. J. Mol. Sci. 2016, 17, 921. [Google Scholar] [CrossRef]
- Jantan, I.; Haque, M.A.; Arshad, L.; Harikrishnan, H.; Septama, A.W.; Mohamed-Hussein, Z.A. Dietary polyphenols suppress chronic inflammation by modulation of multiple inflammation-associated cell signaling pathways. J. Nutr. Biochem. 2021, 93, 108634. [Google Scholar] [CrossRef]
- Al-Khayri, J.M.; Sahana, G.R.; Nagella, P.; Joseph, B.V.; Alessa, F.M.; Al-Mssallem, M.Q. Flavonoids as Potential Anti-Inflammatory Molecules: A Review. Molecules 2022, 27, 2901. [Google Scholar] [CrossRef]
- Shukla, S.; Gupta, S. Apigenin: A promising molecule for cancer prevention. Pharm. Res. 2010, 27, 962–978. [Google Scholar] [CrossRef] [Green Version]
- Xu, L.; Zhang, Y.; Tian, K.; Chen, X.; Zhang, R.; Mu, X.; Wu, Y.; Wang, D.; Wang, S.; Liu, F.; et al. Apigenin suppresses PD-L1 expression in melanoma and host dendritic cells to elicit synergistic therapeutic effects. J. Exp. Clin. Cancer Res. 2018, 37, 261. [Google Scholar] [CrossRef] [Green Version]
- Bouhlel Chatti, I.; Krichen, Y.; Maatouk, M.; Lahmar, A.; Ghoul Mazgar, S.; Kammoun, R.; Safta Skhiri, S.; Chekir Ghedira, L.; Krifa, M. Evaluation of Anticancer Potential of Flavones from Rhamnus alaternus against B16F10 Melanoma Cells. Nutr. Cancer 2022, 74, 2265–2275. [Google Scholar] [CrossRef] [PubMed]
- Ding, J.; Polier, G.; Köhler, R.; Giaisi, M.; Krammer, P.H.; Li-Weber, M. Wogonin and related natural flavones overcome tumor necrosis factor-related apoptosis-inducing ligand (TRAIL) protein resistance of tumors by down-regulation of c-FLIP protein and up-regulation of TRAIL receptor 2 expression. J. Biol. Chem. 2012, 287, 641–649. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, S.; Wang, S.; Liu, S.; Li, Z.; Liu, X.; Wu, J. Baicalein Potentiated M1 Macrophage Polarization in Cancer Through Targeting PI3Kγ/NF-κB Signaling. Front. Pharmacol. 2021, 12, 743837. [Google Scholar] [CrossRef] [PubMed]
- Tian, L.; Wang, S.; Jiang, S.; Liu, Z.; Wan, X.; Yang, C.; Zhang, L.; Zheng, Z.; Wang, B.; Li, L. Luteolin as an adjuvant effectively enhances CTL anti-tumor response in B16F10 mouse model. Int. Immunopharmacol. 2021, 94, 107441. [Google Scholar] [CrossRef] [PubMed]
- Schomberg, J.; Wang, Z.; Farhat, A.; Guo, K.L.; Xie, J.; Zhou, Z.; Liu, J.; Kovacs, B.; Liu-Smith, F. Luteolin inhibits melanoma growth in vitro and in vivo via regulating ECM and oncogenic pathways but not ROS. Biochem. Pharmacol. 2020, 177, 114025. [Google Scholar] [CrossRef]
- Peng, D.; Chen, L.; Sun, Y.; Sun, L.; Yin, Q.; Deng, S.; Niu, L.; Lou, F.; Wang, Z.; Xu, Z.; et al. Melanoma suppression by quercein is correlated with RIG-I and type I interferon signaling. Biomed. Pharmacother. 2020, 125, 109984. [Google Scholar] [CrossRef]
- Li, L.; Zhang, M.; Liu, T.; Li, J.; Sun, S.; Chen, J.; Liu, Z.; Zhang, Z.; Zhang, L. Quercetin-ferrum nanoparticles enhance photothermal therapy by modulating the tumor immunosuppressive microenvironment. Acta Biomater. 2022, 154, 454–466. [Google Scholar] [CrossRef]
- Lian, G.Y.; Wang, Q.M.; Mak, T.S.; Huang, X.R.; Yu, X.Q.; Lan, H.Y. Inhibition of tumor invasion and metastasis by targeting TGF-β-Smad-MMP2 pathway with Asiatic acid and Naringenin. Mol. Ther. Oncolytics 2021, 20, 277–289. [Google Scholar] [CrossRef]
- Jiang, S.; Wang, S.; Zhang, L.; Tian, L.; Li, L.; Liu, Z.; Dong, Q.; Lv, X.; Mu, H.; Zhang, Q.; et al. Hesperetin as an adjuvant augments protective anti-tumour immunity responses in B16F10 melanoma by stimulating cytotoxic CD8+ T cells. Scand. J. Immunol. 2020, 91, e12867. [Google Scholar] [CrossRef]
- Basu, P.; Maier, C. Phytoestrogens and breast cancer: In vitro anticancer activities of isoflavones, lignans, coumestans, stilbenes and their analogs and derivatives. Biomed. Pharmacother. 2018, 107, 1648–1666. [Google Scholar] [CrossRef]
- Guo, T.L.; McCay, J.A.; Zhang, L.X.; Brown, R.D.; You, L.; Karrow, N.A.; Germolec, D.R.; White, K.L., Jr. Genistein modulates immune responses and increases host resistance to B16F10 tumor in adult female B6C3F1 mice. J. Nutr. 2001, 131, 3251–3258. [Google Scholar] [CrossRef] [PubMed]
- Venza, I.; Visalli, M.; Oteri, R.; Beninati, C.; Teti, D.; Venza, M. Genistein reduces proliferation of EP3-expressing melanoma cells through inhibition of PGE2-induced IL-8 expression. Int. Immunopharmacol. 2018, 62, 86–95. [Google Scholar] [CrossRef] [PubMed]
- Chen, B.; Zhang, W.; Lin, C.; Zhang, L. A Comprehensive Review on Beneficial Effects of Catechins on Secondary Mitochondrial Diseases. Int. J. Mol. Sci. 2022, 23, 11569. [Google Scholar] [CrossRef] [PubMed]
- Nihal, M.; Ahsan, H.; Siddiqui, I.A.; Mukhtar, H.; Ahmad, N.; Wood, G.S. (-)-Epigallocatechin-3-gallate (EGCG) sensitizes melanoma cells to interferon induced growth inhibition in a mouse model of human melanoma. Cell Cycle 2009, 8, 2057–2063. [Google Scholar] [CrossRef] [Green Version]
- Shen, Q.; Tian, F.; Jiang, P.; Li, Y.; Zhang, L.; Lu, J.; Li, J. EGCG enhances TRAIL-mediated apoptosis in human melanoma A375 cell line. J. Huazhong Univ. Sci. Technolog. Med. Sci. 2009, 29, 771–775. [Google Scholar] [CrossRef]
- Ellis, L.Z.; Liu, W.; Luo, Y.; Okamoto, M.; Qu, D.; Dunn, J.H.; Fujita, M. Green tea polyphenol epigallocatechin-3-gallate suppresses melanoma growth by inhibiting inflammasome and IL-1β secretion. Biochem. Biophys. Res. Commun. 2011, 414, 551–556. [Google Scholar] [CrossRef] [Green Version]
- Xu, A.; Lee, J.; Zhao, Y.; Wang, Y.; Li, X.; Xu, P. Potential effect of EGCG on the anti-tumor efficacy of metformin in melanoma cells. J. Zhejiang Univ. Sci. B 2021, 22, 548–562. [Google Scholar] [CrossRef]
- Kang, T.H.; Lee, J.H.; Song, C.K.; Han, H.D.; Shin, B.C.; Pai, S.I.; Hung, C.F.; Trimble, C.; Lim, J.S.; Kim, T.W.; et al. Epigallocatechin-3-gallate enhances CD8+ T cell-mediated antitumor immunity induced by DNA vaccination. Cancer Res. 2007, 67, 802–811. [Google Scholar] [CrossRef] [Green Version]
- Ravindran Menon, D.; Li, Y.; Yamauchi, T.; Osborne, D.G.; Vaddi, P.K.; Wempe, M.F.; Zhai, Z.; Fujita, M. EGCG Inhibits Tumor Growth in Melanoma by Targeting JAK-STAT Signaling and Its Downstream PD-L1/PD-L2-PD1 Axis in Tumors and Enhancing Cytotoxic T-Cell Responses. Pharmaceuticals 2021, 14, 1081. [Google Scholar] [CrossRef]
- Bunea, A.; Rugină, D.; Sconţa, Z.; Pop, R.M.; Pintea, A.; Socaciu, C.; Tăbăran, F.; Grootaert, C.; Struijs, K.; VanCamp, J. Anthocyanin determination in blueberry extracts from various cultivars and their antiproliferative and apoptotic properties in B16-F10 metastatic murine melanoma cells. Phytochemistry 2013, 95, 436–444. [Google Scholar] [CrossRef]
- Wang, E.; Liu, Y.; Xu, C.; Liu, J. Antiproliferative and proapoptotic activities of anthocyanin and anthocyanidin extracts from blueberry fruits on B16-F10 melanoma cells. Food Nutr. Res. 2017, 61, 1325308. [Google Scholar] [CrossRef]
- Diaconeasa, Z.; Știrbu, I.; Xiao, J.; Leopold, N.; Ayvaz, Z.; Danciu, C.; Ayvaz, H.; Stǎnilǎ, A.; Nistor, M.; Socaciu, C. Anthocyanins, Vibrant Color Pigments, and Their Role in Skin Cancer Prevention. Biomedicines 2020, 8, 336. [Google Scholar] [CrossRef] [PubMed]
- Afaq, F.; Saleem, M.; Krueger, C.G.; Reed, J.D.; Mukhtar, H. Anthocyanin- and hydrolyzable tannin-rich pomegranate fruit extract modulates MAPK and NF-kappaB pathways and inhibits skin tumorigenesis in CD-1 mice. Int. J. Cancer 2005, 113, 423–433. [Google Scholar] [CrossRef]
- Afaq, F.; Katiyar, S.K. Polyphenols: Skin photoprotection and inhibition of photocarcinogenesis. Mini Rev. Med. Chem. 2011, 11, 1200–1215. [Google Scholar] [CrossRef] [Green Version]
- Ma, E.Z.; Khachemoune, A. Flavonoids and their therapeutic applications in skin diseases. Arch. Dermatol. Res. 2022, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Lu, Y.; Ai, Y.; Xu, X.; Zhu, S.; Zhang, B.; Tang, M.; Zhang, L.; He, T. Glabridin Liposome Ameliorating UVB-Induced Erythema and Lethery Skin by Suppressing Inflammatory Cytokine Production. J. Microbiol. Biotechnol. 2021, 31, 630–636. [Google Scholar] [CrossRef]
- Shyong, E.Q.; Lu, Y.; Lazinsky, A.; Saladi, R.N.; Phelps, R.G.; Austin, L.M.; Lebwohl, M.; Wei, H. Effects of the isoflavone 4’,5,7-trihydroxyisoflavone (genistein) on psoralen plus ultraviolet A radiation (PUVA)-induced photodamage. Carcinogenesis 2002, 23, 317–321. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rajnochová Svobodová, A.; Gabrielová, E.; Michaelides, L.; Kosina, P.; Ryšavá, A.; Ulrichová, J.; Zálešák, B.; Vostálová, J. UVA-photoprotective potential of silymarin and silybin. Arch. Dermatol. Res. 2018, 310, 413–424. [Google Scholar] [CrossRef] [PubMed]
- Meeran, S.M.; Mantena, S.K.; Katiyar, S.K. Prevention of ultraviolet radiation-induced immunosuppression by (-)-epigallocatechin-3-gallate in mice is mediated through interleukin 12-dependent DNA repair. Clin. Cancer Res. 2006, 12, 2272–2280. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Penta, D.; Somashekar, B.S.; Meeran, S.M. Epigenetics of skin cancer: Interventions by selected bioactive phytochemicals. Photodermatol. Photoimmunol. Photomed. 2018, 34, 42–49. [Google Scholar] [CrossRef]
- Greenberg, E.S.; Chong, K.K.; Huynh, K.T.; Tanaka, R.; Hoon, D.S. Epigenetic biomarkers in skin cancer. Cancer Lett. 2014, 342, 170–177. [Google Scholar] [CrossRef] [PubMed]
- Nandakumar, V.; Vaid, M.; Katiyar, S.K. (-)-Epigallocatechin-3-gallate reactivates silenced tumor suppressor genes, Cip1/p21 and p16INK4a, by reducing DNA methylation and increasing histones acetylation in human skin cancer cells. Carcinogenesis 2011, 32, 537–544. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saha, K.; Hornyak, T.J.; Eckert, R.L. Epigenetic cancer prevention mechanisms in skin cancer. AAPS J. 2013, 15, 1064–1071. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prasad, R.; Katiyar, S.K. Polyphenols from green tea inhibit the growth of melanoma cells through inhibition of class I histone deacetylases and induction of DNA damage. Genes Cancer 2015, 6, 49–61. [Google Scholar] [CrossRef] [PubMed]
- Dumitraș, D.A.; Sanda Andrei, S. Recent Advances in the Antiproliferative and Proapoptotic Activity of Various Plant Extracts and Constituents against Murine Malignant Melanoma. Molecules 2022, 27, 2585. [Google Scholar] [CrossRef] [PubMed]
- Palko-Łabuz, A.; Gliszczyńska, A.; Skonieczna, M.; Poła, A.; Wesołowska, O.; Środa-Pomianek, K. Conjugation with Phospholipids as a Modification Increasing Anticancer Activity of Phenolic Acids in Metastatic Melanoma-In Vitro and In Silico Studies. Int. J. Mol. Sci. 2021, 22, 8397. [Google Scholar] [CrossRef]
- Srinivasan, A.; Ekambaram, S.P.; Perumal, S.S.; Aruldhas, J.; Erusappan, T. Chemical characterization and immunostimulatory activity of phenolic acid bound arabinoxylans derived from foxtail and barnyard millets. J. Food Biochem. 2020, 44, e13116. [Google Scholar] [CrossRef]
- Park, J.; Chung, S.W.; Kim, S.H.; Kim, T.S. Up-regulation of interleukin-4 production via NF-AT/AP-1 activation in T cells by biochanin A, a phytoestrogen and its metabolites. Toxicol. Appl. Pharmacol. 2006, 212, 188–199. [Google Scholar] [CrossRef]
- Kauroo, S.; Govinden-Soulange, G.; Ranghoo-Sanmukhiya, V.M.; Miranda, K.; Cotham, W.E.; Walla, M.D.; Nagarkatti, M.; Nagarkatti, P. Extracts of select endemic plants from the Republic of Mauritius exhibiting anti-cancer and immunomodulatory properties. Sci. Rep. 2021, 11, 4272. [Google Scholar] [CrossRef]
- Liu, H.; Gao, H.; Chen, C.; Jia, W.; Xu, D.; Jiang, G. IDO Inhibitor and Gallic Acid Cross-Linked Small Molecule Drug Synergistic Treatment of Melanoma. Front. Oncol. 2022, 12, 904229. [Google Scholar] [CrossRef]
- Prieto, K.; Lozano, M.P.; Urueña, C.; Alméciga-Díaz, C.J.; Fiorentino, S.; Barreto, A. The delay in cell death caused by the induction of autophagy by P2Et extract is essential for the generation of immunogenic signals in melanoma cells. Apoptosis 2020, 25, 875–888. [Google Scholar] [CrossRef] [PubMed]
- Ohno, T.; Inoue, M.; Ogihara, Y. Cytotoxic activity of gallic acid against liver metastasis of mastocytoma cells P-815. Anticancer Res. 2001, 21, 3875–3880. [Google Scholar] [PubMed]
- Pramanik, K.C.; Kudugunti, S.K.; Fofaria, N.M.; Moridani, M.Y.; Srivastava, S.K. Caffeic acid phenethyl ester suppresses melanoma tumor growth by inhibiting PI3K/AKT/XIAP pathway. Carcinogenesis 2013, 34, 2061–2070. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Staniforth, V.; Chiu, L.T.; Yang, N.S. Caffeic acid suppresses UVB radiation-induced expression of interleukin-10 and activation of mitogen-activated protein ki-nases in mouse. Carcinogenesis 2006, 27, 1803–1811. [Google Scholar] [CrossRef] [Green Version]
- Feng, D.; Fang, Z.; Zhang, P. The melanin inhibitory effect of plants and phytochemicals: A systematic review. Phytomedicine 2022, 107, 154449. [Google Scholar] [CrossRef]
- Rodboon, T.; Okada, S.; Suwannalert, P. Germinated Riceberry Rice Enhanced Protocatechuic Acid and Vanillic Acid to Suppress Melanogenesis through Cellular Oxidant-Related Tyrosinase Activity in B16 Cells. Antioxidants 2020, 9, 247. [Google Scholar] [CrossRef] [Green Version]
- Sun, L.; Guo, Y.; Zhang, Y.; Zhuang, Y. Antioxidant and Anti-tyrosinase Activities of Phenolic Extracts from Rape Bee Pollen and Inhibitory Melanogenesis by cAMP/MITF/TYR Pathway in B16 Mouse Melanoma Cells. Front. Pharmacol. 2017, 8, 104. [Google Scholar] [CrossRef] [Green Version]
- Maack, A.; Pegard, A. Populus nigra (Salicaceae) absolute rich in phenolic acids, phenylpropanoids and flavonoids as a new potent tyrosinase inhibitor. Fitoterapia 2016, 111, 95–101. [Google Scholar] [CrossRef]
- Liu, J.; Jiang, R.; Zhou, J.; Xu, X.; Sun, Z.; Li, J.; Chen, X.; Li, Z.; Yan, X.; Zhao, D.; et al. Salicylic acid in ginseng root alleviates skin hyperpigmentation disorders by inhibiting melanogenesis and melanosome transport. Eur. J. Pharmacol. 2021, 910, 174458. [Google Scholar] [CrossRef]
- Chao, E.; Tian, J.; Fan, L.; Zhang, T. Drying methods influence the physicochemical and functional properties of seed-use pumpkin. Food Chem. 2022, 369, 130937. [Google Scholar] [CrossRef]
- Hu, X.; Yang, Z.; Liu, W.; Pan, Z.; Zhang, X.; Li, M.; Liu, X.; Zheng, Q.; Li, D. The Anti-tumor Effects of p-Coumaric Acid on Melanoma A375 and B16 Cells. Front. Oncol. 2020, 10, 558414. [Google Scholar] [CrossRef] [PubMed]
- Catalgol, B.; Batirel, S.; Taga, Y.; Ozer, N.K. Resveratrol: French paradox revisited. Front. Pharmacol. 2012, 3, 141. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Malaguarnera, L. Influence of Resveratrol on the Immune Response. Nutrients 2019, 11, 946. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Davoodvandi, A.; Darvish, M.; Borran, S.; Nejati, M.; Mazaheri, S.; Reza Tamtaji, O.; Hamblin, M.R.; Masoudian, N.; Mirzaei, H. The therapeutic potential of resveratrol in a mouse model of melanoma lung metastasis. Int. Immunopharmacol. 2020, 88, 106905. [Google Scholar] [CrossRef] [PubMed]
- De Sá Coutinho, D.; Pacheco, M.T.; Frozza, R.L.; Bernardi, A. Anti-Inflammatory Effects of Resveratrol: Mechanistic Insights. Int. J. Mol. Sci. 2018, 19, 1812. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Misawa, T.; Saitoh, T.; Kozaki, T.; Park, S.; Takahama, M.; Akira, S. Resveratrol inhibits the acetylated α-tubulin-mediated assembly of the NLRP3-inflammasome. Int. Immunol. 2015, 27, 425–434. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leischner, C.; Burkard, M.; Pfeiffer, M.M.; Lauer, U.M.; Busch, C.; Venturelli, S. Nutritional immunology: Function of natural killer cells and their modulation by resveratrol for cancer prevention and treatment. Nutr. J. 2016, 15, 47. [Google Scholar] [CrossRef] [Green Version]
- Feng, L.; Yasmeen, R.; Schoene, N.W.; Lei, K.Y.; Wang, T.T.Y. Resveratrol differentially modulates immune responses in human THP-1 monocytes and macrophages. Nutr. Res. 2019, 72, 57–69. [Google Scholar] [CrossRef]
- Lee-Chang, C.; Bodogai, M.; Martin-Montalvo, A.; Wejksza, K.; Sanghvi, M.; Moaddel, R.; de Cabo, R.; Biragyn, A. Inhibition of breast cancer metastasis by resveratrol-mediated inactivation of tumor-evoked regulatory B cells. J. Immunol. 2013, 191, 4141–4151. [Google Scholar] [CrossRef] [Green Version]
- Doh, K.C.; Kim, B.M.; Kim, K.W.; Chung, B.H.; Yang, C.W. Effects of resveratrol on Th17 cell-related immune responses under tacrolimus-based immunosuppression. BMC Complement. Altern. Med. 2019, 19, 54. [Google Scholar] [CrossRef] [Green Version]
- Verdura, S.; Cuyàs, E.; Cortada, E.; Brunet, J.; Lopez-Bonet, E.; Martin-Castillo, B.; Bosch-Barrera, J.; Encinar, J.A.; Menendez, J.A. Resveratrol targets PD-L1 glycosylation and dimerization to enhance antitumor T-cell immunity. Aging 2020, 12, 8–34. [Google Scholar] [CrossRef] [PubMed]
- Nakagami, Y.; Suzuki, S.; Espinoza, J.L.; Vu Quang, L.; Enomoto, M.; Takasugi, S.; Nakamura, A.; Nakayama, T.; Tani, H.; Hanamura, I.; et al. Immunomodulatory and Metabolic Changes after Gnetin-C Supplementation in Humans. Nutrients 2019, 11, 1403. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kuo, I.M.; Lee, J.J.; Wang, Y.S.; Chiang, H.C.; Huang, C.C.; Hsieh, P.J.; Han, W.; Ke, C.H.; Liao, A.T.C.; Lin, C.S. Potential enhancement of host immunity and anti-tumor efficacy of nanoscale curcumin and resveratrol in colorectal cancers by modulated electro-hyperthermia. BMC Cancer 2020, 20, 603. [Google Scholar] [CrossRef]
- Li, Z.; Kong, D.; Liu, Y.; Mingkai Lia, M. Pharmacological perspectives and molecular mechanisms of coumarin derivatives against virus disease. Genes Dis. 2022, 9, 80–94. [Google Scholar] [CrossRef] [PubMed]
- Bhattarai, N.; Kumbhar, A.A.; Pokharel, Y.R.; Yadav, P.N. Anticancer Potential of Coumarin and its Derivatives. Mini Rev. Med. Chem. 2021, 21, 2996–3029. [Google Scholar] [CrossRef] [PubMed]
- Albiero, L.R.; Fernandes de Andrade, M.; Marchi, L.F.; Landi-Librandi, A.P.; Garcia de Figueiredo-Rinhel, A.S.; Carvalho, C.A.; Kabeya, L.M.; de Oliveira, R.D.R.; Azzolini, A.E.C.S.; Pupo, M.T.; et al. Immunomodulating action of the 3-phenylcoumarin derivative 6,7-dihydroxy-3-[3’,4’-methylenedioxyphenyl]-coumarin in neutrophils from patients with rheumatoid arthritis and in rats with acute joint inflammation. Inflamm. Res. 2020, 69, 115–130. [Google Scholar] [CrossRef]
- Marshall, M.E.; Rhoades, J.L.; Mattingly, C.; Jennings, C.D. Coumarin (1,2-benzopyrone) enhances DR and DQ antigen expressions by peripheral blood mononuclear cells in vitro. Mol. Biother. 1991, 3, 204–206. [Google Scholar]
- Song, B.; Wang, Z.; Liu, Y.; Xu, S.; Huang, G.; Xiong, Y.; Zhang, S.; Xu, L.; Deng, X.; Guan, S. Immunosuppressive activity of daphnetin, one of coumarin derivatives, is mediated through suppression of NF-κB and NFAT signaling pathways in mouse T cells. PLoS ONE 2014, 9, e96502. [Google Scholar] [CrossRef]
- Maghraby, A.; Bahgat, M. Immunostimulatory effect of coumarin derivatives before and after infection of mice with the parasite Schistosoma mansoni. Arzneimittelforschung 2004, 54, 545–550. [Google Scholar] [CrossRef]
- Marrelli, M.; Perri, M.R.; Amodeo, V.; Giordano, F.; Statti, G.A.; Panno, M.L.; Conforti, F. Assessment of Photo-Induced Cytotoxic Activity of Cachrys sicula and Cachrys libanotis Enriched-Coumarin Extracts against Human Melanoma Cells. Plants 2021, 10, 123. [Google Scholar] [CrossRef]
- Jeon, Y.J.; Jang, J.Y.; Shim, J.H.; Myung, P.K.; Chae, J.I. Esculetin, a Coumarin Derivative, Exhibits Anti-proliferative and Pro-apoptotic Activity in G361 Human Malignant Melanoma. J. Cancer Prev. 2015, 20, 106–112. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Charmforoshan, E.; Karimi, E.; Oskoueian, E.; Iranshahi, M. Antibacterial, Antioxidant and Melanogenesis Inhibitory Activity of Auraptene, a Coumarin from Ferula szowitsiana Root. Nutr. Cancer. 2022, 74, 1829–1836. [Google Scholar] [CrossRef] [PubMed]
- Fontes, S.S.; Nogueira, M.L.; Dias, R.B.; Rocha, C.A.-G.; Soares, M.B.P.; Vannier-Santos, M.A.; Bezerra, D.P. Combination Therapy of Curcumin and Disulfiram Synergistically Inhibits the Growth of B16-F10 Melanoma Cells by Inducing Oxidative Stress. Biomolecules 2022, 12, 1600. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Lu, S.; Ma, C. Anti-proliferative and pro-apoptotic effects of curcumin on skin cutaneous melanoma: Bioinformatics analysis and in vitro experimental studies. Front. Genet. 2022, 13, 983943. [Google Scholar] [CrossRef] [PubMed]
- Chiu, Y.J.; Yang, J.S.; Tsai, F.J.; Chiu, H.Y.; Juan, Y.N.; Lo, Y.H.; Chiang, J.H. Curcumin suppresses cell proliferation and triggers apoptosis in vemurafenib-resistant melanoma cells by downregulating the EGFR signaling pathway. Environ. Toxicol. 2022, 37, 868–879. [Google Scholar] [CrossRef] [PubMed]
- Mirzaei, H.; Naseri, G.; Rezaee, R.; Mohammadi, M.; Banikazemi, Z.; Mirzaei, H.R.; Salehi, H.; Peyvandi, M.; Pawelek, J.M.; Sahebkar, A. Curcumin: A new candidate for melanoma therapy? Int. J. Cancer 2016, 139, 1683–1695. [Google Scholar] [CrossRef]
- Tang, Y.; Cao, Y. Curcumin Inhibits the Growth and Metastasis of Melanoma via miR-222-3p/SOX10/Notch Axis. Dis. Markers 2022, 2022, 3129781. [Google Scholar] [CrossRef]
- Wang, D.; Sang, Y.; Sun, T.; Kong, P.; Zhang, L.; Dai, Y.; Cao, Y.; Tao, Z.; Liu, W. Emerging roles and mechanisms of microRNA-222-3p in human cancer. Int. J. Oncol. 2021, 58, 20. [Google Scholar] [CrossRef]
- Allegra, A.; Mirabile, G.; Ettari, R.; Pioggia, G.; Gangemi, S. The Impact of Curcumin on Immune Response: An Immunomodulatory Strategy to Treat Sepsis. Int. J. Mol. Sci. 2022, 23, 14710. [Google Scholar] [CrossRef]
- Tu, C.X.; Lin, M.; Lu, S.S.; Qi, X.Y.; Zhang, R.X.; Zhang, Y.Y. Curcumin inhibits melanogenesis in human melanocytes. Phytother. Res. 2012, 26, 174–179. [Google Scholar] [CrossRef]
- Zhu, H.; Zhang, H.; Hou, B.; Xu, B.; Ji, L.; Wu, Y. Curcumin Regulates Gut Microbiota and Exerts a Neuroprotective Effect in the MPTP Model of Parkinson’s Disease. Evid. Based Complement. Alternat. Med. 2022, 2022, 9110560. [Google Scholar] [CrossRef] [PubMed]
- Li, C.Y.; Wang, Q.; Wang, X.M.; Li, G.X.; Shen, S.; Wei, X.L. Gambogic acid exhibits anti-metastatic activity on malignant melanoma mainly through inhibition of PI3K/Akt and ERK signaling pathways. Eur. J. Pharmacol. 2019, 864, 172719. [Google Scholar] [CrossRef]
- Saha, S.; Sadhukhan, P.; Sil, P.C. Mangiferin: A xanthonoid with multipotent anti-inflammatory potential. Biofactors 2016, 42, 459–474. [Google Scholar] [CrossRef] [PubMed]
- Takeda, T.; Tsubaki, M.; Sakamoto, K.; Ichimura, E.; Enomoto, A.; Suzuki, Y.; Itoh, T.; Imano, M.; Tanabe, G.; Muraoka, O.; et al. Mangiferin, a novel nuclear factor kappa B-inducing kinase inhibitor, suppresses metastasis and tumor growth in a mouse metastatic melanoma model. Toxicol. Appl. Pharmacol. 2016, 306, 105–112. [Google Scholar] [CrossRef]
- Delgado-Hernández, R.; Hernández-Balmaseda, I.; Rodeiro-Guerra, I.; Cesar Rodriguez Gonzalez, J.; De Wever, O.; Logie, E.; Declerck, K.; Pérez-Novo, C.; Vanden Berghe, W. Anti-angiogenic effects of mangiferin and mechanism of action in metastatic melanoma. Melanoma Res. 2020, 30, 39–51. [Google Scholar] [CrossRef]
- Imran, M.; Arshad, M.S.; Butt, M.S.; Kwon, J.H.; Arshad, M.U.; Sultan, M.T. Mangiferin: A natural miracle bioactive compound against lifestyle related disorders. Lipids Health Dis. 2017, 16, 84. [Google Scholar] [CrossRef] [PubMed]
- Hisa, T.; Kimura, Y.; Takada, K.; Suzuki, F.; Takigawa, M. Shikonin, an ingredient of Lithospermum erythrorhizon, inhibits angiogenesis in vivo and in vitro. Anticancer Res. 1998, 18, 783–790. [Google Scholar]
- Chen, H.M.; Wang, P.H.; Aravindaram, K.; Chen, Y.H.; Yu, H.H.; Yang, W.C.; Yang, N.S. Shikonin enhances efficacy of a gene-based cancer vaccine via induction of RANTES. J. Biomed. Sci. 2012, 19, 42. [Google Scholar] [CrossRef] [Green Version]
- Sanders, B.; Ray, A.M.; Goldberg, S.; Clark, T.; McDaniel, H.R.; Atlas, S.E.; Farooqi, A.; Konefal, J.; Lages, L.C.; Lopez, J.; et al. Anti-cancer effects of aloe-emodin: A systematic review. J. Clin. Transl. Res. 2017, 3, 283–296. [Google Scholar]
- Dong, X.; Zeng, Y.; Liu, Y.; You, L.; Yin, X.; Fu, J.; Ni, J. Aloe-emodin: A review of its pharmacology, toxicity, and pharmacokinetics. Phytother. Res. 2020, 34, 270–281. [Google Scholar] [CrossRef]
- Huang, P.H.; Huang, C.Y.; Chen, M.C.; Lee, Y.T.; Yue, C.H.; Wang, H.Y.; Lin, H. Emodin and Aloe-Emodin Suppress Breast Cancer Cell Proliferation through ER α Inhibition. Evid. Based Complement. Alternat. Med. 2013, 2013, 376123. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stringaro, A.; Serra, S.; Gori, A.; Calcabrini, A.; Colone, M.; Dupuis, M.L.; Spadaro, F.; Cecchetti, S.; Vitali, A. Peptide-Mediated Targeted Delivery of Aloe-Emodin as Anticancer Drug. Molecules 2022, 27, 4615. [Google Scholar] [CrossRef] [PubMed]
- De Carvalho Lima, E.N.; Barros Martins, G.L.; Diaz, R.S.; Schechter, M.; Piqueira, J.R.C.; Justo, J.F. Effects of Carbon Nanomaterials and Aloe vera on Melanomas-Where Are We? Recent Updates. Pharmaceutics 2022, 14, 2004. [Google Scholar] [CrossRef] [PubMed]
- Radovic, J.; Maksimovic-Ivanic, D.; Timotijevic, G.; Popadic, S.; Ramic, Z.; Trajkovic, V.; Miljkovic, D.; Stosic-Grujicic, S.; Mijatovic, S. Cell-type dependent response of melanoma cells to aloe emodin. Food Chem. Toxicol. 2012, 50, 3181–3189. [Google Scholar] [CrossRef]
- Tabolacci, C.; Lentini, A.; Mattioli, P.; Provenzano, B.; Oliverio, S.; Carlomosti, F.; Beninati, S. Antitumor properties of aloe-emodin and induction of transglutaminase 2 activity in B16-F10 melanoma cells. Life Sci. 2010, 87, 316–324. [Google Scholar] [CrossRef]
- Tabolacci, C.; Cordella, M.; Turcano, L.; Rossi, S.; Lentini, A.; Mariotti, S.; Nisini, R.; Sette, G.; Eramo, A.; Piredda, L.; et al. Aloe-emodin exerts a potent anticancer and immunomodulatory activity on BRAF-mutated human melanoma cells. Eur. J. Pharmacol. 2015, 762, 283–292. [Google Scholar] [CrossRef] [PubMed]
- Lin, S.Y.; Yang, J.H.; Hsia, T.C.; Lee, J.H.; Chiu, T.H.; Wei, Y.H.; Chung, J.G. Effect of inhibition of aloe-emodin on N-acetyltransferase activity and gene expression in human malignant melanoma cells (A375.S2). Melanoma Res. 2005, 15, 489–494. [Google Scholar] [CrossRef]
- Ahluwalia, B.; Magnusson, M.K.; Larsson, F.; Savolainen, O.; Ross, A.B.; Öhman, L. Differences in Metabolite Composition of Aloe barbadensis Mill. Extracts Lead to Differential Effects on Human Blood T Cell Activity In Vitro. Molecules 2022, 27, 6643. [Google Scholar] [CrossRef]
- Budai, M.M.; Varga, A.; Milesz, S.; Tőzsér, J.; Benkő, S. Aloe vera downregulates LPS-induced inflammatory cytokine production and expression of NLRP3 inflammasome in human macrophages. Mol. Immunol. 2013, 56, 471–479. [Google Scholar] [CrossRef] [Green Version]
- Das, A.; Dutta, S. Binding Studies of Aloe-Active Compounds with G-Quadruplex Sequences. ACS Omega 2021, 6, 18344–18351. [Google Scholar] [CrossRef]
- Tanaka, K.; Ishikawa, S.; Matsui, Y.; Tamesada, M.; Harashima, N.; Harada, M. Oral ingestion of Lentinula edodes mycelia extract inhibits B16 melanoma growth via mitigation of regulatory T cell-mediated immunosuppression. Cancer Sci. 2011, 102, 516–521. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.B.; Yamagishi, C.; Hayashi, K.; Hayashi, T. Antiviral and immunostimulating effects of lignin-carbohydrate-protein complexes from Pimpinella anisum. Biosci. Biotechnol. Biochem. 2011, 75, 459–465. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sakagami, H.; Kushida, T.; Oizumi, T.; Nakashima, H.; Makino, T. Distribution of lignin-carbohydrate complex in plant kingdom and its functionality as alternative medicine. Pharmacol Ther. 2010, 128, 91–105. [Google Scholar] [CrossRef] [PubMed]
- De Silva, S.F.; Alcorn, J. Flaxseed Lignans as Important Dietary Polyphenols for Cancer Prevention and Treatment: Chemistry, Pharmacokinetics, and Molecular Targets. Pharmaceuticals 2019, 12, 68. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Zhang, Z.; Zhang, K.; Xue, Z.; Li, Y.; Zhang, Z.; Zhang, L.; Gu, C.; Zhang, Q.; Hao, J.; et al. Arctigenin Suppress Th17 Cells and Ameliorates Experimental Autoimmune Encephalomyelitis Through AMPK and PPAR-γ/ROR-γt Signaling. Mol. Neurobiol. 2016, 53, 5356–5366. [Google Scholar] [CrossRef]
- Cho, J.Y.; Park, J.; Kim, P.S.; Yoo, E.S.; Baik, K.U.; Park, M.H. Savinin, a lignan from Pterocarpus santalinus inhibits tumor necrosis factor-alpha production and T cell proliferation. Biol. Pharm. Bull. 2001, 24, 167–171. [Google Scholar] [CrossRef] [Green Version]
- Kortesoja, M.; Karhu, E.; Olafsdottir, E.S.; Freysdottir, J.; Hanski, L. Impact of dibenzocyclooctadiene lignans from Schisandra chinensis on the redox status and activation of human innate immune system cells. Free Radic. Biol. Med. 2019, 131, 309–317. [Google Scholar] [CrossRef]
- Ma, Z.; Xu, G.; Shen, Y.; Hu, S.; Lin, X.; Zhou, J.; Zhao, W.; Liu, J.; Wang, J.; Guo, J. Schisandrin B-mediated TH17 cell differentiation attenuates bowel inflammation. Pharmacol. Res. 2021, 166, 105459. [Google Scholar] [CrossRef]
- Li, X.; Gao, J.; Yu, Z.; Jiang, W.; Sun, W.; Yu, C.; Sun, J.; Wang, C.; Chen, J.; Jing, S.; et al. Regulatory Effect of Anwulignan on the Immune Function Through Its Antioxidation and Anti-Apoptosis in D-Galactose-Induced Aging Mice. Clin. Interv. Aging 2020, 15, 97–110. [Google Scholar] [CrossRef] [Green Version]
- Shimoyoshi, S.; Takemoto, D.; Kishimoto, Y.; Amano, A.; Sato, A.; Ono, Y.; Rogi, T.; Shibata, H.; Ishigami, A. Sesame lignans suppress age-related disorders of the kidney in mice. Eur. Rev. Med. Pharmacol. Sci. 2020, 24, 5140–5147. [Google Scholar] [CrossRef]
- Majdalawieh, A.F.; Yousef, S.M.; Abu-Yousef, I.A.; Nasrallah, G.K. Immunomodulatory and anti-inflammatory effects of sesamin: Mechanisms of action and future directions. Crit. Rev. Food Sci. Nutr. 2022, 62, 5081–5112. [Google Scholar] [CrossRef] [PubMed]
- Cox-Georgian, D.; Ramadoss, N.; Dona, C.; Basu, C. Therapeutic and medicinal uses of terpenes. In Medicinal Plants: From Farm to Pharmacy; Springer: Berlin/Heidelberg, Germany, 2019. [Google Scholar]
- Kim, T.; Song, B.; Cho, K.S.; Lee, I.S. Therapeutic Potential of Volatile Terpenes and Terpenoids from Forests for Inflammatory Diseases. Int. J. Mol. Sci. 2020, 21, 2187. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kamran, S.; Sinniah, A.; Abdulghani, M.A.M.; Alshawsh, M.A. Therapeutic Potential of Certain Terpenoids as Anticancer Agents: A Scoping Review. Cancers 2022, 14, 1100. [Google Scholar] [CrossRef] [PubMed]
- Richard, S.A. Exploring the Pivotal Immunomodulatory and Anti-Inflammatory Potentials of Glycyrrhizic and Glycyrrhetinic Acids. Mediat. Inflamm. 2021, 2021, 6699560. [Google Scholar] [CrossRef] [PubMed]
- Raphael, T.J.; Kuttan, G. Effect of naturally occurring triterpenoids glycyrrhizic acid, ursolic acid, oleanolic acid and nomilin on the immune system. Phytomedicine 2003, 10, 483–489. [Google Scholar] [CrossRef] [Green Version]
- Raphael, T.J.; Kuttan, G. Effect of naturally occurring triterpenoids ursolic acid and glycyrrhizic acid on the cell-mediated immune responses of metastatic tumor-bearing animals. Immunopharmacol. Immunotoxicol. 2008, 30, 243–255. [Google Scholar] [CrossRef]
- Jeong, H.G.; Kim, Y.K. Induction of inducible nitric oxide synthase expression by 18beta-glycyrrhetinic acid in macrophages. FEBS Lett. 2002, 513, 208–212. [Google Scholar] [CrossRef] [Green Version]
- Abe, N.; Ebina, T.; Ishida, N. Interferon induction by glycyrrhizin and glycyrrhetinic acid in mice. Microbiol. Immunol. 1982, 26, 535–539. [Google Scholar] [CrossRef] [Green Version]
- Mlala, S.; Oyedeji, A.O.; Gondwe, M.; Oyedeji, O.O. Ursolic Acid and Its Derivatives as Bioactive Agents. Molecules 2019, 24, 2751. [Google Scholar] [CrossRef] [Green Version]
- Manu, K.A.; Kuttan, G. Ursolic acid induces apoptosis by activating p53 and caspase-3 gene expressions and suppressing NF-kappaB mediated activation of bcl-2 in B16F-10 melanoma cells. Int. Immunopharmacol. 2008, 8, 974–981. [Google Scholar] [CrossRef]
- Pratheeshkumar, P.; Kuttan, G. Nomilin inhibits metastasis via induction of apoptosis and regulates the activation of transcription factors and cytokine profile in B16F-10 cells. Integr. Cancer Ther. 2012, 11, 48–60. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rajagopal, S.; Kumar, R.A.; Deevi, D.S.; Satyanarayana, C.; Rajagopalan, R. Andrographolide, a potential cancer therapeutic agent isolated from Andrographis paniculata. J. Exp. Ther. Oncol. 2003, 3, 147–158. [Google Scholar] [CrossRef] [PubMed]
- Sun, J. D-Limonene: Safety and clinical applications. Altern. Med. Rev. 2007, 12, 259–264. [Google Scholar] [PubMed]
- Kuttan, G.; Pratheeshkumar, P.; Manu, K.A.; Kuttan, R. Inhibition of tumor progression by naturally occurring terpenoids. Pharm. Biol. 2011, 49, 995–1007. [Google Scholar] [CrossRef]
- Pratheeshkumar, P.; Kuttan, G. Vernolide-A, a sesquiterpene lactone from Vernonia cinerea, induces apoptosis in B16F-10 melanoma cells by modulating p53 and caspase-3 gene expressions and regulating NF-κB-mediated bcl-2 activation. Drug. Chem. Toxicol. 2011, 34, 261–270. [Google Scholar] [CrossRef]
- Bozzuto, G.; Colone, M.; Toccacieli, L.; Stringaro, A.; Molinari, A. Tea tree oil might combat melanoma. Planta Med. 2011, 77, 54–56. [Google Scholar] [CrossRef]
- Calcabrini, A.; Stringaro, A.; Toccacieli, L.; Meschini, S.; Marra, M.; Colone, M.; Salvatore, G.; Mondello, F.; Arancia, G.; Molinari, A. Terpinen-4-ol, the main component of Melaleuca alternifolia (tea tree) oil inhibits the in vitro growth of human melanoma cells. J. Investig. Dermatol. 2004, 122, 349–360. [Google Scholar] [CrossRef] [Green Version]
- Di Martile, M.; Garzoli, S.; Sabatino, M.; Valentini, E.; D’Aguanno, S.; Ragno, R.; Del Bufalo, D. Antitumor effect of Melaleuca alternifolia essential oil and its main component terpinen-4-ol in combination with target therapy in melanoma models. Cell Death Discov. 2021, 7, 127. [Google Scholar] [CrossRef]
- Bozzuto, G.; Mariano, F.; Costa, I.; Calcabrini, A.; Molinari, A. Tea Tree Oil and Terpinen-4-Ol Induce Cytoskeletal Reorganization of Human Melanoma Cells. Planta Med. Int. Open 2021, 8, e34–e53. [Google Scholar] [CrossRef]
- Aslam, S.; Younis, W.; Malik, M.N.H.; Jahan, S.; Alamgeer Uttra, A.M.; Munir, M.U.; Roman, M. Pharmacological evaluation of anti-arthritic potential of terpinen-4-ol using in vitro and in vivo assays. Inflammopharmacology 2022, 30, 945–959. [Google Scholar] [CrossRef]
- Hart, P.H.; Brand, C.; Carson, C.F.; Riley, T.V.; Prager, R.H.; Finlay-Jones, J.J. Terpinen-4-ol, the main component of the essential oil of Melaleuca alternifolia (tea tree oil), suppresses inflammatory mediator production by activated human monocytes. Inflamm. Res. 2000, 49, 619–626. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.; Zhou, Q.H.; Xu, K. Are isothiocyanates potential anti-cancer drugs? Acta Pharmacol. Sin. 2009, 30, 501–512. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lawson, A.P.; Long, M.J.C.; Coffey, R.T.; Qian, Y.; Weerapana, E.; El Oualid, F.; Hedstrom, L. Naturally Occurring Isothiocyanates Exert Anticancer Effects by Inhibiting Deubiquitinating Enzymes. Cancer Res. 2015, 75, 5130–5142. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mitsiogianni, M.; Koutsidis, G.; Mavroudis, N.; Trafalis, D.T.; Botaitis, S.; Franco, R.; Zoumpourlis, V.; Amery, T.; Galanis, A.; Pappa, A.; et al. The Role of Isothiocyanates as Cancer Chemo-Preventive, Chemo-Therapeutic and Anti-Melanoma Agents. Antioxidants 2019, 8, 106. [Google Scholar] [CrossRef]
- Sundaram, M.K.; Preetha, R.; Haque, S.; Akhter, N.; Khan, S.; Ahmad, S.; Hussain, A. Dietary isothiocyanates inhibit cancer progression by modulation of epigenome. Semin. Cancer Biol. 2022, 83, 353–376. [Google Scholar] [CrossRef]
- Hamsa, T.P.; Thejass, P.; Kuttan, G. Induction of apoptosis by sulforaphane in highly metastatic B16F-10 melanoma cells. Drug Chem. Toxicol. 2011, 34, 332–340. [Google Scholar] [CrossRef]
- Pradhan, S.J.; Mishra, R.; Sharma, P.; Kundu, G.C. Quercetin and sulforaphane in combination suppress the progression of melanoma through the down-regulation of matrix metalloproteinase-9. Exp. Ther. Med. 2010, 1, 915–920. [Google Scholar] [CrossRef] [Green Version]
- Eom, Y.S.; Shah, F.H.; Kim, S.J. Sulforaphane induces cell differentiation, melanogenesis and also inhibit the proliferation of melanoma cells. Eur. J. Pharmacol. 2022, 921, 174894. [Google Scholar] [CrossRef]
- Lee, J.J.; Murphy, G.F.; Lian, C.G. Melanoma epigenetics: Novel mechanisms, markers, and medicines. Lab. Investig. 2014, 94, 822–838. [Google Scholar] [CrossRef] [Green Version]
- Su, Z.Y.; Zhang, C.; Lee, J.H.; Shu, L.; Wu, T.Y.; Khor, T.O.; Conney, A.H.; Lu, Y.P.; Kong, A.N. Requirement and epigenetics reprogramming of Nrf2 in suppression of tumor promoter TPA-induced mouse skin cell transformation by sulforaphane. Cancer Prev. Res. 2014, 7, 319–329. [Google Scholar] [CrossRef] [Green Version]
- Kuo, H.D.; Wu, R.; Li, S.; Yang, A.Y.; Kong, A.N. Anthocyanin Delphinidin Prevents Neoplastic Transformation of Mouse Skin JB6 P+ Cells: Epigenetic Re-activation of Nrf2-ARE Pathway. AAPS J. 2019, 21, 83. [Google Scholar] [CrossRef] [PubMed]
- Dickinson, S.E.; Rusche, J.J.; Bec, S.L.; Horn, D.J.; Janda, J.; Rim, S.H.; Smith, C.L.; Bowden, G.T. The effect of sulforaphane on histone deacetylase activity in keratinocytes: Differences between in vitro and in vivo analyses. Mol. Carcinog. 2015, 54, 1513–1520. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Patya, M.; Zahalka, M.A.; Vanichkin, A.; Rabinkov, A.; Miron, T.; Mirelman, D.; Wilchek, M.; Harry, M.; Lander, H.M.; Novogrodsky, A. Allicin stimulates lymphocytes and elicits an antitumor effect: A possible role of p21ras. Int. Immunol. 2004, 16, 275–281. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jobani, B.M.; Najafzadeh, N.; Mazani, M.; Arzanlou, M.; Vardin, M.M. Molecular mechanism and cytotoxicity of allicin and all-trans retinoic acid against CD44+ versus CD117+ melanoma cells. Phytomedicine 2018, 48, 161–169. [Google Scholar] [CrossRef]
- Wang, H.C.; Pao, J.; Lin, S.Y.; Sheen, L.Y. Molecular mechanisms of garlic-derived allyl sulfides in the inhibition of skin cancer progression. Ann. N. Y. Acad. Sci. 2012, 1271, 44–52. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Fu, R.; Tu, S.; Qu, D.; Liu, C.; Li, X.; Zhao, Y.; Ma, C.; Chen, Y.; Li Tao, L. Extracellular Microparticles Encapsulated with Diallyl Trisulfide Interfere with the Inflammatory Tumor Microenvironment and Lung Metastasis of Invasive Melanoma. Mol. Pharm. 2021, 18, 822–835. [Google Scholar] [CrossRef]
- Nakagawa, C.; Suzuki-Karasaki, M.; Suzuki-Karasaki, M.; Ochiai, T.; Suzuki-Karasaki, Y. The Mitochondrial Ca2+ Overload via Voltage-Gated Ca2+ Entry Contributes to an Anti-Melanoma Effect of Diallyl Trisulfide. Int. J. Mol. Sci. 2020, 21, 491. [Google Scholar] [CrossRef] [Green Version]
- Hung, F.M.; Shang, H.S.; Tang, N.Y.; Lin, J.J.; Lu, K.W.; Lin, J.P.; Ko, Y.C.; Yu, C.C.; Wang, H.L.; Liao, J.C.; et al. Effects of diallyl trisulfide on induction of apoptotic death in murine leukemia WEHI-3 cells in vitro and alterations of the immune responses in normal and leukemic mice in vivo. Environ. Toxicol. 2015, 30, 1343–1353. [Google Scholar] [CrossRef]
- Tudu, C.K.; Dutta, T.; Ghorai, M.; Biswas, P.; Samanta, D.; Oleksak, P.; Jha, N.K.; Kumar, M.; Radha; Proćków, J.; et al. Traditional uses, phytochemistry, pharmacology and toxicology of garlic (Allium sativum), a storehouse of diverse phytochemicals: A review of research from the last decade focusing on health and nutritional implications. Rev. Front. Nutr. 2022, 9, 949554. [Google Scholar] [CrossRef]
- Saleh, J. Murine models of melanoma. Pathol. Res. Pract. 2018, 214, 1235–1238. [Google Scholar] [CrossRef]
- Micucci, M.; Bolchi, C.; Budriesi, R.; Cevenini, M.; Maroni, L.; Capozza, S.; Chiarini, A.; Pallavicini, M.; Angeletti, A. Antihypertensive phytocomplexes of proven efficacy and well-established use: Mode of action and individual characterization of the active constituents. Phytochemistry 2020, 170, 112222. [Google Scholar] [CrossRef] [PubMed]
- Rahman, M.M.; Rahaman, M.S.; Islam, M.R.; Hossain, M.E.; Mannan Mithi, F.; Ahmed, M.; Saldías, M.; Akkol, E.K.; Sobarzo-Sánchez, E. Multifunctional Therapeutic Potential of Phytocomplexes and Natural Extracts for Antimicrobial Properties. Antibiotics 2021, 10, 1076. [Google Scholar] [CrossRef]
- Wojtunik-Kulesza, K.; Oniszczuk, A.; Oniszczuk, T.; Combrzyński, M.; Nowakowska, D.; Matwijczuk, A. Influence of In Vitro Digestion on Composition, Bioaccessibility and Antioxidant Activity of Food Polyphenols-A Non-Systematic Review. Nutrients 2020, 12, 1401. [Google Scholar] [CrossRef] [PubMed]
- Bertelli, A.; Biagi, M.; Corsini, M.; Baini, G.; Cappellucci, G.; Miraldi, E. Polyphenols: From Theory to Practice. Foods 2021, 10, 2595. [Google Scholar] [CrossRef] [PubMed]
- Yin, S.Y.; Yang, N.S.; Lin, T.J. Phytochemicals Approach for Developing Cancer Immunotherapeutics. Front. Pharmacol. 2017, 8, 386. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ali Reza, A.S.M.; Nasrin, M.S.; Hossen, M.A.; Rahman, M.A.; Jantan, I.; Haque, M.A.; Sobarzo-Sánchez, E. Mechanistic insight into immunomodulatory effects of food-functioned plant secondary metabolites. Crit. Rev. Food Sci. Nutr. 2021, 27, 1–31. [Google Scholar] [CrossRef] [PubMed]
- Maheshwari, S.; Kumar, V.; Bhadauria, G.; Mishra, A. Immunomodulatory potential of phytochemicals and other bioactive compounds of fruits: A review. Food Front. 2022, 3, 221–238. [Google Scholar] [CrossRef]
- Plundrich, D.; Chikhladze, S.; Fichtner-Feigl, S.; Feuerstein, R.; Briquez, P.S. Molecular Mechanisms of Tumor Immunomodulation in the Microenvironment of Colorectal Cancer. Int. J. Mol. Sci. 2022, 23, 2782. [Google Scholar] [CrossRef]
- Merecz-Sadowska, A.; Sitarek, P.; Stelmach, J.; Zajdel, K.; Kucharska, E.; Zajdel, R. Plants as Modulators of Melanogenesis: Role of Extracts, Pure Compounds and Patented Compositions in Therapy of Pigmentation Disorders. Int. J. Mol. Sci. 2022, 23, 14787. [Google Scholar] [CrossRef]










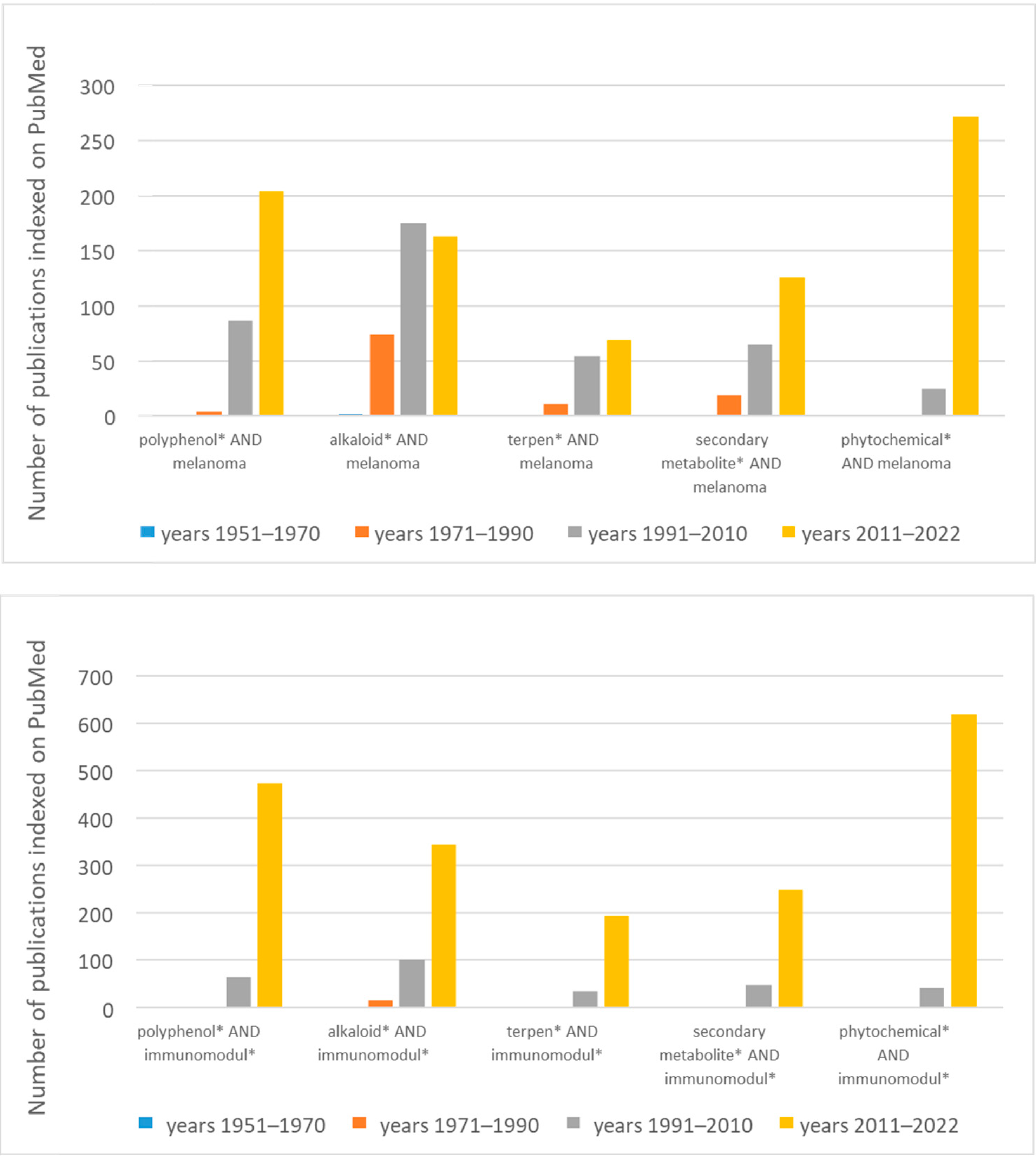{kind=link}
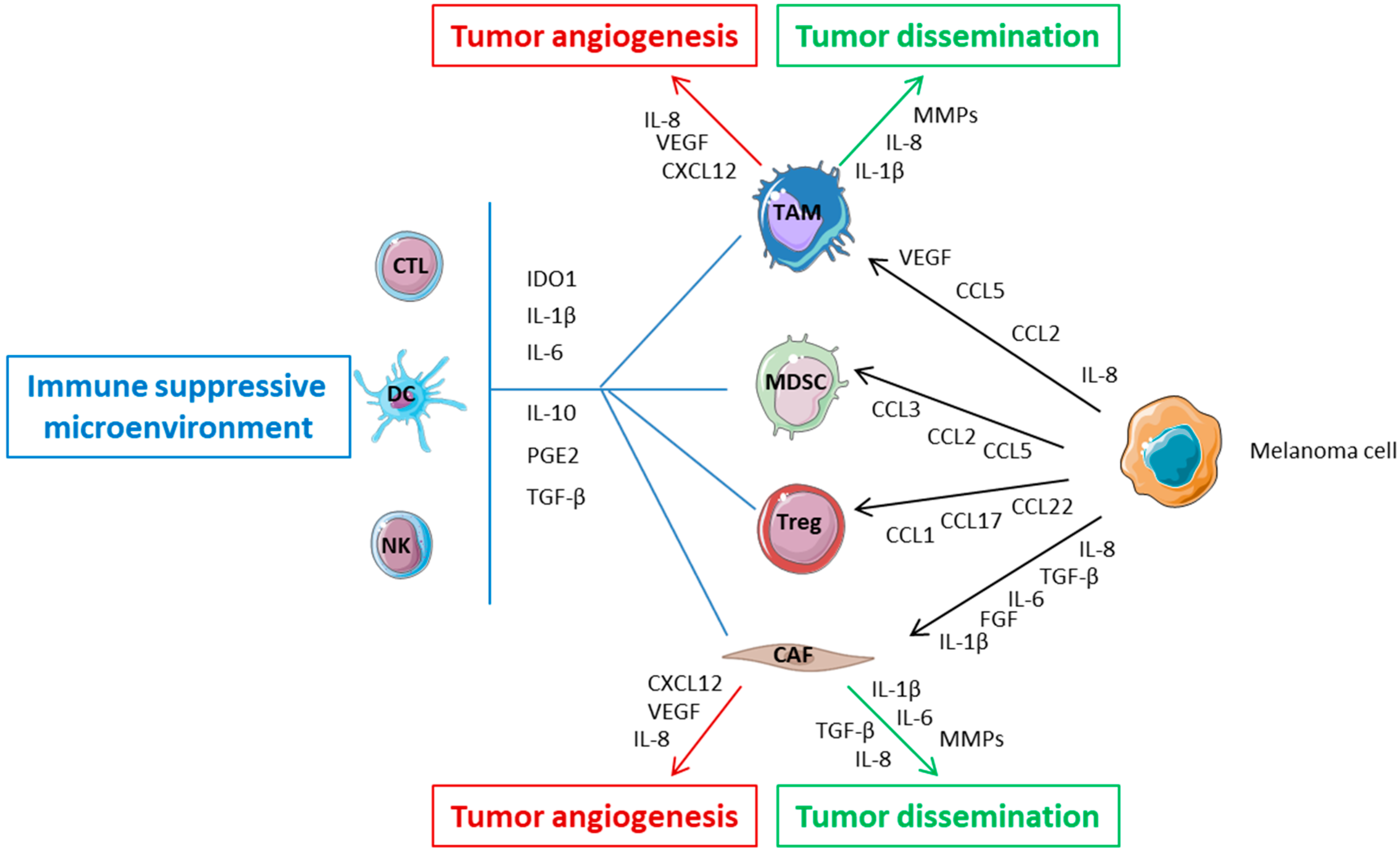{kind=link}
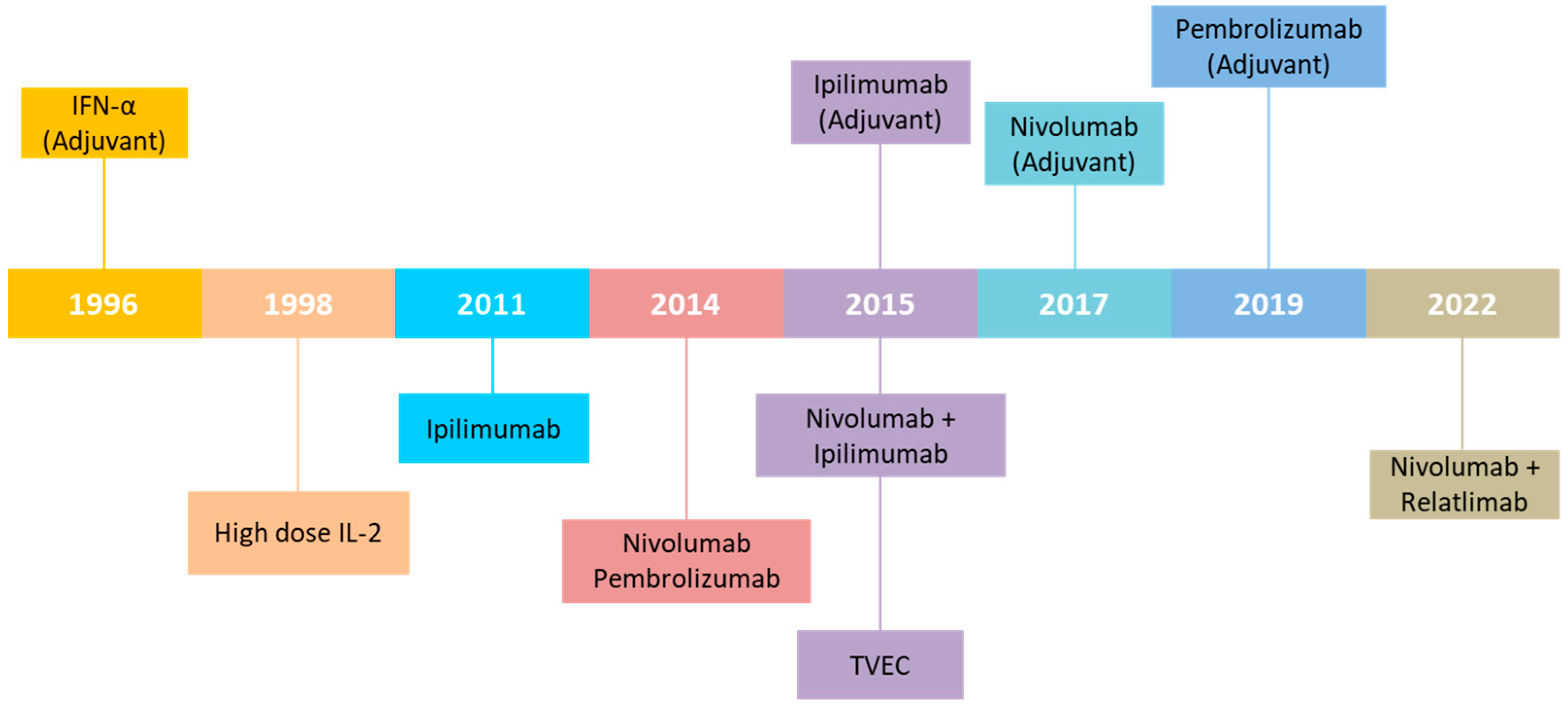{kind=link}
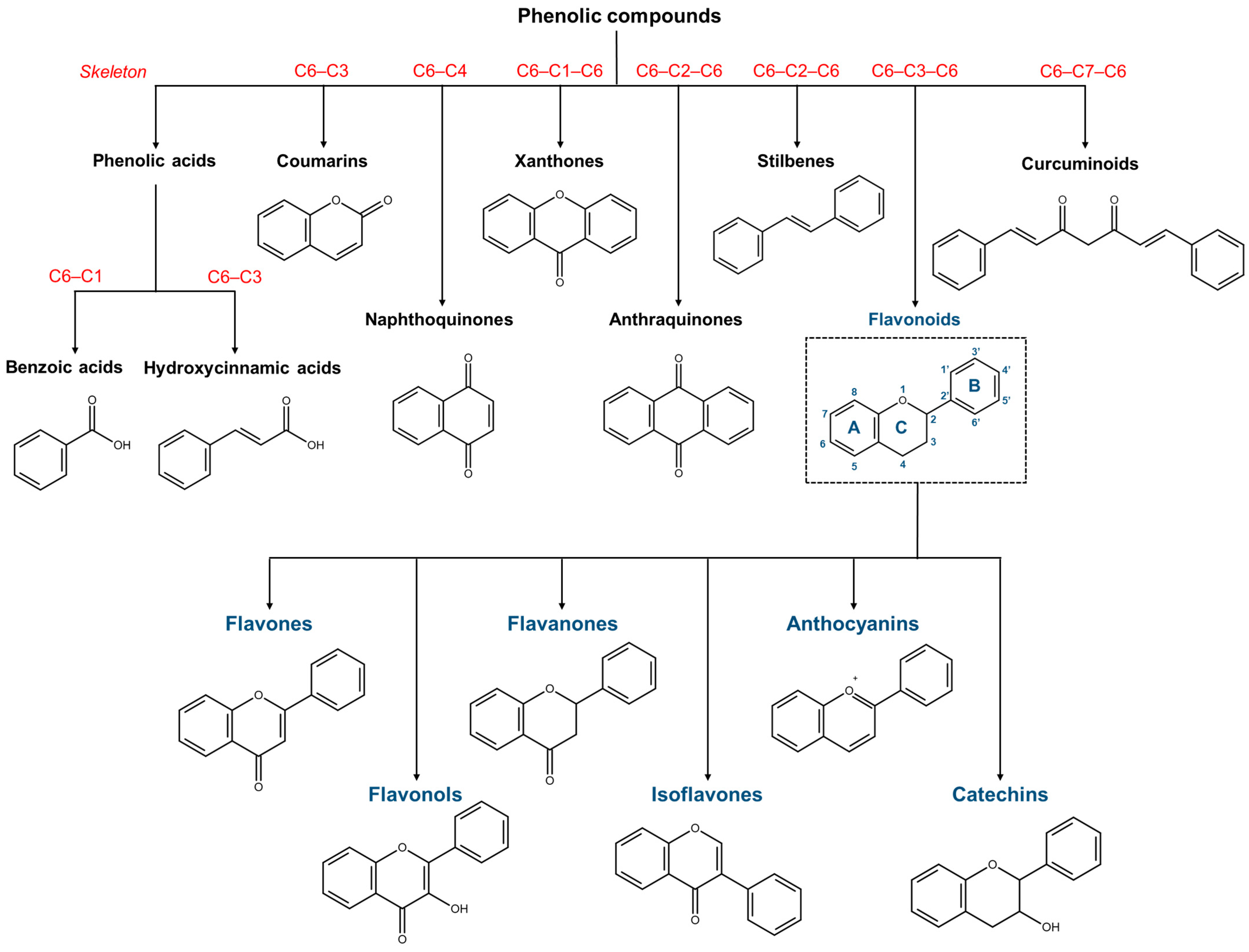{kind=link}
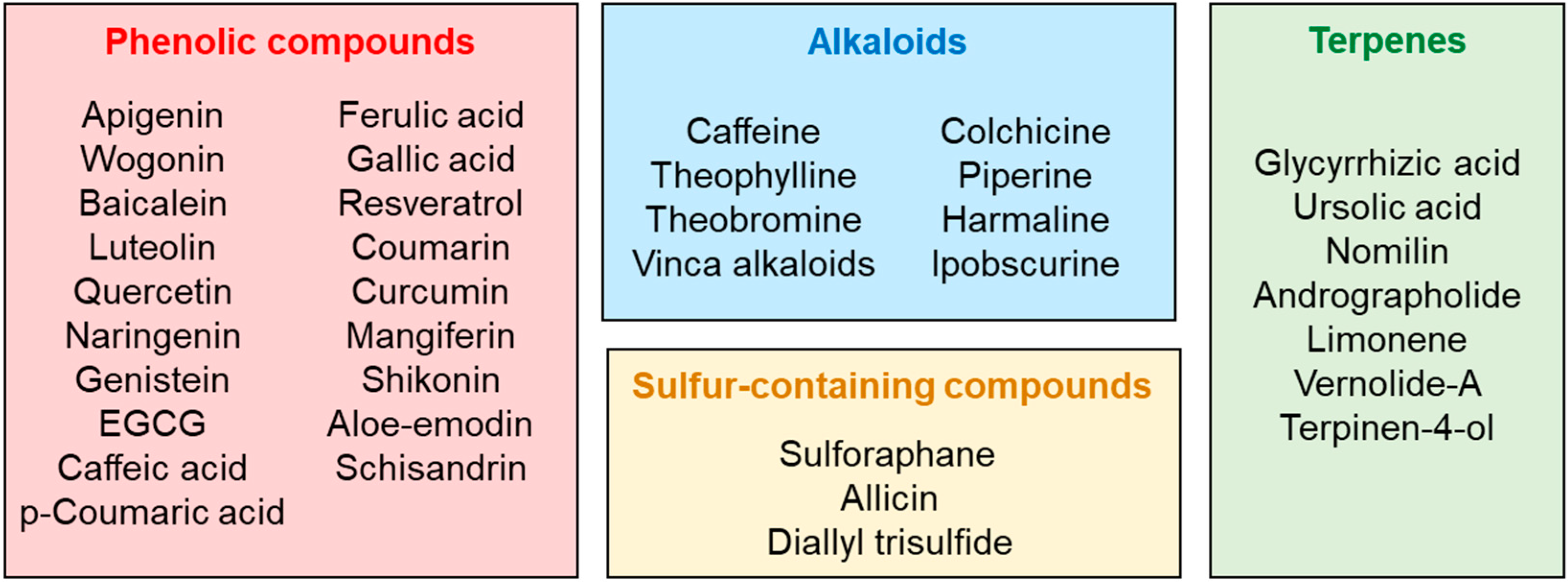{kind=link}
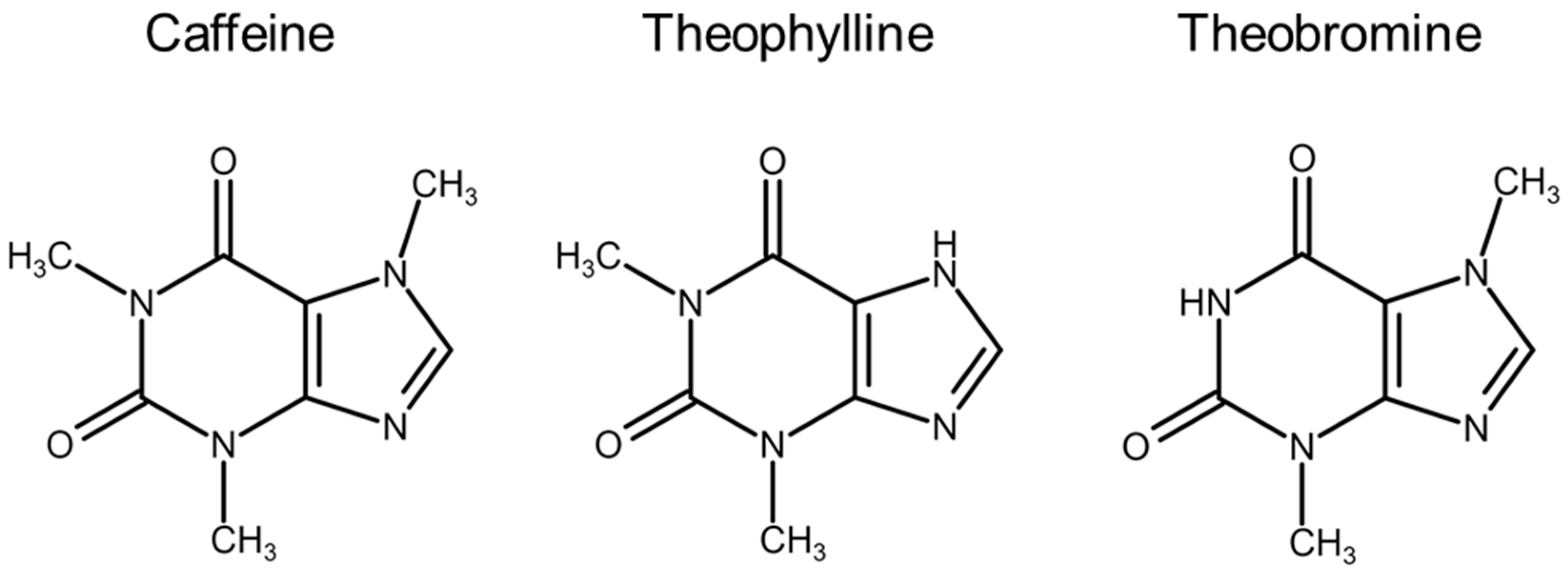{kind=link}
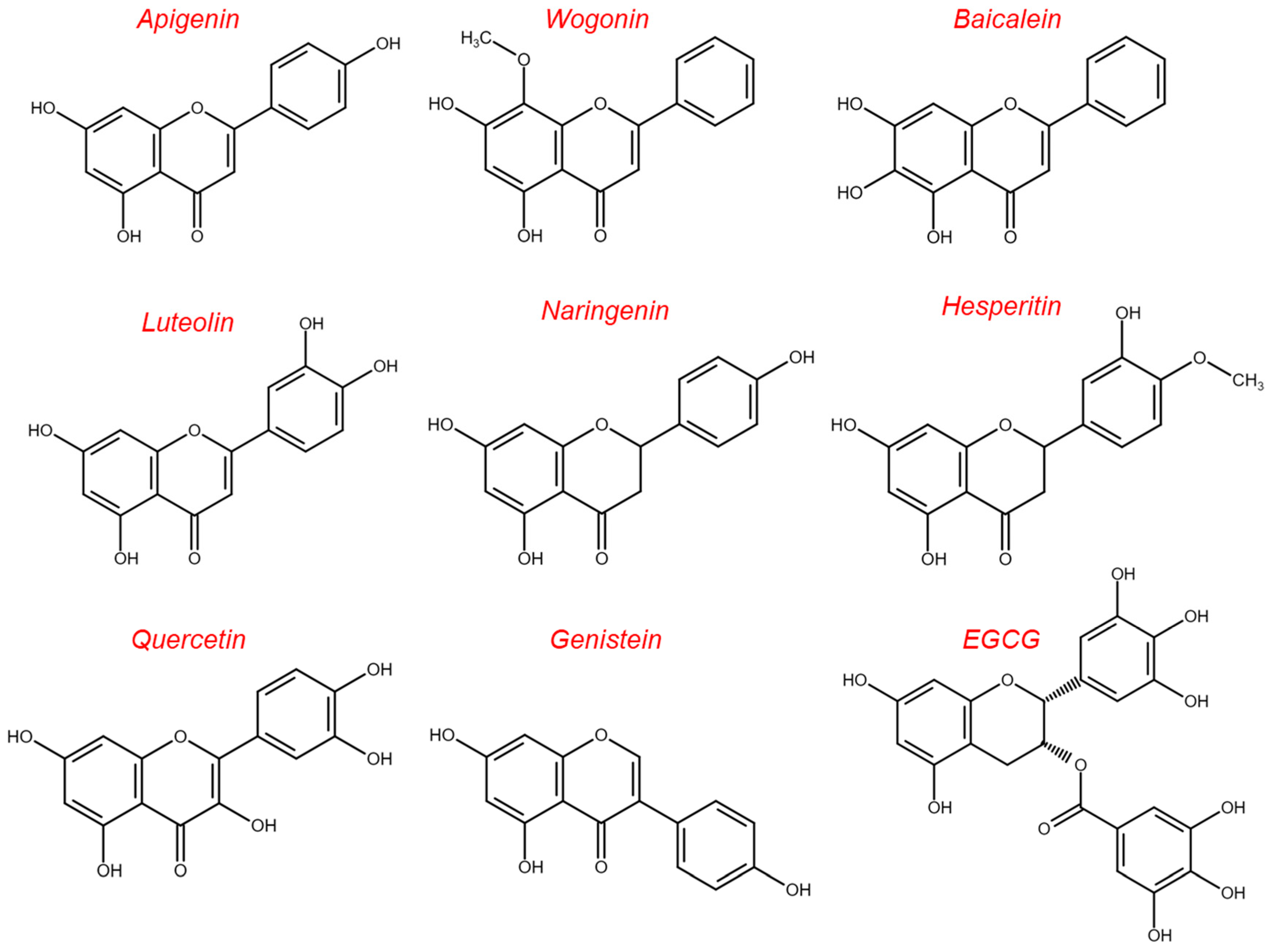{kind=link}
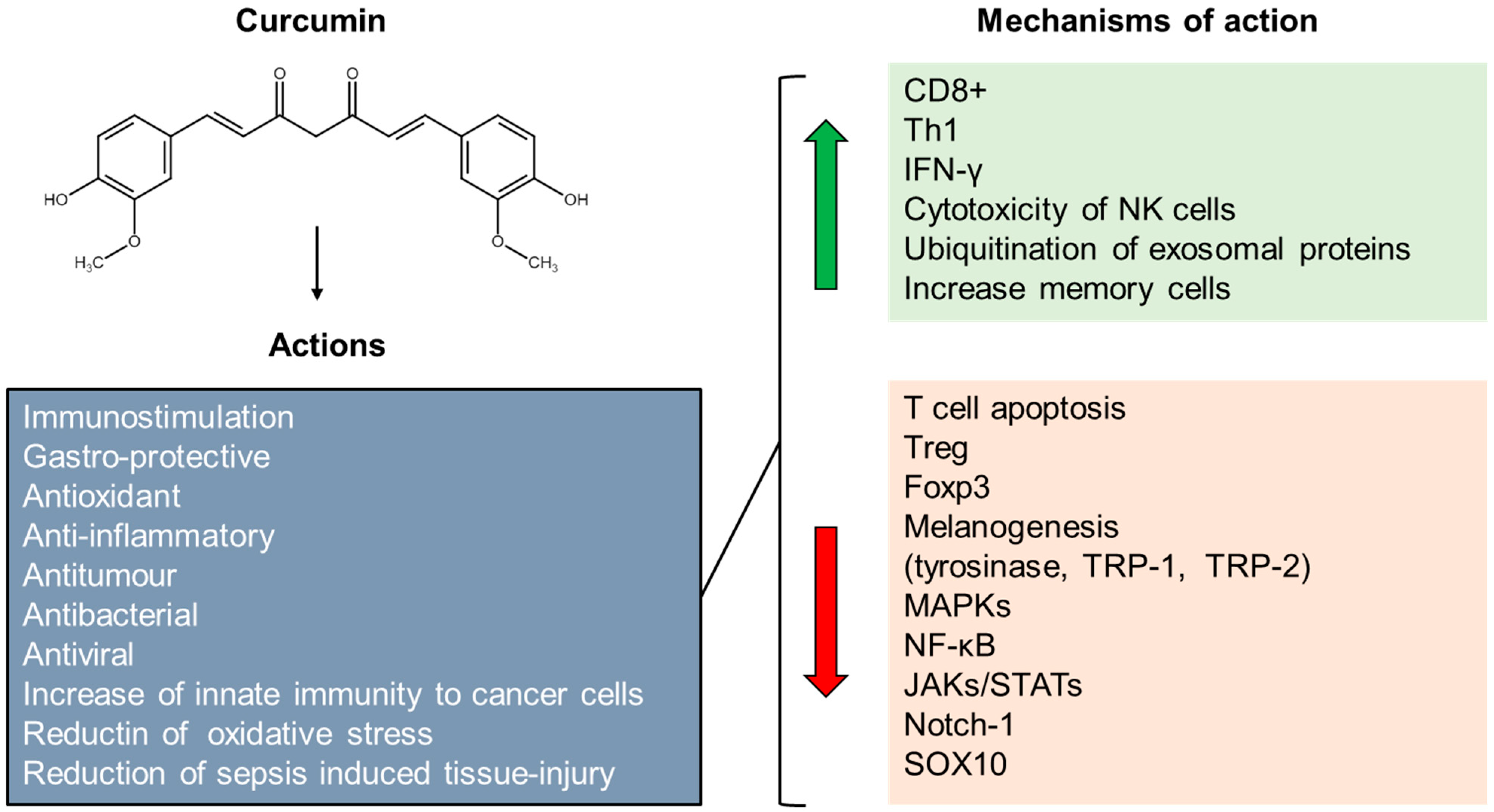{kind=link}
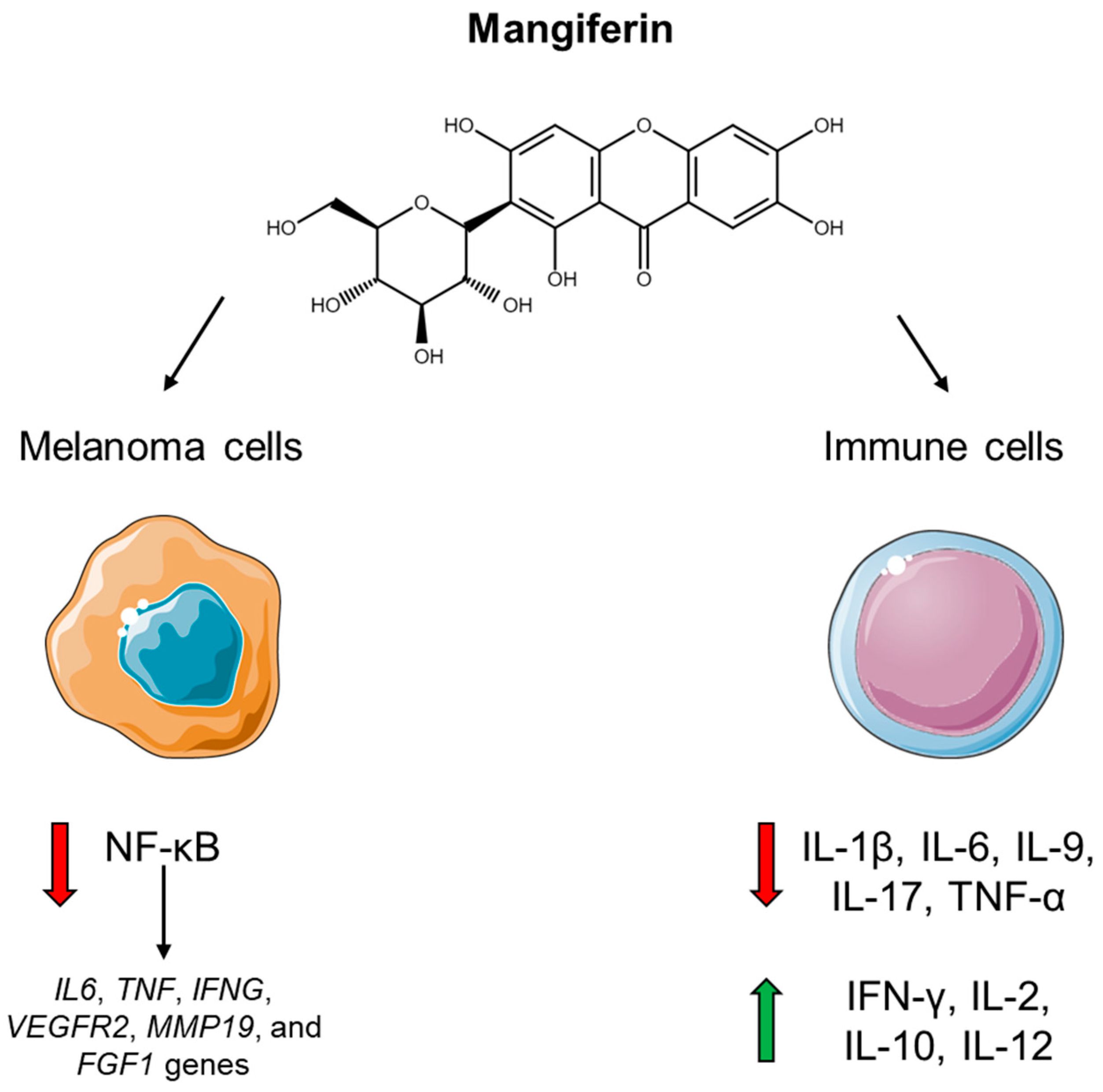{kind=link}
| Secondary Metabolites | Extraction Method | References |
|---|---|---|
| Alkaloids | Extraction using a “acid–base shakeout” (only for basic alkaloids) | [101] |
| Extraction using alcoholic solvents (methanol or ethanol) or apolar solvents (e.g., chloroform) at room temperature or by refluxing, with the main drawbacks to extract also not alkaloid compounds and to use harmful solvents. | ||
| Extraction by supercritical CO2 (SF-CO2) | [102] | |
| Extraction by ionic liquids | [103] | |
| Extraction by natural deep eutectic solvents | [104] | |
| Extraction by microwave-assisted extractions | [105] | |
| Phenolic compounds | Conventional methods, i.e., solid-liquid extractions, can be used. The recovery of phenolics from plant matrices is strictly influenced by solvent, extraction time and temperature, all factors which may affect extraction efficiency due the low stability at high temperature and in alkaline medium where phenols easily oxidize | [106] |
| Alternative methods requiring ultrasounds, microwaves, supercritical fluids, pressurized liquids, or enzymes can be applied | [84] | |
| Terpenes | Considering their apolar character, terpenes are traditionally extracted by apolar solvents, such as chloroform and n-hexane. | |
| Extraction by SF-CO2 | [107,108] | |
| Volatiles compounds can be also extracted by hydro distillation or steam distillation | [109] | |
| Glucosinolates | The enzymatic degration of glucosinolates must be taken into consideration during the extraction, where myrosinase denaturation is a crucial step. Therefore, the most common procedure to isolate glucosinolates from plant consists of extraction in 70% methanol for 10 min at 75 °C or boiling water in order to denaturate the enzyme | [110] |
| Thiosulfinates | Allicin extraction from garlic can be carried out by solvent extraction, which requires long time extraction. Alternatively, SF-CO2, Ultrasonic-Assisted Extraction (UAE) or Pressurized Liquid Extraction (PLE) can be used although they requires two separated processes, the enzymatic and extraction ones. Therefore, subcritical water extraction (SWE) is a good option in order to have in a single-step the enzymatic and extraction process in a close system | [111] |
| Inflammatory Mediator | Secondary Metabolites | References |
|---|---|---|
| GM-CSF | Ipobscurine | [148] |
| Aloe-emodin | [253] | |
| Glycyrrhizic acid | [274] | |
| Ursolic acid | [274] | |
| Nomilin | [279] | |
| Andrographolide | [280] | |
| Vernolide-A | [283] | |
| Sulforaphane | [294] | |
| IFN-α | Quercetin | [163] |
| IFN-β | Quercetin | [163] |
| IFN-γ | Aloe-emodin | [253] |
| IL-1β | Theophylline | [126] |
| Piperine | [140] | |
| Ipobscurine | [148] | |
| EGCG | [172,173] | |
| Nomilin | [279] | |
| Andrographolide | [280] | |
| Vernolide-A | [283] | |
| Sulforaphane | [294] | |
| IL-2 | Ipobscurine | [148] |
| Aloe-emodin | [253] | |
| Glycyrrhizic acid | [274] | |
| Ursolic acid | [274] | |
| IL-6 | Piperine | [140] |
| Ipobscurine | [148] | |
| Naringenin | [166] | |
| Glabridin | [183] | |
| Glycyrrhizic acid | [274] | |
| Ursolic acid | [274] | |
| Nomilin | [279] | |
| Andrographolide | [280] | |
| Vernolide-A | [283] | |
| Sulforaphane | [294] | |
| IL-10 | Glabridin | [183] |
| Caffeic acid | [201] | |
| IL-12 | Theophylline | [126] |
| Aloe-emodin | [253] | |
| Sulforaphane | [294] | |
| IL-17 | Resveratrol | [217] |
| IP-10 | Caffeine | [125] |
| Theophylline | [126] | |
| MCP-1 | Theophylline | [126] |
| MIP-1α | Caffeine | [125] |
| Theophylline | [126] | |
| MIP-1β | Caffeine | [125] |
| Theophylline | [126] | |
| RANTES | Caffeine | [125] |
| Theophylline | [126] | |
| Shikonin | [243] | |
| TNF-α | Methylxanthines | [117] |
| Piperine | [140] | |
| Ipobscurine | [148] | |
| Glabridin | [183] | |
| Shikonin | [243] | |
| Nomilin | [279] | |
| Andrographolide | [280] | |
| Vernolide-A | [283] | |
| Sulforaphane | [294] | |
| VEGF | Theophylline | [126] |
| Ipobscurine | [148] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tabolacci, C.; De Vita, D.; Facchiano, A.; Bozzuto, G.; Beninati, S.; Failla, C.M.; Di Martile, M.; Lintas, C.; Mischiati, C.; Stringaro, A.; et al. Phytochemicals as Immunomodulatory Agents in Melanoma. Int. J. Mol. Sci. 2023, 24, 2657. https://doi.org/10.3390/ijms24032657
Tabolacci C, De Vita D, Facchiano A, Bozzuto G, Beninati S, Failla CM, Di Martile M, Lintas C, Mischiati C, Stringaro A, et al. Phytochemicals as Immunomodulatory Agents in Melanoma. International Journal of Molecular Sciences. 2023; 24(3):2657. https://doi.org/10.3390/ijms24032657
Chicago/Turabian StyleTabolacci, Claudio, Daniela De Vita, Antonio Facchiano, Giuseppina Bozzuto, Simone Beninati, Cristina Maria Failla, Marta Di Martile, Carla Lintas, Carlo Mischiati, Annarita Stringaro, and et al. 2023. "Phytochemicals as Immunomodulatory Agents in Melanoma" International Journal of Molecular Sciences 24, no. 3: 2657. https://doi.org/10.3390/ijms24032657