
Semi-Targeted Profiling of Bile Acids by High-Resolution Mass Spectrometry in a Rat Model of Drug-Induced Liver Injury


Abstract
:1. Introduction
2. Results
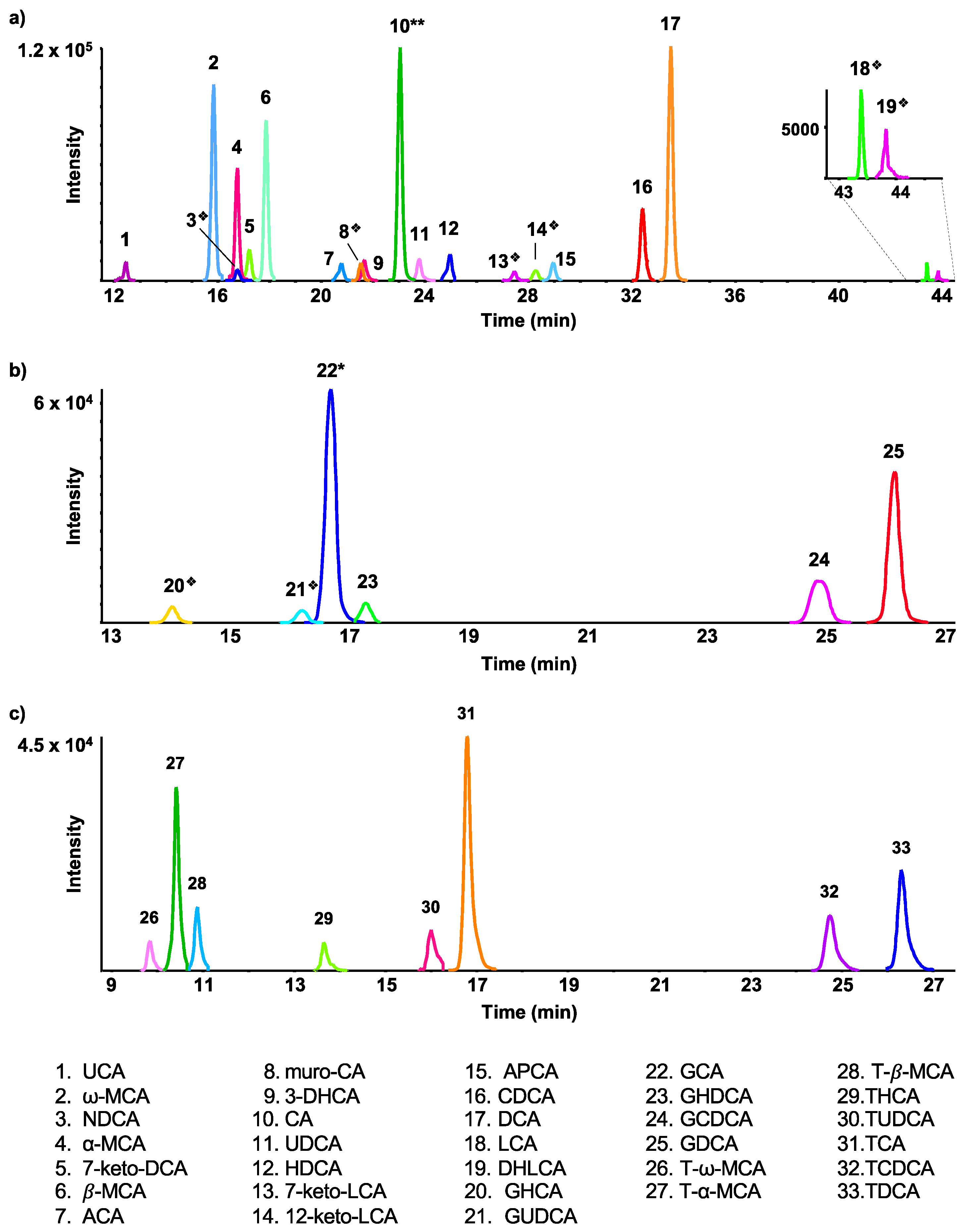
2.1. Assigned Bile Acids
2.2. Putatively Identified Bile Acid Isomers
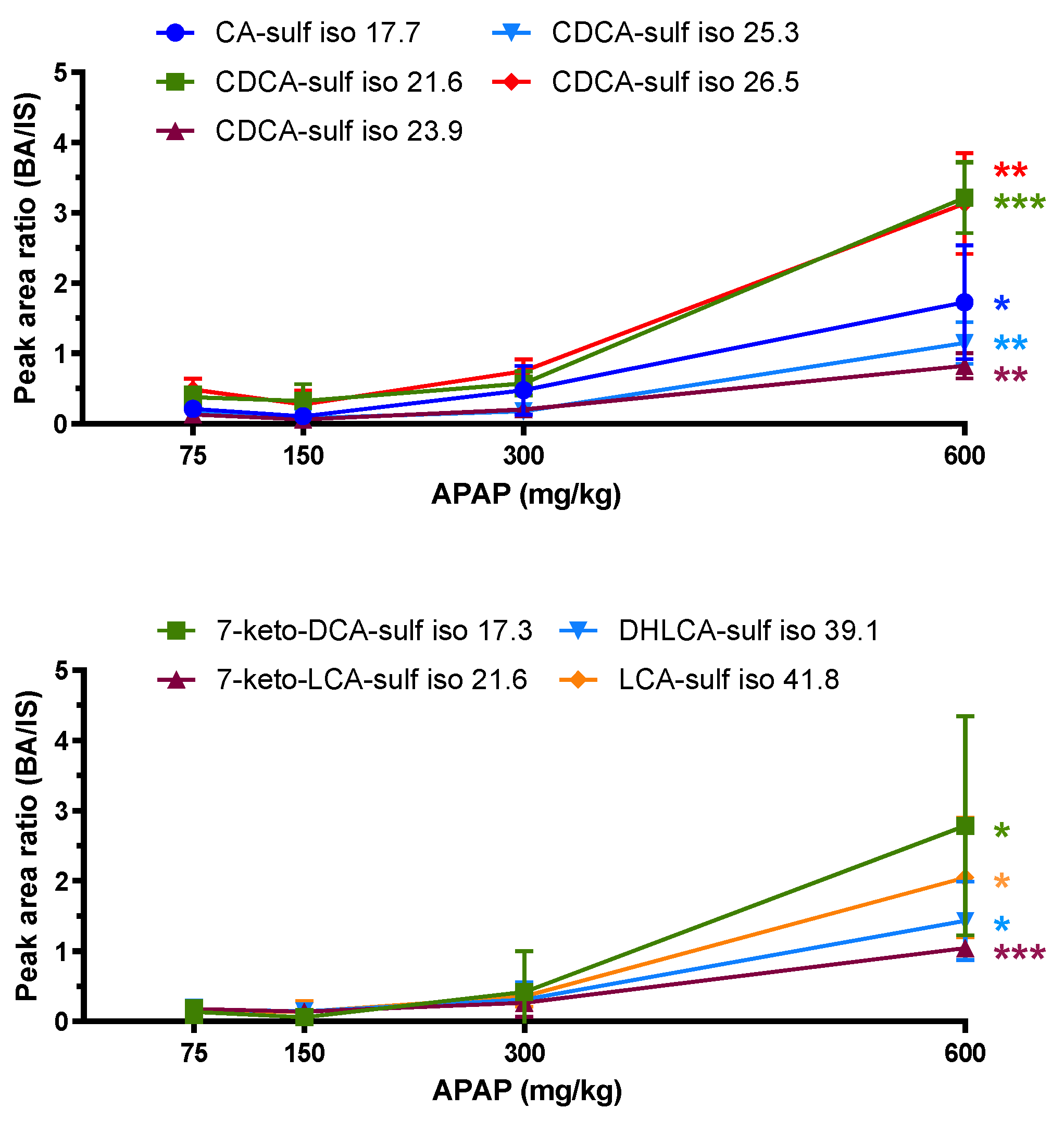
2.3. Glucuronide- and Sulfate-Conjugated Bile Acids
2.4. In Vitro Metabolism Incubations
3. Discussion
4. Materials and Methods
4.1. Chemicals and Reagents
4.2. Sample Preparation
4.2.1. Standard and Internal Standard Mix
4.2.2. Extraction of Plasma Samples
4.3. In Vitro Incubations
4.4. LC-MS/MS Analysis
4.5. Data Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Ticho, A.L.; Malhotra, P.; Dudeja, P.K.; Gill, R.K.; Alrefai, W.A. Intestinal Absorption of Bile Acids in Health and Disease. Compr. Physiol. 2019, 10, 21–56. [Google Scholar] [CrossRef] [PubMed]
- Ashby, K.; Almario, E.E.N.; Tong, W.; Borlak, J.; Mehta, R.; Chen, M. Review article: Therapeutic bile acids and the risks for hepatotoxicity. Aliment. Pharmacol. Ther. 2018, 47, 1623–1638. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kiriyama, Y.; Nochi, H. The Biosynthesis, Signaling, and Neurological Functions of Bile Acids. Biomolecules 2019, 9, 232. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, T.; Apte, U. Bile Acid Metabolism and Signaling in Cholestasis, Inflammation, and Cancer. Adv. Pharmacol. 2015, 74, 263–302. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.; Hong, J.-Y.; Rockwell, C.E.; Copple, B.L.; Jaeschke, H.; Klaassen, C.D. Effect of bile duct ligation on bile acid composition in mouse serum and liver. Liver Int. 2012, 32, 58–69. [Google Scholar] [CrossRef] [Green Version]
- Perez, M.-J.; Briz, O. Bile-acid-induced cell injury and protection. World J. Gastroenterol. 2009, 15, 1677–1689. [Google Scholar] [CrossRef]
- Perreault, M.; Białek, A.; Trottier, J.; Verreault, M.; Caron, P.; Milkiewicz, P.; Barbier, O. Role of Glucuronidation for Hepatic Detoxification and Urinary Elimination of Toxic Bile Acids during Biliary Obstruction. PLoS ONE 2013, 8, e80994. [Google Scholar] [CrossRef]
- Alnouti, Y. Bile Acid Sulfation: A Pathway of Bile Acid Elimination and Detoxification. Toxicol. Sci. 2009, 108, 225–246. [Google Scholar] [CrossRef] [Green Version]
- Yamazaki, M.; Miyake, M.; Sato, H.; Masutomi, N.; Tsutsui, N.; Adam, K.-P.; Alexander, D.C.; Lawton, K.A.; Milburn, M.V.; Ryals, J.A.; et al. Perturbation of bile acid homeostasis is an early pathogenesis event of drug induced liver injury in rats. Toxicol. Appl. Pharmacol. 2013, 268, 79–89. [Google Scholar] [CrossRef]
- Woolbright, B.L.; McGill, M.R.; Staggs, V.S.; Winefield, R.D.; Gholami, P.; Olyaee, M.; Sharpe, M.R.; Curry, S.C.; Lee, W.M.; Jaeschke, H.; et al. Glycodeoxycholic Acid Levels as Prognostic Biomarker in Acetaminophen-Induced Acute Liver Failure Patients. Toxicol. Sci. 2014, 142, 436–444. [Google Scholar] [CrossRef] [Green Version]
- Luo, L.; Aubrecht, J.; Li, D.; Warner, R.L.; Johnson, K.J.; Kenny, J.; Colangelo, J.L. Assessment of serum bile acid profiles as biomarkers of liver injury and liver disease in humans. PLoS ONE 2018, 13, e0193824. [Google Scholar] [CrossRef] [PubMed]
- Luo, L.; Schomaker, S.; Houle, C.; Aubrecht, J.; Colangelo, J.L. Evaluation of Serum Bile Acid Profiles as Biomarkers of Liver Injury in Rodents. Toxicol. Sci. 2014, 137, 12–25. [Google Scholar] [CrossRef] [Green Version]
- Mazaleuskaya, L.L.; Sangkuhl, K.; Thorn, C.F.; FitzGerald, G.A.; Altman, R.B.; Klein, T.E. PharmGKB summary: Pathways of acetaminophen metabolism at the therapeutic versus toxic doses. Pharm. Genom. 2015, 25, 416–426. [Google Scholar] [CrossRef] [Green Version]
- Hodgman, M.J.; Garrard, A.R. A Review of Acetaminophen Poisoning. Crit. Care Clin. 2012, 28, 499–516. [Google Scholar] [CrossRef] [PubMed]
- Dahlin, D.C.; Miwa, G.T.; Lu, A.Y.; Nelson, S.D. N-acetyl-p-benzoquinone imine: A cytochrome P-450-mediated oxidation product of acetaminophen. Proc. Natl. Acad. Sci. USA 1984, 81, 1327–1331. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wood, D.; Kalsi, S.S.; Waring, W.S.; Dargan, P. A review of the evidence concerning hepatic glutathione depletion and susceptibility to hepatotoxicity after paracetamol overdose. Open Access Emerg. Med. 2011, 3, 87–96. [Google Scholar] [CrossRef] [Green Version]
- Davis, D.C.; Potter, W.Z.; Jollow, D.J.; Mitchell, J.R. Species differences in hepatic glutathione depletion, covalent binding and hepatic necrosis after acetaminophen. Life Sci. 1974, 14, 2099–2109. [Google Scholar] [CrossRef] [PubMed]
- Nevada Department of Health and Human Services. FDA Acetaminophen Dosage Announcement 2012. Available online: https://www.medicaid.nv.gov/Downloads/provider/web_announcement_468_20120425.pdf (accessed on 23 December 2022).
- Larson, A.M.; Polson, J.; Fontana, R.J.; Davern, T.J.; Lalani, E.; Hynan, L.S.; Reisch, J.S.; Schiødt, F.V.; Ostapowicz, G.; Shakil, A.O.; et al. Acetaminophen-induced acute liver failure: Results of a United States multicenter, prospective study. Hepatology 2005, 42, 1364–1372. [Google Scholar] [CrossRef]
- Lee, W.M. Acetaminophen and the U.S. acute liver failure study group: Lowering the risks of hepatic failure. Hepatology 2004, 40, 6–9. [Google Scholar] [CrossRef]
- Bhushan, B.; Apte, U. Liver Regeneration after Acetaminophen Hepatotoxicity: Mechanisms and Therapeutic Opportunities. Am. J. Pathol. 2019, 189, 719–729. [Google Scholar] [CrossRef] [Green Version]
- Beckett, G.J.; Chapman, B.J.; Dyson, E.H.; Hayes, J.D. Plasma glutathione S-transferase measurements after paracetamol overdose: Evidence for early hepatocellular damage. Gut 1985, 26, 26–31. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prinville, V.; Ohlund, L.; Sleno, L. Targeted Analysis of 46 Bile Acids to Study the Effect of Acetaminophen in Rat by LC-MS/MS. Metabolites 2020, 10, 26. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dutta, M.; Cai, J.; Gui, W.; Patterson, A.D. A review of analytical platforms for accurate bile acid measurement. Anal. Bioanal. Chem. 2019, 411, 4541–4549. [Google Scholar] [CrossRef] [PubMed]
- García-Cañaveras, J.C.; Donato, M.T.; Castell, J.V.; Lahoz, A. Targeted profiling of circulating and hepatic bile acids in human, mouse, and rat using a UPLC-MRM-MS-validated method. J. Lipid Res. 2012, 53, 2231–2241. [Google Scholar] [CrossRef] [Green Version]
- Gómez, C.; Stücheli, S.; Kratschmar, D.V.; Bouitbir, J.; Odermatt, A. Development and Validation of a Highly Sensitive LC-MS/MS Method for the Analysis of Bile Acids in Serum, Plasma, and Liver Tissue Samples. Metabolites 2020, 10, 282. [Google Scholar] [CrossRef]
- Shefer, S.; Salen, G.; Hauser, S.; Dayal, B.; Batta, A. Metabolism of iso-bile acids in the rat. J. Biol. Chem. 1982, 257, 1401–1406. [Google Scholar] [CrossRef]
- Yousef, I.; Mignault, D.; Weber, A.; Tuchweber, B. Influence of Dehydrocholic Acid on the Secretion of Bile Acids and Biliary Lipids in Rats. Digestion 1990, 45, 40–51. [Google Scholar] [CrossRef]
- Thakare, R.; Alamoudi, J.A.; Gautam, N.; Rodrigues, A.D.; Alnouti, Y. Species differences in bile acids I. Plasma and urine bile acid composition. J. Appl. Toxicol. 2018, 38, 1323–1335. [Google Scholar] [CrossRef]
- Dossa, A.Y.; Escobar, O.; Golden, J.; Frey, M.R.; Ford, H.R.; Gayer, C.P. Bile acids regulate intestinal cell proliferation by modulating EGFR and FXR signaling. Am. J. Physiol. Liver Physiol. 2016, 310, G81–G92. [Google Scholar] [CrossRef] [Green Version]
- Powell, A.A.; LaRue, J.M.; Batta, A.K.; Martinez, J.D. Bile acid hydrophobicity is correlated with induction of apoptosis and/or growth arrest in HCT116 cells. Biochem. J. 2001, 356, 481–486. [Google Scholar] [CrossRef]
- Song, M.; Yang, Q.; Zhang, F.; Chen, L.; Su, H.; Yang, X.; He, H.; Liu, F.; Zheng, J.; Ling, M.; et al. Hyodeoxycholic acid (HDCA) suppresses intestinal epithelial cell proliferation through FXR-PI3K/AKT pathway, accompanied by alteration of bile acids metabolism profiles induced by gut bacteria. FASEB J. 2020, 34, 7103–7117. [Google Scholar] [CrossRef] [PubMed]
- Monte, M.J.; El-Mir, M.Y.; Sainz, G.R.; Bravo, P.; Marin, J.J. Bile acid secretion during synchronized rat liver regeneration. Biochim. Biophys. Acta (BBA)—Mol. Basis Dis. 1997, 1362, 56–66. [Google Scholar] [CrossRef] [Green Version]
- Monte, M.J.; Martinez-Diez, M.C.; El-Mir, M.Y.; E Mendoza, M.; Bravo, P.; Bachs, O.; Marin, J.J. Changes in the pool of bile acids in hepatocyte nuclei during rat liver regeneration. J. Hepatol. 2002, 36, 534–542. [Google Scholar] [CrossRef] [PubMed]
- Bhushan, B.; Borude, P.; Edwards, G.; Walesky, C.; Cleveland, J.; Li, F.; Ma, X.; Apte, U. Role of Bile Acids in Liver Injury and Regeneration following Acetaminophen Overdose. Am. J. Pathol. 2013, 183, 1518–1526. [Google Scholar] [CrossRef] [Green Version]
- Stamp, D.; Jenkins, G. An overview of bile-acid synthesis, chemistry and function. In Bile Acids: Toxicology and Bioactivity; Issues in Toxicology; RSC Publishing: Cambridge, UK, 2008; pp. 1–13. [Google Scholar]
- James, L.; Yan, K.; Pence, L.; Simpson, P.; Bhattacharyya, S.; Gill, P.; Letzig, L.; Kearns, G.; Beger, R. Comparison of Bile Acids and Acetaminophen Protein Adducts in Children and Adolescents with Acetaminophen Toxicity. PLoS ONE 2015, 10, e0131010. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- LeBlanc, A.; Shiao, T.C.; Roy, R.; Sleno, L. Absolute Quantitation of NAPQI-Modified Rat Serum Albumin by LC–MS/MS: Monitoring Acetaminophen Covalent Binding in Vivo. Chem. Res. Toxicol. 2014, 27, 1632–1639. [Google Scholar] [CrossRef]
- Shulpekova, Y.; Shirokova, E.; Zharkova, M.; Tkachenko, P.; Tikhonov, I.; Stepanov, A.; Sinitsyna, A.; Izotov, A.; Butkova, T.; Shulpekova, N.; et al. A Recent Ten-Year Perspective: Bile Acid Metabolism and Signaling. Molecules 2022, 27, 1983. [Google Scholar] [CrossRef]
- Choudhuri, S.; Klaassen, C.D. Molecular Regulation of Bile Acid Homeostasis. Drug Metab. Dispos. 2022, 50, 425–455. [Google Scholar] [CrossRef]
- Brandon, E.F.; Raap, C.D.; Meijerman, I.; Beijnen, J.H.; Schellens, J.H. An update on in vitro test methods in human hepatic drug biotransformation research: Pros and cons. Toxicol. Appl. Pharmacol. 2003, 189, 233–246. [Google Scholar] [CrossRef] [PubMed]
- Thakare, R.; Alamoudi, J.A.; Gautam, N.; Rodrigues, A.D.; Alnouti, Y. Species differences in bile acids II. Bile acid metabolism. J. Appl. Toxicol. 2018, 38, 1336–1352. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Bile Acid | Formula | RT (min) | p-Value | Fold Change * (600/75 dose) |
|---|---|---|---|---|
| keto-LCA isomer | C24H38O4 | 23.6 | 0.0082 | 3.1 |
| 25.9 | 0.032 | 14.9 | ||
| 30.3 | 0.015 | 3.1 | ||
| keto-DCA isomer | C24H38O5 | 10.2 | 0.027 | 23.9 |
| 15.3 | 0.028 | 23.0 | ||
| 15.8 | 0.0018 | 12.0 | ||
| 17.8 | 0.035 | 13.3 | ||
| 18.5 | 0.010 | 5.3 | ||
| 20.2 | 0.018 | 9.1 | ||
| 23.0 | 0.013 | 2.6 | ||
| CA isomer | C24H40O5 | 14.3 | 0.031 | 20.0 |
| 18.9 | 0.014 | 14.7 | ||
| GCA isomer | C26H43NO6 | 6.7 | 0.043 | 6.1 |
| 8.4 | 0.0057 | 8.9 | ||
| 9.5 | 0.0078 | 14.0 | ||
| 10.8 | 0.036 | 5.6 | ||
| 11.8 | 0.021 | 8.0 | ||
| GCDCA isomer | C26H43NO5 | 13.8 | 0.043 | 20.5 |
| 19.4 | 0.0026 | 8.0 | ||
| CDCA isomer | C24H40O4 | 22.5 | 0.031 | 60.0 |
| 25.3 | 0.030 | 19.5 | ||
| 26.7 | 0.010 | 8.6 |
| Bile Acid (Sulf-Isomer) | Formula | m/z [M-H]− | ppm | RT (min) | p-Value | Fold Change* (600/75 dose) |
|---|---|---|---|---|---|---|
| keto-DCA+SO3 | C24H38O8S | 485.2212 | −0.5 | 17.3 | 0.035 | 20.7 |
| CA+SO3 | C24H40O8S | 487.2363 | −1.7 | 17.7 | 0.025 | 8.2 |
| keto-LCA+SO3 | C24H38O7S | 469.2278 | 2.7 | 21.6 | 0.000085 | 5.9 |
| CDCA+SO3 | C24H40O7S | 471.2412 | −2.1 | 21.6 | 0.00025 | 8.5 |
| 471.2425 | 0.6 | 23.9 | 0.0015 | 6.3 | ||
| 471.2405 | −3.6 | 25.3 | 0.0026 | 6.9 | ||
| 471.2411 | −2.3 | 26.5 | 0.0017 | 6.5 | ||
| DHLCA+SO3 | C24H38O6S | 453.2330 | 3.0 | 39.1 | 0.013 | 8.3 |
| LCA+SO3 | C24H40O6S | 455.2481 | 1.8 | 41.8 | 0.013 | 14.0 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mireault, M.; Prinville, V.; Ohlund, L.; Sleno, L. Semi-Targeted Profiling of Bile Acids by High-Resolution Mass Spectrometry in a Rat Model of Drug-Induced Liver Injury. Int. J. Mol. Sci. 2023, 24, 2489. https://doi.org/10.3390/ijms24032489
Mireault M, Prinville V, Ohlund L, Sleno L. Semi-Targeted Profiling of Bile Acids by High-Resolution Mass Spectrometry in a Rat Model of Drug-Induced Liver Injury. International Journal of Molecular Sciences. 2023; 24(3):2489. https://doi.org/10.3390/ijms24032489
Chicago/Turabian StyleMireault, Myriam, Vivaldy Prinville, Leanne Ohlund, and Lekha Sleno. 2023. "Semi-Targeted Profiling of Bile Acids by High-Resolution Mass Spectrometry in a Rat Model of Drug-Induced Liver Injury" International Journal of Molecular Sciences 24, no. 3: 2489. https://doi.org/10.3390/ijms24032489