
The Effects of the Follicle-Stimulating Hormone on Human Follicular Fluid-Derived Stromal Cells
 , ,
, ,
Abstract
:1. Introduction
2. Results
2.1. Study Population, the Clinical Data
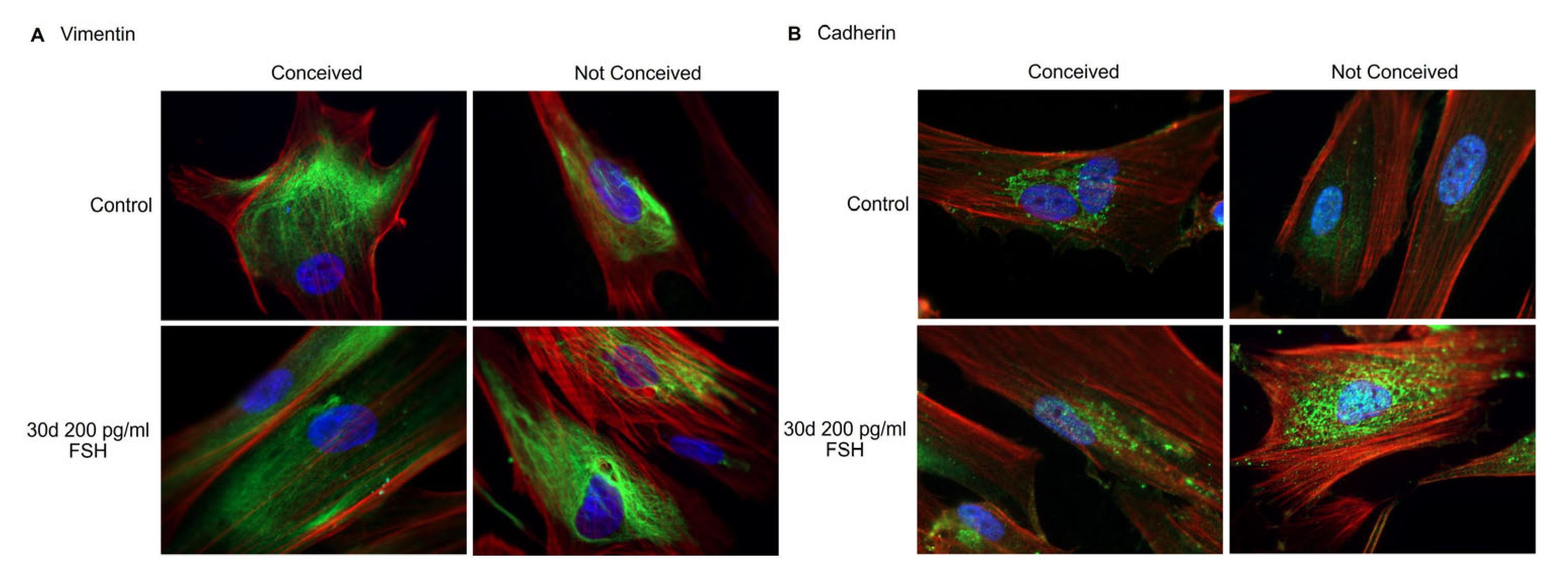
2.2. Characterization of Follicular Fluid-Derived Stromal Cells Treated with FSH
2.3. Analysis of the Protein Expression in FF Stromal Cells
2.4. Assessment of Gene Expression in FF Sediment Cells and FSH-Treated FF Stromal Cells
2.5. Changes in FF Stromal Cells Secreted Proteins during FSH Treatment
3. Discussion
4. Materials and Methods
4.1. Study Population and Data Collection

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Characteristic | Conceived | Not Conceived |
|---|---|---|
| Patient count (n) | 8 | 8 |
| Average Age (years) | 30.6 ± 4.7 | 34.4 ± 2.7 |
| Duration of infertility (years) | 5.5 ± 3.8 | 5.5 ± 3.4 |
| FSH U/L | 5.9 ± 1.6 | 6.0 ± 1.7 |
| AMH µg/L | 4.6 ± 2.5 | 2.1 ± 1.5 |
| LH U/L | 4.5 ± 1.4 | 3.4 ± 0.7 |
| Estradiol pmol/L | 146.6 ± 86.1 | 132.0 ± 47.1 |
| Average of oocytes retrieved (n) | 16.8 ± 4.9 | 11.6 ± 4.2 |
| Average of zygotes (n) | 11.9 ± 3.2 | 7.8 ± 4.3 |
| BMI > 25 (n) | 2 | 1 |
| Diagnosis (n) | Tubal factor infertility (2) Unexplained infertility (5) Male factor infertility (1) | Tubal factor infertility (2) Unexplained infertility (4) Male factor infertility (2) |
| Endometriosis (n) | Stage I (2) | Stage I (1) Stage II (1) |
4.2. Collection of Follicular Fluid, Extraction of Sediment Cells and Cultivation, Proliferation and FSH Treatment of Follicular Fluid-Derived Stromal Cells
4.3. Evaluation of Cell Surface Markers via Flow Cytometry Analysis
4.4. Gene Expression Analysis via RT-qPCR
4.5. Proteins Analysis by Fluorescent Microscopy
4.6. LC-MS-Based Protein Identification
4.7. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Sun, H.; Gong, T.-T.; Jiang, Y.-T.; Zhang, S.; Zhao, Y.-H.; Wu, Q.-J. Global, Regional, and National Prevalence and Disability-Adjusted Life-Years for Infertility in 195 Countries and Territories, 1990–2017: Results from a Global Burden of Disease Study, 2017. Aging 2019, 11, 10952–10991. [Google Scholar] [CrossRef]
- Datta, J.; Palmer, M.J.; Tanton, C.; Gibson, L.J.; Jones, K.G.; Macdowall, W.; Glasier, A.; Sonnenberg, P.; Field, N.; Mercer, C.H.; et al. Prevalence of Infertility and Help Seeking among 15 000 Women and Men. Hum. Reprod. 2016, 31, 2108–2118. [Google Scholar] [CrossRef] [Green Version]
- Mascarenhas, M.N.; Cheung, H.; Mathers, C.D.; Stevens, G.A. Measuring Infertility in Populations: Constructing a Standard Definition for Use with Demographic and Reproductive Health Surveys. Popul. Health Metr. 2012, 10, 17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kushnir, V.A.; Barad, D.H.; Albertini, D.F.; Darmon, S.K.; Gleicher, N. Systematic Review of Worldwide Trends in Assisted Reproductive Technology 2004–2013. Reprod. Biol. Endocrinol. 2017, 15, 6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dobson, R. Proportion of Babies Born in Europe after IVF Varies 20-Fold. BMJ 2006, 332, 626. [Google Scholar] [CrossRef] [Green Version]
- ART Success Rates|CDC. Available online: https://www.cdc.gov/art/artdata/index.html (accessed on 21 December 2022).
- Dumesic, D.A.; Meldrum, D.R.; Katz-Jaffe, M.G.; Krisher, R.L.; Schoolcraft, W.B. Oocyte Environment: Follicular Fluid and Cumulus Cells Are Critical for Oocyte Health. Fertil. Steril. 2015, 103, 303–316. [Google Scholar] [CrossRef]
- Rodgers, R.J.; Irving-Rodgers, H.F. Formation of the Ovarian Follicular Antrum and Follicular Fluid1. Biol. Reprod. 2010, 82, 1021–1029. [Google Scholar] [CrossRef] [Green Version]
- Revelli, A.; Piane, L.D.; Casano, S.; Molinari, E.; Massobrio, M.; Rinaudo, P. Follicular Fluid Content and Oocyte Quality: From Single Biochemical Markers to Metabolomics. Reprod. Biol. Endocrinol. 2009, 7, 40. [Google Scholar] [CrossRef] [Green Version]
- Orlowski, M.; Sarao, M.S. Physiology, Follicle Stimulating Hormone. In StatPearls; StatPearls Publishing: Treasure Island, FL, USA, 2022. [Google Scholar]
- Rosen, M.P.; Zamah, A.M.; Shen, S.; Dobson, A.T.; McCulloch, C.E.; Rinaudo, P.F.; Lamb, J.D.; Cedars, M.I. The Effect of Follicular Fluid Hormones on Oocyte Recovery after Ovarian Stimulation: FSH Level Predicts Oocyte Recovery. Reprod. Biol. Endocrinol. 2009, 7, 35. [Google Scholar] [CrossRef] [Green Version]
- Hennet, M.L.; Combelles, C.M.H. The Antral Follicle: A Microenvironment for Oocyte Differentiation. Int. J. Dev. Biol. 2013, 56, 819–831. [Google Scholar] [CrossRef]
- Riva, F.; Omes, C.; Bassani, R.; Nappi, R.E.; Mazzini, G.; Cornaglia, A.I.; Casasco, A. In-Vitro Culture System for Mesenchymal Progenitor Cells Derived from Waste Human Ovarian Follicular Fluid. Reprod. Biomed. Online 2014, 29, 457–469. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smitz, J.; Wolfenson, C.; Chappel, S.; Ruman, J. Follicle-Stimulating Hormone: A Review of Form and Function in the Treatment of Infertility. Reprod. Sci. 2016, 23, 706–716. [Google Scholar] [CrossRef] [PubMed]
- Song, K.; Dai, L.; Long, X.; Wang, W.; Di, W. Follicle-Stimulating Hormone Promotes the Proliferation of Epithelial Ovarian Cancer Cells by Activating Sphingosine Kinase. Sci. Rep. 2020, 10, 13834. [Google Scholar] [CrossRef] [PubMed]
- Ji, Q.; Liu, P.I.; Chen, P.K.; Aoyama, C. Follicle Stimulating Hormone-Induced Growth Promotion and Gene Expression Profiles on Ovarian Surface Epithelial Cells. Int. J. Cancer 2004, 112, 803–814. [Google Scholar] [CrossRef] [PubMed]
- Allan, C.M.; Garcia, A.; Spaliviero, J.; Zhang, F.-P.; Jimenez, M.; Huhtaniemi, I.; Handelsman, D.J. Complete Sertoli Cell Proliferation Induced by Follicle-Stimulating Hormone (FSH) Independently of Luteinizing Hormone Activity: Evidence from Genetic Models of Isolated FSH Action. Endocrinology 2004, 145, 1587–1593. [Google Scholar] [CrossRef]
- Bi, Y.; Yang, S.; Wang, H.; Chang, G.; Chen, G. Follicle-Stimulating Hormone Is Expressed in Ovarian Follicles of Chickens and Promotes Ovarian Granulosa Cell Proliferation. J. Integr. Agric. 2021, 20, 2749–2757. [Google Scholar] [CrossRef]
- Li, X.; Lin, J.; Chen, Y.; Wang, L.; Han, B.; Jia, B.; Wu, Y.; Huang, J. FSH Promotes the Proliferation of Sheep Granulosa Cells by Inhibiting the Expression of TSP1. Anim. Biotechnol. 2022, 33, 260–272. [Google Scholar] [CrossRef]
- Du, L.; Gu, T.; Zhang, Y.; Huang, Z.; Wu, N.; Zhao, W.; Chang, G.; Xu, Q.; Chen, G. Transcriptome Profiling to Identify Key Mediators of Granulosa Cell Proliferation upon FSH Stimulation in the Goose (Anser cygnoides). Br. Poult. Sci. 2018, 59, 416–421. [Google Scholar] [CrossRef]
- Shi, X.; Qiu, S.; Zhuang, W.; Wang, C.; Zhang, S.; Yuan, N.; Yuan, F.; Qiao, Y. Follicle-Stimulating Hormone Inhibits Cervical Cancer via NF-ΚB Pathway. OncoTargets Ther. 2018, 11, 8107–8115. [Google Scholar] [CrossRef] [Green Version]
- Chandramohan, Y.; Jeganathan, K.; Sivanesan, S.; Koka, P.; Amritha, T.M.S.; Vimalraj, S.; Dhanasekaran, A. Assessment of Human Ovarian Follicular Fluid Derived Mesenchymal Stem Cells in Chitosan/PCL/Zn Scaffold for Bone Tissue Regeneration. Life Sci. 2021, 264, 118502. [Google Scholar] [CrossRef]
- Omes, C.; Fassina, L.; Van Vlierberghe, S.; Magenes, G.; Dubruel, P.; Vaghi, P.; Reguzzoni, M.; Riva, F. A Case of Successful Interaction between Cells Derived from Human Ovarian Follicular Liquid and Gelatin Cryogel for Biotech and Medical Applications. Annu. Int. Conf. IEEE Eng. Med. Biol. Soc. 2013, 2013, 6240–6243. [Google Scholar] [CrossRef] [PubMed]
- Somigliana, E.; Viganò, P.; La Sala, G.B.; Balasini, M.; Candiani, M.; Incerti, L.; Busacca, M.; Vignali, M. Follicular Fluid as a Favourable Environment for Endometrial and Endometriotic Cell Growth in Vitro. Hum. Reprod. 2001, 16, 1076–1080. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hollingsworth, J.; Lau, A.; Tone, A.; Kollara, A.; Allen, L.; Colgan, T.J.; Dube, V.; Rosen, B.; Murphy, K.J.; Greenblatt, E.M.; et al. BRCA1 Mutation Status and Follicular Fluid Exposure Alters NFκB Signaling and ISGylation in Human Fallopian Tube Epithelial Cells. Neoplasia 2018, 20, 697–709. [Google Scholar] [CrossRef]
- da Luz, C.M.; da Broi, M.G.; Donabela, F.C.; Paro de Paz, C.C.; Meola, J.; Navarro, P.A. PTGS2 Down-Regulation in Cumulus Cells of Infertile Women with Endometriosis. Reprod. Biomed. Online 2017, 35, 379–386. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Niringiyumukiza, J.D.; Cai, H.; Xiang, W. Prostaglandin E2 Involvement in Mammalian Female Fertility: Ovulation, Fertilization, Embryo Development and Early Implantation. Reprod. Biol. Endocrinol. RBE 2018, 16, 43. [Google Scholar] [CrossRef] [PubMed]
- Sayutti, N.; Abu, M.A.; Ahmad, M.F. PCOS and Role of Cumulus Gene Expression in Assessing Oocytes Quality. Front. Endocrinol. 2022, 13, 843867. [Google Scholar] [CrossRef] [PubMed]
- França, M.M.; Mendonca, B.B. Genetics of Ovarian Insufficiency and Defects of Folliculogenesis. Best Pract. Res. Clin. Endocrinol. Metab. 2022, 36, 101594. [Google Scholar] [CrossRef]
- Osiński, M.; Wirstlein, P.; Wender-Ożegowska, E.; Mikołajczyk, M.; Jagodziński, P.P.; Szczepańska, M. HSD3B2, HSD17B1, HSD17B2, ESR1, ESR2 and AR Expression in Infertile Women with Endometriosis. Ginekol. Pol. 2018, 89, 125–134. [Google Scholar] [CrossRef] [Green Version]
- Richani, D.; Constance, K.; Lien, S.; Agapiou, D.; Stocker, W.A.; Hedger, M.P.; Ledger, W.L.; Thompson, J.G.; Robertson, D.M.; Mottershead, D.G.; et al. Cumulin and FSH Cooperate to Regulate Inhibin B and Activin B Production by Human Granulosa-Lutein Cells In Vitro. Endocrinology 2019, 160, 853–862. [Google Scholar] [CrossRef]
- Owens, L.A.; Kristensen, S.G.; Lerner, A.; Christopoulos, G.; Lavery, S.; Hanyaloglu, A.C.; Hardy, K.; Yding Andersen, C.; Franks, S. Gene Expression in Granulosa Cells From Small Antral Follicles From Women With or Without Polycystic Ovaries. J. Clin. Endocrinol. Metab. 2019, 104, 6182–6192. [Google Scholar] [CrossRef]
- Bhartiya, D.; Patel, H. An Overview of FSH-FSHR Biology and Explaining the Existing Conundrums. J. Ovarian Res. 2021, 14, 144. [Google Scholar] [CrossRef] [PubMed]
- De Conto, E.; Matte, U.; Cunha-Filho, J.S. BMP-6 and SMAD4 Gene Expression Is Altered in Cumulus Cells from Women with Endometriosis-Associated Infertility. Acta Obstet. Gynecol. Scand. 2021, 100, 868–875. [Google Scholar] [CrossRef] [PubMed]
- Law, N.C.; Weck, J.; Kyriss, B.; Nilson, J.H.; Hunzicker-Dunn, M. Lhcgr Expression in Granulosa Cells: Roles for PKA-Phosphorylated β-Catenin, TCF3, and FOXO1. Mol. Endocrinol. 2013, 27, 1295–1310. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Z.; Wu, L.; Diao, F.; Chen, B.; Fu, J.; Mao, X.; Yan, Z.; Li, B.; Mu, J.; Zhou, Z.; et al. Novel Mutations in LHCGR (Luteinizing Hormone/Choriogonadotropin Receptor): Expanding the Spectrum of Mutations Responsible for Human Empty Follicle Syndrome. J. Assist. Reprod. Genet. 2020, 37, 2861–2868. [Google Scholar] [CrossRef] [PubMed]
- Zamah, A.M.; Hassis, M.E.; Albertolle, M.E.; Williams, K.E. Proteomic Analysis of Human Follicular Fluid from Fertile Women. Clin. Proteom. 2015, 12, 5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Regiani, T.; Cordeiro, F.B.; da Costa, L.d.V.T.; Salgueiro, J.; Cardozo, K.; Carvalho, V.M.; Perkel, K.J.; Zylbersztejn, D.S.; Cedenho, A.P.; Lo Turco, E.G. Follicular Fluid Alterations in Endometriosis: Label-Free Proteomics by MSE as a Functional Tool for Endometriosis. Syst. Biol. Reprod. Med. 2015, 61, 263–276. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ambekar, A.S.; Kelkar, D.S.; Pinto, S.M.; Sharma, R.; Hinduja, I.; Zaveri, K.; Pandey, A.; Prasad, T.S.K.; Gowda, H.; Mukherjee, S. Proteomics of Follicular Fluid from Women with Polycystic Ovary Syndrome Suggests Molecular Defects in Follicular Development. J. Clin. Endocrinol. Metab. 2015, 100, 744–753. [Google Scholar] [CrossRef]






| Name | Primer Sequence | Product Size, bp |
|---|---|---|
| RPL13A | F: GTTGATGCCTTCACAGCGTA | 128 |
| R: AGATGGCGGAGGTGCAG | ||
| STAR | F: GGCTCAGGAAGGACGAAGAACC | 188 |
| R: ATCACAGCCTGTTGCCTCAGC | ||
| INHBA | F: CCTCGGAGATCATCACG | 238 |
| R: CCCTTTAAGCCCACTTCCTC | ||
| INHBB | F: CCTGAAACTCCTGCCCTACG | 104 |
| R: CCACCATGTTCCACCTGTCA | ||
| HAS2 | F: AGCCTTCAGAGCACTGGGACGA | 81 |
| R: ACAGATGAGGCTGGGTCAAGCA | ||
| PTGS2 | F: TGAAACCCACTCCAAACACA | 198 |
| R: AGGAGAGGTTAGAGAAGGCT | ||
| LHCGR | F: TGGAGAAGATGCACAATGGA | 122 |
| R: GGCAATTAGCCTCTGAATGG | ||
| HSD3B2 | F: TGCCTTGTGACAGGAGCAGG | 238 |
| R: TACAGGCGGTGTGGATGACG | ||
| CYP11A1 | F: GTGATGACCTGTTCCGCTTTGC | 155 |
| R: AAGGTTGAGCATGGGGACGC | ||
| CYP19A1 | F: GCTGGACACCTCTAACACGCT | 289 |
| R: CAGGTCACCACGTTTCTCTGCT | ||
| AMH | F: CGCCTGGTGGTCCTACAC | 60 |
| R: GAACCTCAGCGAGGGTGTT | ||
| FSHR | F: TGGGCTCAGGATGTCATCATCGGA | 145 |
| R: TGGATGACTCGAAGCTTGGTGAGG |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Skliutė, G.; Vaigauskaitė-Mažeikienė, B.; Kaupinis, A.; Valius, M.; Kazėnaitė, E.; Navakauskienė, R. The Effects of the Follicle-Stimulating Hormone on Human Follicular Fluid-Derived Stromal Cells. Int. J. Mol. Sci. 2023, 24, 2450. https://doi.org/10.3390/ijms24032450
Skliutė G, Vaigauskaitė-Mažeikienė B, Kaupinis A, Valius M, Kazėnaitė E, Navakauskienė R. The Effects of the Follicle-Stimulating Hormone on Human Follicular Fluid-Derived Stromal Cells. International Journal of Molecular Sciences. 2023; 24(3):2450. https://doi.org/10.3390/ijms24032450
Chicago/Turabian StyleSkliutė, Giedrė, Brigita Vaigauskaitė-Mažeikienė, Algirdas Kaupinis, Mindaugas Valius, Edita Kazėnaitė, and Rūta Navakauskienė. 2023. "The Effects of the Follicle-Stimulating Hormone on Human Follicular Fluid-Derived Stromal Cells" International Journal of Molecular Sciences 24, no. 3: 2450. https://doi.org/10.3390/ijms24032450