

Establishment of a Pseudovirus Platform for Neuraminidase Inhibiting Antibody Analysis

 , ,
, ,
Abstract
:1. Introduction
2. Results
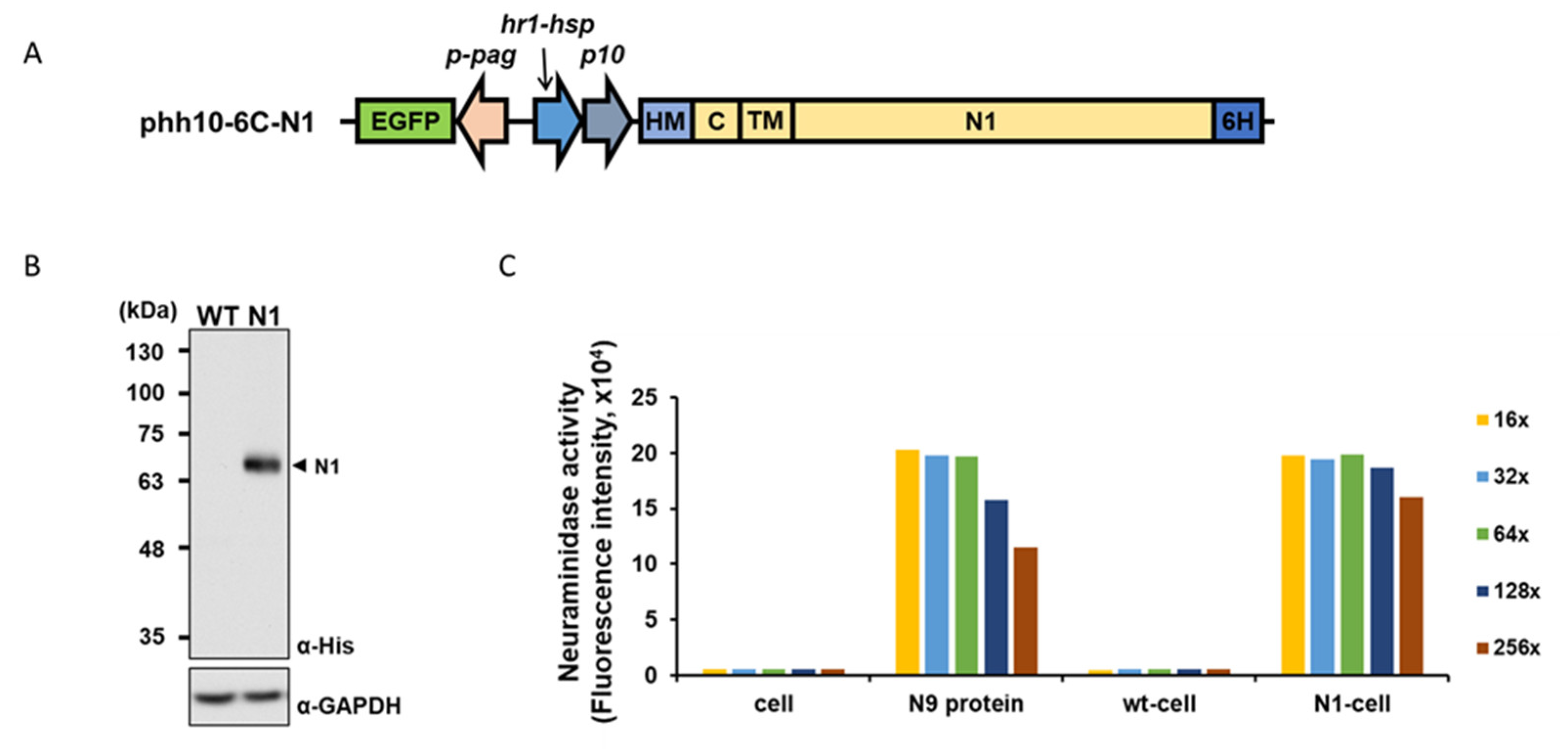
2.1. Production of a Recombinant Baculovirus Expressing Functional Influenza Virus NA
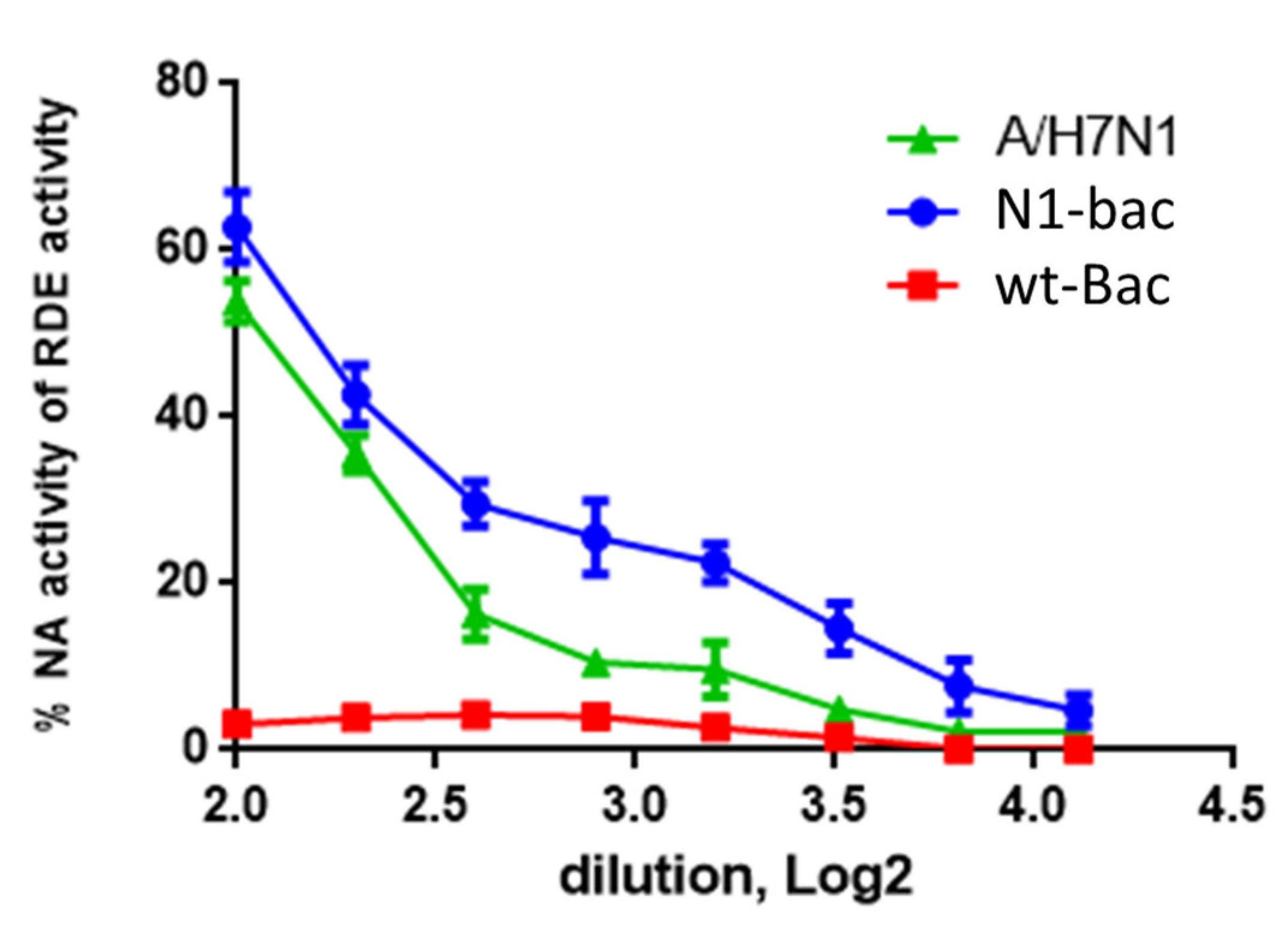
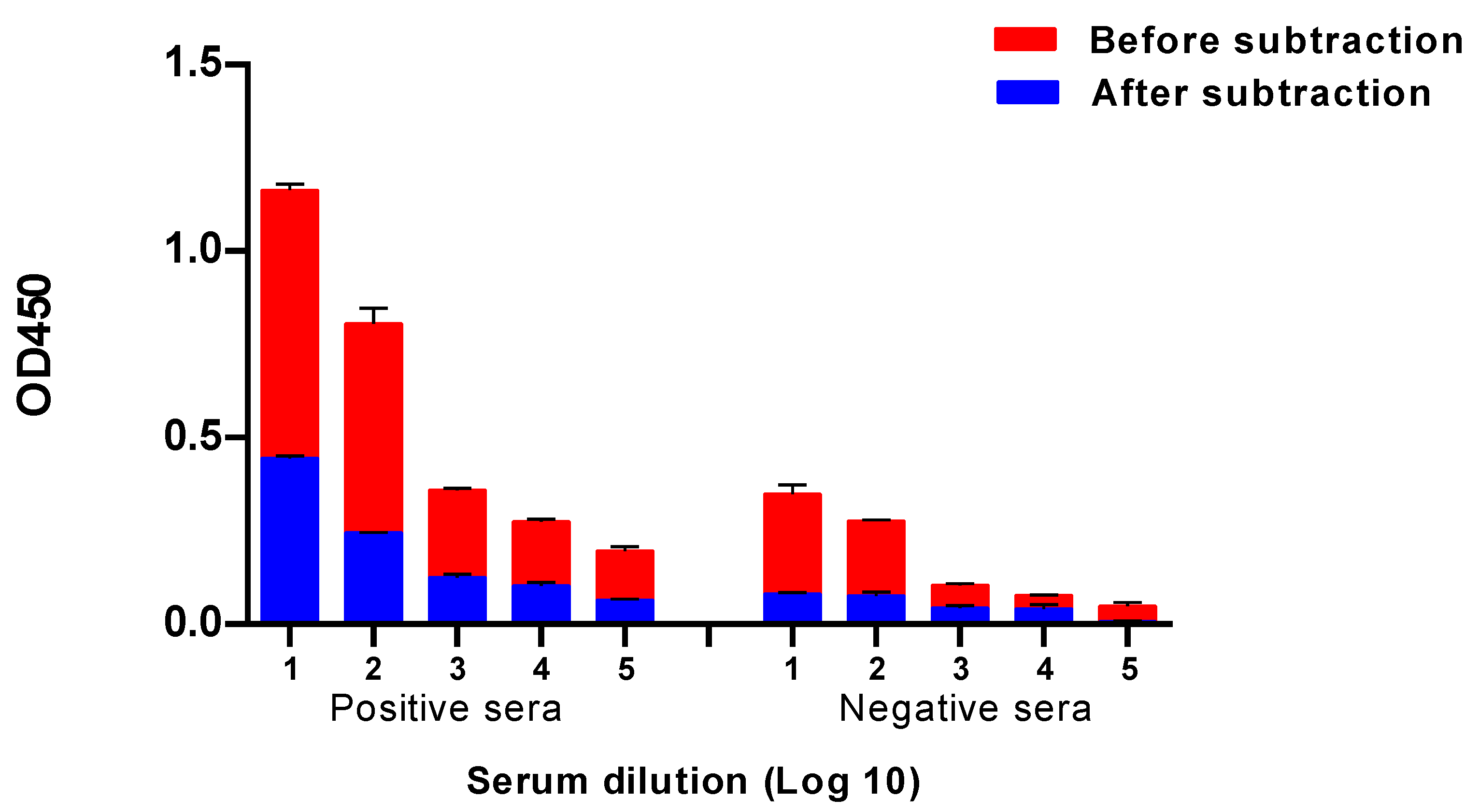
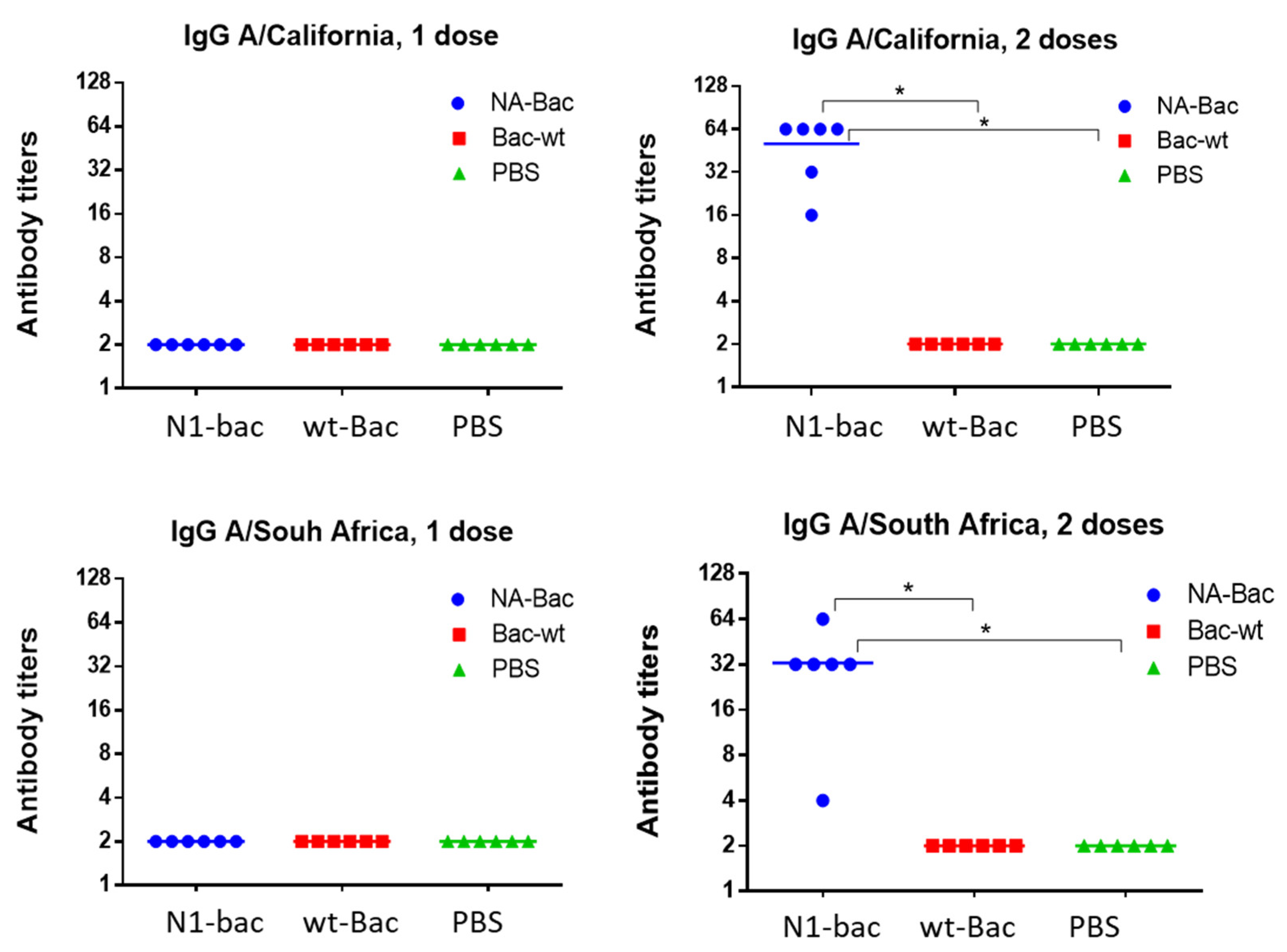
2.2. Development of a Baculovirus Platform for Assessing Antibodies to Influenza Virus NA
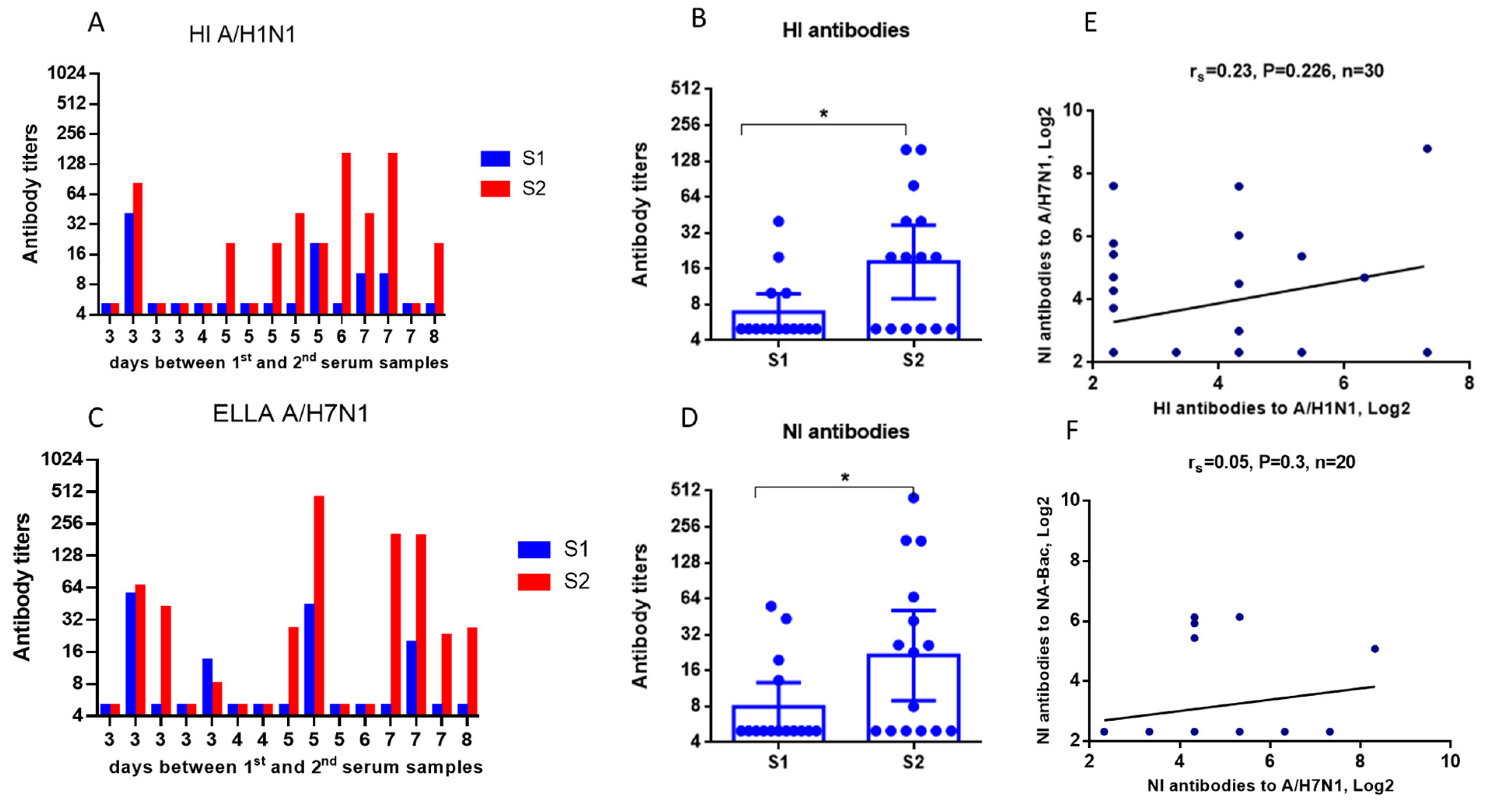
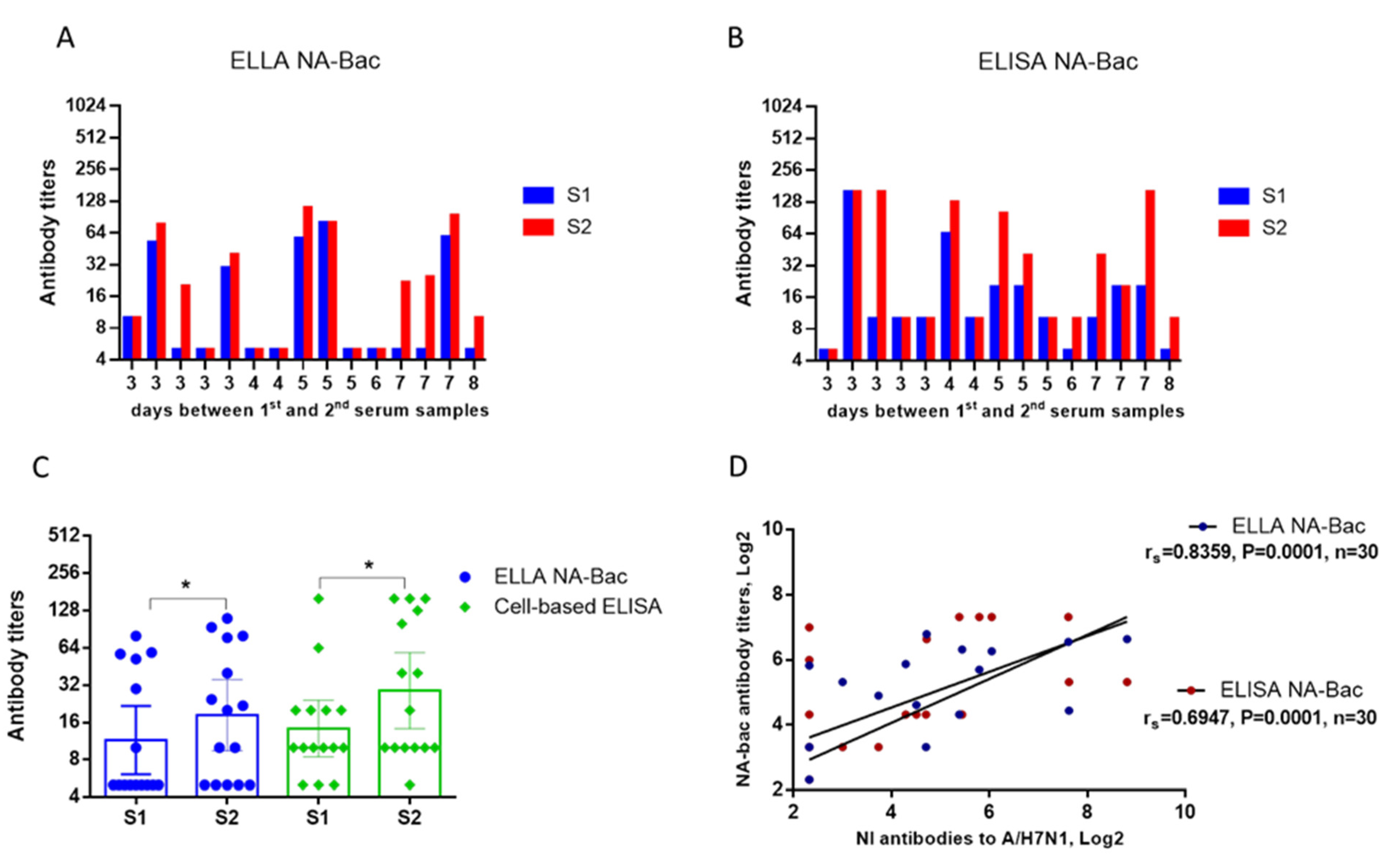
2.3. Evaluation of NA Antibodies in the Paired Sera of Patients with an Influenza Infection
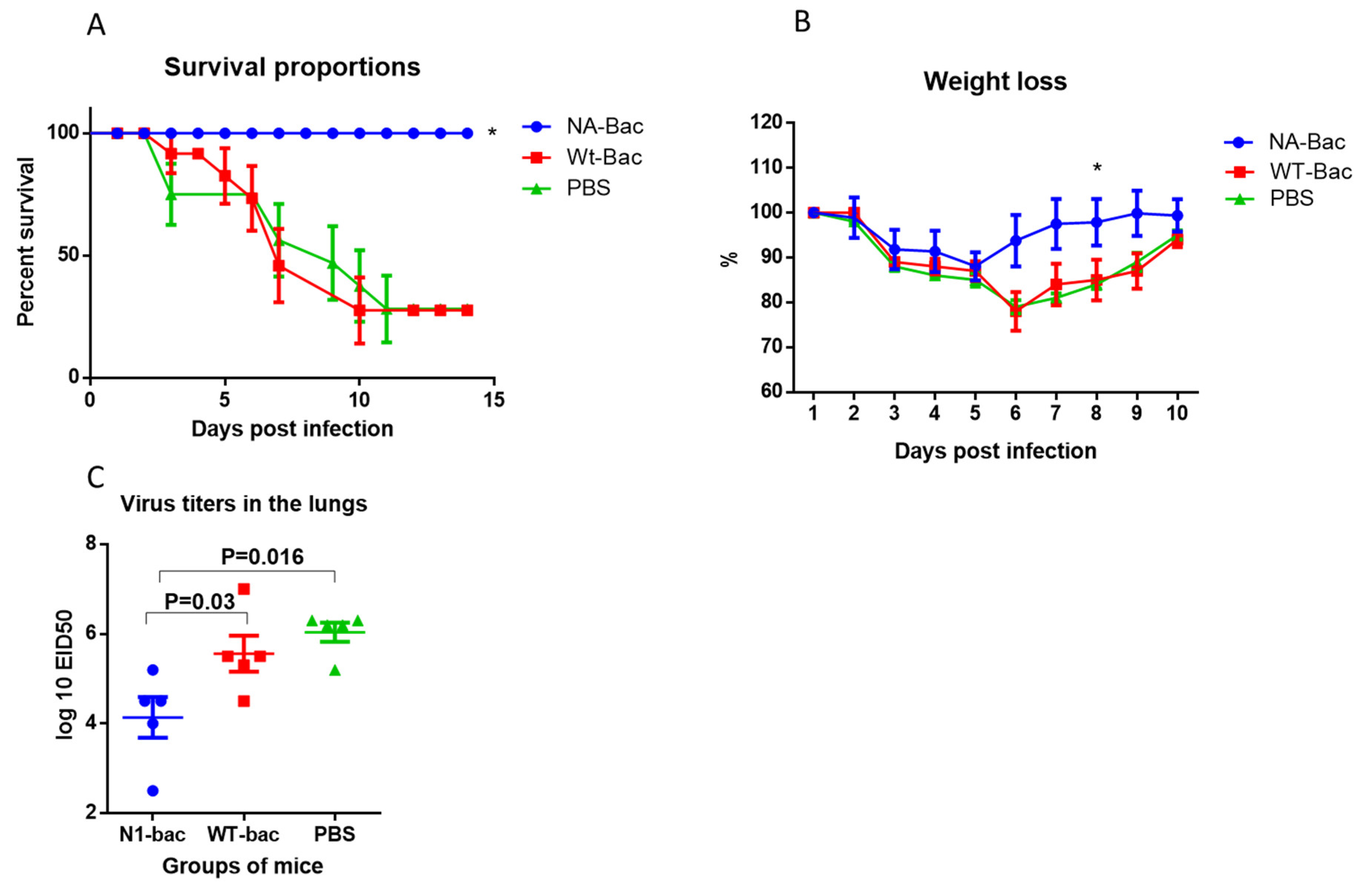
2.4. Protective Properties of Antibodies to Influenza Virus NA in a Mouse Model of Influenza Infection
3. Discussion
4. Materials and Methods
4.1. Ethic Statements
4.2. Viruses
4.3. Production of a Recombinant Baculovirus N1-Bac
4.4. Western Blotting Analysis
4.5. Fluorescence-Based NA Activity Assay
4.6. NA Activity Assay
4.7. Enzyme-Linked Lectin Assay (ELLA)
4.8. Cell-Based ELISA
4.9. Hemagglutination Inhibition (HI) Test
4.10. Mouse Immunization and Challenge
4.11. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Murphy, B.R.; Kasel, J.A.; Chanock, R.M. Association of serum anti-neuraminidase antibody with resistance to influenza in man. N. Engl. J. Med. 1972, 286, 1329–1332. [Google Scholar] [CrossRef]
- Maier, H.E.; Nachbagauer, R.; Kuan, G.; Ng, S.; Lopez, R.; Sanchez, N.; Stadlbauer, D.; Gresh, L.; Schiller, A.; Rajabhathor, A.; et al. Pre-existing antineuraminidase antibodies are associated with shortened duration of influenza A (H1N1) pdm virus shedding and illness in naturally infected adults. Clin. Infect. Dis. 2020, 70, 2290–2297. [Google Scholar] [CrossRef] [PubMed]
- McCullers, J.A.; Huber, V.C. Correlates of vaccine protection from influenza and its complications. Hum. Vaccines Immunother. 2012, 8, 34–44. [Google Scholar] [CrossRef] [Green Version]
- Eichelberger, M.C.; Monto, A.S. Neuraminidase, the forgotten surface antigen, emerges as an influenza vaccine target for broadened protection. J. Infect. Dis. 2019, 219, S75–S80. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bright, R.A.; Neuzil, K.M.; Pervikov, Y.; Palkonyay, L. WHO meeting on the role of neuraminidase in inducing protective immunity against influenza infection, Vilamoura, Portugal, September 14, 2008. Vaccine 2009, 27, 6366–6369. [Google Scholar] [CrossRef] [PubMed]
- Krammer, F.; Fouchier, R.A.; Eichelberger, M.C.; Webby, R.J.; Shaw-Saliba, K.; Wan, H.; Wilson, P.C.; Compans, R.W.; Skountzou, I.; Monto, A.S. NAction! How can neuraminidase-based immunity contribute to better influenza virus vaccines? MBio 2018, 9, e02332-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krammer, F.; Palese, P. Advances in the development of influenza virus vaccines. Nat. Rev. Drug Discov. 2015, 14, 167–182. [Google Scholar] [CrossRef] [PubMed]
- Toon, K.; Bentley, E.M.; Mattiuzzo, G. More than just gene therapy vectors: Lentiviral vector pseudotypes for serological investigation. Viruses 2021, 13, 217. [Google Scholar] [CrossRef] [PubMed]
- Schlimgen, R.; Howard, J.; Wooley, D.; Thompson, M.; Baden, L.R.; Yang, O.O.; Christiani, D.C.; Mostoslavsky, G.; Diamond, D.V.; Duane, E.G.; et al. Risks Associated With Lentiviral Vector Exposures and Prevention Strategies. J. Occup. Environ. Med. 2016, 58, 1159–1166. [Google Scholar] [CrossRef] [Green Version]
- Assenberg, R.; Wan, P.T.; Geisse, S.; Mayr, L.M. Advances in recombinant protein expression for use in pharmaceutical research. Curr. Opin. Struct. Biol. 2013, 23, 393–402. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.Y.; Wang, C.H.; Hsiao, W.K.; Lo, H.R.; Wu, C.P.; Chao, Y.C. RING and coiled-coil domains of baculovirus IE2 are critical in strong activation of the cytomegalovirus major immediate-early promoter in mammalian cells. J. Virol. 2009, 83, 3604–3616. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chang, C.Y.; Hsu, W.T.; Chao, Y.C.; Chang, H.W. Display of Porcine Epidemic Diarrhea Virus Spike Protein on Baculovirus to Improve Immunogenicity and Protective Efficacy. Viruses 2018, 10, 346. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wei, S.C.; Hsu, W.T.; Chiu, C.H.; Chang, F.Y.; Lo, H.R.; Liao, C.Y.; Yang, H.I.; Chou, Y.C.; Tsai, C.H.; Chao, Y.C. An Integrated Platform for Serological Detection and Vaccination of COVID-19. Front. Immunol. 2021, 12, 771011. [Google Scholar] [CrossRef] [PubMed]
- Liao, C.C.; Tsai, C.H.; Lo, H.R.; Lin, P.R.; Lin, C.C.; Chao, Y.C. Development of a Scrub Typhus Diagnostic Platform Incorporating Cell-Surface Display Technology. Front. Immunol. 2021, 12, 4230. [Google Scholar] [CrossRef]
- Hsu, W.T.; Chang, C.Y.; Tsai, C.H.; Wei, S.C.; Lo, H.R.; Lamis, R.J.; Chang, H.W.; Chao, Y.C. PEDV Infection Generates Conformation-Specific Antibodies That Can Be Effectively Detected by a Cell-Based ELISA. Viruses 2021, 13, 303. [Google Scholar] [CrossRef]
- Desheva, Y.; Sychev, I.; Smolonogina, T.; Rekstin, A.; Ilyushina, N.; Lugovtsev, V.; Samsonova, A.; Go, A.; Lerner, A. Anti-neuraminidase antibodies against pandemic A/H1N1 influenza viruses in healthy and influenza-infected individuals. PLoS ONE 2018, 13, e0196771. [Google Scholar] [CrossRef]
- Couzens, L.; Gao, J.; Westgeest, K.; Sandbulte, M.; Lugovtsev, V.; Fouchier, R.; Eichelberger, M. An optimized enzyme-linked lectin assay to measure influenza A virus neuraminidase inhibition antibody titers in human sera. J. Virol. Methods 2014, 210, 7–14. [Google Scholar] [CrossRef] [Green Version]
- Krammer, F.; Palese, P. Influenza virus hemagglutinin stalk-based antibodies and vaccines. Curr. Opin. Virol. 2013, 3, 521–530. [Google Scholar] [CrossRef] [Green Version]
- Rajendran, M.; Nachbagauer, R.; Ermler, M.E.; Bunduc, P.; Amanat, F.; Izikson, R.; Cox, M.; Palese, P.; Eichelberger, M.; Krammer, F. Analysis of Anti-Influenza Virus Neuraminidase Antibodies in Children, Adults, and the Elderly by ELISA and Enzyme Inhibition: Evidence for Original Antigenic Sin. mBio 2017, 8, e02281-16. [Google Scholar] [CrossRef] [Green Version]
- Paterson, R.G.; Lamb, R.A. Conversion of a class II integral membrane protein into a soluble and efficiently secreted protein: Multiple intracellular and extracellular oligomeric and conformational forms. J. Cell. Biol. 1990, 110, 999–1011. [Google Scholar] [CrossRef]
- Yee, C.M.; Zak, A.J.; Hill, B.D.; Wen, F. The Coming Age of Insect Cells for Manufacturing and Development of Protein Therapeutics. Ind. Eng. Chem. Res. 2018, 57, 10061–10070. [Google Scholar] [CrossRef] [PubMed]
- Harrison, R.L.; Jarvis, D.L. Protein N-Glycosylation in the Baculovirus–Insect Cell Expression System and Engineering of Insect Cells to Produce “Mammalianized” Recombinant Glycoproteins. Adv. Virus Res. 2006, 68, 159. [Google Scholar] [CrossRef] [PubMed]
- Shi, X.; Jarvis, D.L. Protein N-Glycosylation in the Baculovirus-Insect Cell System. Curr. Drug Targets 2007, 8, 1116. [Google Scholar] [CrossRef] [Green Version]
- Lin, S.C.; Jan, J.T.; Dionne, B.; Butler, M.; Huang, M.H.; Wu, C.Y.; Wong, C.H.; Wu, S.C. Different Immunity Elicited by Recombinant H5N1 Hemagglutinin Proteins Containing Pauci-Mannose, High-Mannose, or Complex Type N-Glycans. PLoS ONE 2013, 8, e66719. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Vries, R.P.; Smit, C.H.; de Bruin, E.; Rigter, A.; de Vries, E.; Cornelissen, L.A.; Eggink, D.; Chung, N.P.; Moore, J.P.; Sanders, R.W.; et al. Glycan-Dependent Immunogenicity of Recombinant Soluble Trimeric Hemagglutinin. J. Virol. 2012, 86, 11735. [Google Scholar] [CrossRef] [Green Version]
- Shan, M.; Klasse, P.J.; Banerjee, K.; Dey, A.K.; Iyer, S.P.N.; Dionisio, R.; Charles, D.; Campbell-Gardener, L.; Olson, W.C.; Sanders, R.W.; et al. HIV-1 Gp120 Mannoses Induce Immunosuppressive Responses from Dendritic Cells. PLoS Pathog. 2007, 3, e169. [Google Scholar] [CrossRef] [Green Version]
- Webster, R.; Cox, N.; Stohr, K. WHO Manual on Animal Influenza Diagnosis and Surveillance; WHO/CDS/CDR/2002.5; World Health Organization; Department of Communicable Disease Surveillance and Response: Geneva, Switzerland, 2002; Volume 1, pp. 48–54. [Google Scholar]
- Playford, E.G.; Dwyer, D.E. Laboratory diagnosis of influenza virus infection. Pathology 2002, 34, 115–125. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group Score | Influenza Infected Patients |
|---|---|
| Number in group | 15 |
| Timing of sampling | 13 January 2018–9 February 2018 |
| Mean time between first and second blood draw (days) | 4.8 |
| Age (Me; Q1, Q3) | 43.00 (22.00; 50.00) |
| Male | 8 (53.3) |
| Female | 7 (46.7%) |
| 4-fold HI seroconversions to H1 | 7 (46.7%) |
| 2-fold NI seroconversions N1 | 7 (46.7%) |
| Both HI and NI seroconversions | 6 (40%) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Desheva, Y.; Petkova, N.; Losev, I.; Guzhov, D.; Go, A.; Chao, Y.-C.; Tsai, C.-H. Establishment of a Pseudovirus Platform for Neuraminidase Inhibiting Antibody Analysis. Int. J. Mol. Sci. 2023, 24, 2376. https://doi.org/10.3390/ijms24032376
Desheva Y, Petkova N, Losev I, Guzhov D, Go A, Chao Y-C, Tsai C-H. Establishment of a Pseudovirus Platform for Neuraminidase Inhibiting Antibody Analysis. International Journal of Molecular Sciences. 2023; 24(3):2376. https://doi.org/10.3390/ijms24032376
Chicago/Turabian StyleDesheva, Yulia, Nadezhda Petkova, Igor Losev, Dmitry Guzhov, Alexey Go, Yu-Chan Chao, and Chih-Hsuan Tsai. 2023. "Establishment of a Pseudovirus Platform for Neuraminidase Inhibiting Antibody Analysis" International Journal of Molecular Sciences 24, no. 3: 2376. https://doi.org/10.3390/ijms24032376