
Phycobilisomes and Phycobiliproteins in the Pigment Apparatus of Oxygenic Photosynthetics: From Cyanobacteria to Tertiary Endosymbiosis

Abstract
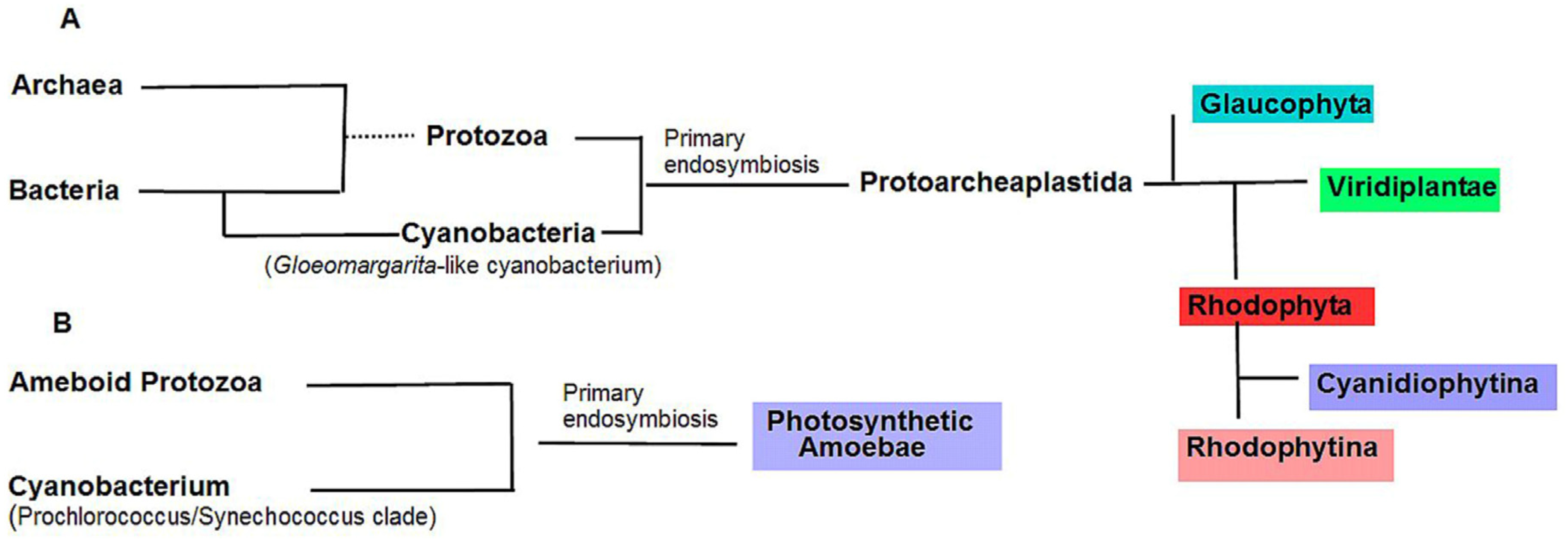
:1. Introduction
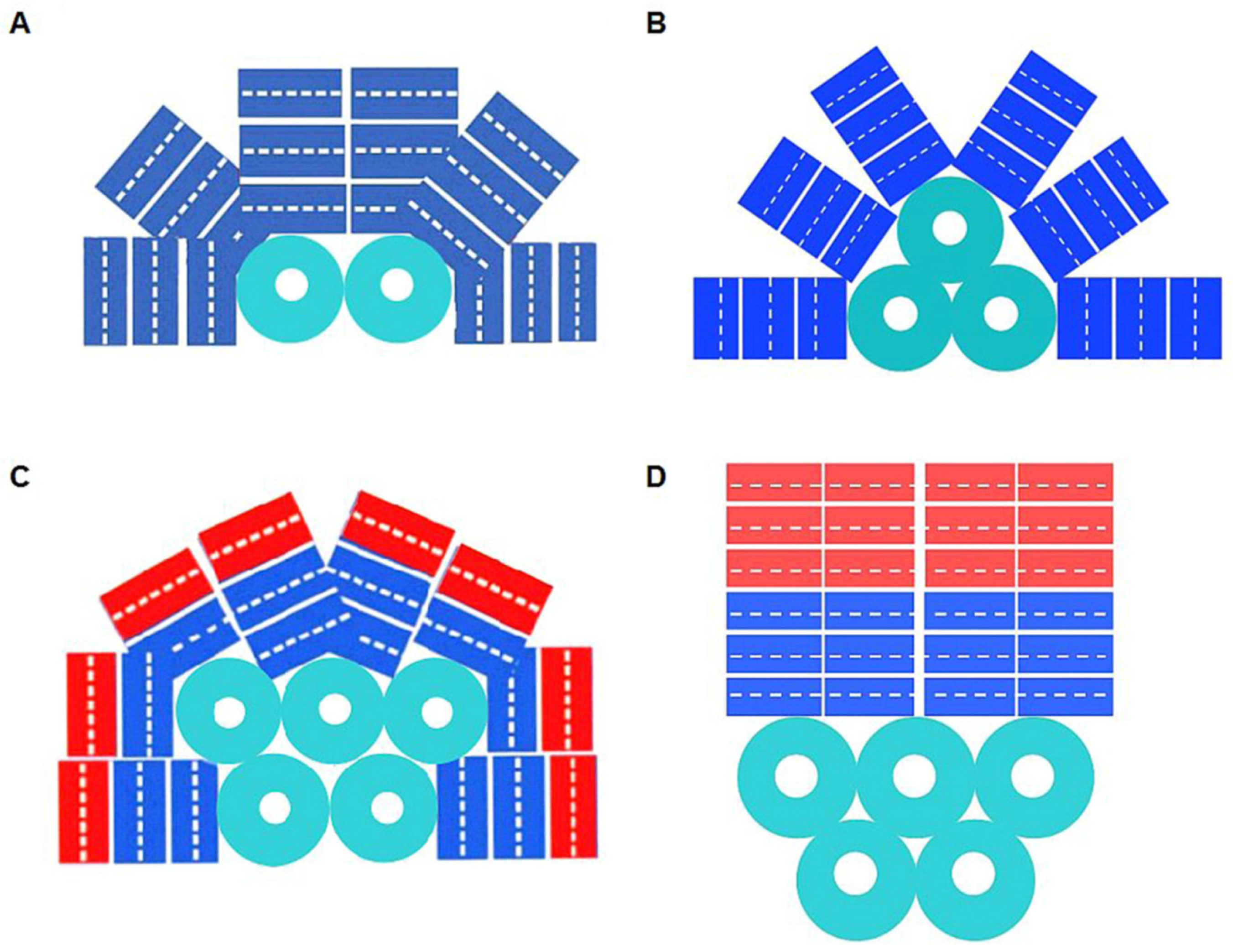
2. Phycobiliproteins and Phycobilisomes of Cyanobacteria
2.1. Prochlorophyta: Chlorophyll b-Containing Cyanobacteria
2.2. Chlorophyll d-Containing Cyanobacteria, Acaryochloris marina
2.3. Chlorophyll f-Containing Cyanobacteria and Their Involvement in the Process of Photoacclimation to Far Red Light
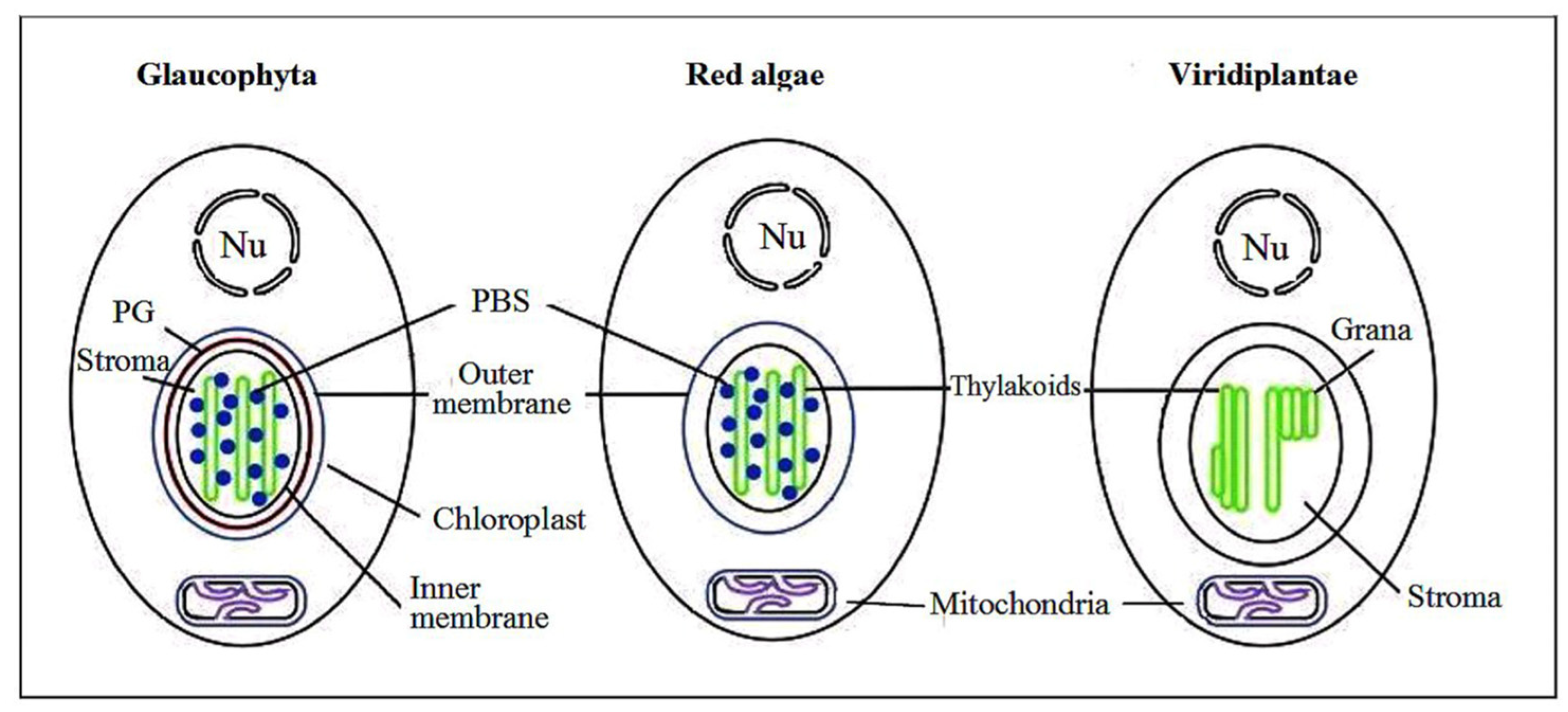
3. Red Algae Phycobilisomes
4. Glaucophytic algae
5. Photosynthetic Amoeba Paulinella
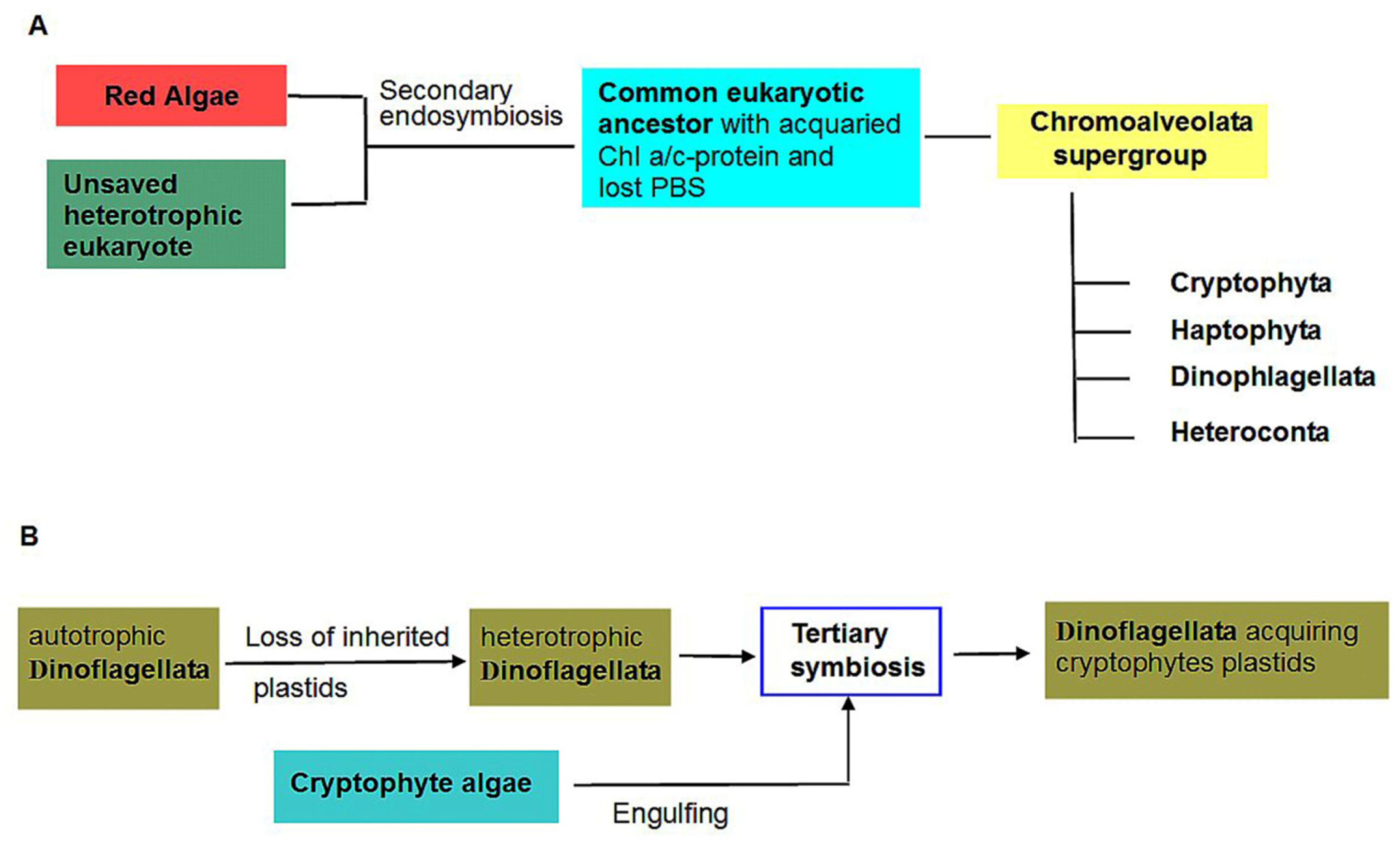
6. Cryptophyte algae
7. Tertiary Endosymbioses Involving Cryptophyte algae
8. Advantages and Disadvantages of the Phycobiliprotein Antennae and Loss of PBS
8.1. Functional Advantages of PBS over Other Antennae
8.2. Disadvantages of PBS Antennae
8.3. Role of Cell Wall Formation
9. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Schirrmeister, B.E.; Gugger, M.; Donoghue, P.C.J. Cyanobacteria and the great oxidation event: Evidence from genes and fossils. Paleontology 2015, 58, 769–785. [Google Scholar] [CrossRef] [Green Version]
- Yutin, N.; Wolf, M.Y.; Wolf, Y.I.; Koonin, E.V. The origins of phagocytosis and eukaryogenesis. Biol. Direct 2009, 4, 9. [Google Scholar] [CrossRef] [Green Version]
- Martijn, J.; Vosseberg, J.; Guy, L.; Offre, P.; Ettema, T.J.G. Deep mitochondrial origin outside the sampled alphaproteobacteria. Nature 2018, 557, 101–105. [Google Scholar] [CrossRef]
- López-Garcìa, P.; Moreira, D. The syntrophy hypothesis for the origin of eukaryotes revisited. Nat. Microbiol. 2020, 5, 655–667. [Google Scholar] [CrossRef] [PubMed]
- Stadnichuk, I.N.; Kusnetsov, V.V. Endosymbiotic origin of chloroplasts in plant cells evolution. Rus. J. Plant Physiol. 2021, 68, 1–16. [Google Scholar] [CrossRef]
- Archibald, J.M. Endosymbiosis and eukaryotic cell evolution. Curr. Biol. 2015, 25, R911–R921. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bhattacharya, D.; Qiu, H.; Lee, J.M.; Yoon, H.S.; Weber, A.P.N.; Price, D.C. When less is more: Red algae as models for studying gene loss and genome evolution in eukaryotes. Crit. Rev. Plant Sci. 2018, 37, 81–99. [Google Scholar] [CrossRef]
- Figueroa-Martinez, F.; Christopher, J.; Reyes-Prieto, A. Plastid genomes from diverse Glaucophyte genera reveal a largely conserved gene content and limited architectural diversity. Genome Biol. Evol. 2019, 11, 174–188. [Google Scholar] [CrossRef] [Green Version]
- Donoghue, P.; Paps, J. Plant evolution: Assembling land plants. Curr. Biol. 2020, 30, R81–R83. [Google Scholar] [CrossRef]
- Cavalier-Smith, T. Kingdom Chromista and its eight phyla: A new synthesis emphasising periplastid protein targeting, cytoskeletal and periplastid evolution, and ancient divergences. Protoplasma 2018, 255, 297–357. [Google Scholar] [CrossRef] [Green Version]
- Stadnichuk, I.N. Phycobiliproteins: Determination of chromophore composition and content. Phytochem. Analysis 1995, 6, 281–288. [Google Scholar] [CrossRef]
- Stadnichuk, I.N.; Tropin, I.V. Phycobiliproteins: Structure, functions and biotechnological applications. Appl. Biochem. Microbiol. 2017, 53, 1–10. [Google Scholar] [CrossRef]
- Betz, M. One century of protein crystallography: The phycobiliproteins. Biol. Chem. 1997, 378, 167–176. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blot, N.; Wu, X.-J.; Thomas, J.-C.; Zhang, J.; Garczarek, L.; Böhm, S.; Tu, J.-M.; Zhou, M.; Plöscher, M.; Eichacker, L.; et al. Phycourobilin in trichromatic phycocyanin from oceanic cyanobacteria is formed post-translationally by a phycoerythrobilin lyase-isomerase. J. Biol. Chem. 2009, 284, 9290–9298. [Google Scholar] [CrossRef] [Green Version]
- Bernát, G.; Schreiber, U.; Sendtko, E.; Stadnichuk, I.N.; Rexroth, S.; Rögner, M.; Koenig, F. Unique properties vs. common themes: The atypical cyanobacterium Gloeobacter violaceus PCC 7421 is capable of state transitions and blue-light induced fluorescence quenching. Plant Cell Physiol. 2012, 53, 528–542. [Google Scholar] [CrossRef]
- Kondo, K.; Ochiai, Y.; Katayama, M.; Ikeuchi, M. The membrane-associated CpcG2-phycobilisome in Synechocystis: A new photosystem I antenna. Plant Physiol. 2007, 144, 1200–1210. [Google Scholar] [CrossRef] [Green Version]
- Elanskaya, I.V.; Zlenko, D.V.; Lukashev, E.P.; Suzina, N.E.; Kononova, I.A.; Stadnichuk, I.N. Phycobilisomes from the mutant cyanobacterium Synechocystis sp. PCC 6803 missing chromophore domain of ApcE. Biochim. Biophys. Acta (BBA)-Bioenerg. 2018, 1859, 280–291. [Google Scholar] [CrossRef]
- Grébert, T.; Garczarek, L.; Daubin, V.; Humily, F.; Marie, D.; Ratin, M.; Devailly, A.; Farrant, G.K.; Mary, I.; Mella-Flores, D.; et al. Diversity and evolution of pigment types in marine Synechococcus cyanobacteria. Genome Biol. Evol. 2022, 14, evac035. [Google Scholar] [CrossRef] [PubMed]
- Gindt, Y.M.; Zhou, J.; Bryant, D.A.; Sauer, K. Spectroscopic studies of phycobilisome subcore preparations lacking key core chromophores: Assignment of excited state energies to the Lcm, β18 and αAP-B chromophores. Biochim. Biophys. Acta (BBA)-Bioenerg. 1994, 1186, 153–162. [Google Scholar] [CrossRef]
- Dong, C.; Tang, A.; Zhao, J.; Mullineaux, C.W.; Shen, G.; Bryant, D.A. ApcD is necessary for efficient energy transfer from phycobilisomes to photosystem I and helps to prevent photoinhibition in the cyanobacterium Synechococcus sp. PCC 7002. Biochim. Biophys. Acta 2009, 1787, 1122–1128. [Google Scholar] [CrossRef] [Green Version]
- Zheng, L.; Zheng, Z.; Li, X.; Wang, G.; Zhang, K.; Wei, P.; Zhao, J.; Ga, N. Structural insight into the mechanism of energy transfer in cyanobacterial phycobilisomes. Nat. Commun. 2021, 2, 5497. [Google Scholar] [CrossRef] [PubMed]
- Bald, D.; Kruip, J.; Rögner, M. Supramolecular architecture of cyanobacterial thylakoid membranes: How is the phycobilisome connected with the photosystems? Photosynth. Res. 1996, 49, 103–118. [Google Scholar] [CrossRef] [PubMed]
- Rakhimberdieva, M.G.; Boichenko, V.A.; Karapetyan, N.V.; Stadnichuk, I.N. Interaction of phycobilisomes with photosystem II dimers and photosystem I monomers and trimers in the cyanobacterium Spirulina platensis. Biochemistry 2001, 40, 15780–15788. [Google Scholar] [CrossRef]
- Bryant, D.A.; Canniffe, D.P. How nature designs light-harvesting antenna systems: Design principles and functional realization in chlorophototrophic prokaryotes. J. Phys. B At. Mol. Opt. Phys. 2018, 51, 033001. [Google Scholar] [CrossRef]
- Arteni, A.A.; Ajlani, G.; Boekema, E.J. Structural organization of phycobilisomes from Synechocystis strain PCC 6803 and their interaction with the membrane. Biochim. Biophys. Acta 2009, 1787, 272–279. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chang, L.; Liu, X.; Li, Y.; Liu, C.; Yang, F. Structural organization of an intact phycobilisome and its association with photosystem II. Cell Res. 2015, 25, 726–737. [Google Scholar] [CrossRef] [Green Version]
- Chitnis, V.; Chitnis, P. Psal subunit is required for the formation of photosystem I trimers in the cyanobacterium Synechocystis sp. PCC 6803. FEBS Lett. 1993, 336, 330–334. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kruip, J.; Chitnis, P.; Lagoutte, B.; Rögner, M.; Boekema, E. Structural organization of the major subunits in cyanobacterial photosystem I. Localization of subunits PsaC, -D, -E, -F, and –J. J. Biol. Chem. 1997, 272, 17061–17069. [Google Scholar] [CrossRef] [Green Version]
- Zlenko, D.V.; Krasilnikov, P.M.; Stadnichuk, I.N. Structural modeling of the phycobilisome core and its association with the photosystems. Photosynth. Res. 2016, 130, 347–356. [Google Scholar] [CrossRef]
- Burger-Wiersma, T.; Post, A.F. Functional Analysis of the photosynthetic apparatus of Prochlorothrix hollandica (Prochlorales), a chlorophyll b containing procaryote. Plant Physiol. 1989, 91, 770–774. [Google Scholar] [CrossRef] [Green Version]
- Averina, S.G.; Velichko, N.V.; Pinevich, A.A.; Senatskaya, E.V.; Pinevich, A.V. Non-a chlorophylls in cyanobacteria. Photosynthetica 2019, 57, 1109–1118. [Google Scholar] [CrossRef] [Green Version]
- Lewin, R.A. A marine Synechocystis (Cyanophyta, Chroococcales) epizoic on ascidians. Phycologia 1975, 14, 153–160. [Google Scholar] [CrossRef]
- La Roche, J.; Van der Staay, G.; Partensky, F.; Ducret, A.; Aebersold, R.; Li, R.; Golden, S.S.; Hiller, R.; Wrench, P.M.; Larkum, A.W.D.; et al. Independent evolution of the prochlorophyte and green plant chlorophyll a/b light harvesting proteins. Proc. Natl. Acad. Sci. USA 1996, 93, 15244–15248. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, M.; Li, Y.Q.; Birch, D.; Willows, R.D. A cyanobacterium that contains chlorophyll f-a red-absorbing photopigment. FEBS Lett. 2012, 586, 3249–3254. [Google Scholar] [CrossRef] [Green Version]
- Giddings, T.H., Jr.; Withers, N.W.; Staehelin, L.A. Supramolecular structure of stacked and unstacked regions of the photosynthetic membranes of Prochloron sp., a prokaryote. Proc. Natl. Acad. Sci. USA 1980, 77, 352–356. [Google Scholar] [CrossRef] [Green Version]
- Boichenko, V.A.; Pinevich, A.V.; Stadnichuk, I.N. Association of chlorophyll a/b-binding Pcb proteins with photosystems I and II in Prochlorothrix hollandica. Biochim. Biophys. Acta (BBA)-Bioenerg. 2007, 1767, 801–806. [Google Scholar] [CrossRef] [Green Version]
- Barrera-Rojas, J.; González de la Vara, L.E.; Ríos-Castro, E. The distribution of divinyl chlorophylls a and b and the presence of ferredoxin-NADP+ reductase in Prochlorococcus marinus MIT9313 thylakoid membranes. Helion 2018, 4, e01100. [Google Scholar] [CrossRef] [Green Version]
- Larkum, A.W.D.; Scaramuzzi, C.; Cox, G.C.; Hiller, R.G.; Turner, A.G. Light harvesting chlorophyll c-like pigment in Prochloron. Proc. Natl. Acad. Sci. USA 1994, 91, 678–683. [Google Scholar] [CrossRef] [Green Version]
- Hess, W.R.; Partensky, F.; Van der Staay, G.W.M.; Garcia-Fernandez, J.M.; Börner, T.; Vaulot, D. Coexistence of phycoerythrin and a chlorophyll a/b antenna in a marine prokaryote. Proc. Natl. Acad. Sci. USA 1996, 93, 11126–11130. [Google Scholar] [CrossRef] [Green Version]
- Penno, S.; Campbell, L.; Hess, W.R. Presence of phycoerythrin in two strains of Prochlorococcus isolated from the sub-tropical North Pacific Ocean. J. Phycol. 2000, 36, 723–729. [Google Scholar] [CrossRef]
- Beale, S.I. Photosynthetic pigments: Perplexing persistent prevalence of ‘superfluous’ pigment production. Curr. Biol. 2008, 18, R342–R343. [Google Scholar] [CrossRef] [Green Version]
- Moore, L.R.; Rocap, G.; Chisholm, S.W. Physiology and molecular phylogeny of coexisting Prochlorococcus ecotypes. Nature 1998, 393, 464–467. [Google Scholar] [CrossRef] [PubMed]
- Wiethaus, J.; Busch, A.W.U.; Dammeyer, T.; Frankenberg-Dinkel, N. Phycobiliproteins in Prochlorococcus marinus: Biosynthesis of pigments and their assembly into proteins. Eur. J. Cell Biol. 2010, 89, 1005–1010. [Google Scholar] [CrossRef]
- Hess, W.R.; Steglich, C.; Lichtlé, C.; Partensky, F. Phycoerythrins of the oxyphotobacterium Prochlorococcus marinus are associated to the thylakoid membrane and are encoded by a single large gene cluster. Plant Mol. Biol. 1999, 40, 507–521. [Google Scholar] [CrossRef] [PubMed]
- Ting, C.S.; Rocap, G.; King, J.; Chisholm, S.W. Phycobiliprotein genes of the marine photosynthetic prokaryote Prochlorococcus: Evidence for rapid evolution of genetic heterogeneity. Microbiology 2001, 147, 3171–3182. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sonani, R.R.; Rastogi, R.P.; Joshi, M.; Madamwar, D. A stable and functional single peptide phycoerythrin (15.45 kDa) from Lyngbya sp. A09DM. Int. J. Biol. Macromol. 2015, 74, 29–35. [Google Scholar] [CrossRef] [PubMed]
- Ikeuchi, M.; Ishizuka, T. Cyanobacteriochromes: A new superfamily of tetrapyrrole-binding photoreceptors in cyanobacteria. Photochem. Photobiol. Sci. 2008, 7, 1159–1167. [Google Scholar] [CrossRef] [PubMed]
- Miyashita, H.; Ikemoto, H.; Kurano, N.; Adachi, K.; Chihara, M.; Miyachi, S. Chlorophyll d as a major pigment. Nature 1996, 383, 402. [Google Scholar] [CrossRef]
- Miller, S.R.; Abresch, H.E.; Baroch, J.J.; Miller, C.K.F.; Garber, A.I.; Oman, A.R.; Ulrich, N.J. Genomic and functional variation of the chlorophyll d-producing cyanobacterium Acaryochloris marina. Microorganisms 2022, 10, 569. [Google Scholar] [CrossRef]
- Tomo, T.; Okubo, T.; Akimoto, S.; Yokono, M.; Miyashita, H.; Tsuchiya, T.; Noguchi, T.; Mimuro, M. Identification of the special pair of photosystem II in a chlorophyll d-dominated cyanobacterium. Proc. Natl. Acad. Sci. USA 2007, 104, 7283–7288. [Google Scholar] [CrossRef] [Green Version]
- Chen, M.; Quinnell, R.G.; Larkum, A.W.D. The major light-harvesting pigment protein of Acaryochloris marina. FEBS Lett. 2002, 514, 149–152. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamamoto, H.; Uesaka, K.; Tsuzuki, Y.; Yamakawa, H.; Itoh, S.; Fujita, Y. Comparative genomic analysis of the marine cyanobacterium Acaryochloris marina MBIC10699 reveals the impact of phycobiliprotein reacquisition and the diversity of Acaryochloris plasmids. Microorganisms 2022, 10, 1374. [Google Scholar] [CrossRef] [PubMed]
- Theiss, C.; Schmitt, F.-J.; Pieper, J.; Nganou, C.; Grehna, M.; Vitali, M.; Olliges, R.; Eichler, H.J.; Eckert, H.-J. Excitation energy transfer in intact cells and in the phycobiliprotein antennae of the chlorophyll d containing cyanobacterium Acaryochloris marina. J. Plant Physiol. 2011, 168, 1473–1487. [Google Scholar] [CrossRef]
- Marquardt, J.; Senger, H.; Miyashita, H.; Miyachi, S.; Mörschel, E. Isolation and characterization of biliprotein aggregates from Acaryochloris marina, a Prochloron-like prokaryote containing mainly chlorophyll d. FEBS Lett. 1997, 410, 428–432. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, Q.; Marquardt, J.; Iwasaki, I.; Miyashita, H.; Kurano, N.; Mörschel, E.; Miyachi, S. Molecular structure, localization and function of biliproteins in the chlorophyll a/d containing oxygenic photosynthetic prokaryote Acaryochloris marina. Biochim. Biophys. Acta 1999, 1412, 250–261. [Google Scholar] [CrossRef] [Green Version]
- Boichenko, V.A.; Klimov, V.V.; Miyashita, H.; Miyachi, S. Functional characteristics of chlorophyll d-predominating photosynthetic apparatus in intact cells of Acaryochloris marina. Photosynth. Res. 2000, 65, 269–277. [Google Scholar] [CrossRef]
- Ulrich, N.J.; Uchida, H.; Kanesaki, Y.; Hirose, E.; Murakami, A.; Miller, S.R. Reacquisition of light-harvesting genes in a marine cyanobacterium confers a broader solar niche. Curr. Biol. 2021, 31, 1539–1546. [Google Scholar] [CrossRef] [PubMed]
- Ho, M.-Y.; Soulier, N.T.; Canniffe, D.P.; Shen, G.; Bryant, D.A. Light regulation of pigment and photosystem biosynthesis in cyanobacteria. Curr. Opin. Plant Biol. 2017, 37, 24–33. [Google Scholar] [CrossRef] [Green Version]
- Gan, F.; Zhang, S.; Rockwell, N.C.; Martin, S.S.; Lagarias, J.C.; Bryant, D.A. Extensive remodeling of a cyanobacterial photosynthetic apparatus in far-red light. Science 2014, 345, 1312–1317. [Google Scholar] [CrossRef] [Green Version]
- Larkum, A.W.D.; Ritchie, R.J.; Raven, J.A. Living off the Sun: Chlorophylls, bacteriochlorophylls and rhodopsins. Photosynthetica 2018, 56, 11–43. [Google Scholar] [CrossRef]
- Gisriel, C.J.; Shen, G.; Ho, M.-Y.; Kurashov, V.; Fleshe, D.A.; Wang, J.; Armstrong, W.H.; Golbeck, J.H.; Gunner, M.R.; Vinyard, D.J.; et al. Structure of a monomeric photosystem II core complex from a cyanobacterium acclimated to far-red light reveals the functions of chlorophylls d and f. J. Biol. Chem. 2022, 298, 101424. [Google Scholar] [CrossRef]
- Kato, K.; Shinoda, T.; Nagao, R.; Akimoto, S.; Suzuki, T.; Dohmae, N.; Chen, M.; Allakhverdiev, S.I.; Shen, J.-R.; Akita, F.; et al. Structural basis for the adaptation and function of chlorophyll f in photosystem I. Nat. Commun. 2020, 11, 238. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Antonaru, L.A.; Cardona, T.; Larkum, A.W.D.; Nürnberg, D.J. Global distribution of a chlorophyll f cyanobacterial marker. ISME J. 2020, 14, 2275–2287. [Google Scholar] [CrossRef] [PubMed]
- Stadnichuk, I.N.; Tropin, I.V. Cyanidiales as polyextreme eukaryotes. Biochemistry 2022, 87, 472–487. [Google Scholar] [CrossRef]
- Koller, K.P.; Wehrmeyer, W.; Mörschel, E. Biliprotein assembly in the discshaped phycobilisomes of Rhodella violacea. Eur. J. Biochem. 1978, 91, 57–63. [Google Scholar] [CrossRef] [PubMed]
- Glazer, A.N.; West, J.A.; Chan, C. Phycoerythrins as chemotaxonomic markers in red algae: A survey. Biochem. System. Ecol. 1982, 10, 203–215. [Google Scholar] [CrossRef]
- Arteni, A.A.; Liu, L.-N.; Aartsma, T.J.; Zhang, Z.Y.; Zhou, B.C.; Boekema, E.J. Structure and organization of phycobilisomes on membranes of the red alga Porphyridium cruentum. Photosynth. Res. 2008, 95, 169–174. [Google Scholar] [CrossRef] [Green Version]
- Gantt, E.; Lipschultz, C.A. Structure and phycobiliprotein composition of phycobilisomes from Griffitshia pacifica (Rhodophyceae). J. Phycol. 1980, 16, 394–398. [Google Scholar] [CrossRef]
- Ma, J.; You, X.; Sun, S.; Wang, X.; Qin, S.; Sui, S.F. Structural basis of energy transfer in Porphyridium purpureum phycobilisome. Nature 2020, 579, 146–151. [Google Scholar] [CrossRef]
- Zhang, J.; Ma, J.; Liu, D.; Qin, S.; Sun, S.; Zhao, J.; Sui, S.-F. Structure of phycobilisome from the red alga Griffitshia pacifica. Nature 2017, 551, 57–63. [Google Scholar] [CrossRef]
- Ago, H.; Adachi, H.; Umena, Y.; Tashiro, T.; Kewakami, K.; Kamiya, L.; Tian, G.; Han, T.; Liu, Z.; Wang, F.; et al. Novel features of eukaryotic photosystem II revealed by its crystal structure analysis from a red alga. J. Biol. Chem. 2016, 291, 5676–5687. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Antoshvili, M.; Caspy, I.; Hippler, M.; Nelson, N. Structure and function of photosystem I in Cyanidioschyzon merolae. Photosynth. Res. 2019, 139, 499–508. [Google Scholar] [CrossRef] [PubMed]
- Schenk, H.E.A. Cyanophora paradoxa: Anagenetic model or missing link of plastid evolution? Endocytobiosis Cell Res. 1993, 10, 87–106. [Google Scholar]
- Price, D.C.; Steiner, J.M.; Yoon, H.S.; Bhattacharya, D.; Löffelhard, W. Glaucophyta. In Handbook of the Protists; Archibald, J.M., Simpson, A.G.B., Slamovits, C.H., Margulis, L., Melkonian, M., Chapman, D.J., Corliss, J.O., Eds.; Springer International Publishing AG: Cham, Switzerland, 2016; pp. 1–65. [Google Scholar] [CrossRef]
- Jackson, C.; Clayden, S.; Reyes-Prieto, A. The Glaucophyta: The blue-green plants in a nutshell. Acta Soc. Bot. Pol. 2015, 84, 149–165. [Google Scholar] [CrossRef] [Green Version]
- Ball, S.; Colleoni, C.; Cenci, U.; Raj, J.N.; Tirtiaux, C. The evolution of glycogen and starch metabolism in eukaryotes gives molecular clues to understand the establishment of plastid endosymbiosis. J. Exp. Bot. 2011, 62, 1775–1801. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Löffelhardt, W.; Bohnert, H.J. The cyanelle (muroplast) of Cyanophora paradoxa: A paradigm for endosymbiotic organelle evolution. In Symbiosis. Mechanisms and Model Systems. Cellular Origin, Life in Extreme Habitats and Astrobiology; Seckbach, J., Ed.; Kluwer Academic Publishers: Dordrecht, The Netherlands, 2001; Volume 4, pp. 111–130. [Google Scholar]
- Steiner, J.M.; Löffelhardt, W. Protein translocation into and within cyanelles (review). Mol. Membr. Biol. 2005, 22, 123–132. [Google Scholar] [CrossRef] [PubMed]
- Kovacevic, G.; Steiner, J.M.; Löffelhardt, W. Cyanobacterial and algal symbioses. In Intracellular Niches of Microbes; Schaible, U.E., Haas, A., Eds.; Wiley-VCH Verlag: Weinheim, Germany, 2009; pp. 527–545. [Google Scholar]
- Floener, L.; Bothe, H. Metabolic activities in Cyanophora paradoxa and its cyanelles: II. Photosynthesis and respiration. Planta 1982, 156, 78–83. [Google Scholar] [CrossRef]
- Burnap, R.L.; Trench, R.K. The biogenesis of the cyanellae of Cyanophora paradoxa. I. Polypeptide composition of the cyanellae. Proc. R. Soc. Lond. B 1989, 238, 53–72. [Google Scholar] [CrossRef]
- Shibata, M.; Kashino, Y.; Satoh, K.; Koike, H. Isolation and characterization of oxygen-evolving thylakoid membranes and photosystem II particles from a glaucocystophyte, Cyanophora paradoxa. Plant Cell Physiol. 2001, 42, 733–741. [Google Scholar] [CrossRef] [Green Version]
- Bhattacharya, D.; Price, D.C.; Chan, C.X.; Gross, J. Analysis of the genome of Cyanophora paradoxa: An algal model for understanding primary endosymbiosis. In Endosymbiosis; Löffelhardt, W., Ed.; Springer: Vienna, Austria, 2014; pp. 135–148. [Google Scholar]
- Koike, H.; Shibata, M.; Yasutomi, K.; Kashino, Y.; Satoh, K. Identification of Photosystem I components from a glaucocystophyte, Cyanophora paradoxa: The PsaD protein has an N-terminal stretch homologous to higher plants. Photosynth. Res. 2000, 65, 207–217. [Google Scholar] [CrossRef]
- Watanabe, M.; Kubota, H.; Wada, H.; Narikawa, R.; Ikeuchi, M. Novel supercomplex organization of photosystem I in Anabaena and Cyanophora paradoxa. Plant Cell Physiol. 2011, 52, 162–168. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kato, K.; Nagao, R.; Ueno, Y.; Yokono, M.; Suzuki, T.; Jiang, T.-Y.; Dohmae, N.; Akita, F.; Akimoto, S.; Miyazaki, N.; et al. Structure of a tetrameric photosystem I from a glaucophyte alga Cyanophora paradoxa. Nat. Commun. 2022, 13, 1679. [Google Scholar] [CrossRef] [PubMed]
- Giddings, T.H.; Wasmann, C.; Staehelin, L.A. Structure of the thylakoids and envelope membranes of the cyanelles of Cyanophora paradoxa. Plant Physiol. 1983, 71, 409–419. [Google Scholar] [CrossRef] [Green Version]
- Watanabe, M.; Sato, M.; Kondo, K.; Narikawa, R.; Ikeuchi, M. Phycobilisome model with novel skeleton-like structures in a glaucocystophyte Cyanophora paradoxa. Biochim. Biophys. Acta (BBA)-Bioenerg. 2012, 1817, 1428–1435. [Google Scholar] [CrossRef] [Green Version]
- Nicholls, K.H. Six new marine species of the genus Paulinella (Rhizopoda: Filosea, or Rhizaria: Cercozoa). J. Mar. Biol. Assoc. UK 2009, 89, 1415–1425. [Google Scholar] [CrossRef]
- Kies, L.; Kremer, B.P. Function of cyanelles in thecamoeba Paulinella chromatophora. Naturwissenschaften 1979, 66, 578–579. [Google Scholar] [CrossRef]
- Douzery, E.J.P.; Snell, E.A.; Bapteste, E.; Delsuc, F.; Philippe, H. The timing of eukaryotic evolution: Does a relaxed molecular clock reconcile proteins and fossils? Proc. Natl. Acad. Sci. USA 2004, 101, 15836–15839. [Google Scholar] [CrossRef] [Green Version]
- Nowack, E.C.M. Paulinella chromatophora—Rethinking the transition from endosymbiont to organelle. Acta. Soc. Bot. Pol. 2014, 83, 387–397. [Google Scholar] [CrossRef] [Green Version]
- Delaye, L.; Valadez-Cano, C.; Perez-Zamorano, B. How really ancient is Paulinella chromatophora? PLoS Curr. 2016, 8. [Google Scholar] [CrossRef]
- Gabr, A.; Grossman, A.R.; Bhattacharya, D. Paulinella, a model for understanding plastid primary endosymbiosis. J. Phycol. 2020, 56, 837–843. [Google Scholar] [CrossRef]
- Burki, F.; Roge, A.J.; Brown, M.W.; Simpson, G.B. The new tree of eukaryotes. Trends Ecol. Evol. 2019, 35, 43–55. [Google Scholar] [CrossRef] [Green Version]
- Lhee, D.; Lee, J.; Ettahi, K.; Cho, C.H.; Ha, J.-S.; Chan, Y.-F.; Zelzion, U.; Stephens, T.G.; Price, D.C.; Gabr, A.; et al. Amoeba genome reveals dominant host contribution to plastid endosymbiosis. Mol. Biol. Evol. 2021, 38, 344–357. [Google Scholar] [CrossRef] [PubMed]
- Zhang, R.; Nowack, E.C.; Price, D.C.; Bhattacharya, D.; Grossman, A.R. Impact of light intensity and quality on chromatophore and nuclear gene expression in Paulinella chromatophora, an amoeba with nascent photosynthetic organelles. Plant J. 2017, 90, 221–234. [Google Scholar] [CrossRef] [Green Version]
- Hill, D.R.; Rowan, K.S. The biliproteins of the cryptophyceae. Phycologia 1989, 28, 455–463. [Google Scholar] [CrossRef]
- Glazer, A.N.; Wedemayer, G.J. Cryptomonad biliproteins: An evolutionary perspective. Photosynth. Res. 1995, 46, 93–105. [Google Scholar] [CrossRef] [PubMed]
- Sidler, W.A. Phycobilisome and phycobiliprotein structures. In The Molecular Biology of Cyanobacteria. Advances in Photosynthesis; Bryant, D.A., Ed.; Kluwer Academic Publishers: Dordrecht, The Netherlands, 1994; Volume 1, pp. 39–216. [Google Scholar] [CrossRef]
- Toole, C.; Allnut, F.C.T. Red, cryptomonad and glaucocystophyte algal phycobiliproteins. In Photosynthesis in Algae; Larkum, A.W.D., Douglas, S.E., Raven, J.A., Eds.; Kluwer Academic Publishers: Dordrecht, The Netherlands, 2003; pp. 305–334. [Google Scholar]
- Hoef-Emden, K.; Archibald, J.M. Cryptophyta (Cryptomonads). In Handbook of the Protists, 2nd ed.; Archibald, J.M., Simpson, A.G.B., Slamovitz, C.H., Eds.; Springer: Cham, Switzerland, 2017; pp. 851–891. [Google Scholar] [CrossRef]
- Cunningham, B.R.; Greenwold, M.J.; Lachenmyer, E.M.; Heidenreich, K.M.; Davis, A.C.; Dudycha, J.L.; Richardson, T.L. Light capture and pigment diversity in marine and freshwater cryptophytes. J. Phycol. 2019, 55, 552–564. [Google Scholar] [CrossRef] [PubMed]
- Richardson, T.L. The colorful world of cryptophyte phycobiliproteins. J. Plankton 2022, 44, 806–818. [Google Scholar] [CrossRef]
- Lora-Vilchis, M.C. Cryptophyte: Biology, Culture, and Biotechnological Applications. In Progress in Microalgae Research—A Path for Shaping Sustainable Futures; Zepka, L.Q., Jacob-Lopes, E., Deprá, M.C., Eds.; IntechOpen: Rijeka, Croatia, 2022; Volume 53. [Google Scholar] [CrossRef]
- Neilson, J.A.D.; Durnford, D.G. Structural and functional diversification of the light-harvesting complexes in photosynthetic eukaryotes. Photosynth. Res. 2010, 106, 57–71. [Google Scholar] [CrossRef]
- Hoffman, G.E.; Sanchez-Puerta, M.V.; Delwiche, C.F. Evolution of light harvesting complex proteins from Chl c-containing algae. BMC Evol. Biol. 2011, 11, 101. [Google Scholar] [CrossRef] [Green Version]
- Corbella, M.; Cupellini, L.; Lipparini, F.; Scholes, G.D.; Curutchet, C. Spectral variability in phycocyanin cryptophyte antenna complexes is controlled by changes in the α-polypeptide chains. ChemPhotoChem 2019, 3, 945–956. [Google Scholar] [CrossRef]
- Greenwold, M.J.; Cunningham, B.R.; Lachenmyer, E.M.; Pullman, J.M.; Richardson, T.L.; Dudycha, J.L. Diversification of light capture ability was accompanied by the evolution of phycobiliproteins in cryptophyte algae. Proc. R. Soc. B 2019, 286, 20190655. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mörschel, E.; Wehrmeyer, W. Elektronen-mikroskopische Feinstrukturanalyse von nativen Biliproteidaggregaten und deren räumliche Ordnung. Ber. Deutsch. Bot. Ges. 1979, 92, 393–402. [Google Scholar] [CrossRef]
- Vesk, M.; Dwarte, D.; Fowler, S.; Hiller, R.G. Freeze fracture immunocytochemistry of light-harvesting pigment complexes in a cryptophyte. Protoplasma 1992, 170, 166–176. [Google Scholar] [CrossRef]
- Rathbone, H.W.; Michie, K.A.; Landsberg, M.J.; Green, B.R.; Curmi, P.M.G. Scaffolding proteins guide the evolution of algal light harvesting proteins. Nat. Commun. 2012, 12, 1890. [Google Scholar] [CrossRef]
- Apt, K.E.; Collier, J.L.; Grossman, A.R. Evolution of the phycobiliproteins. J. Mol. Biol. 1995, 248, 79–96. [Google Scholar] [CrossRef]
- Broughton, M.J.; Howe, C.J.; Hiller, R.G. Distinctive organization of genes for light harvesting proteins in the cryptophyte alga Rhodomonas. Gene 2006, 369, 72–79. [Google Scholar] [CrossRef]
- Kieselbach, T.; Cheregi, O.; Green, B.R.; Funk, C. Proteomic analysis of the phycobiliprotein antenna of the cryptophyte alga Guillardia theta cultured under different light intensities. Photosynth. Res. 2018, 135, 149–163. [Google Scholar] [CrossRef] [PubMed]
- Harrop, S.J.; Wilka, K.E.; Dinshaw, R.; Collinic, E.; Mirkovic, T.; Teng, C.Y.; Oblinsky, D.G.; Green, B.R.; Hoef-Emden, K.; Hiller, R.G.; et al. Single-residue insertion switches the quaternary structure and exciton states of cryptophytes light-harvesting proteins. Proc. Natl. Acad. Sci. USA 2014, 111, E2666–E2675. [Google Scholar] [CrossRef] [Green Version]
- Ludwig, M.; Gibbs, S.P. Localization of phycoerythrin at the lumenal surface of the thylakoid membrane in Rhodomonas lens. J. Cell Biol. 1989, 108, 875884. [Google Scholar] [CrossRef] [PubMed]
- Kereiche, S.; Kouřil, R.; Oostergetel, G.T.; Fusetti, F.; Boekema, E.J.; Doust, A.B.; Van der Weij-de Wit, C.D.; Dekker, J.P. Association of chlorophyll a/c2 complexes to photosystem I and photosystem II in the cryptophyte Rhodomonas CS24. Biochim. Biophys. Acta 2008, 1777, 1122–1128. [Google Scholar] [CrossRef] [Green Version]
- Kuthanová-Trsková, E.; Bína, D.; Santabarbara, S.; Sobotka, R.; Kaňa, R.; Belgio, E. Isolation and characterization of CAC antenna proteins and photosystem I supercomplex from the cryptophytic alga Rhodomonas salina. Physiol. Plant 2019, 166, 309–319. [Google Scholar] [CrossRef] [PubMed]
- Bruce, D.; Biggins, J.; Steiner, T.; Thewalt, M. Excitation energy transfer in the cryptophytes. Fluorescence excitation spectra and picosecond time-resolved emission spectra of intact algae at 77 K. Photochem. Photobiol. 1986, 44, 519–525. [Google Scholar] [CrossRef]
- Wedemayer, G.J.; Wemmer, D.E.; Glazer, A.N. Phycobilins of cryptophycean algae: Structures of novel bilins with acryloyl substituents from phycoerythrin 566. J. Biol. Chem. 1991, 266, 4731–4741. [Google Scholar] [CrossRef]
- MacColl, R.; Eisele, L.E.; Dhar, M.; Ecuyer, J.-P.; Hopkins, S.; Marrone, J.; Barnard, R.; Malak, H.; Lewitus, A.J. Bilin organization on cryptomonad biliproteins. Biochemistry 1999, 28, 4097–4105. [Google Scholar] [CrossRef] [PubMed]
- Van der Weij-De Wit, C.D.; Doust, A.B.; Van Stokkum, I.H.M.; Dekker, J.P.; Wilk, K.E.; Curmi, P.M.G.; Scholes, G.D.; Van Grondelle, R. How energy funnels from the phycoerythrin antenna complex to photosystem I and photosystem II in cryptophytes Rhodomonas CS24 cells. J. Phys. Chem. B 2006, 110, 25066–25073. [Google Scholar] [CrossRef] [PubMed]
- Van der Weij-De Wit, C.D.; Doust, A.B.; van Stokkum, I.H.M.; Dekker, J.P.; Wilk, K.E.; Curmi, P.G.; van Grondelle, R. Phycocyanin Sensitizes both Photosystem I and Photosystem II in Cryptophyte Chroomonas CCMP270 Cells. Biophys. J. 2008, 94, 2423–2433. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stadnichuk, I.N. Role of phycobiliprotein antenna of cyanobacteria, red and cryptophyte algae in association with photosystems I and II. J. Plant Biochem. Physiol. 2020, 8, 1–4. [Google Scholar] [CrossRef]
- Stadnichuk, I.N.; Novikova, T.M.; Miniuk, G.S.; Boichenko, V.A.; Bolychevtseva, Y.V.; Gusev, E.S.; Lukashev, E.P. Phycoerythrin Association with Photosystem II in the cryptophyte alga Rhodomonas salina. Biochemistry 2020, 85, 679–688. [Google Scholar] [CrossRef]
- Jeffrey, S.W. The occurence of chlorophylls c1 and c2 in algae. J. Phycol. 1976, 12, 349–354. [Google Scholar] [CrossRef]
- Schimek, C.; Stadnichuk, I.N.; Knaust, R.; Wehrmeyer, W. Detection of chlorophyll c1 and magnesium-2,4-divinylpheoporphyrin a5 monomethylester in cryptophytes. J. Phycol. 1994, 30, 621–627. [Google Scholar] [CrossRef]
- Larkum, A.W.D. Light-harvesting systems in algae. In Photosynthesis of Algae; Larkum, A.W.D., Douglas, S.E., Raven, J.A., Eds.; Kluwer Academic Publishers: Dordrecht, The Netherlands, 2003; pp. 277–304. [Google Scholar] [CrossRef]
- Burki, F.; Okamoto, N.; Pombert, J.F.; Keeling, P.J. The evolutionary history of haptophytes and cryptophytes: Phylogenomic evidence for separate origins. Proc. Biol. Sci. 2012, 279, 2246–2254. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Johnson, M.D. The acquisition of phototrophy: Adaptive strategies of hosting endosymbionts and organelles. Photosynth. Res. 2011, 107, 117–132. [Google Scholar] [CrossRef]
- Gagat, P.; Bodył, A.; Mackiewicz, P.; Stiller, J.W. Tertiary plastid endosymbioses in dinoflagellates. In Endosymbiosis; Löffelhardt, W., Ed.; Springer: Vienna, Austria, 2014; pp. 233–290. [Google Scholar] [CrossRef]
- Stoecker, D.K.; Hansen, P.J.; Caron, D.A.; Mitra, A. Mixotrophy in the marine plankton. Ann. Rev. Mar. Sci. 2017, 9, 311–335. [Google Scholar] [CrossRef] [Green Version]
- Hackett, J.D.; Maranda, L.; Yoon, H.S.; Bhattacharya, D. Phylogenetic evidence for the cryptophyte origin of the plastid of Dinophysis (Dinophysiales, Dinophyceae). J. Phycol. 2003, 39, 440–448. [Google Scholar] [CrossRef] [Green Version]
- Hewes, C.D.; Mitchell, B.G.; Moisan, T.A.; Vernet, M.; Reid, F.M.H. The phycobiln signatures of chloroplasts from three dinoflagellate species: A microanalytical study of Dinophysis caudata, D. fortii, and D. acuminata (Dinophysiales, Dinophyceae). J. Phycol. 1998, 34, 945–951. [Google Scholar] [CrossRef]
- Vesk, M.; Dibbayawan, T.P.; Vesk, P.A. Immunogold localization of phycoerythrin in chloroplasts of Dinophysis acuminate and D. fortii (Dinophysiales, Dinophyta). Phycologia 1996, 35, 234–238. [Google Scholar] [CrossRef]
- Schnepf, E.; Elbrächter, M. Dinophyte chloroplasts and phylogeny—A review. Grana 1999, 38, 81–97. [Google Scholar] [CrossRef]
- Croce, R.; van Amerongen, H. Natural strategies for photosynthetic light harvesting. Nat. Chem. Biol. 2014, 10, 492–501. [Google Scholar] [CrossRef]
- Kirilovsky, D.; Büchel, C. Evolution and function of light-harvesting antenna in oxygenic photosynthesis. In Advances in Botanical Research; Academic Press: Cambridge, MA, USA, 2019. [Google Scholar] [CrossRef]
- Glazer, A.N. Light guides. Directional energy transfer in a photosynthetic antenna. J. Biol. Chem. 1989, 264, 1–4. [Google Scholar] [CrossRef] [PubMed]
- Nishio, J.N. Why are higher plants green? Evolution of the higher plant photosynthetic pigment complement. Plant Cell Environ. 2000, 23, 599–648. [Google Scholar] [CrossRef]
- Saunders, G.W.; Hommersand, M.H. Assessing red algal supraordinal diversity and taxonomy in the context of contemporary systematic data. Am. J. Bot. 2004, 91, 1494–1507. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Raven, J.A. Photosynthesis in watercolours. Nature 2007, 448, 418. [Google Scholar] [CrossRef] [PubMed]
- Adir, N. Elucidation of the molecular structures of components of phycobilisome: Reconstructing a giant. Photosynth. Res. 2005, 85, 15–32. [Google Scholar] [CrossRef] [PubMed]
- Scheer, H.; Zhao, K.-H. Biliprotein maturation: The chromophore attachment. Mol. Microbiol. 2008, 68, 263–276. [Google Scholar] [CrossRef] [Green Version]
- Raven, J.A. A cost-benefit analysis of photon absorption by photosynthetic unicells. New Phytol. 1984, 98, 274–285. [Google Scholar] [CrossRef]
- Marosvölgyi, M.A.; van Gorkom, H.J. Cost and color of photosynthesis. Photosynth. Res. 2010, 103, 105–109. [Google Scholar] [CrossRef] [Green Version]
- Westermann, M.; Reuter, W.; Schimek, C.; Wehrmeyer, W. Presence of both hemidiscoidal and hemiellipsoidal phycobilisomes in a Phormidium species (cyanobacteria). Z. Naturforsch. 1993, 48, 28–34. [Google Scholar] [CrossRef]
- Niklas, K.J. The cell walls that bind the tree of life. BioScience 2004, 54, 831–841. [Google Scholar] [CrossRef] [Green Version]
- Sarkar, P.; Bosneaga, E.; Auer, M. Plant cell walls throughout evolution: Towards a molecular understanding of their design principles. J. Exp. Bot. 2009, 60, 3615–3635. [Google Scholar] [CrossRef] [Green Version]
- Yong, W.; Link, B.; O’Malley, R.; Tewari, J.; Hunter, C.T.; Lu, C.-A.; Li, X.; Bleecker, A.B.; Koch, K.E.; McCann, M.C.; et al. Genomics of plant cell wall biogenesis. Planta 2005, 221, 747–751. [Google Scholar] [CrossRef]
- Rosen, R. Optimality Principles in Biology; Butterworths: London, UK, 1967; 198p. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Phycobiliprotein | Presence in Photosynthetics |
|---|---|
| Allophycocyanin (PC, or APC) | Cyanobacteria, red, glaucophytic algae, and photosynthetic amoebae |
| C-phycocyanin (PC, or C-PC) | Cyanobacteria, glaucophytic algae, photosynthetic amoebae, and some red algae |
| Phycoerythrocyanin (PEC) | Some cyanobacteria |
| R-phycocyanin (R-PC, or R-PC I) | Red algae |
| R-PC II, R-PC III, R-PC IV and R-PC V | Cyanobacteria |
| C-phycoerythrin (C-PE) | Cyanobacteria |
| CU-phycoerythrin (CU-PE) | Cyanobateria |
| B- and b-phycoerythrins (B-PE and b-PE) | Red algae |
| R-phycoerythrin (R-PE) | Red algae |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Stadnichuk, I.N.; Kusnetsov, V.V. Phycobilisomes and Phycobiliproteins in the Pigment Apparatus of Oxygenic Photosynthetics: From Cyanobacteria to Tertiary Endosymbiosis. Int. J. Mol. Sci. 2023, 24, 2290. https://doi.org/10.3390/ijms24032290
Stadnichuk IN, Kusnetsov VV. Phycobilisomes and Phycobiliproteins in the Pigment Apparatus of Oxygenic Photosynthetics: From Cyanobacteria to Tertiary Endosymbiosis. International Journal of Molecular Sciences. 2023; 24(3):2290. https://doi.org/10.3390/ijms24032290
Chicago/Turabian StyleStadnichuk, Igor N., and Victor V. Kusnetsov. 2023. "Phycobilisomes and Phycobiliproteins in the Pigment Apparatus of Oxygenic Photosynthetics: From Cyanobacteria to Tertiary Endosymbiosis" International Journal of Molecular Sciences 24, no. 3: 2290. https://doi.org/10.3390/ijms24032290