
Plasma-Generated Nitric Oxide Water Mediates Environmentally Transmitted Pathogenic Bacterial Inactivation via Intracellular Nitrosative Stress
 , , , , ,
, , , , ,  and
and
Abstract
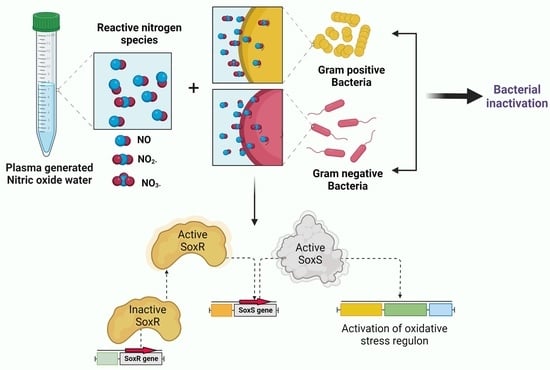
:
1. Introduction
2. Results
2.1. Physiochemical Characterization of PG-NOW
2.2. Antibacterial Activity of PG-NOW
2.3. Analysis of Intracellular RNS Generated by PG-NOW in Bacteria
2.4. Plasma-Activated NO Water Upregulates Nitrosative Stress Genes’ Response
2.5. SEM and EDS Analysis
3. Discussion
4. Materials and Methods
4.1. Microwave Plasma System and PG-NOW Preparation
4.2. Characterization of PG-NOW
4.3. Determination of Cell Inhibition with Colony-Forming Unit Assay
4.4. Live–Dead Assay
4.5. Detection of Intracellular RNS
4.6. Quantitative Real-Time PCR Analysis
4.7. SEM and EDS
4.8. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Denegre, A.A.; Mbah, M.L.N.; Myers, K.; Fefferman, N.H. Emergence of Antibiotic Resistance in Immunocompromised Host Populations: A Case Study of Emerging Antibiotic Resistant Tuberculosis in AIDS Patients. PLoS ONE 2019, 14, e0212969. [Google Scholar] [CrossRef] [PubMed]
- Larsson, D.G.J.; Flach, C.F. Antibiotic Resistance in the Environment. Nat. Rev. Microbiol. 2022, 20, 257–269. [Google Scholar] [CrossRef] [PubMed]
- Aljeldah, M.M. Antimicrobial Resistance and Its Spread Is a Global Threat. Antibiotics 2022, 11, 1082. [Google Scholar] [CrossRef] [PubMed]
- Onanuga, A.; Temedie, T.C. Multidrug-Resistant Intestinal Staphylococcus Aureus among Self-Medicated Healthy Adults in Amassoma, South-South, Nigeria. J. Health Popul. Nutr. 2011, 29, 446–453. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mohsen, S.; Dickinson, J.A.; Somayaji, R. Update on the Adverse Effects of Antimicrobial Therapies in Community Practice. Can. Fam. Physician 2020, 66, 651. [Google Scholar]
- Calderwood, S.B.; Moellering, R.C. Common Adverse Effects of Antibacterial Agents on Major Organ Systems. Surg. Clin. N. Am. 1980, 60, 65–81. [Google Scholar] [CrossRef]
- Song, C.H.; Attri, P.; Ku, S.K.; Han, I.; Bogaerts, A.; Choi, E.H. Cocktail of Reactive Species Generated by Cold Atmospheric Plasma: Oral Administration Induces Non-Small Cell Lung Cancer Cell Death. J. Phys. D. Appl. Phys. 2021, 54, 185202. [Google Scholar] [CrossRef]
- Jones-Carson, J.; Yahashiri, A.; Kim, J.S.; Liu, L.; Fitzsimmons, L.F.; Weiss, D.S.; Vázquez-Torres, A. Nitric Oxide Disrupts Bacterial Cytokinesis by Poisoning Purine Metabolism. Sci. Adv. 2020, 6, eaaz0260. [Google Scholar] [CrossRef] [Green Version]
- Ma, R.; Wang, G.; Tian, Y.; Wang, K.; Zhang, J.; Fang, J. Non-Thermal Plasma-Activated Water Inactivation of Food-Borne Pathogen on Fresh Produce. J. Hazard. Mater. 2015, 300, 643–651. [Google Scholar] [CrossRef]
- Kaushik, N.K.; Bhartiya, P.; Kaushik, N.; Shin, Y.; Nguyen, L.N.; Park, J.S.; Kim, D.; Choi, E.H. Nitric-Oxide Enriched Plasma-Activated Water Inactivates 229E Coronavirus and Alters Antiviral Response Genes in Human Lung Host Cells. Bioact. Mater. 2023, 19, 569–580. [Google Scholar] [CrossRef]
- Kang, M.H.; Jeon, S.S.; Shin, S.M.; Veerana, M.; Ji, S.H.; Uhm, H.S.; Choi, E.H.; Shin, J.H.; Park, G. Dynamics of Nitric Oxide Level in Liquids Treated with Microwave Plasma-Generated Gas and Their Effects on Spinach Development. Sci. Rep. 2019, 9, 1011. [Google Scholar] [CrossRef]
- Poh, W.H.; Rice, S.A. Recent Developments in Nitric Oxide Donors and Delivery for Antimicrobial and Anti-Biofilm Applications. Molecules 2022, 27, 674. [Google Scholar] [CrossRef]
- Julák, J.; Hujacová, A.; Scholtz, V.; Khun, J.; Holada, K. Contribution to the Chemistry of Plasma-Activated Water. Plasma Phys. Rep. 2018, 44, 125–136. [Google Scholar] [CrossRef]
- Stern, A.M.; Zhu, J. An Introduction to Nitric Oxide Sensing and Response in Bacteria. Adv. Appl. Microbiol. 2014, 87, 187–220. [Google Scholar]
- Barraud, N.; Hassett, D.J.; Hwang, S.H.; Rice, S.A.; Kjelleberg, S.; Webb, J.S. Involvement of Nitric Oxide in Biofilm Dispersal of Pseudomonas Aeruginosa. J. Bacteriol. 2006, 188, 7344–7353. [Google Scholar] [CrossRef] [Green Version]
- Figueroa, X.F.; Lillo, M.A.; Gaete, P.S.; Riquelme, M.A.; Sáez, J.C. Diffusion of Nitric Oxide across Cell Membranes of the Vascular Wall Requires Specific Connexin-Based Channels. Neuropharmacology 2013, 75, 471–478. [Google Scholar] [CrossRef]
- Liu, X.; Srinivasan, P.; Collard, E.; Grajdeanu, P.; Zweier, J.L.; Friedman, A. Nitric Oxide Diffusion Rate Is Reduced in the Aortic Wall. Biophys. J. 2008, 94, 1880. [Google Scholar] [CrossRef] [Green Version]
- Haselden, W.D.; Kedarasetti, R.T.; Drew, P.J. Spatial and Temporal Patterns of Nitric Oxide Diffusion and Degradation Drive Emergent Cerebrovascular Dynamics. PLoS Comput. Biol. 2020, 16, e1008069. [Google Scholar] [CrossRef]
- Möller, M.N.; Denicola, A. Diffusion of Nitric Oxide and Oxygen in Lipoproteins and Membranes Studied by Pyrene Fluorescence Quenching. Free Radic. Biol. Med. 2018, 128, 137–143. [Google Scholar] [CrossRef]
- Fang, F.C.; Vázquez-Torres, A. Reactive Nitrogen Species in Host–Bacterial Interactions. Curr. Opin. Immunol. 2019, 60, 96–102. [Google Scholar] [CrossRef]
- Schairer, D.O.; Chouake, J.S.; Nosanchuk, J.D.; Friedman, A.J. The Potential of Nitric Oxide Releasing Therapies as Antimicrobial Agents. Virulence 2012, 3, 271–279. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, H.R.; Lee, Y.S.; You, Y.S.; Huh, J.Y.; Kim, K.; Hong, Y.C.; Kim, C.H. Antimicrobial Effects of Microwave Plasma-Activated Water with Skin Protective Effect for Novel Disinfectants in Pandemic Era. Sci. Rep. 2022, 12, 5968. [Google Scholar] [CrossRef] [PubMed]
- Privett, B.J.; Broadnax, A.D.; Bauman, S.J.; Riccio, D.A.; Schoenfisch, M.H. Examination of Bacterial Resistance to Exogenous Nitric Oxide. Nitr. Oxide—Biol. Chem. 2012, 26, 169–173. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaper, J.B.; Nataro, J.P.; Mobley, H.L.T. Pathogenic Escherichia coli. Nat. Rev. Microbiol. 2004, 2, 123–140. [Google Scholar] [CrossRef] [PubMed]
- Qin, S.; Xiao, W.; Zhou, C.; Pu, Q.; Deng, X.; Lan, L.; Liang, H.; Song, X.; Wu, M. Pseudomonas Aeruginosa: Pathogenesis, Virulence Factors, Antibiotic Resistance, Interaction with Host, Technology Advances and Emerging Therapeutics. Signal Transduct. Target. Ther. 2022, 7, 199. [Google Scholar] [CrossRef]
- Cheung, G.Y.C.; Bae, J.S.; Otto, M. Pathogenicity and Virulence of Staphylococcus Aureus. Virulence 2021, 12, 547–569. [Google Scholar] [CrossRef]
- Man, C.; Zhang, C.; Fang, H.; Zhou, R.; Huang, B.; Xu, Y.; Zhang, X.; Shao, T. Nanosecond-Pulsed Microbubble Plasma Reactor for Plasma-Activated Water Generation and Bacterial Inactivation. Plasma Process. Polym. 2022, 19, 2200004. [Google Scholar] [CrossRef]
- Goss, S.P.A.; Singh, R.J.; Hogg, N.; Kalyanaraman, B. Reactions of ·NO, ·NO2 and Peroxynitrite in Membranes: Physiological Implications. Free Radic. Res. 1999, 31, 597–606. [Google Scholar] [CrossRef]
- Schönafinger, K. Heterocyclic NO Prodrugs. Farmaco 1999, 54, 316–320. [Google Scholar] [CrossRef]
- Abraham, M.H.; Gola, J.M.R.; Cometto-Muniz, J.E.; Cain, W.S. The Solvation Properties of Nitric Oxide. J. Chem. Soc. Perkin Trans. 2000, 10, 2067–2070. [Google Scholar] [CrossRef] [Green Version]
- Anes, J.; Dever, K.; Eshwar, A.; Nguyen, S.; Cao, Y.; Sivasankaran, S.K.; Sakalauskaitė, S.; Lehner, A.; Devineau, S.; Daugelavičius, R.; et al. Analysis of the Oxidative Stress Regulon Identifies Soxs as a Genetic Target for Resistance Reversal in Multidrug-Resistant Klebsiella Pneumoniae. mBio 2021, 12, e00867-21. [Google Scholar] [CrossRef]
- Fourie, K.R.; Wilson, H.L. Understanding Groel and Dnak Stress Response Proteins as Antigens for Bacterial Diseases. Vaccines 2020, 8, 773. [Google Scholar] [CrossRef]
- Fayet, O.; Ziegelhoffer, T.; Georgopoulos, C. The GroES and GroEL Heat Shock Gene Products of Escherichia coli Are Essential for Bacterial Growth at All Temperatures. J. Bacteriol. 1989, 171, 1379–1385. [Google Scholar] [CrossRef] [Green Version]
- Calloni, G.; Chen, T.; Schermann, S.M.; Chang, H.C.; Genevaux, P.; Agostini, F.; Tartaglia, G.G.; Hayer-Hartl, M.; Hartl, F.U. DnaK Functions as a Central Hub in the E. coli Chaperone Network. Cell Rep. 2012, 1, 251–264. [Google Scholar] [CrossRef] [Green Version]
- Staerck, C.; Gastebois, A.; Vandeputte, P.; Calenda, A.; Larcher, G.; Gillmann, L.; Papon, N.; Bouchara, J.P.; Fleury, M.J.J. Microbial Antioxidant Defense Enzymes. Microb. Pathog. 2017, 110, 56–65. [Google Scholar] [CrossRef]
- Fasnacht, M.; Polacek, N. Oxidative Stress in Bacteria and the Central Dogma of Molecular Biology. Front. Mol. Biosci. 2021, 8, 671037. [Google Scholar] [CrossRef]
- Imlay, J.A. Transcription Factors That Defend Bacteria against Reactive Oxygen Species. Annu. Rev. Microbiol. 2015, 69, 93–108. [Google Scholar] [CrossRef] [Green Version]
- Christman, M.F.; Storz, G.; Ames, B.N. OxyR, a Positive Regulator of Hydrogen Peroxide-Inducible Genes in Escherichia coli and Salmonella Typhimurium, Is Homologous to a Family of Bacterial Regulatory Proteins. Proc. Natl. Acad. Sci. USA 1989, 86, 3484–3488. [Google Scholar] [CrossRef] [Green Version]
- Devane, M.L.; Moriarty, E.; Weaver, L.; Cookson, A.; Gilpin, B. Fecal Indicator Bacteria from Environmental Sources; Strategies for Identification to Improve Water Quality Monitoring. Water Res. 2020, 185, 116204. [Google Scholar] [CrossRef]
- Ringen, L.M.; Drake, C.H. A Study of the Incidence of Pseudomonas Aeruginosa from Various Natural Sources. J. Bacteriol. 1952, 64, 841–845. [Google Scholar] [CrossRef] [Green Version]
- O’Gara, J.P. Into the Storm: Chasing the Opportunistic Pathogen Staphylococcus aureus from Skin Colonisation to Life-Threatening Infections. Environ. Microbiol. 2017, 19, 3823–3833. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choi, E.H.; Kaushik, N.K.; Hong, Y.J.; Lim, J.S.; Choi, J.S.; Han, I. Plasma Bioscience for Medicine, Agriculture and Hygiene Applications. J. Korean Phys. Soc. 2022, 80, 817–851. [Google Scholar] [CrossRef] [PubMed]
- Choi, E.H.; Uhm, H.S.; Kaushik, N.K. Plasma Bioscience and Its Application to Medicine. AAPPS Bull. 2021, 31, 10. [Google Scholar] [CrossRef]
- Kaushik, N.K.; Bekeschus, S.; Tanaka, H.; Lin, A.; Choi, E.H. Plasma Medicine Technologies. Appl. Sci. 2021, 11, 4584. [Google Scholar] [CrossRef]
- Patinglag, L.; Melling, L.M.; Whitehead, K.A.; Sawtell, D.; Iles, A.; Shaw, K.J. Non-Thermal Plasma-Based Inactivation of Bacteria in Water Using a Microfluidic Reactor. Water Res. 2021, 201, 117321. [Google Scholar] [CrossRef]
- Tomić, S.; Petrović, A.; Puač, N.; Škoro, N.; Bekić, M.; Petrović, Z.L.; Čolić, M. Plasma-Activated Medium Potentiates the Immunogenicity of Tumor Cell Lysates for Dendritic Cell-Based Cancer Vaccines. Cancers 2021, 13, 1626. [Google Scholar] [CrossRef]
- Deupree, S.M.; Schoenfisch, M.H. Morphological Analysis of the Antimicrobial Action of Nitric Oxide on Gram-Negative Pathogens Using Atomic Force Microscopy. Acta Biomater. 2009, 5, 1405–1415. [Google Scholar] [CrossRef] [Green Version]
- Burney, S.; Caulfield, J.L.; Niles, J.C.; Wishnok, J.S.; Tannenbaum, S.R. The Chemistry of DNA Damage from Nitric Oxide and Peroxynitrite. Mutat. Res. Mol. Mech. Mutagen. 1999, 424, 37–49. [Google Scholar] [CrossRef]
- Fang, F.C. Perspectives Series: Host/Pathogen Interactions. J. Clin. Investig. 1997, 99, 2818–2825. [Google Scholar] [CrossRef] [Green Version]
- Hetrick, E.M.; Shin, J.H.; Stasko, N.A.; Johnson, C.B.; Wespe, D.A.; Holmuhamedov, E.; Schoenfisch, M.H. Bactericidal Efficacy of Nitric Oxide-Releasing Silica Nanoparticles. ACS Nano 2008, 2, 235–246. [Google Scholar] [CrossRef]
- Ischiropoulos, H.; Zhu, L.; Chen, J.; Tsai, M.; Martin, J.C.; Smith, C.D.; Beckman, J.S. Peroxynitrite-Mediated Tyrosine Nitration Catalyzed by Superoxide Dismutase. Arch. Biochem. Biophys. 1992, 298, 431–437. [Google Scholar] [CrossRef]
- Pryor, W.A.; Jin, X.; Squadrito, G.L. One- and Two-Electron Oxidations of Methionine by Peroxynitrite. Proc. Natl. Acad. Sci. USA 1994, 91, 11173–11177. [Google Scholar] [CrossRef]
- Radi, R.; Beckman, J.S.; Bush, K.M.; Freeman, B.A. Peroxynitrite-Induced Membrane Lipid Peroxidation: The Cytotoxic Potential of Superoxide and Nitric Oxide. Arch. Biochem. Biophys. 1991, 288, 481–487. [Google Scholar] [CrossRef]
- King, P.A.; Jamison, E.; Strahs, D.; Anderson, V.E.; Brenowitz, M. ‘Footprinting’ Proteins on DNA with Peroxonitrous Acid. Nucleic Acids Res. 1993, 21, 2473–2478. [Google Scholar] [CrossRef] [Green Version]
- Castro, L.; Rodriguez, M.; Radi, R. Aconitase Is Readily Inactivated by Peroxynitrite, but Not by Its Precursor, Nitric Oxide. J. Biol. Chem. 1994, 269, 29409–29415. [Google Scholar] [CrossRef]
- Floris, R.; Piersma, S.R.; Yang, G.; Jones, P.; Wever, R. Interaction of Myeloperoxidase with Peroxynitrite. Eur. J. Biochem. 1993, 215, 767–775. [Google Scholar] [CrossRef]
- Denicola, A.; Souza, J.M.; Radi, R.; Lissi, E. Nitric Oxide Diffusion in Membranes Determined by Fluorescence Quenching. Arch. Biochem. Biophys. 1996, 328, 208–212. [Google Scholar] [CrossRef]
- Marchettini, N.; Rustici, M.; Branca, M.; Culeddu, N.; Fruianu, M.; Serra, M.V.; Tiezzi, E. Solubility of Nitric Oxide (NO) in Lipid Aggregates as Monitored by Nuclear Magnetic Resonance. Colloids Surf. A Physicochem. Eng. Asp. 1998, 1, 1–6. [Google Scholar] [CrossRef]
- Rubbo, H.; Radi, R.; Anselmi, D.; Kirk, M.; Barnes, S.; Butler, J.; Eiserich, J.P.; Freeman, B.A. Nitric Oxide Reaction with Lipid Peroxyl Radicals Spares α-Tocopherol during Lipid Peroxidation. J. Biol. Chem. 2000, 275, 10812–10818. [Google Scholar] [CrossRef] [Green Version]
- Rouillard, K.R.; Novak, O.P.; Pistiolis, A.M.; Yang, L.; Ahonen, M.J.R.; McDonald, R.A.; Schoenfisch, M.H. Exogenous Nitric Oxide Improves Antibiotic Susceptibility in Resistant Bacteria. ACS Infect. Dis. 2021, 7, 23–33. [Google Scholar] [CrossRef]
- Mai-Prochnow, A.; Zhou, R.; Zhang, T.; Ostrikov, K.; Mugunthan, S.; Rice, S.A.; Cullen, P.J. Interactions of Plasma-Activated Water with Biofilms: Inactivation, Dispersal Effects and Mechanisms of Action. npj Biofilms Microbiom. 2021, 7, 11. [Google Scholar] [CrossRef] [PubMed]
- Han, Q.Y.; He, Z.Y.; Zhong, C.S.; Wen, X.; Ni, Y.Y. The Optimization of Plasma Activated Water (PAW) Generation and the Inactivation Mechanism of PAW on Escherichia coli. J. Food Process. Preserv. 2022, 46, e17120. [Google Scholar] [CrossRef]
- Lu, Y.; Slomberg, D.L.; Schoenfisch, M.H. Nitric Oxide-Releasing Chitosan Oligosaccharides as Antibacterial Agents. Biomaterials 2014, 35, 1716–1724. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Z.; Demple, B. SoxS, an Activator of Superoxide Stress Genes in Escherichia coli. Purification and Interaction with DNA. J. Biol. Chem. 1994, 269, 18371–18377. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.; Weiss, B. Two Divergently Transcribed Genes, SoxR and SoxS, Control a Superoxide Response Regulon of Escherichia coli. J. Bacteriol. 1991, 173, 2864–2871. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Demple, B. Redox Signaling and Gene Control in the Escherichia coli SoxRS Oxidative Stress Regulon—A Review. Gene 1996, 179, 53–57. [Google Scholar] [CrossRef]
- Seo, S.W.; Kim, D.; Szubin, R.; Palsson, B.O. Genome-Wide Reconstruction of OxyR and SoxRS Transcriptional Regulatory Networks under Oxidative Stress in Escherichia coli K-12 MG1655. Cell Rep. 2015, 12, 1289–1299. [Google Scholar] [CrossRef] [Green Version]
- Imlay, J.A. Cellular Defenses against Superoxide and Hydrogen Peroxide. Annu. Rev. Biochem. 2008, 77, 755–776. [Google Scholar] [CrossRef] [Green Version]
- Mayer, M.P. The Hsp70-Chaperone Machines in Bacteria. Front. Mol. Biosci. 2021, 8, 1–22. [Google Scholar] [CrossRef]
- Nakamura, T.; Tanaka, M.; Maruya, A.; Higashi, Y.; Kurusu, Y. A Nonconserved Carboxy-Terminal Segment of GroEL Contributes to Reaction Temperature. Biosci. Biotechnol. Biochem. 2004, 68, 2498–2504. [Google Scholar] [CrossRef] [Green Version]
- Lund, P.A. Microbial Molecular Chaperones. Adv. Microb. Physiol. 2001, 44, 93–140. [Google Scholar] [CrossRef]
- Ryabova, N.A.; Marchenkov, V.V.; Marchenkova, S.Y.; Kotova, N.V.; Semisotnov, G.V. Molecular Chaperone GroEL/ES: Unfolding and Refolding Processes. Biochemistry 2013, 78, 1405–1414. [Google Scholar] [CrossRef]
- Zhang, H.; Gong, W.; Wu, S.; Perrett, S. Hsp70 in Redox Homeostasis. Cells 2022, 11, 829. [Google Scholar] [CrossRef]
- Khor, H.K.; Fisher, M.T.; Schöneich, C. Potential Role of Methionine Sulfoxide in the Inactivation of the Chaperone GroEL by Hypochlorous Acid (HOCl) and Peroxynitrite (ONOO-). J. Biol. Chem. 2004, 279, 19486–19493. [Google Scholar] [CrossRef] [Green Version]
- Xiang, Q.; Kang, C.; Niu, L.; Zhao, D.; Li, K.; Bai, Y. Antibacterial Activity and a Membrane Damage Mechanism of Plasma-Activated Water against Pseudomonas Deceptionensis CM2. LWT 2018, 96, 395–401. [Google Scholar] [CrossRef]
- Shen, J.; Tian, Y.; Li, Y.; Ma, R.; Zhang, Q.; Zhang, J.; Fang, J. Bactericidal Effects against S. Aureus and Physicochemical Properties of Plasma Activated Water Stored at Different Temperatures. Sci. Rep. 2016, 6, 28505. [Google Scholar] [CrossRef] [Green Version]
- Malik, M.A. Nitric Oxide Production by High Voltage Electrical Discharges for Medical Uses: A Review. Plasma Chem. Plasma Process. 2016, 36, 737–766. [Google Scholar] [CrossRef]
- Tenover, F.C. Antimicrobial Susceptibility Testing. In Encyclopedia of Microbiology; Academic Press: Cambridge, MA, USA, 2019; pp. 166–175. ISBN 9780128117378. [Google Scholar]
- Chautrand, T.; Souak, D.; Chevalier, S.; Duclairoir-Poc, C. Gram-Negative Bacterial Envelope Homeostasis under Oxidative and Nitrosative Stress. Microorganisms 2022, 10, 924. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain | Gram Nature | Pathogenic Properties | Origin |
|---|---|---|---|
| E. coli (K-12) | Negative | Cause illness, diarrhoea, dysentery, meningitis, urinary tract infection, etc. | Waterborne; foodborne |
| P. aeruginosa | Negative | Cause nosocomial infections such as pneumonia, urinary tract infections, surgical wound infection, bacteraemia, etc. | Waterborne; soilborne |
| S. aureus | Positive | Develops multiple antibiotic resistance, bacteraemia, infective endocarditis, osteoarticular, skin and soft tissue, pleuropulmonary, etc. | Waterborne; Airborne; foodborne |
| Gene Name | Sequence (5′-3′) | Size (bp) |
|---|---|---|
| gapA-forward | CACGCTACTACCGCTACTCA | 205 |
| gapA -reverse | CGGTCAGGTCAACTACGGAT | |
| sorR-forward | GTATCGGCGCTGCATTTCTA | 208 |
| soxR-reverse | GCTGTTTCCACTCTTTCGCA | |
| soxS-forward | GCATATTGACCAGCCGCTTA | 245 |
| soxS-reverse | GATCAAACTGCCGACGGAAA | |
| oxyR-forward | AGTTGGACCGTACCTGCTAC | 161 |
| oxyR-reverse | CTTCGCTCTCTTTCACCAGC | |
| groL-forward | TCCGTACCATGCTCTGACTC | 178 |
| groL-reverse | GCATACCTTCAACCACGTCC | |
| dnaK-forward | ATCGAACTGTCTTCCGCTCA | 156 |
| dnaK-reverse | AACTTTCAGCGGCTCAATGG |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Borkar, S.B.; Negi, M.; Kaushik, N.; Abdul Munnaf, S.; Nguyen, L.N.; Choi, E.H.; Kaushik, N.K. Plasma-Generated Nitric Oxide Water Mediates Environmentally Transmitted Pathogenic Bacterial Inactivation via Intracellular Nitrosative Stress. Int. J. Mol. Sci. 2023, 24, 1901. https://doi.org/10.3390/ijms24031901
Borkar SB, Negi M, Kaushik N, Abdul Munnaf S, Nguyen LN, Choi EH, Kaushik NK. Plasma-Generated Nitric Oxide Water Mediates Environmentally Transmitted Pathogenic Bacterial Inactivation via Intracellular Nitrosative Stress. International Journal of Molecular Sciences. 2023; 24(3):1901. https://doi.org/10.3390/ijms24031901
Chicago/Turabian StyleBorkar, Shweta B., Manorma Negi, Neha Kaushik, Shaik Abdul Munnaf, Linh Nhat Nguyen, Eun Ha Choi, and Nagendra Kumar Kaushik. 2023. "Plasma-Generated Nitric Oxide Water Mediates Environmentally Transmitted Pathogenic Bacterial Inactivation via Intracellular Nitrosative Stress" International Journal of Molecular Sciences 24, no. 3: 1901. https://doi.org/10.3390/ijms24031901