1. Introduction
The cell wall is widely present in non-animal species and plays an important role in the maintenance of life. In eukaryotic algae and plants, there are significant similarities in the structure and composition of cell walls. Deeply understanding the synthesis mechanism of cell walls is of great significance for elucidating the basic laws of cell activity and the evolution process from algae to plants. Extensin was initially discovered in plant cell walls, and more and more studies have shown that hydroxyproline-rich glycoproteins (HRGPs) play similar roles in eukaryotic algal cells, but the function and synthesis mechanism of HRGPs in the synthesis of algal cell walls are still unclear. Therefore, the model microalgae Chlamydomonas reinhardtii with a clear genetic background was used as the research object to study the mechanisms related to HRGP synthesis, laying the foundation for elucidating the role of extensin in cell wall synthesis and the cell evolution of algae towards plants.
C. reinhardtii is a unicellular eukaryotic green alga belonging to Chlorophyta, Volvocales. It is an important link in the evolution of multicellular algae and higher plants.
C. reinhardtii also has the characteristics of protists, so early scholars mainly used it as the study object of cell structure [
1]. After that, various mutant types of
C. reinhardtii were obtained by physical and chemical methods, among which the discovery of cell wall deficient strains laid the foundation for the study of cell wall structure and provided favorable conditions for the successful establishment of an efficient genetic transformation system in
C. reinhardtii. This history makes
C. reinhardtii one of the important model organisms in biological research [
2], and it is widely used in microalgae biotechnology [
3].
In the early days, the cell wall composition of
Chlamydomonas is considered to be cellulose [
4]; however, a growing number of studies have shown that the cell wall of
C. reinhardtii is a kind of glycoprotein lattice structure rich in hydroxyproline, similar to the plant extensin of higher plants. The analysis of amino acid composition showed that the cell wall protein was rich in hydroxyproline residues, and the chromatographic analysis showed that the glycogroup components in the cell wall were mainly arabinose, galactose, and a small amount of mannose [
5,
6,
7,
8,
9,
10]. In addition, when the defective cell wall strains of
C. reinhardtii were taken as the research object, it was found that the glycoprotein layer of the cell wall may have a self-assembly process [
11,
12,
13,
14]. Recently, Cronmiller et al. (2019) have carried out functional studies on some cell wall genes, including some cell wall protein processing genes and structural protein genes. Concurrently, it was found that these cell wall genes were partially activated after lysin treatment and participated in the cell wall integrity signal regulation in
C. reinhardtii [
15]. In addition, it has been shown that the arabinosyltransferase encoded by
RRA was involved in protein O-glycosylation during root hair development of model organism
Arabidopsis thaliana, and played an important role in stabilizing the helical conformation of cell wall glycoprotein [
16]. Importantly, homologous genes belonging to the glycosyltransferase family (GT77) were also found in
C. reinhardtii, which confirmed that the cell wall glycoproteins of green algae shared a common ancestor with the extensin of higher plants in the origin and evolution of green algal cell walls [
17].
RNA sequencing is a technology that comprehensively studies gene function and reveals molecular mechanisms in specific biological processes [
18]. The transcriptome is a collection of RNAs transcribed by a specific tissue or cell at a certain development stage or functional state. By studying the transcriptome, the gene expression level can be understood at the overall level, and the molecular mechanism of gene expression regulation can be revealed [
19]. At present, the molecular studies on the functional components in
C. reinhardtii cell wall are in the development stage. The functions of some cell wall genes have been studied, and the self-assembly process in
C. reinhardtii cell wall has been confirmed. However, the key genes and their roles in the cell wall synthesis process are still unclear, and more studies are needed at the molecular level to truly understand the assembly mechanism of
C. reinhardtii cell wall. In the Chlamydomonas Resource Center (University of Minnesota), there were many mutant cultures of
C. reinhardtii. Among them, CC-849 is a cell wall deficient strain with no other significant differences compared to the wild type, which lays a good foundation for researching the differences between cell wall synthesis and assembly. Therefore, this work aims to explore the key genes involved in cell wall assembly in the cell wall defective strain of
C. reinhardtii by transcriptome sequencing technology and then research the function of arabinosyltransferase on the formation of cell wall glycoprotein through gene knockout, which lays a foundation for further reveal the role of extensin in cell wall synthesis and the evolution of
C. reinhardtii cell wall glycoprotein and higher plants extensin.
3. Discussion
C. reinhardtii has the advantages of simple culture conditions, a short growth cycle, and a clear genetic background, which makes it a good material for many transgenic technology studies. The successful screening of cell-deficient strains created more favorable conditions for the genetic transformation of
C. reinhardtii and laid a solid foundation for the study of cell structure at the same time. The cell wall of
C. reinhardtii is dominated by glycoproteins rich in hydroxyproline. During its formation, the fiber network of the inner layer of W1 and the outer layer of W7 is first synthesized. After the initial complex is constructed, other cell wall layers are gradually assembled in it [
11]. The hydroxyprolin-rich glycoprotein (HRGPs) in the inner layer is initially synthesized in the endoplasmic reticulum and Golgi apparatus, transported with vesicles to the shrinking vacuole, and released to the cell surface for self-assembly [
12]. Finally, the glycoproteins interact and cross-link to the inner and outer fiber layers. The highly organized HRGPs in algal cell walls are ancient and primitive and have evolved into a superfamily containing multiple groups in plant cell wall extensin [
24]. In the study concerning the origin and evolution of cell walls in green algae, it has been stated that the extensin of higher plants is a cell wall protein that shares a common ancestor with the HRGPs of the cell wall of green algae [
17]. From the hydroxyproline-rich glycoproteins found in the algal cell wall to the development of higher plant extensin, it is the secular evolution of plant cell wall structure and function.
In this study, based on the comparative analysis of the cell wall component synthesis between the cell wall defective strain and normal cell wall strain, the differentially expressed genes were mainly related to the extensin synthesis pathway and synthesis and degradation pathways of glycoskeleton in
C. reinhardtii. In CC-849, there were up-regulated
XEG113 and
RRA (arabinosyltransferase gene) and downregulated
SGT1 (peptidyserine α-galactosyltransferase gene) in the process of plant extensin synthesis. Additionally, some cell wall structural protein genes were up-regulated. So, the expression of various regulatory genes was speculated to cooperate to maintain the level of cell wall component metabolism. However, these genes are generally up-regulated, indicating active cell wall metabolism, which seems to be inconsistent with the defect of the cell wall in CC-849. It is speculated that the defect of the CC-849 cell wall may not be entirely caused by the expression of these genes, which are up-regulated to synthesize missing cell walls to compensate for the CC-849 cell wall defect. A previous study demonstrated that gamete-specific (
GAS) Hyp-rich pherophorin-encoding genes (
GAS28,
GAS30,
GAS31), cell wall pherophorin gene (
PHC19) and protein processing genes (
SEC61G,
AraGT1,
RHM1) related to translation or glycosylation indicated a substantial increase in gene expression upon g-lysin treatment, which in the same way suggested that cell wall removal up-regulated cell wall genes expression [
15].
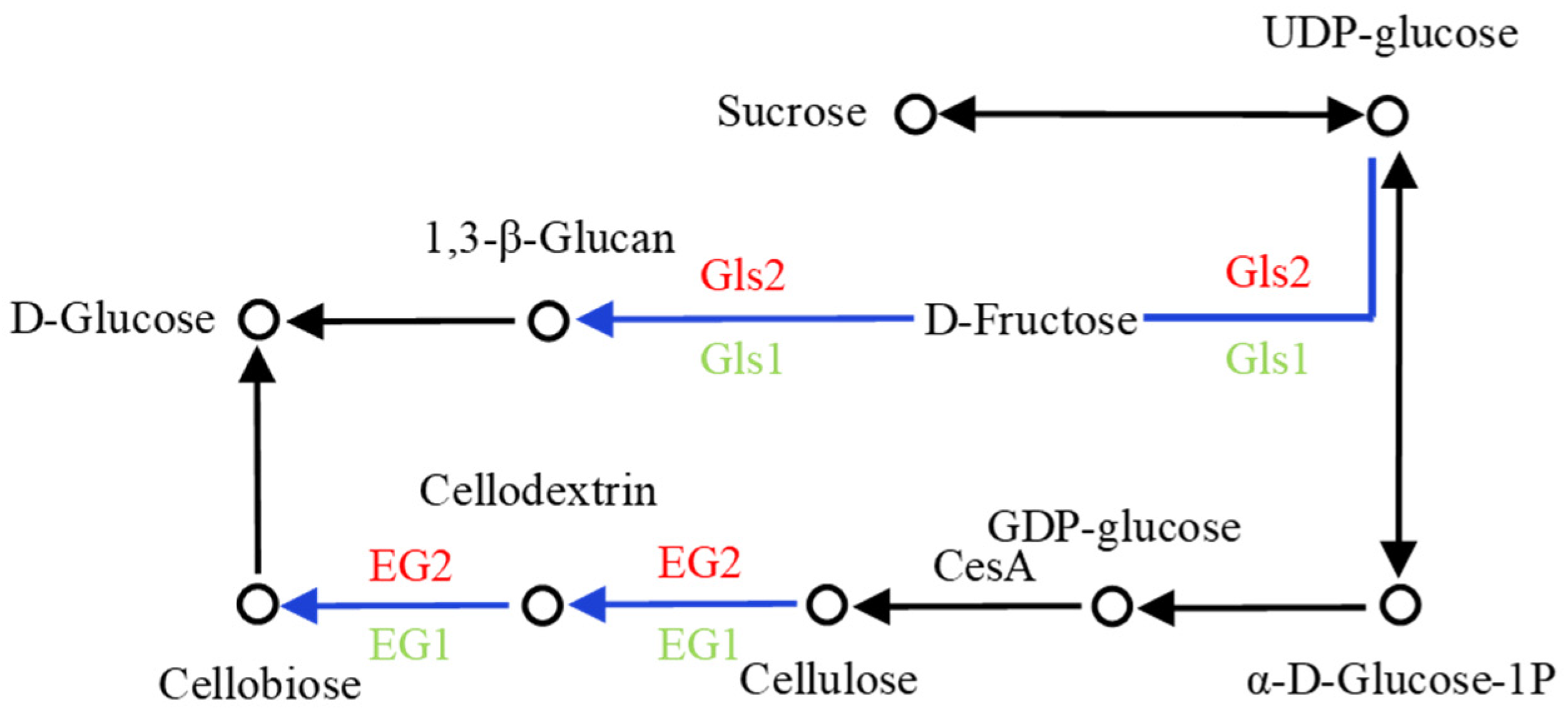
Hydroxyproline-rich glycoproteins in higher plant cells, including extensin (EXTs), are secreted into the cell walls after synthesis and shaped by posttranslational modifications. There are O-glycosylation chains with up to 4 or 5 linear arabinosyl units on each Hyp and monogalactosylation of Ser residues in Ser (Hyp)
4 repeats of EXTs (
Figure 11). Typically, during this process, there are three groups of arabinosyltransferases genes (
AraTs), hydroxyproline O-arabinosyltransferases genes (
HPAT1-HAPT3; GT8 family), reduced residual arabinosyltransferase genes (
RRA1-RRA3) and xyloglucanase gene (
XEG113; GT77 family), and a peptidyl-Ser galactosyltransferase gene (
SERGT1; GT96 family) adding a single galactopyranose to each Ser residue in Ser (Hyp)
4 motifs. Ultimately, the glycosylated EXTs may be cross-linked with the peroxidases at the Tyr residue to construct a three-dimensional network that may interact with other components of the cell walls. Some glycosylation modifying enzymes have been shown to play a role in root hair development in the model organism
Arabidopsis thaliana and influence self-assembly of the cell wall EXTs [
25]. Studies have found that
Chlamydomonas also had multiple proline hydroxylase genes (
P4H) that were essential for the correct assembly of the cell wall. By knocking out the
P4H gene in
Chlamydomonas, the cell wall layers were also unclear, and it was speculated that the disappearance of W2 and W4 layers resulted in the merger of W1 and W7 layers. It was speculated that
P4H might affect the hydroxylation of proline, reduce the content of Hyp, and further affect its glycosylation, thus affecting the cross-linking and stability of the fiber layer [
26]. However, the function of some O-glycosylation modifying enzymes is rarely reported in the research of algae.
In the present study, as
RRA gene coding glycosyltransferase is one of the most crucial genes related to the synthesis of plant extensin found by transcriptome sequencing, arabinosyltransferase inhibitor was found to prevent the algal cell growth and increase cell permeability. At the same time, after arabinosyltransferase gene (
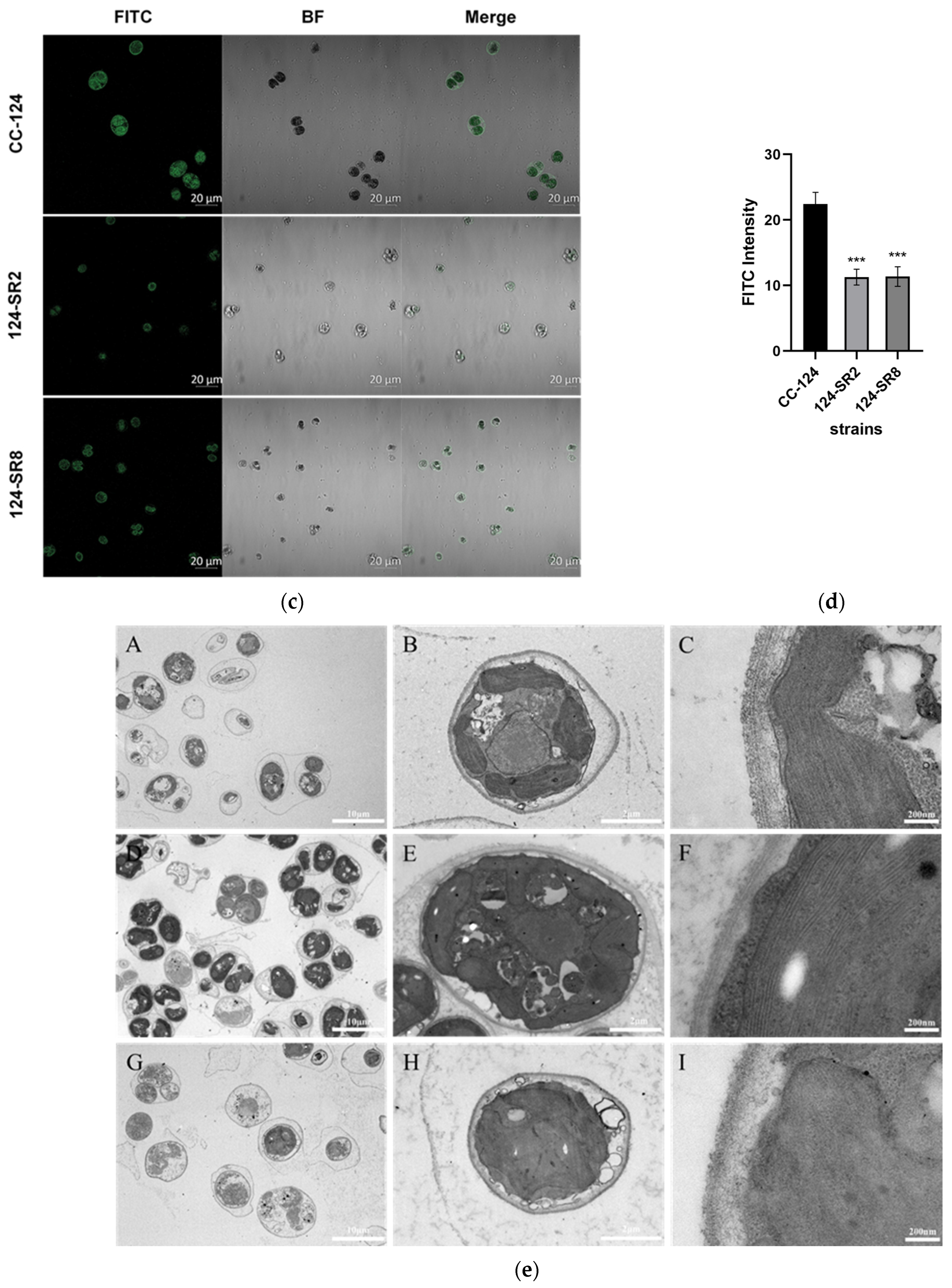
RRA) was knocked out, the cell wall showed blurred boundaries and reduced structural levels. The structure of W2 and W6 layers in the cell walls of mutant strains did not seem to disappear, but the boundary is not clear. Additionally, the reason for the looseness of the cell wall structure may be that the W7 layer is not cross-linked, and neither is the W1 layer. Similar to the CC-849 cell wall defective type, there was a vague structure layer on the outside of the plasma membrane, which may resemble the remnants of the W1 and W7 layers. Therefore, arabinosyltransferase may play an important role in the glycosylation modification and conformational stability of cell wall glycoprotein, which further affects the structural assembly of cell wall glycoprotein in
C. reinhardtii. Similarly, during the root hair development of
A. thaliana, the knockout of arabinosyltransferase genes (
RRA1 and
RRA2) revealed reduced arabinose content in the cell wall, demonstrating that these genes involved in the process of protein O-glycosylation and played an important role in stabilizing the helical conformation of cell wall extensin [
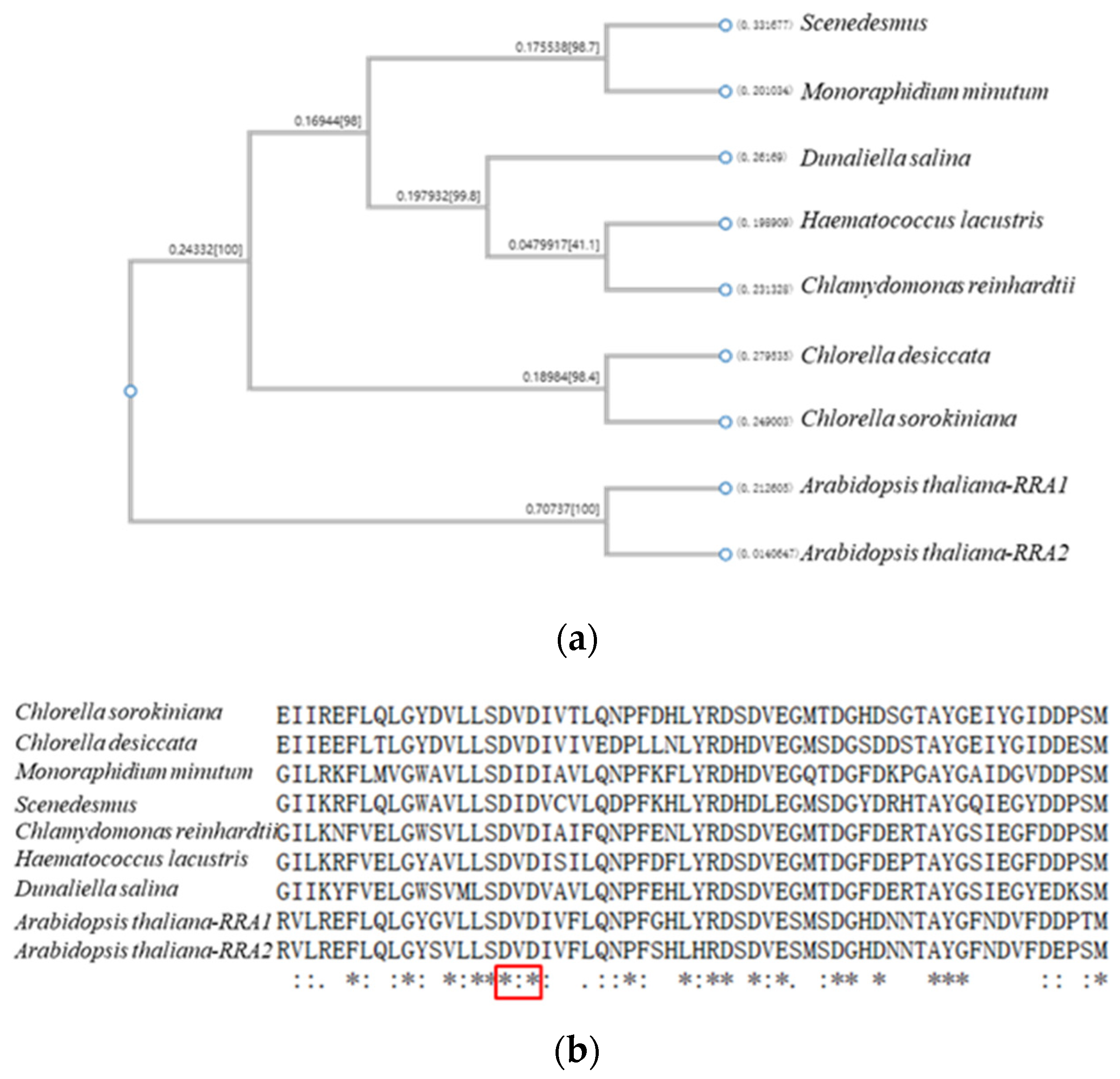
27]. At the same time, the glycosyltransferase family was investigated, and homologous genes were found in
C. reinhardtii with the same conserved domain, which confirmed the evolution trend of HRGPs from green algae cell wall to plant cell wall extensin.
4. Materials and Methods
4.1. Cultivation and Treatment of Algae
C. reinhardtii was obtained from the Chlamydomonas Resource Center (University of Minnesota laboratory), and the algal species number of the cell wall deficient strain is CC-849, and the wild-type strain with normal cell wall is CC-124. The cells of the two algal strains were cultured in a Tris-acetoacetate-phosphate (TAP) medium (pH 6.8). The illumination was 50 μmol·m−2 s−1, the temperature was 23 ± 1 °C, the light period was 12L:12D, and the rotating speed was 100 rpm. The two algal strains were set with the same initial cell density and cultured under the same conditions for three days, and the algal strains in the stable stage were sampled. After freezing with liquid nitrogen, the samples were stored at a low temperature of −80 °C for RNA extraction. The experimental groups were as follows: cell wall deficient strain CC-849 was the experimental group, denoted as CW-D, normal cell wall strain CC-124 was the control, denoted as CW, and each group consisted of triplicate.
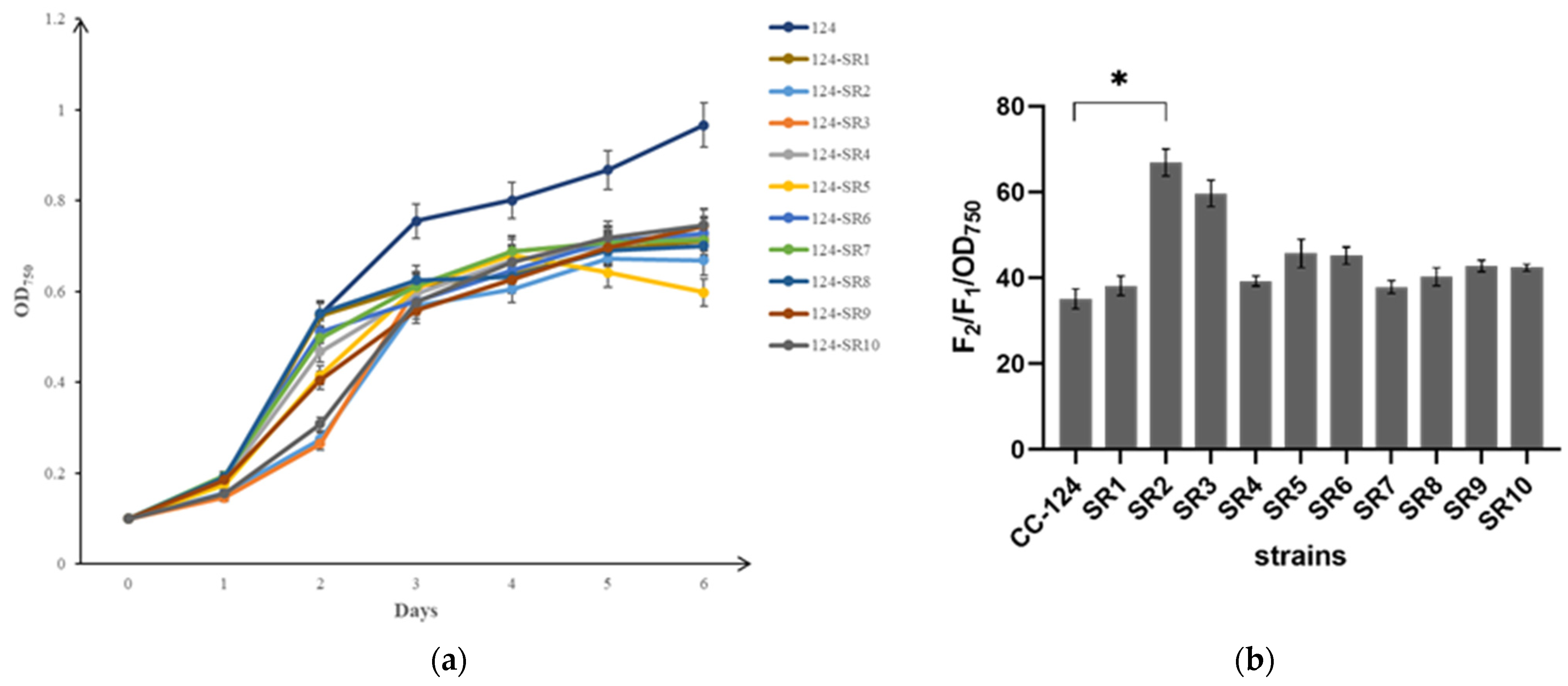
4.2. Permeability and Integrity Analysis of Cell Wall
The biomass of
C. reinhardtii was characterized by the number of cells per milliliter of algal culture and calculated from the absorbance OD
750. According to the experimental method of Yali Wang (2016) [
28], the lysing agent NP-40 (Solarbio, Beijing, China) with the main composition of Tris (pH7.4), NaCl, and 1% NP-40 was used to compare cell wall permeability of the two algal cells. The fluorescence emission value F
1 was measured at 680 nm under an excitation light of 435 nm by taking a one-milliliter suspension from the algal culture of CC-849 and CC-124, which were in stable stages. After treatment with NP-40 for 1 h and centrifugation at 3000×
g for 5 min, the fluorescence value F
2 was measured by taking a one-milliliter supernatant at the same set wavelength. The ratio of F
2/F
1 was positively correlated with the cell wall permeability.
In addition to the measurement of physiological parameters, the integrity of the
C. reinhardtii cell wall was analyzed from the following two aspects. Firstly, fluorescein-labeled ricinus communis agglutinin I (FITC-RCA I, Qiyue, Xian, China) was used to show the location of the glycogroup on the surface of algal cells, which can bind each other specifically and directly. In reference to the method of Juan Lin (2020) [
29], the algal culture at the same period and concentration was incubated with FITC-RCA I in the dark for 1 h, rinsed three times with PBS after centrifugation, and then green fluorescence was observed under a confocal laser scanning microscope (CLSM). Apart from observing the glycogroup structure of the algal cell walls, the overall structure level of the algal cell walls was further observed using transmission electron microscopy (TEM).
4.3. Extraction of Total RNA and Construction of cDNA Library
RNA was extracted from CC-849 and CC-124 by using an RNA extraction kit (Biomarker, Beijing, China). RNA concentration and purity were measured using Nanodrop 2000 and RNA integrity was assessed using the RNA Nano 6000 Assay Kit of the Agilent Bioanalyzer 2100 system to ensure transcriptome sequencing with qualified RNA samples. After the samples were qualified, transcriptome sequencing libraries were generated using the NEBNext UltraTM RNA Library Prep Kit for Illumina (NEB, Ipswich, MA, USA) following the manufacturer’s recommendations, and index codes were added to attribute sequences to each sample.
4.4. Sequencing, Quality Control and Comparative Analysis
After the library was constructed, the effective concentration of the library was accurately quantified by qRT-PCR (the effective concentration of the library was higher than 2 nM) to ensure the quality of the library. After the database check was qualified, the clustering of the index-coded samples was performed on a cBot Cluster Generation System using TruSeq PE Cluster Kit v4-cBot-HS (Illumina, San Diego, CA, USA). After cluster generation, the library preparations were sequenced on an Illumina platform, and paired-end reads were generated.
The raw reads were further processed with a bioinformatic pipeline tool, BMKCloud (
www.biocloud.net accessed on 15 December 2023) online platform. Raw data (raw reads) of FastQ format were first processed through in-house Perl scripts. In this step, clean data (clean reads) were obtained by removing reads containing adapter, reads containing ploy-N, and low-quality reads from raw data. At the same time, Q20, Q30, GC-content, and sequence duplication levels of the clean data were calculated. All the downstream analyses were based on clean data with high quality. These clean reads were then mapped to the reference genome sequences using the Hisat2 tools. Based on the reference genome of
C. reinhardtii (
https://www.ncbi.nlm.nih.gov/genome/147 accessed on 15 December 2023), only reads with a perfect match or one mismatch were further analyzed and annotated.
4.5. Differentially Expressed Gene Analysis
The FPKM value (number of fragments per kilobase length per million fragments from a gene) was used to determine the expression pattern of differentially expressed genes under different experimental conditions [
30]. For samples with biological duplication, DESeq2 was suitable for differential expression analysis between sample groups to obtain differential expression gene sets between two biological conditions. In the process of differential expression gene detection, Fold Change ≥ 2 and error discovery rate (FDR) < 0.01 were used as screening criteria, both of which were obtained by correcting the
p-value of difference significance. To investigate the differential expression genes of CC-849 and CC-124 in cell wall synthesis, a comparative combination of CW-D1&CW-D2&CW-D3 vs. CW1&CW2&CW3 was set.
The above-obtained genes were functionally annotated in seven databases (Nr, Nt, Pfam, KOG/COG, Swiss-prot, KEGG, and GO). According to the functional annotation results, the classification methods of different databases can be classified. Through GO annotation, gene functions were classified into three categories (biological processes, cell components, and molecular functions). After KO annotation, gene annotations were classified according to the KEGG metabolic pathway in which they participate.
4.6. Quantitative PCR Validation of Gene Expression
Based on the transcriptome results, differentially expressed genes related to the synthesis of cell wall components in glucose metabolism, lipid metabolism, and amino acid metabolism were selected for quantitative validation of gene expression using qRT-PCR. β-tubulin gene was selected as the internal reference gene.
RNA extraction was conducted using the E.Z.N.A.
® Plant RNA Kit (OMEGA, Germany), and the integrity of the 28S and 18S bands were checked by agarose gel electrophoresis to verify the RNA quality. The total RNA was reverse transcribed into cDNA, and the nucleic acid concentration was determined by Nanodrop. qPCR was performed using the TB Green Premix Ex Taq II Kit (Tli RNaseH Plus, Takara, Dalian, China), and each sample was set up in triplicate. The experimental data were analyzed and plotted according to the RQ = 2
−ΔΔCt. qPCR genes and primers are shown in
Table 5.
4.7. Treatment with Arabinosyltransferase Inhibitor
According to the differential expression genes obtained through transcriptome sequencing (4.5) and their validation experiments (4.6), the difference in arabinosyltransferase gene (RRA) between the two algal strains was significant. To research its role in the synthesis of cell walls in C. reinhardtii, the inhibitor-ethambutol dihydrochloride (Sparkjade, Shandong, China) is used to inhibit the activity of arabinosyltransferase.
According to pre-experimental results, the inhibitor began to affect the growth of normal cell wall strain CC-124 at a concentration of 20 mg/L. Thus, the inhibitor concentration gradient was set to 0, 10, 20, 30, 40, and 50 mg/L to treat the cell wall deficient strain CC-849 and normal cell wall strain CC-124, respectively. The two algal strains were set with the same initial cell density, cultured under the same conditions for three days, and sampled in a 10 mL TAP medium containing different concentrations of inhibitor. Finally, the algal cells affected by the inhibitor were observed microscopically, and the permeability of the cell wall was measured according to
Section 4.2.
4.8. Transformation of Arabinosyltransferase Gene
4.8.1. Construction of Carrier
The arabinosyltransferase gene (RRA) is one of the most crucial function genes related to the synthesis of plant extensin. According to the results of transcriptome sequencing (4.5, 4.6) and enzyme activity inhibition experiment (4.7), arabinosyltransferase may play an important regulatory role in the cell wall synthesis in C. reinhardtii. Therefore, the RRA gene was knocked out to study its function.
The construction of the RRA knockout vector 007S-RRA-Hyg was based on the pClone007S vector. First, the 1107 bp RRA gene was inserted into the vector to obtain the plasmid 007S-RRA. Then, the 007S-RRA plasmid was digested with restriction endonuclease SacI and BsiwI. Concurrently, T4 ligase was used to link the β-tubulin promoter of C. reinhardtii, the aph7 gene, and the 3’ untranslated region of the rbcS2 to form the Hyg gene expression framework. Finally, by inserting the expression framework of the Hyg resistance gene with the homologous arm of the 007S-RRA plasmid into the RRA gene sequence using a DNA recombinant cloning kit (Vazyme, Nanjing, China), the knockout vector 007S-RRA-Hyg was obtained. The obtained plasmid 007S-RRA-Hyg was sequenced to verify that the sequence was correct and that the components in the cassette were connected correctly.
4.8.2. Glass Bead Transformation in C. reinhardtii
The transformation process in
C. reinhardtii is based on the method described in Kindle, which used sterilized glass microbeads to transform [
31]. The cells were cultured to a cell density of 3–5 × 10
6 cells per milliliter and then harvested by centrifugation at 3000×
g for 10 min. The cell density of 10
8 cells per milliliter was obtained by re-suspension precipitation using a TAP medium. The cell re-suspension was then swirled for 45 s on a vortex oscillator with 400 μg sterilized glass beads (diameter 425–600 μm) and 3–10 μg circular plasmid DNA. The mixture was inoculated into 5 mL fresh TAP medium, oscillated at 50 rpm at 23 °C, and cultured in a dark environment for 18 h. The algal fluid was then concentrated by harvesting the cells by centrifugation. The 100 μL concentrated
C. reinhardtii solution was gently and evenly spread on a TAP medium plate containing 1.5% Agar (containing 10 μg/mL hygromycin B). The plates were cultured upside down for 10–15 days, then monoclones were selected from the emerging algal colonies.
4.8.3. Screening of Mutants by Polymerase Chain Reaction
Monoclones were selected from the plate and inoculated into a 5 mL TAP liquid medium containing hygromycin B. The culture oscillated for three days, and then primers were designed according to the arabinosyltransferase gene and hygromycin B gene for PCR (
Table 6). According to the bands displayed on 1% agarose gel electrophoresis and gene sequencing, the transgenic algal strains, which were named 124-SR, were verified.
The permeability and integrity of the
C. reinhardtii cell wall were analyzed according to
Section 4.2.
5. Conclusions
Transcriptome sequencing technology was used to compare the differential expression genes related to cell wall component synthesis in cell wall deficient strain CC-849 in contrast to the normal cell wall strain CC-124. Similar to the glycosylation of the cell wall glycoprotein in the synthesis pathway of higher plant extensin, there is a metabolic pathway in C. reinhardtii, in which the expression of the arabinosyltransferase gene is up-regulated. Further restraining the action pathway of this enzyme by inhibitor, it was found that the cell wall of defective algal strain was not affected, while the permeability of the cell wall in normal algal strain was affected. Finally, utilizing genetic engineering, RRA was knocked out from CC-124, and the cell walls of the mutant strains showed unclear boundaries, loose structure, and reduced structure levels. It was speculated that inhibiting arabinosyltransferase would affect the glycosylation modification of cell wall proteins and further affect the assembly of cell wall glycoproteins.
The study of the structure and function of cell wall glycoproteins, as well as the further exploration of the cross-linking and assembly mechanism of cell wall components, has important guiding significance for understanding the evolution of cell wall glycoproteins in algae and higher plants. Based on a deep comprehension of the functions of these genes, gene transformation, gene editing, and other genetic engineering methods will provide new ideas for using these genes to construct more cell wall deficient strains suitable for different purposes such as aquatic feed, health food, drug delivery and so on.














{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}