
Comparative Analysis of Bivalve and Sea Urchin Genetics and Development: Investigating the Dichotomy in Bilateria


Abstract
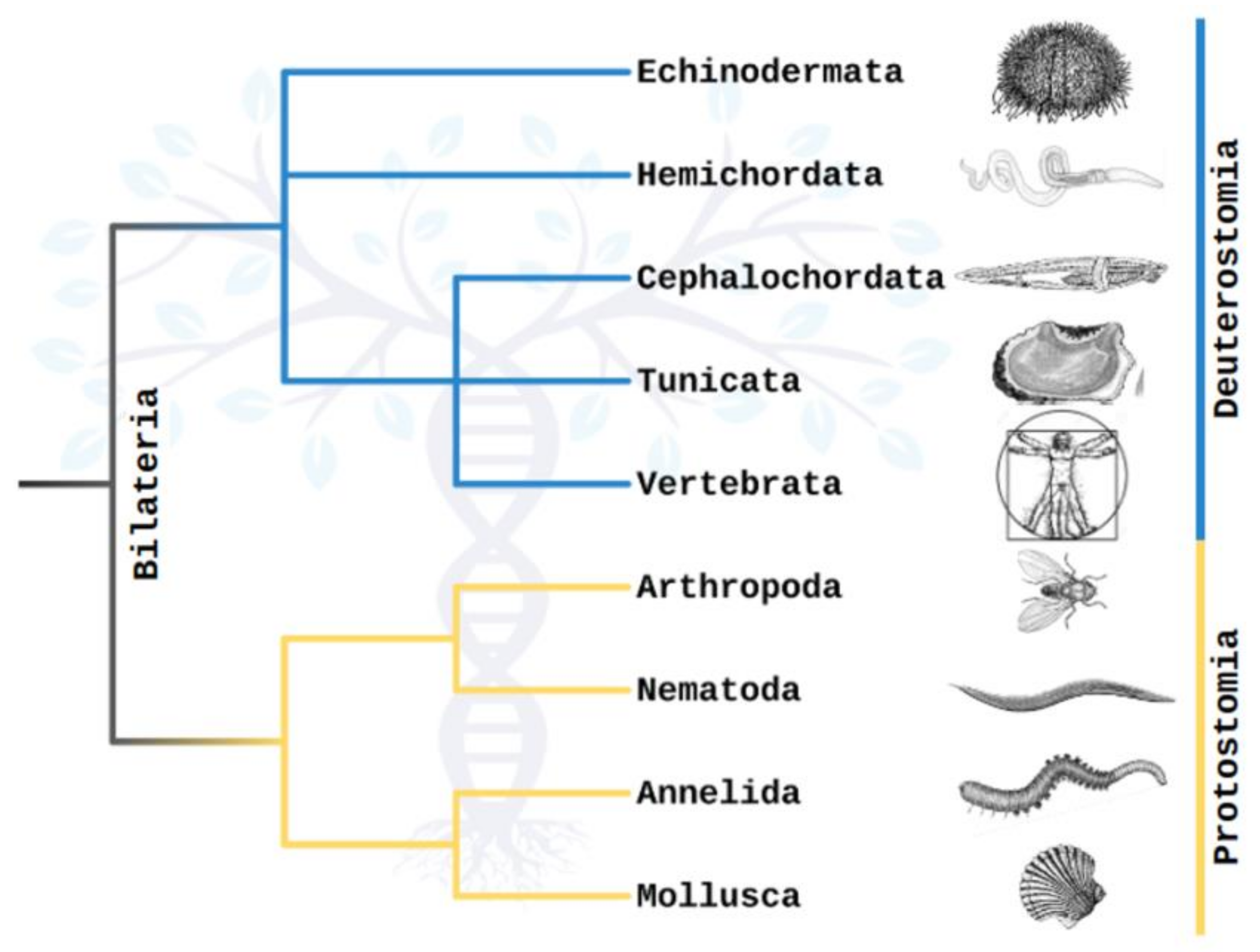
:1. Introduction
2. Comparative Embryogenesis of Bivalves and Sea Urchins: Insights and Distinctive Characteristics
3. Post-Genomic Era of Bivalve and Sea Urchins Studying
3.1. Genome Size and Complexity
3.2. Gene Organization and Density
4. Repetitive Elements and Non-Coding RNAs
5. Comparative Genomic Analyses
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Martindale, M.Q. Evolution of Development: The Details Are in the Entrails. Curr. Biol. 2013, 23, R25–R28. [Google Scholar] [CrossRef] [PubMed]
- Lochab, A.K.; Extavour, C.G. Bone Morphogenetic Protein (BMP) Signaling in Animal Reproductive System Development and Function. Dev. Biol. 2017, 427, 258–269. [Google Scholar] [CrossRef] [PubMed]
- Schulreich, S.M.; Salamanca-Díaz, D.A.; Zieger, E.; Calcino, A.D.; Wanninger, A. A Mosaic of Conserved and Novel Modes of Gene Expression and Morphogenesis in Mesoderm and Muscle Formation of a Larval Bivalve. Org. Divers. Evol. 2022, 22, 893–913. [Google Scholar] [CrossRef] [PubMed]
- Ayala, F.J.; Rzhetsky, A.; Ayala, F.J. Origin of the Metazoan Phyla: Molecular Clocks Confirm Paleontological Estimates. Proc. Natl. Acad. Sci. USA 1998, 95, 606–611. [Google Scholar] [CrossRef]
- Blair, J.E.; Hedges, S.B. Molecular Phylogeny and Divergence Times of Deuterostome Animals. Mol. Biol. Evol. 2005, 22, 2275–2284. [Google Scholar] [CrossRef] [PubMed]
- Giribet, G.; Edgecombe, G.D. The Invertebrate Tree of Life; Princeton University Press: Princeton, NJ, USA, 2020; ISBN 978-0-691-17025-1. [Google Scholar]
- Valentine, J.W. Cleavage Patterns and the Topology of the Metazoan Tree of Life. Proc. Natl. Acad. Sci. USA 1997, 94, 8001–8005. [Google Scholar] [CrossRef]
- Henry, J.Q.; Martindale, M.Q. Evolution of Cleavage Programs in Relationship to Axial Specification and Body Plan Evolution. Biol. Bull. 1998, 195, 363–366. [Google Scholar] [CrossRef]
- Gilbert, S.F. An Introduction to Early Developmental Processes. In Developmental Biology, 6th ed.; Sinauer Associates: Sunderland, MA, USA, 2000. [Google Scholar]
- Hejnol, A. A Twist in Time—The Evolution of Spiral Cleavage in the Light of Animal Phylogeny. Integr. Comp. Biol. 2010, 50, 695–706. [Google Scholar] [CrossRef]
- Gillies, T.E.; Cabernard, C. Cell Division Orientation in Animals. Curr. Biol. CB 2011, 21, R599–R609. [Google Scholar] [CrossRef]
- Kondo, M.; Akasaka, K. Current Status of Echinoderm Genome Analysis—What Do We Know? Curr. Genom. 2012, 13, 134–143. [Google Scholar] [CrossRef]
- Schuster, H.C.; Hirth, F. Phylogenetic Tracing of Midbrain-Specific Regulatory Sequences Suggests Single Origin of Eubilaterian Brains. Sci. Adv. 2023, 9, eade8259. [Google Scholar] [CrossRef]
- Martí-Solans, J.; Børve, A.; Bump, P.; Hejnol, A.; Lynagh, T. Peripheral and Central Employment of Acid-Sensing Ion Channels during Early Bilaterian Evolution. eLife 2023, 12, e81613. [Google Scholar] [CrossRef] [PubMed]
- Cavallo, A.; Clark, M.S.; Peck, L.S.; Harper, E.M.; Sleight, V.A. Evolutionary Conservation and Divergence of the Transcriptional Regulation of Bivalve Shell Secretion across Life-History Stages. R. Soc. Open Sci. 2022, 9, 221022. [Google Scholar] [CrossRef] [PubMed]
- Nicolini, F.; Ghiselli, F.; Luchetti, A.; Milani, L. Bivalves as Emerging Model Systems to Study the Mechanisms and Evolution of Sex Determination: A Genomic Point of View. Genome Biol. Evol. 2023, 15, evad181. [Google Scholar] [CrossRef]
- Ben-Tabou de-Leon, S.; Davidson, E.H. Experimentally Based Sea Urchin Gene Regulatory Network and the Causal Explanation of Developmental Phenomenology. Wiley Interdiscip. Rev. Syst. Biol. Med. 2009, 1, 237–246. [Google Scholar] [CrossRef]
- Martik, M.L.; Lyons, D.C.; McClay, D.R. Developmental Gene Regulatory Networks in Sea Urchins and What We Can Learn from Them. F1000Research 2016, 5, F1000. [Google Scholar] [CrossRef]
- Gudo, M.; Syed, T. 100 Years of Deuterostomia (Grobben, 1908): Cladogenetic and Anagenetic Relations within the Notoneuralia Domain. arXiv 2008, arXiv:0811.2189. [Google Scholar]
- Lovtrup, S. Validity of the Protostomia-Deuterostomia Theory. Syst. Zool. 1975, 24, 96–108. [Google Scholar] [CrossRef]
- Drozdov, A.L. Cytological Bases of Regulatory Development in Echinoderms and Determination in Mollusks. Biol. Bull. 2021, 48, 530–535. [Google Scholar] [CrossRef]
- Grobben, K. Die Systematische Einteilung des Tierreichs. Verh. Zool. Bot. Ges. Wien 1908, 58, 49l–511. [Google Scholar]
- Driesch, H. Die Isolirten Blastomeren des Echinidenkeimes. Arch. Entwicklungsmechanik Org. 1900, 10, 361–410. [Google Scholar] [CrossRef]
- Loeb, J. Ober Chemichen Bedingungen fur die Entstehumg Einluger Zwiliinge Beim Seeigel, Entwicklungsmech. Organismen 1909, 27, 119–140. [Google Scholar]
- Harvey, E.B. A New Method of Producing Twins, Triplets and Quadruplets in Arbacia punctulata, and Their Development. Biol. Bull. 1940, 78, 202–216. [Google Scholar] [CrossRef]
- Harvey, E.B. Some Surface Phenomena in the Fertilized Sea Urchin Egg as Influenced by Centrifugal Force. Biol. Bull. 1935, 69, 298–304. [Google Scholar] [CrossRef]
- Drozdov, A.L.; Svyatogor, G.P. Morphogenetic role of the cortex in the development of twins in sea urchins. Ontogenez 1987, 18, 540–546. [Google Scholar] [PubMed]
- Wilson, E.B. The Cell in Development and Heredity; Macmillan: New York, NY, USA, 1925. [Google Scholar]
- Drozdov, A.L. Ultrastructure of gametes and fertilization in Gray’s mussel. In Biology of Gray’s Mussel; Nauka: Novosibirsk, Russia, 1983; pp. 22–28. [Google Scholar]
- Drozdov, A.L.; Zavarzina, E.G.; Kulikova, V.A. Embryonic and larval development of Gray’s mussel. In Biology of Gray’s Mussel; Nauka: Novosibirsk, Russia, 1983; pp. 35–41. [Google Scholar]
- Malakhov, V.V. Embryonic Development of Bivalve Mollusks under Normal Conditions and under the Influence of Heavy Metals; Nauka: Novosibirsk, Russia, 1991; ISBN 978-5-02-005481-3. [Google Scholar]
- Devries, J. The formation and fate of the embryonic layers in the earthworm Eisinia foetida (annelida, oligocheta). Arch. Anat. Microsc. Morphol. Exp. 1973, 62, 15–38. [Google Scholar] [PubMed]
- Devries, J. The determination of the mesodermic metamerism specificity in the embryo of earthworm Eisenia foetida Sav. (Effect of bromodeoxyuridine. Arch. Biol. 1983, 94, 23–40. [Google Scholar]
- Peltrera, A. Le Capacità Regolative Dell’uovo di Aplysia limacina, L. Studiate con la Centrifugazione e con le Reazioni Vitali; Stazione Zoologica Anton Dohrn: Napoli, Italy, 1940. [Google Scholar]
- Zoja, R. Sullo Svillupo dei Blastomeri Isolati dale Uova di alcuni Medusae (e di altri Organismi). Wilhelm Rouxs Arch. 1895, 1, 578–595. [Google Scholar]
- Wolpert, L. From Soil Mechanics to Chick Development. Int. J. Dev. Biol. 2018, 62, 35–41. [Google Scholar] [CrossRef]
- Wolpert, L.; Hornbruch, A. Positional Signalling and the Development of the Humerus in the Chick Limb Bud. Dev. Camb. Engl. 1987, 100, 333–338. [Google Scholar] [CrossRef]
- Towers, M.; Wolpert, L.; Tickle, C. Gradients of Signalling in the Developing Limb. Curr. Opin. Cell Biol. 2012, 24, 181–187. [Google Scholar] [CrossRef]
- Vargesson, N.; Clarke, J.D.; Vincent, K.; Coles, C.; Wolpert, L.; Tickle, C. Cell Fate in the Chick Limb Bud and Relationship to Gene Expression. Dev. Camb. Engl. 1997, 124, 1909–1918. [Google Scholar] [CrossRef]
- Vargesson, N. Positional Information—A Concept Underpinning Our Understanding of Developmental Biology. Dev. Dyn. 2020, 249, 298–312. [Google Scholar] [CrossRef]
- Davidson, P.L.; Guo, H.; Swart, J.S.; Massri, A.J.; Edgar, A.; Wang, L.; Berrio, A.; Devens, H.R.; Koop, D.; Cisternas, P.; et al. Recent Reconfiguration of an Ancient Developmental Gene Regulatory Network in Heliocidaris Sea Urchins. Nat. Ecol. Evol. 2022, 6, 1907–1920. [Google Scholar] [CrossRef]
- Raven, C.H.R.P. Morphogenetic Analysis of Spiralian Development. Am. Zool. 1976, 16, 395–403. [Google Scholar] [CrossRef]
- Isaeva, V.V. Morphogenetic role of the cortical cytoskeleton and plasma membrane of the ovum. Tsitologiia 1984, 26, 5–13. [Google Scholar]
- Sawada, T.; Osanai, K. Cortical Contraction and Ooplasmic Movement in Centrifuged or Artificially Constricted Eggs of Ciona intestinalis. Wilhelm Rouxs Arch. Dev. Biol. 1984, 193, 127–132. [Google Scholar] [CrossRef]
- Sardet, C.; Speksnijder, J.; Inoue, S.; Jaffe, L. Fertilization and Ooplasmic Movements in the Ascidian Egg. Dev. Camb. Engl. 1989, 105, 237–249. [Google Scholar] [CrossRef]
- Shimizu, T. Cytoskeletal Mechanisms of Ooplasmic Segregation in Annelid Eggs. Int. J. Dev. Biol. 1999, 43, 11–18. [Google Scholar]
- Chung, J.T.; Downey, B.R.; Casper, R.F.; Chian, R.C. Effect of Centrifugation on Early Embryonic Development and Parthenogenetic Activation of Bovine Oocytes Matured in Vitro. Reprod. Fertil. Dev. 2001, 13, 383–388. [Google Scholar] [CrossRef]
- Tran, S.L.; Welte, M.A. In-Vivo Centrifugation of Drosophila Embryos. J. Vis. Exp. JoVE 2010, 40, e2005. [Google Scholar] [CrossRef]
- Drozdov, A.L. The Cortical Cytoskeleton of Non-Fertilized and Fertilized Surf Clam (Mactra chireusis). Citologiâ 1989, 31, 652–656. [Google Scholar]
- Zhang, G.; Fang, X.; Guo, X.; Li, L.; Luo, R.; Xu, F.; Yang, P.; Zhang, L.; Wang, X.; Qi, H.; et al. The Oyster Genome Reveals Stress Adaptation and Complexity of Shell Formation. Nature 2012, 490, 49–54. [Google Scholar] [CrossRef]
- The Genome of the Sea Urchin Strongylocentrotus purpuratus. Science 2006, 314, 941–952. [CrossRef]
- Materna, S.C.; Howard-Ashby, M.; Gray, R.F.; Davidson, E.H. The C2H2 Zinc Finger Genes of Strongylocentrotus purpuratus and Their Expression in Embryonic Development. Dev. Biol. 2006, 300, 108–120. [Google Scholar] [CrossRef]
- García-Deister, V. The Old Man and the Sea Urchin Genome: Theory and Data in the Work of Eric Davidson, 1969–2006. Hist. Philos. Life Sci. 2011, 33, 147–163. [Google Scholar]
- Cameron, R.A.; Rowen, L.; Nesbitt, R.; Bloom, S.; Rast, J.P.; Berney, K.; Arenas-Mena, C.; Martinez, P.; Lucas, S.; Richardson, P.M.; et al. Unusual Gene Order and Organization of the Sea Urchin Hox Cluster. J. Exp. Zool. B Mol. Dev. Evol. 2006, 306, 45–58. [Google Scholar] [CrossRef]
- Tu, Q.; Brown, C.T.; Davidson, E.H.; Oliveri, P. Sea Urchin Forkhead Gene Family: Phylogeny and Embryonic Expression. Dev. Biol. 2006, 300, 49–62. [Google Scholar] [CrossRef]
- Adonin, L.; Drozdov, A.; Barlev, N.A. Sea Urchin as a Universal Model for Studies of Gene Networks. Front. Genet. 2021, 11, 627259. [Google Scholar] [CrossRef]
- Canapa, A.; Barucca, M.; Biscotti, M.A.; Forconi, M.; Olmo, E. Transposons, Genome Size, and Evolutionary Insights in Animals. Cytogenet. Genome Res. 2015, 147, 217–239. [Google Scholar] [CrossRef]
- Yang, Z.; Zhang, L.; Hu, J.; Wang, J.; Bao, Z.; Wang, S. The Evo-Devo of Molluscs: Insights from a Genomic Perspective. Evol. Dev. 2020, 22, 409–424. [Google Scholar] [CrossRef] [PubMed]
- Yan, X.; Nie, H.; Huo, Z.; Ding, J.; Li, Z.; Yan, L.; Jiang, L.; Mu, Z.; Wang, H.; Meng, X.; et al. Clam Genome Sequence Clarifies the Molecular Basis of Its Benthic Adaptation and Extraordinary Shell Color Diversity. iScience 2019, 19, 1225–1237. [Google Scholar] [CrossRef] [PubMed]
- Calcino, A.D.; de Oliveira, A.L.; Simakov, O.; Schwaha, T.; Zieger, E.; Wollesen, T.; Wanninger, A. The Quagga Mussel Genome and the Evolution of Freshwater Tolerance. DNA Res. Int. J. Rapid Publ. Rep. Genes Genomes 2019, 26, 411–422. [Google Scholar] [CrossRef] [PubMed]
- Leffler, E.M.; Bullaughey, K.; Matute, D.R.; Meyer, W.K.; Ségurel, L.; Venkat, A.; Andolfatto, P.; Przeworski, M. Revisiting an Old Riddle: What Determines Genetic Diversity Levels within Species? PLoS Biol. 2012, 10, e1001388. [Google Scholar] [CrossRef] [PubMed]
- Corrochano-Fraile, A.; Davie, A.; Carboni, S.; Bekaert, M. Evidence of Multiple Genome Duplication Events in Mytilus Evolution. BMC Genom. 2022, 23, 340. [Google Scholar] [CrossRef]
- Rast, J.P.; Smith, L.C.; Loza-Coll, M.; Hibino, T.; Litman, G.W. Genomic Insights into the Immune System of the Sea Urchin. Science 2006, 314, 952–956. [Google Scholar] [CrossRef]
- Gildor, T.; Winter, M.R.; Layous, M.; Hijaze, E.; Ben-Tabou de-Leon, S. The Biological Regulation of Sea Urchin Larval Skeletogenesis—From Genes to Biomineralized Tissue. J. Struct. Biol. 2021, 213, 107797. [Google Scholar] [CrossRef]
- Amir, Y.; Insler, M.; Giller, A.; Gutman, D.; Atzmon, G. Senescence and Longevity of Sea Urchins. Genes 2020, 11, 573. [Google Scholar] [CrossRef]
- Marlétaz, F.; Couloux, A.; Poulain, J.; Labadie, K.; Da Silva, C.; Mangenot, S.; Noel, B.; Poustka, A.J.; Dru, P.; Pegueroles, C.; et al. Analysis of the P. lividus Sea Urchin Genome Highlights Contrasting Trends of Genomic and Regulatory Evolution in Deuterostomes. Cell Genom. 2023, 3, 100295. [Google Scholar] [CrossRef]
- Koop, D.; Cisternas, P.; Morris, V.B.; Strbenac, D.; Yang, J.Y.H.; Wray, G.A.; Byrne, M. Nodal and BMP Expression during the Transition to Pentamery in the Sea Urchin Heliocidaris erythrogramma: Insights into Patterning the Enigmatic Echinoderm Body Plan. BMC Dev. Biol. 2017, 17, 4. [Google Scholar] [CrossRef]
- Morris, V.B.; Kable, E.; Koop, D.; Cisternas, P.; Byrne, M. Early Development of the Feeding Larva of the Sea Urchin Heliocidaris tuberculata: Role of the Small Micromeres. Dev. Genes Evol. 2019, 229, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Takeuchi, T.; Koyanagi, R.; Gyoja, F.; Kanda, M.; Hisata, K.; Fujie, M.; Goto, H.; Yamasaki, S.; Nagai, K.; Morino, Y.; et al. Bivalve-Specific Gene Expansion in the Pearl Oyster Genome: Implications of Adaptation to a Sessile Lifestyle. Zool. Lett. 2016, 2, 3. [Google Scholar] [CrossRef]
- Sun, Y.; Liu, X.; Xie, X.; Bai, Y.; Wang, S.; Teng, W.; Li, D.; Li, H.; Yu, Z.; Zhang, M.; et al. A High-Quality Chromosome-Level Genome Assembly of the Bivalve Mollusk Mactra veneriformis. G3 Genes|Genomes|Genetics 2022, 12, jkac229. [Google Scholar] [CrossRef]
- Feschotte, C. Transposable Elements and the Evolution of Regulatory Networks. Nat. Rev. Genet. 2008, 9, 397–405. [Google Scholar] [CrossRef]
- Zattera, M.L.; Bruschi, D.P. Transposable Elements as a Source of Novel Repetitive DNA in the Eukaryote Genome. Cells 2022, 11, 3373. [Google Scholar] [CrossRef]
- Bourque, G.; Burns, K.H.; Gehring, M.; Gorbunova, V.; Seluanov, A.; Hammell, M.; Imbeault, M.; Izsvák, Z.; Levin, H.L.; Macfarlan, T.S.; et al. Ten Things You Should Know about Transposable Elements. Genome Biol. 2018, 19, 199. [Google Scholar] [CrossRef]
- Wicker, T.; Sabot, F.; Hua-Van, A.; Bennetzen, J.L.; Capy, P.; Chalhoub, B.; Flavell, A.; Leroy, P.; Morgante, M.; Panaud, O.; et al. A Unified Classification System for Eukaryotic Transposable Elements. Nat. Rev. Genet. 2007, 8, 973–982. [Google Scholar] [CrossRef] [PubMed]
- Makałowski, W.; Gotea, V.; Pande, A.; Makałowska, I. Transposable Elements: Classification, Identification, and Their Use as a Tool for Comparative Genomics. Methods Mol. Biol. 2019, 1910, 177–207. [Google Scholar] [CrossRef]
- Kojima, K.K. Structural and Sequence Diversity of Eukaryotic Transposable Elements. Genes Genet. Syst. 2020, 94, 233–252. [Google Scholar] [CrossRef]
- Martelossi, J.; Nicolini, F.; Subacchi, S.; Pasquale, D.; Ghiselli, F.; Luchetti, A. Multiple and Diversified Transposon Lineages Contribute to Early and Recent Bivalve Genome Evolution. BMC Biol. 2023, 21, 145. [Google Scholar] [CrossRef]
- Metzger, M.J.; Paynter, A.N.; Siddall, M.E.; Goff, S.P. Horizontal Transfer of Retrotransposons between Bivalves and Other Aquatic Species of Multiple Phyla. Proc. Natl. Acad. Sci. USA 2018, 115, E4227–E4235. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; He, J.; Tan, P.; Gong, Z.; Qian, S.; Miao, Y.; Zhang, H.-Y.; Tu, G.; Chen, Q.; Zhong, Q.; et al. The Genome of an Apodid Holothuroid (Chiridota heheva) Provides Insights into Its Adaptation to a Deep-Sea Reducing Environment. Commun. Biol. 2022, 5, 224. [Google Scholar] [CrossRef]
- Pappalardo, A.M.; Ferrito, V.; Biscotti, M.A.; Canapa, A.; Capriglione, T. Transposable Elements and Stress in Vertebrates: An Overview. Int. J. Mol. Sci. 2021, 22, 1970. [Google Scholar] [CrossRef] [PubMed]
- Panyushev, N.; Okorokova, L.; Danilov, L.; Adonin, L. Pattern of Repetitive Element Transcription Segregate Cell Lineages during the Embryogenesis of Sea Urchin Strongylocentrotus purpuratus. Biomedicines 2021, 9, 1736. [Google Scholar] [CrossRef]
- Hamilton, A.J.; Baulcombe, D.C. A Species of Small Antisense RNA in Posttranscriptional Gene Silencing in Plants. Science 1999, 286, 950–952. [Google Scholar] [CrossRef] [PubMed]
- Waterhouse, P.M.; Graham, M.W.; Wang, M.B. Virus Resistance and Gene Silencing in Plants Can Be Induced by Simultaneous Expression of Sense and Antisense RNA. Proc. Natl. Acad. Sci. USA 1998, 95, 13959–13964. [Google Scholar] [CrossRef]
- Hadjiargyrou, M.; Delihas, N. The Intertwining of Transposable Elements and Non-Coding RNAs. Int. J. Mol. Sci. 2013, 14, 13307–13328. [Google Scholar] [CrossRef]
- Kapusta, A.; Kronenberg, Z.; Lynch, V.J.; Zhuo, X.; Ramsay, L.; Bourque, G.; Yandell, M.; Feschotte, C. Transposable Elements Are Major Contributors to the Origin, Diversification, and Regulation of Vertebrate Long Noncoding RNAs. PLoS Genet. 2013, 9, e1003470. [Google Scholar] [CrossRef]
- Fort, V.; Khelifi, G.; Hussein, S.M.I. Long Non-Coding RNAs and Transposable Elements: A Functional Relationship. Biochim. Biophys. Acta Mol. Cell Res. 2021, 1868, 118837. [Google Scholar] [CrossRef]
- Zhang, P.; Wu, W.; Chen, Q.; Chen, M. Non-Coding RNAs and Their Integrated Networks. J. Integr. Bioinform. 2019, 16, 20190027. [Google Scholar] [CrossRef]
- Lagos-Quintana, M.; Rauhut, R.; Lendeckel, W.; Tuschl, T. Identification of Novel Genes Coding for Small Expressed RNAs. Science 2001, 294, 853–858. [Google Scholar] [CrossRef]
- Lee, R.C.; Feinbaum, R.L.; Ambros, V.; The, C. Elegans Heterochronic Gene Lin-4 Encodes Small RNAs with Antisense Complementarity to Lin-14. Cell 1993, 75, 843–854. [Google Scholar] [CrossRef] [PubMed]
- Reinhart, B.J.; Slack, F.J.; Basson, M.; Pasquinelli, A.E.; Bettinger, J.C.; Rougvie, A.E.; Horvitz, H.R.; Ruvkun, G. The 21-Nucleotide Let-7 RNA Regulates Developmental Timing in Caenorhabditis Elegans. Nature 2000, 403, 901–906. [Google Scholar] [CrossRef] [PubMed]
- Kozomara, A.; Birgaoanu, M.; Griffiths-Jones, S. miRBase: From microRNA Sequences to Function. Nucleic Acids Res. 2019, 47, D155–D162. [Google Scholar] [CrossRef]
- Ma, L.; Bajic, V.B.; Zhang, Z. On the Classification of Long Non-Coding RNAs. RNA Biol. 2013, 10, 925–933. [Google Scholar] [CrossRef]
- Kornienko, A.E.; Guenzl, P.M.; Barlow, D.P.; Pauler, F.M. Gene Regulation by the Act of Long Non-Coding RNA Transcription. BMC Biol. 2013, 11, 59. [Google Scholar] [CrossRef] [PubMed]
- Campo-Paysaa, F.; Sémon, M.; Cameron, R.A.; Peterson, K.J.; Schubert, M. microRNA Complements in Deuterostomes: Origin and Evolution of microRNAs. Evol. Dev. 2011, 13, 15–27. [Google Scholar] [CrossRef] [PubMed]
- Ozata, D.M.; Gainetdinov, I.; Zoch, A.; O’Carroll, D.; Zamore, P.D. PIWI-Interacting RNAs: Small RNAs with Big Functions. Nat. Rev. Genet. 2019, 20, 89–108. [Google Scholar] [CrossRef]
- Wang, C.; Lin, H. Roles of piRNAs in Transposon and Pseudogene Regulation of Germline mRNAs and lncRNAs. Genome Biol. 2021, 22, 27. [Google Scholar] [CrossRef]
- Nojima, T.; Proudfoot, N.J. Mechanisms of lncRNA Biogenesis as Revealed by Nascent Transcriptomics. Nat. Rev. Mol. Cell Biol. 2022, 23, 389–406. [Google Scholar] [CrossRef]
- Mattick, J.S.; Amaral, P.P.; Carninci, P.; Carpenter, S.; Chang, H.Y.; Chen, L.-L.; Chen, R.; Dean, C.; Dinger, M.E.; Fitzgerald, K.A.; et al. Long Non-Coding RNAs: Definitions, Functions, Challenges and Recommendations. Nat. Rev. Mol. Cell Biol. 2023, 24, 430–447. [Google Scholar] [CrossRef] [PubMed]
- Zhou, W.-Y.; Cai, Z.-R.; Liu, J.; Wang, D.-S.; Ju, H.-Q.; Xu, R.-H. Circular RNA: Metabolism, Functions and Interactions with Proteins. Mol. Cancer 2020, 19, 172. [Google Scholar] [CrossRef] [PubMed]
- Patil, V.S.; Zhou, R.; Rana, T.M. Gene Regulation by Noncoding RNAs. Crit. Rev. Biochem. Mol. Biol. 2014, 49, 16–32. [Google Scholar] [CrossRef] [PubMed]
- Jehn, J.; Gebert, D.; Pipilescu, F.; Stern, S.; Kiefer, J.S.T.; Hewel, C.; Rosenkranz, D. PIWI Genes and piRNAs Are Ubiquitously Expressed in Mollusks and Show Patterns of Lineage-Specific Adaptation. Commun. Biol. 2018, 1, 137. [Google Scholar] [CrossRef]
- Ramat, A.; Simonelig, M. Functions of PIWI Proteins in Gene Regulation: New Arrows Added to the piRNA Quiver. Trends Genet. TIG 2021, 37, 188–200. [Google Scholar] [CrossRef]
- Huang, S.; Yoshitake, K.; Asakawa, S. A Review of Discovery Profiling of PIWI-Interacting RNAs and Their Diverse Functions in Metazoans. Int. J. Mol. Sci. 2021, 22, 11166. [Google Scholar] [CrossRef]
- Yajima, M.; Gustafson, E.A.; Song, J.L.; Wessel, G.M. Piwi Regulates Vasa Accumulation during Embryogenesis in the Sea Urchin. Dev. Dyn. Off. Publ. Am. Assoc. Anat. 2014, 243, 451–458. [Google Scholar] [CrossRef]
- Rosani, U.; Pallavicini, A.; Venier, P. The miRNA Biogenesis in Marine Bivalves. PeerJ 2016, 4, e1763. [Google Scholar] [CrossRef]
- Chen, J.; Zhai, Z.; Lu, L.; Li, S.; Guo, D.; Bai, L.; Yu, D. Identification and Characterization of miRNAs and Their Predicted mRNAs in the Larval Development of Pearl Oyster Pinctada fucata. Mar. Biotechnol. 2022, 24, 303–319. [Google Scholar] [CrossRef]
- Niu, D.; Li, B.; Xie, S.; Dong, Z.; Li, J. Integrated mRNA and Small RNA Sequencing Reveals Regulatory Expression of Larval Metamorphosis of the Razor Clam. Mar. Biotechnol. 2020, 22, 696–705. [Google Scholar] [CrossRef]
- Song, J.L.; Stoeckius, M.; Maaskola, J.; Friedländer, M.; Stepicheva, N.; Juliano, C.; Lebedeva, S.; Thompson, W.; Rajewsky, N.; Wessel, G.M. Select microRNAs Are Essential for Early Development in the Sea Urchin. Dev. Biol. 2012, 362, 104–113. [Google Scholar] [CrossRef] [PubMed]
- Sampilo, N.F.; Stepicheva, N.A.; Zaidi, S.A.M.; Wang, L.; Wu, W.; Wikramanayake, A.; Song, J.L. Inhibition of microRNA Suppression of Dishevelled Results in Wnt Pathway-Associated Developmental Defects in Sea Urchin. Dev. Camb. Engl. 2018, 145, dev167130. [Google Scholar] [CrossRef]
- Okamitsu, Y.; Yamamoto, T.; Fujii, T.; Ochiai, H.; Sakamoto, N. Dicer Is Required for the Normal Development of Sea Urchin, Hemicentrotus pulcherrimus. Zool. Sci. 2010, 27, 477–486. [Google Scholar] [CrossRef]
- Stepicheva, N.A.; Song, J.L. microRNA-31 Modulates Skeletal Patterning in the Sea Urchin Embryo. Dev. Camb. Engl. 2015, 142, 3769–3780. [Google Scholar] [CrossRef]
- Sampilo, N.F.; Stepicheva, N.A.; Song, J.L. microRNA-31 Regulates Skeletogenesis by Direct Suppression of Eve and Wnt1. Dev. Biol. 2021, 472, 98–114. [Google Scholar] [CrossRef]
- Stepicheva, N.; Nigam, P.A.; Siddam, A.D.; Peng, C.F.; Song, J.L. microRNAs Regulate β-Catenin of the Wnt Signaling Pathway in Early Sea Urchin Development. Dev. Biol. 2015, 402, 127–141. [Google Scholar] [CrossRef]
- Pauli, A.; Rinn, J.L.; Schier, A.F. Non-Coding RNAs as Regulators of Embryogenesis. Nat. Rev. Genet. 2011, 12, 136. [Google Scholar] [CrossRef]
- Yu, H.; Zhao, X.; Li, Q. Genome-Wide Identification and Characterization of Long Intergenic Noncoding RNAs and Their Potential Association with Larval Development in the Pacific Oyster. Sci. Rep. 2016, 6, 20796. [Google Scholar] [CrossRef]
- Evensen, K.G.; Robinson, W.E.; Krick, K.; Murray, H.M.; Poynton, H.C. Comparative Phylotranscriptomics Reveals Putative Sex Differentiating Genes across Eight Diverse Bivalve Species. Comp. Biochem. Physiol. Part D Genom. Proteom. 2022, 41, 100952. [Google Scholar] [CrossRef]
- Wang, Q.; Cao, T.; Wang, Y.; Li, X.; Wang, Y. Genome-Wide Identification and Comparative Analysis of Dmrt Genes in Echinoderms. Sci. Rep. 2023, 13, 7664. [Google Scholar] [CrossRef]
- Ottolenghi, C.; Pelosi, E.; Tran, J.; Colombino, M.; Douglass, E.; Nedorezov, T.; Cao, A.; Forabosco, A.; Schlessinger, D. Loss of Wnt4 and Foxl2 Leads to Female-to-Male Sex Reversal Extending to Germ Cells. Hum. Mol. Genet. 2007, 16, 2795–2804. [Google Scholar] [CrossRef]
- Zhang, N.; Xu, F.; Guo, X. Genomic Analysis of the Pacific Oyster (Crassostrea gigas) Reveals Possible Conservation of Vertebrate Sex Determination in a Mollusc. G3 Genes|Genomes|Genetics 2014, 4, 2207–2217. [Google Scholar] [CrossRef] [PubMed]
- Yaguchi, S.; Yaguchi, J.; Angerer, R.C.; Angerer, L.M. A Wnt-FoxQ2-Nodal Pathway Links Primary and Secondary Axis Specification in Sea Urchin Embryos. Dev. Cell 2008, 14, 97–107. [Google Scholar] [CrossRef] [PubMed]
- Rho, H.K.; McClay, D.R. The Control of foxN2/3 Expression in Sea Urchin embryos and Its Function in the Skeletogenic Gene Regulatory Network. Dev. Camb. Engl. 2011, 138, 937–945. [Google Scholar] [CrossRef]
- David, B.; Mooi, R. How Hox Genes Can Shed Light on the Place of Echinoderms among the Deuterostomes. EvoDevo 2014, 5, 22. [Google Scholar] [CrossRef]
- Simakov, O.; Marletaz, F.; Cho, S.-J.; Edsinger-Gonzales, E.; Havlak, P.; Hellsten, U.; Kuo, D.-H.; Larsson, T.; Lv, J.; Arendt, D.; et al. Insights into Bilaterian Evolution from Three Spiralian Genomes. Nature 2013, 493, 526–531. [Google Scholar] [CrossRef]
- Li, Y.; Sun, X.; Hu, X.; Xun, X.; Zhang, J.; Guo, X.; Jiao, W.; Zhang, L.; Liu, W.; Wang, J.; et al. Scallop Genome Reveals Molecular Adaptations to Semi-Sessile Life and Neurotoxins. Nat. Commun. 2017, 8, 1721. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Zhang, J.; Jiao, W.; Li, J.; Xun, X.; Sun, Y.; Guo, X.; Huan, P.; Dong, B.; Zhang, L.; et al. Scallop Genome Provides Insights into Evolution of Bilaterian Karyotype and Development. Nat. Ecol. Evol. 2017, 1, 0120. [Google Scholar] [CrossRef]
- Pérez-Parallé, M.L.; Pazos, A.J.; Mesías-Gansbiller, C.; Sánchez, J.L. Hox, Parahox, Ehgbox, and NK Genes in Bivalve molluscs: Evolutionary Implications. J. Shellfish Res. 2016, 35, 179–190. [Google Scholar] [CrossRef]
- Yan, C.; Xu, M.; Ye, Y.; Gu, Z.; Huang, J.; Guo, B.; Qi, P.; Li, J.; Yan, X. Hox Gene Clusters in the Mussel Mytilus Coruscus: Implications for Bivalves’ Evolution. Ecol. Indic. 2023, 154, 110810. [Google Scholar] [CrossRef]
- Salamanca-Díaz, D.A.; Calcino, A.D.; de Oliveira, A.L.; Wanninger, A. Non-Collinear Hox Gene Expression in Bivalves and the Evolution of Morphological Novelties in Mollusks. Sci. Rep. 2021, 11, 3575. [Google Scholar] [CrossRef] [PubMed]
- Paps, J.; Xu, F.; Zhang, G.; Holland, P.W.H. Reinforcing the Egg-Timer: Recruitment of Novel Lophotrochozoa Homeobox Genes to Early and Late Development in the Pacific Oyster. Genome Biol. Evol. 2015, 7, 677–688. [Google Scholar] [CrossRef] [PubMed]
- Arnone, M.I.; Rizzo, F.; Annunciata, R.; Cameron, R.A.; Peterson, K.J.; Martínez, P. Genetic Organization and Embryonic Expression of the ParaHox Genes in the Sea Urchin S. purpuratus: Insights into the Relationship between Clustering and Colinearity. Dev. Biol. 2006, 300, 63–73. [Google Scholar] [CrossRef] [PubMed]
- Yamaguchi, S.; Hano, Y.; Hayashi, A.; Yamaguchi, M. The Hox Genes of the Direct-Type Developing Sea Urchin Peronella japonica. Zygote Camb. Engl. 2000, 8 (Suppl. S1), S73. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Name | Assembly | GenBank | Genome Size | Assembly Level | |
|---|---|---|---|---|---|
| Sea urchins | S. purpuratus | Spur_5.0 | GCA_000002235.4 | 921.8 Mb | Scaffold |
| E. tribuloides | Etri_1.0 | GCA_001188425.1 | 2.2 Gb | Scaffold | |
| H. pulcherrimus | HpulGenome_v1 | GCA_003118195.1 | 568.9 Mb | Scaffold | |
| Bivalves | C. gigas | cgigas_uk_roslin_v1 | GCA_902806645.1 | 647.9 Mb | Chromosome |
| S. cumingii | ASM2855479v1 | GCA_028554795.1 | 3.4 Gb | Chromosome | |
| L. rhynchaena | LuRhyn_1.0 | GCA_008271625.1 | 543.9 Mb | Contig |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Drozdov, A.; Lebedev, E.; Adonin, L. Comparative Analysis of Bivalve and Sea Urchin Genetics and Development: Investigating the Dichotomy in Bilateria. Int. J. Mol. Sci. 2023, 24, 17163. https://doi.org/10.3390/ijms242417163
Drozdov A, Lebedev E, Adonin L. Comparative Analysis of Bivalve and Sea Urchin Genetics and Development: Investigating the Dichotomy in Bilateria. International Journal of Molecular Sciences. 2023; 24(24):17163. https://doi.org/10.3390/ijms242417163
Chicago/Turabian StyleDrozdov, Anatoliy, Egor Lebedev, and Leonid Adonin. 2023. "Comparative Analysis of Bivalve and Sea Urchin Genetics and Development: Investigating the Dichotomy in Bilateria" International Journal of Molecular Sciences 24, no. 24: 17163. https://doi.org/10.3390/ijms242417163