
Sitagliptin Induces Tolerogenic Human Dendritic Cells
 , , , , and
, , , , and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Cytotoxicity of Sitagliptin in MoDC Cultures
2.2. Sitagliptin Impairs Differentiation and Maturation of MoDCs
2.3. Sitagliptin Up-Regulates the Expression of CD26 on MoDCs
2.4. Sitagliptin Modulates Cytokine Production by MoDCs and Inhibits Their Allostimulatory Capacity
2.5. Sitagliptin Modulates T Helper Polarization Capability of MoDCs
2.6. Tolerogenic Properties of MoDCs Treated with Sitagliptin
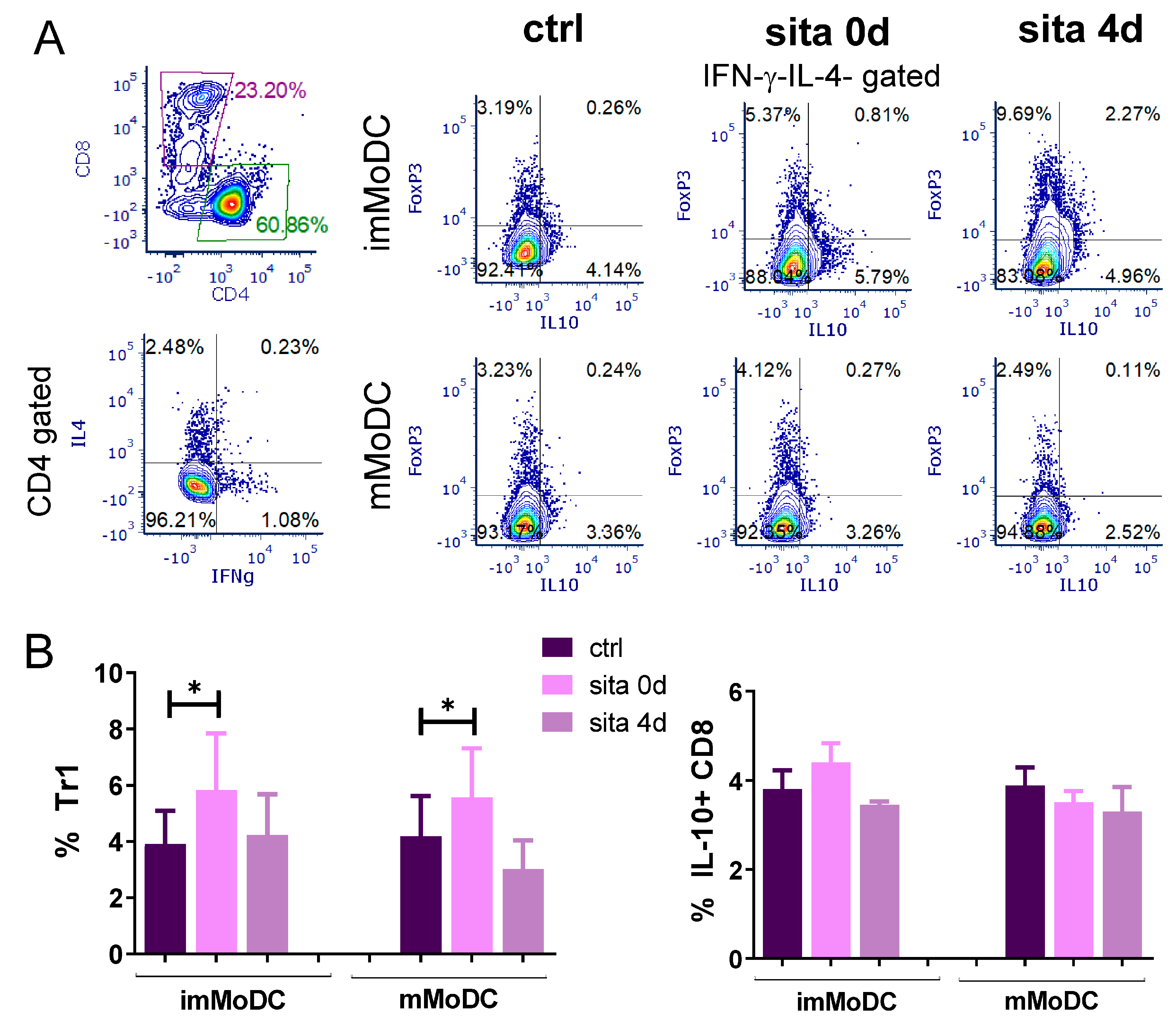
2.7. Induction of Tr1 Cells by Sitagliptin-Treated MoDCs
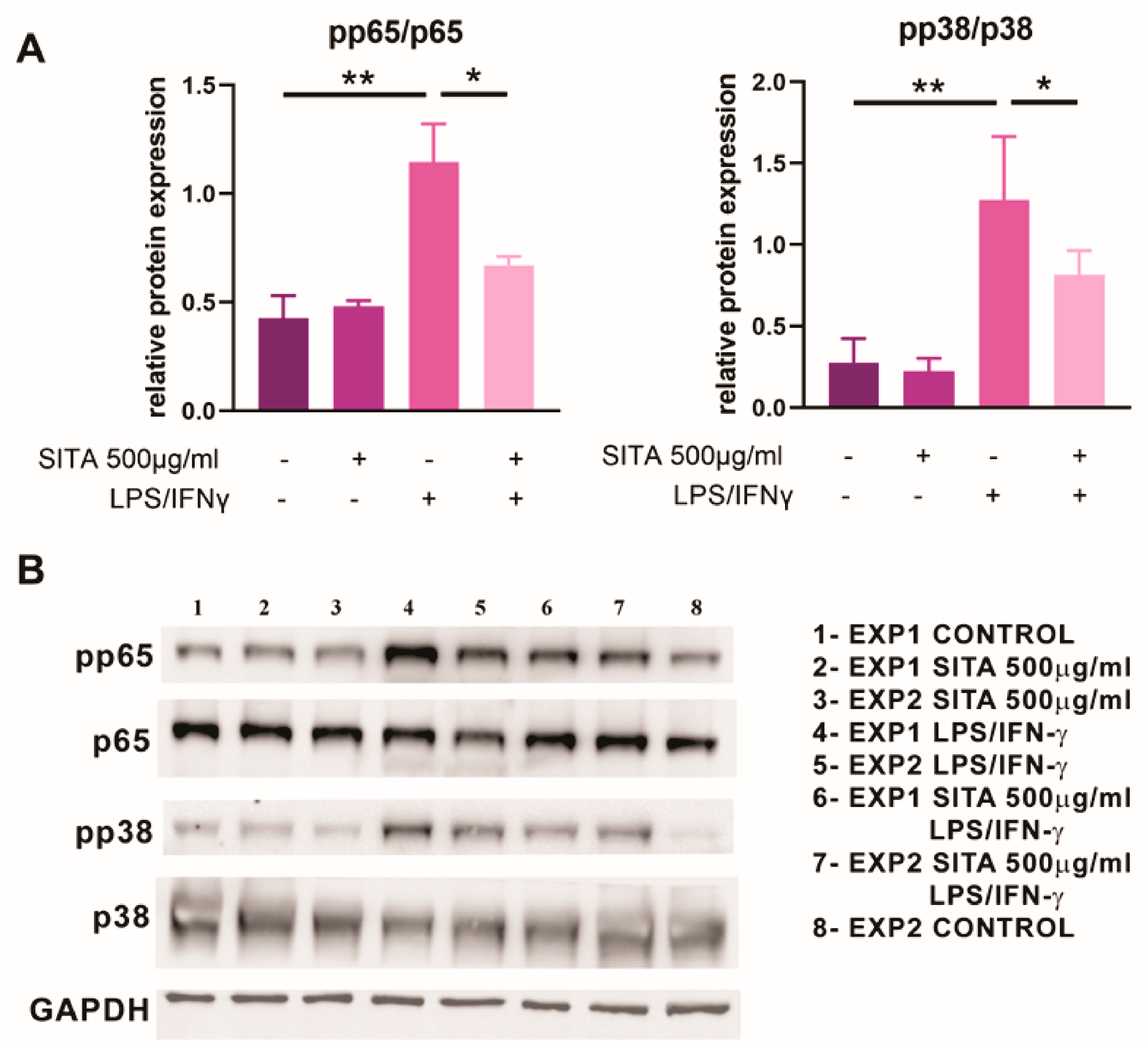
2.8. Sitagliptin Inhibits the Expression of p65 NF-kB and p38 MAPK in MoDCs
3. Discussion
4. Materials and Methods
4.1. Generation of MoDCs
4.2. Cytotoxicity Assays
4.3. Mixed Leukocyte Reaction
4.4. Th Polarization and Tregs Induction
4.5. Cytokine Measurement
4.6. Flow Cytometry
4.7. Western Blot
4.8. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| DPP-4 | Dipeptidyl peptidase-4 |
| GLP1 | Glucagon-like peptide 1 |
| GIP | Glucose-dependent insulinotropic peptide |
| T2DM | Type 2 Diabetes Mellitus |
| PBMCs | Human peripheral blood mononuclear cells |
| Ig | Immunoglobulin |
| APCs | Antigen-presenting cells |
| DCs | Dendritic cells |
| MoDCs | Human DCs differentiated from monocytes in vitro |
| ImMoDCs | Immature MoDCs |
| mMoDCs | Mature MoDCs |
| Treg | Regulatory T cell |
| Th1 | Type 1 T helper cell |
| Th2 | Type 2 T helper cell |
| Th17 | Type 17 T helper cell |
| CTLs | Cytotoxic T lymphocytes |
| NF-kB | Nuclear Factor kappa B |
| FoxP3 | Forkhead box Protein 3 |
| PHA | Phytohemagglutinin |
| TGF-β | Transforming Growth Factor-β |
| LPS | Lipopolysaccharide |
| TNF-α | Tumor Necrosis Factor-α |
| IL | Interleukin |
| IFN-γ | Interferon-γ |
| ILT4 | Immunoglobulin-Like Transcript 4 |
| ILT3 | Immunoglobulin-Like Transcript 3 |
| IDO1 | Indoleamine 2,3-Dioxygenase 1 |
| MAPK | Mitogen-Activated Protein Kinase |
| CTLA-4 | Cytotoxic T Lymphocyte Antigen 4 |
| MLR | Mixed leukocyte reaction |
| GM-CSF | Granulocyte-Macrophage Colony Stimulating Factor |
| SDS | Sodium dodecyl sulfate |
| BCL6 | B-Cell Lymphoma 6 |
| ELISA | Enzyme-linked immunosorbent assay |
| MACS | Magnetic-activated cell sorting |
| LADA | Latent Autoimmune Diabetes of Adults |
| HLA-DR | Human Leukocyte Antigen-DR isotype |
| CD | Cluster of differentiation |
| NLRP3 | NLR family pyrin domain containing 3 |
| ICOS-L | Inducible Costimulator Ligand |
| ADA | Adenosine Deaminase |
| PD-1 | Programmed cell death protein 1 |
| PD-L1 | Programmed death-ligand 1 |
| RAR | Retinoic Acid Receptor |
| RORγt | RAR-related orphan receptor gamma |
| HUVECs | Human umbilical vein endothelial cells |
| FITC | Fluorescein isothiocyanate |
| APC | Allophycocyanin |
| PE | Phycoerythrin |
| 7-AAD | 7-Aminoactinomycin D |
| PerCP/Cy | Peridinin chlorophyll protein-Cyanine |
| PMA | Phorbol 12-myristate 13-acetate |
| FMO | Fluorescence minus one |
| FSC | Forward scatter |
| SSC | Side scatter |
| OD | Optical density |
References
- Hopsu-Havu, V.K.; Glenner, G.G. A new dipeptide naphthylamidase hydrolyzing glycyl-prolyl-beta-naphthylamide. Histochemie 1966, 7, 197–201. [Google Scholar] [CrossRef] [PubMed]
- Vincenzo, F.; Manfredi, T.; Carmine, C.; Mario, R. CD26: A multi-purpose pharmacological target. Curr. Clin. Pharmacol. 2014, 9, 157–164. [Google Scholar] [CrossRef] [PubMed]
- Dang, N.H.; Morimoto, C. CD26: An expanding role in immune regulation and cancer. Histol. Histopathol. 2002, 17, 1213–1226. [Google Scholar]
- Klemann, C.; Wagner, L.; Stephan, M.; von Hosten, S. Cut to the chase: A review of CD26/dipeptidyl peptidase-4’s (DPP4) entanglement in the immune system. Clin. Exp. Immunol. 2016, 185, 1–21. [Google Scholar] [CrossRef] [PubMed]
- Wagner, L.; Klemann, C.; Stephan, M.; von Horsten, S. Unravelling the immunological roles of dipeptidyl peptidase 4 (DPP4) activity and/or structure homologue (DASH) proteins. Clin. Exp. Immunol. 2016, 184, 265–283. [Google Scholar] [CrossRef] [PubMed]
- Gschwandtner, M.; Paultischke, V.; Midner, M.; Brunner, P.M.; Hacker, S.; Eisenwort, G.; Sperr, W.R.; Valent, P.; Gerner, C.; Tschachler, E. Proteome analysis identifies L1CAM/CD171 and DPP4/CD26 as novel markers of human skin mast cells. Allergy 2017, 72, 85–97. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Grigo, C.; Steinbeck, J.; von Horsten, S.; Amann, K.; Daniel, C. Soluble DPP4 originates in part from bone marrow cells and not from the kidney. Peptides 2014, 57, 109–117. [Google Scholar] [CrossRef] [PubMed]
- Gorrell, M.D.; Gysbers, V.; McCaughan, G.W. CD26: A multifunctional integral membrane and secreted protein of activated lymphocytes. Scand. J. Immunol. 2001, 54, 249–264. [Google Scholar] [CrossRef]
- Nauck, M.A.; Quast, D.R.; Wefers, J.; Pfeiffer, A.F.H. The evolving story of incretins (GIP and GLP-1) in metabolic and cardiovascular disease: A pathophysiological update. Diabetes Obes. Metab. 2021, 23 (Suppl. 3), 5–29. [Google Scholar] [CrossRef]
- Gilbert, M.P.; Pratley, R.E. GLP-1 Analogs and DPP-4 Inhibitors in Type 2 Diabetes Therapy: Review of Head-to-Head Clinical Trials. Front. Endocrinol. 2020, 11, 178. [Google Scholar] [CrossRef]
- Deacon, C.F. Peptide degradation and the role of DPP-4 inhibitors in the treatment of type 2 diabetes. Peptides 2018, 100, 150–157. [Google Scholar] [CrossRef] [PubMed]
- Bohannon, N. Overview of the gliptin class (dipeptidyl peptidase-4 inhibitors) in clinical practice. Postgrad. Med. 2009, 121, 40–45. [Google Scholar] [CrossRef] [PubMed]
- Choy, M.; Lam, S. Sitagliptin: A novel drug for the treatment of type 2 diabetes. Cardiol. Rev. 2007, 15, 264–271. [Google Scholar] [CrossRef] [PubMed]
- Scott, L.J. Sitagliptin: A Review in Type 2 Diabetes. Drugs 2017, 77, 209–224. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.; Kim, Y.; Yang, E.; Kwon, E.J.; Kim, S.; Kang, S.; Park, D.B.; Oh, B.C.; Kim, J.; Heo, S.; et al. CD26/DPP4 levels in Peripheral Blood and T cells in Patients with Type 2 Diabetes Mellitus. J. Clin. Endocrinol. Metab. 2013, 98, 2553–2561. [Google Scholar] [CrossRef] [PubMed]
- Makdissi, A.; Ghanim, H.; Vora, M.; Green, K.; Abuaysheh, S.; Chaudhuri, A.; Dhindsa, S.; Dandona, P. Sitagliptin exerts an antinflammatory action. J. Clin. Endocrinol. Metab. 2012, 97, 3333–3341. [Google Scholar] [CrossRef] [PubMed]
- Zhuge, F.; Ni, Y.; Nagashimada, M.; Nagata, N.; Xu, L.; Mukaida, N.; Kaneko, S.; Ota, T. DPP-4 Inhibition by Linagliptin Attenuates Obesity-Related Inflammation and Insulin Resistance by Regulating M1/M2 Macrophage Polarization. Diabetes 2016, 65, 2966–2979. [Google Scholar] [CrossRef] [PubMed]
- Li, B.; Luo, Y.R.; Zhang, Q.; Fu, S.H.; Chen, Y.D.; Tian, J.W.; Guo, Y. Sitagliptin, a dipeptidyl peptidase-4 inhibitor, attenuates apoptosis of vascular smooth muscle cells and reduces atherosclerosis in diabetic apolipoprotein E-deficient mice. Vasc. Pharmacol. 2021, 140, 106854. [Google Scholar] [CrossRef]
- Telikani, Z.; Sheikh, V.; Zamani, A.; Borzouei, S.; Salehi, I.; Amirzargar, M.A.; Alahgholi-Hajibehzad, M. Effects of sitagliptin and vitamin D3 on T helper cell transcription factors and cytokine production in clinical subgroups of type 2 diabetes mellitus: Highlights upregulation of FOXP3 and IL-37. Immunopharmacol. Immunotoxicol. 2019, 41, 299–311. [Google Scholar] [CrossRef]
- Pinheiro, M.M.; Stoppa, C.L.; Valduga, C.J.; Okuyama, C.E.; Gorjao, R.; Pereira, R.M.S.; Diniz, S.N. Sitagliptin inhibit human lymphocytes proliferation and Th1/Th17 differentiation in vitro. Eur. J. Pharm. Sci. 2017, 100, 17–24. [Google Scholar] [CrossRef]
- Yan, S.; Marguet, D.; Dobers, J.; Reutter, W.; Fan, H. Deficiency of CD26 results in a change of cytokine and immunoglobulin secretion after stimulation by pokeweed mitogen. Eur. J. Immunol. 2003, 33, 1519–1527. [Google Scholar] [CrossRef] [PubMed]
- Waisman, A.; Lukas, D.; Clausen, B.E.; Yogev, N. Dendritic cells as gatekeepers of tolerance. Semin. Immunopathol. 2017, 39, 153–163. [Google Scholar] [CrossRef] [PubMed]
- Møller, S.H.; Wang, L.; Ho, P.-C. Metabolic programming in dendritic cells tailors immune responses and homeostasis. Cell. Mol. Immunol. 2022, 19, 370–383. [Google Scholar] [CrossRef] [PubMed]
- Gliddon, D.R.; Howard, C.J. CD26 is expressed on a restricted subpopulation of dendritic cells in vivo. European Journal of Immunology, 2002, 32, 1472–1481. [Google Scholar] [CrossRef] [PubMed]
- Zhong, J.; Rao, X.; Deiuliis, J.; Braunstein, Z.; Narula, V.; Hazey, J.; Mikami, D.; Needleman, B.; Satoskar, A.R.; Rajagopalan, S. A potential role for dendritic cell/macrophage-expressing DPP4 in obesity-induced visceral inflammation. Diabetes 2013, 62, 149–157. [Google Scholar] [CrossRef] [PubMed]
- Schütz, F.; Hackstein, H. Identification of novel dendritic cell subset markers in human blood. Biochem. Biophys. Res. Commun. 2014, 443, 453–457. [Google Scholar] [CrossRef] [PubMed]
- Peng, H.; Guerau-de-Arellano, M.; Yang, Y.; Huss, D.J.; Papenfuss, T.L.; Lovett-Racke, A.E.; Racke, M.K. Dimethyl Fumarate Inhibits Dendritic Cell Maturation via Nuclear Factor κB (NF-κB) and Extracellular Signal-regulated Kinase 1 and 2 (ERK1/2) and Mitogen Stress-activated Kinase 1 (MSK1) Signaling. J. Biol. Chem. 2012, 287, 28017–28026. [Google Scholar] [CrossRef] [PubMed]
- Lingappan, K. NF-κB in Oxidative Stress. Curr. Opin. Toxicol. 2018, 7, 81–86. [Google Scholar] [CrossRef]
- Majeed Khan, H.; Nawaz, H.; Aerooj, H.; Sharif, A. Genotoxic and cytotoxic assessment of sitagliptin and simvastatin alone and in combination. Pak. J. Pharm. Sci. 2021, 34 (Suppl. S5), 1939–1944. [Google Scholar]
- You, F.; Li, C.; Zhang, S.; Zhang, Q.; Hu, Z.; Wang, Y.; Zhang, T.; Meng, Q.; Yu, R.; Gao, S. Sitagliptin inhibits the survival, stemness and autophagy of glioma cells, and enhances temozolomide cytotoxicity. Biomed. Pharmacother. 2023, 162, 114555. [Google Scholar] [CrossRef]
- Herman, G.A.; Stevens, C.; Van Dyck, K.; Bergman, A.; Yi, B.; De Smet, M.; Snyder, K.; Hillard, D.; Tanen, M.; Tanaka, W.; et al. Pharmacokinetics and pharmacodynamics of sitagliptin, an inhibitor of dipeptidyl peptidase IV, in healthy subjects: Results from two randomized, double-blind, placebo-controlled studies with single oral doses. Clin. Pharmacol. Ther. 2005, 78, 675–688. [Google Scholar] [CrossRef] [PubMed]
- Farag, S.S.; Abu Zaid, M.; Schwartz, J.E.; Thakrar, T.C.; Blakley, A.J.; Abonour, R.; Robertson, M.J.; Broxmeyer, H.E.; Zhang, S. Dipeptidyl Peptidase 4 Inhibition for Prophylaxis of Acute Graft-versus-Host Disease. N. Engl. J. Med. 2021, 384, 11–19. [Google Scholar] [CrossRef] [PubMed]
- Sallusto, F.; Lanzavecchia, A. Efficient presentation of soluble antigen by cultured human dendritic cells is maintained by granulocyte/macrophage colony-stimulating factor plus interleukin 4 and downregulated by tumor necrosis factor alpha. J. Exp. Med. 1994, 179, 1109–1118. [Google Scholar] [CrossRef] [PubMed]
- Cabeza-Cabrerizo, M.; Cardoso, A.; Minutti, C.M.; Pereira da Costa, M.; Reis e Sousa, C. Dendritic Cells Revisited. Annu. Rev. Immunol. 2021, 39, 131–166. [Google Scholar] [CrossRef] [PubMed]
- Bekić, M.; Vasiljević, M.; Stojanović, D.; Kokol, V.; Mihajlović, D.; Vučević, D.; Uskoković, P.; Čolić, M.; Tomić, S. Phosphonate-Modified Cellulose Nanocrystals Potentiate the Th1 Polarising Capacity of Monocyte-Derived Dendritic Cells via GABA-B Receptor. Int. J. Nanomed. 2022, 17, 3191–3216. [Google Scholar] [CrossRef] [PubMed]
- Gogolak, P.; Rethi, B.; Szatmari, I.; Lanyi, A.; Dezso, B.; Nagy, L.; Rajnavolgyi, E. Differentiation of CD1a- and CD1a+ monocyte-derived dendritic cells is biased by lipid environment and PPARγ. Blood 2007, 109, 643–652. [Google Scholar] [CrossRef] [PubMed]
- Patente, T.A.; Pinho, M.P.; Oliveira, A.A.; Evangelista, G.C.M.; Bergami-Santos, P.C.; Barbuto, J.A.M. Human Dendritic Cells: Their Heterogeneity and Clinical Application Potential in Cancer Immunotherapy. Front. Immunol. 2019, 9, 3176. [Google Scholar] [CrossRef] [PubMed]
- Zanna, M.Y.; Yasmin, A.R.; Omar, A.R.; Arshad, S.S.; Mariatulqabtiah, A.R.; Nur-Fazila, S.H.; Mahiza, M.I.N. Review of Dendritic Cells, Their Role in Clinical Immunology, and Distribution in Various Animal Species. Int. J. Mol. Sci. 2021, 22, 8044. [Google Scholar] [CrossRef]
- Han, T.H.; Jin, P.; Ren, J.; Slezak, S.; Marincola, F.M.; Stroncek, D.F. Evaluation of 3 clinical dendritic cell maturation protocols containing lipopolysaccharide and interferon-gamma. J. Immunother. 2009, 32, 399–407. [Google Scholar] [CrossRef]
- Gardner, J.K.; Cornwall, S.M.J.; Musk, A.W.; Alvarez, J.; Mamotte, C.D.S.; Jackman, C.; Nowak, A.K.; Nelson, D.J. Elderly dendritic cells respond to LPS/IFN-γ and CD40L stimulation despite incomplete maturation. PLoS ONE 2018, 13, e0195313. [Google Scholar] [CrossRef]
- Qu, C.; Brinck-Jensen, N.S.; Zang, M.; Chen, K. Monocyte-derived dendritic cells: Targets as potent antigen-presenting cells for the design of vaccines against infectious diseases. Int. J. Infect. Dis. 2014, 19, 1–5. [Google Scholar] [CrossRef]
- Fang, Y.; Yu, S.; Ellis, J.S.; Shrav, T.; Braley-Mullen, H. Comparison of sensitivity of Th1, Th2, and Th17 cells to Fas-mediated apoptosis. J. Leukoc. Biol. 2010, 87, 1019–1028. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Y.; Yao, Y.; Li, J.; Wang, Y.; Cheng, J.; Zhu, Y. Functional Dissection of CD26 and Its Pharmacological Inhibition by Sitagliptin During Skin Wound Healing. Med. Sci. Monit. 2021, 27, e928933. [Google Scholar] [CrossRef] [PubMed]
- Price, J.D.; Linder, G.; Li, W.P.; Zimmermann, B.; Rother, K.I.; Malek, R.; Alattar, M.; Tarbell, K.V. Effects of short-term sitagliptin treatment on immune parameters in healthy individuals, a randomized placebo-controlled study. Clin. Exp. Immunol. 2013, 174, 120–128. [Google Scholar] [CrossRef] [PubMed]
- Sato, Y.; Kamada, T.; Yamauchi, A. The role of dipeptidyl peptidase 4 (DPP4) in the preservation of renal function: DPP4 involvement in hemoglobin expression. J. Endocrinol. 2014, 223, 133–142. [Google Scholar] [CrossRef] [PubMed]
- Valls, P.O.; Esposito, A. Signalling dynamics, cell decisions, and homeostatic control in health and disease. Curr. Opin. Cell Biol. 2022, 75, 102066. [Google Scholar] [CrossRef] [PubMed]
- Sauvat, A.; Ciccosanti, F.; Colavita, F.; Di Rienzo, M.; Castilletti, C.; Capobianchi, M.R.; Kepp, O.; Zitvogel, L.; Fimia, G.M.; Piacentini, M.; et al. On-target versus off-target effects of drugs inhibiting the replication of SARS-CoV-2. Cell Death Dis. 2020, 11, 656. [Google Scholar] [CrossRef] [PubMed]
- Gong, Y.; Zhang, Z. Alternative signaling pathways: When, where and why? FEBS Lett. 2005, 579, 5265–5274. [Google Scholar] [CrossRef] [PubMed]
- Luo, W.; Hu, J.; Xu, W.; Dong, J. Distinct spatial and temporal roles for Th1, Th2, and Th17 cells in asthma. Front. Immunol. 2022, 13, 974066. [Google Scholar] [CrossRef]
- Brogdon, J.L.; Xu, Y.; Szabo, S.J.; An, S.; Buxton, F.; Cohen, D.; Huang, Q. Histone deacetylase activities are required for innate immune cell control of Th1 but not Th2 effector cell function. Blood 2007, 109, 1123–1130. [Google Scholar] [CrossRef]
- Lin, T.; Cai, Y.; Tang, L.; Lian, Y.; Liu, M.; Liu, C. Efficacy and safety of sitagliptin and insulin for latent autoimmune diabetes in adults: A systematic review and meta-analysis. J. Diabetes Investig. 2022, 13, 1506–1519. [Google Scholar] [CrossRef] [PubMed]
- Mahabadi-Ashtiyani, E.; Sheikh, V.; Borzouei, S.; Salehi, I.; Alahgholi-Hajibehzad, M. The increased T helper cells proliferation and inflammatory responses in patients with type 2 diabetes mellitus is suppressed by sitagliptin and vitamin D3 in vitro. Inflamm. Res. 2019, 68, 857–866. [Google Scholar] [CrossRef] [PubMed]
- Leung, S.; Liu, X.; Fang, L.; Chen, X.; Guo, T.; Zhang, J. The cytokine milieu in the interplay of pathogenic Th1/Th17 cells and regulatory T cells in autoimmune disease. Cell. Mol. Immunol. 2010, 7, 182–189. [Google Scholar] [CrossRef] [PubMed]
- Zhuang, Q.; Cai, H.; Cao, Q.; Li, Z.; Liu, S.; Ming, Y. Tolerogenic Dendritic Cells: The Pearl of Immunotherapy in Organ Transplantation. Front. Immunol. 2020, 11, 552988. [Google Scholar] [CrossRef] [PubMed]
- Morante-Palacios, O.; Fondelli, F.; Ballestar, E.; Martinez-Cáceres, E.M. Tolerogenic Dendritic Cells in Autoimmunity and Inflammatory Diseases. Trends Immunol. 2021, 42, 59–75. [Google Scholar] [CrossRef] [PubMed]
- Gao, A.; Sun, Y.; Peng, G. ILT4 functions as a potential checkpoint molecule for tumor immunotherapy. Biochim. Biophys. Acta Rev. Cancer 2018, 1869, 278–285. [Google Scholar] [CrossRef] [PubMed]
- Sharma, M.D.; Pacholczyk, R.; Shi, H.; Berrong, Z.J.; Zakharia, Y.; Greco, A.; Chang, C.S.; Eathiraj, S.; Kennedy, E.; Cash, T.; et al. Inhibition of the BTK-IDO-mTOR axis promotes differentiation of monocyte-lineage dendritic cells and enhances anti-tumor T cell immunity. Immunity 2021, 54, 2354–2371. [Google Scholar] [CrossRef] [PubMed]
- Gargaro, M.; Scalisi, G.; Manni, G.; Briseno, C.G.; Bagadia, P.; Durai, V.; Theisen, D.J.; Kim, S.; Castelli, M.; Xu, C.A.; et al. Indoleamine 2,3-dioxygenase 1 activation in mature cDC1 promotes tolerogenic education of inflammatory cDC2 via metabolic communication. Immunity 2022, 55, 1032–1050. [Google Scholar] [CrossRef]
- Brenk, M.; Scheler, M.; Koch, S.; Neumann, J.; Takikawa, O.; Häcker, G.; Bieber, T.; von Bubnoff, D. Tryptophan Deprivation Induces Inhibitory Receptors ILT3 and ILT4 on Dendritic Cells Favoring the Induction of Human CD4+CD25+ Foxp3+ T Regulatory Cells1. J. Immunol. 2009, 183, 145–154. [Google Scholar] [CrossRef]
- Underwood, D.C.; Osborn, R.R.; Bochnowicz, S.; Webb, E.F.; Rieman, D.J.; Lee, J.C.; Romanic, A.M.; Adams, J.L.; Hay, D.W.; Griswold, D.E. SB 239063, a p38 MAPK inhibitor, reduces neutrophilia, inflammatory cytokines, MMP-9, and fibrosis in lung. Am. J. Physiol. Lung Cell. Mol. Physiol. 2000, 279, L895–L902. [Google Scholar] [CrossRef]
- Lu, Y.; Zhang, M.; Wang, S.; Hong, B.; Wang, Z.; Li, H.; Zheng, Y.; Yang, J.; Davis, R.E.; Qian, J.; et al. p38 MAPK-inhibited dendritic cells induce superior antitumour immune responses and overcome regulatory T-cell-mediated immunosuppression. Nat. Commun. 2014, 5, 4229. [Google Scholar] [CrossRef] [PubMed]
- Ade, N.; Antonios, D.; Kerdine-Romer, S.; Boisleve, F.; Rousset, F.; Pallardy, M. NF-kappaB plays a major role in the maturation of human dendritic cells induced by NiSO4 but not by DNCB. Toxicol. Sci. 2007, 99, 488–501. [Google Scholar] [CrossRef] [PubMed]
- Hasegawa, H.; Nakamura, Y.; Tsuji, M.; Ono, R.; Oguchi, T.; Oguchi, K.; Kiuchi, Y.; Ohsawa, I.; Gotoh, H.; Goto, Y.; et al. Sitagliptin Inhibits the Lipopolysaccharide-Induced Inflammation. J. Pharm. Drug Deliv. Res. 2016, 5, 2. [Google Scholar] [CrossRef]
- Nakahara, T.; Moroi, Y.; Uchi, H.; Furue, M. Differential role of MAPK signaling in human dendritic cell maturation and Th1/Th2 engagement. J. Dermatol. Sci. 2006, 42, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Shi, Q.; Cheng, L.; Liu, Z.; Hu, K.; Ran, J.; Ge, D.; Fu, J. The p38 MAPK inhibitor SB203580 differentially modulates LPS-induced interleukin 6 expression in macrophages. Cent. Eur. J. Immunol. 2015, 40, 276–282. [Google Scholar] [CrossRef] [PubMed]
- Ohnuma, K.; Yamochi, T.; Uchiyama, M.; Nishibashi, K.; Yoshikawa, N.; Shimizu, N.; Iwata, S.; Tanaka, H.; Dang, N.H.; Morimoto, C. CD26 up-regulates expression of CD86 on antigen-presenting cells by means of caveolin-1. Proc. Natl. Acad. Sci. USA 2004, 101, 14186–14191. [Google Scholar] [CrossRef] [PubMed]
- Ohnuma, K.; Uchiyama, M.; Yamochi, T.; Nishibashi, K.; Hosono, O.; Takahashi, N.; Kina, S.; Tanaka, H.; Lin, X.; Dang, N.H.; et al. Caveolin-1 triggers T-cell activation via CD26 in association with CARMA1. J. Biol. Chem. 2007, 282, 10117–10131. [Google Scholar] [CrossRef] [PubMed]
- Busek, P.; Duke-Cohan, J.S.; Sedo, A. Does DPP-IV Inhibition Offer New Avenues for Therapeutic Intervention in Malignant Disease? Cancers 2022, 14, 2072. [Google Scholar] [CrossRef]
- Herholz, M.; Cepeda, E.; Baumann, L.; Kukat, A.; Hermeling, J.; Maciej, S.; Szczepanowska, K.; Pavlenko, V.; Frommolt, P.; Trifunovic, A. KLF-1 orchestrates a xenobiotic detoxification program essential for longevity of mitochondrial mutants. Nat. Commun. 2019, 10, 3323. [Google Scholar] [CrossRef]












Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Drakul, M.; Tomić, S.; Bekić, M.; Mihajlović, D.; Vasiljević, M.; Rakočević, S.; Đokić, J.; Popović, N.; Bokonjić, D.; Čolić, M. Sitagliptin Induces Tolerogenic Human Dendritic Cells. Int. J. Mol. Sci. 2023, 24, 16829. https://doi.org/10.3390/ijms242316829
Drakul M, Tomić S, Bekić M, Mihajlović D, Vasiljević M, Rakočević S, Đokić J, Popović N, Bokonjić D, Čolić M. Sitagliptin Induces Tolerogenic Human Dendritic Cells. International Journal of Molecular Sciences. 2023; 24(23):16829. https://doi.org/10.3390/ijms242316829
Chicago/Turabian StyleDrakul, Marija, Sergej Tomić, Marina Bekić, Dušan Mihajlović, Miloš Vasiljević, Sara Rakočević, Jelena Đokić, Nikola Popović, Dejan Bokonjić, and Miodrag Čolić. 2023. "Sitagliptin Induces Tolerogenic Human Dendritic Cells" International Journal of Molecular Sciences 24, no. 23: 16829. https://doi.org/10.3390/ijms242316829