

Quantum Biology and the Potential Role of Entanglement and Tunneling in Non-Targeted Effects of Ionizing Radiation: A Review and Proposed Model

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction

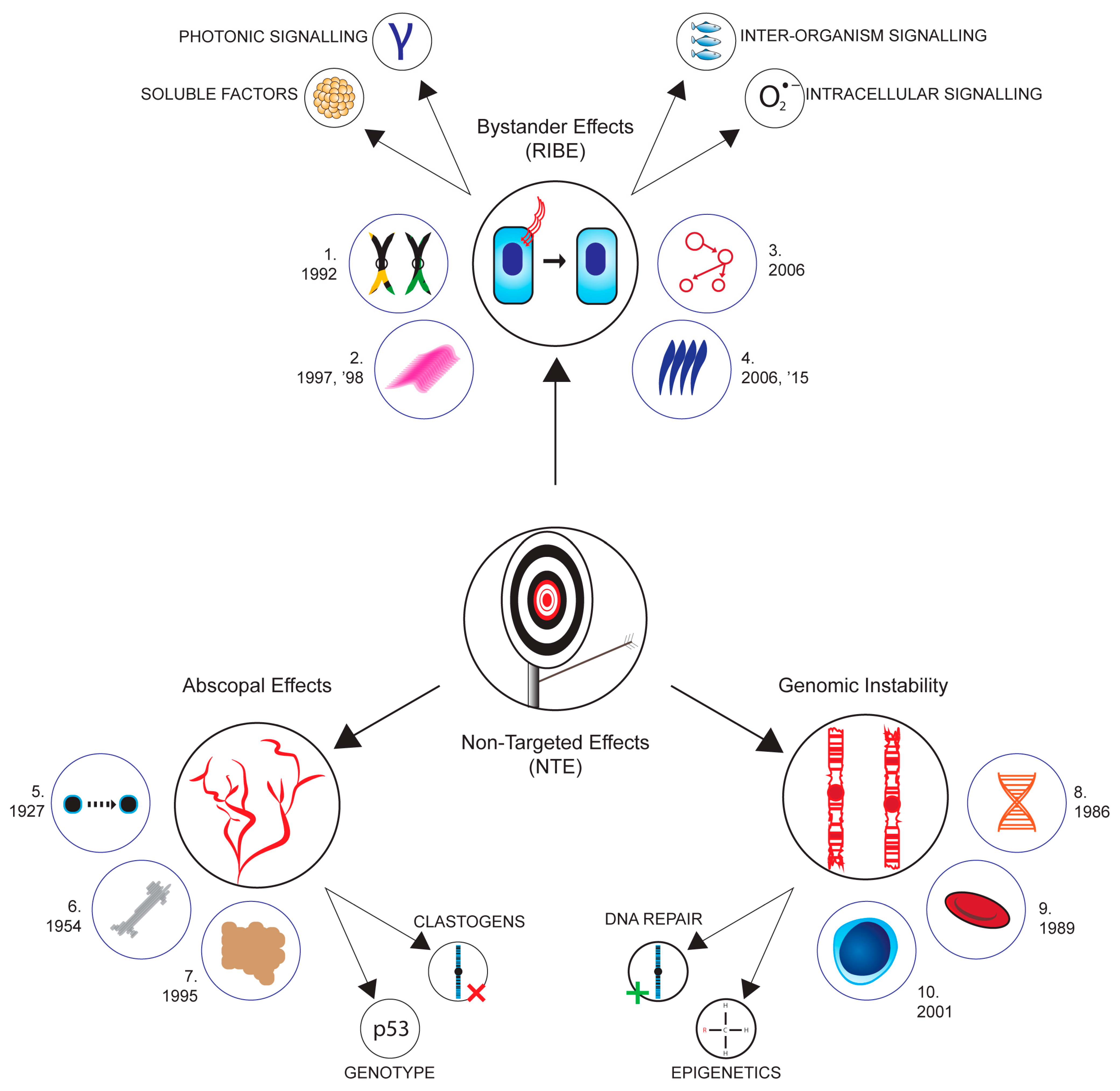
2. Non-Targeted Effects (NTE) Gaps in Our Knowledge
3. Quantum Biology and NTE
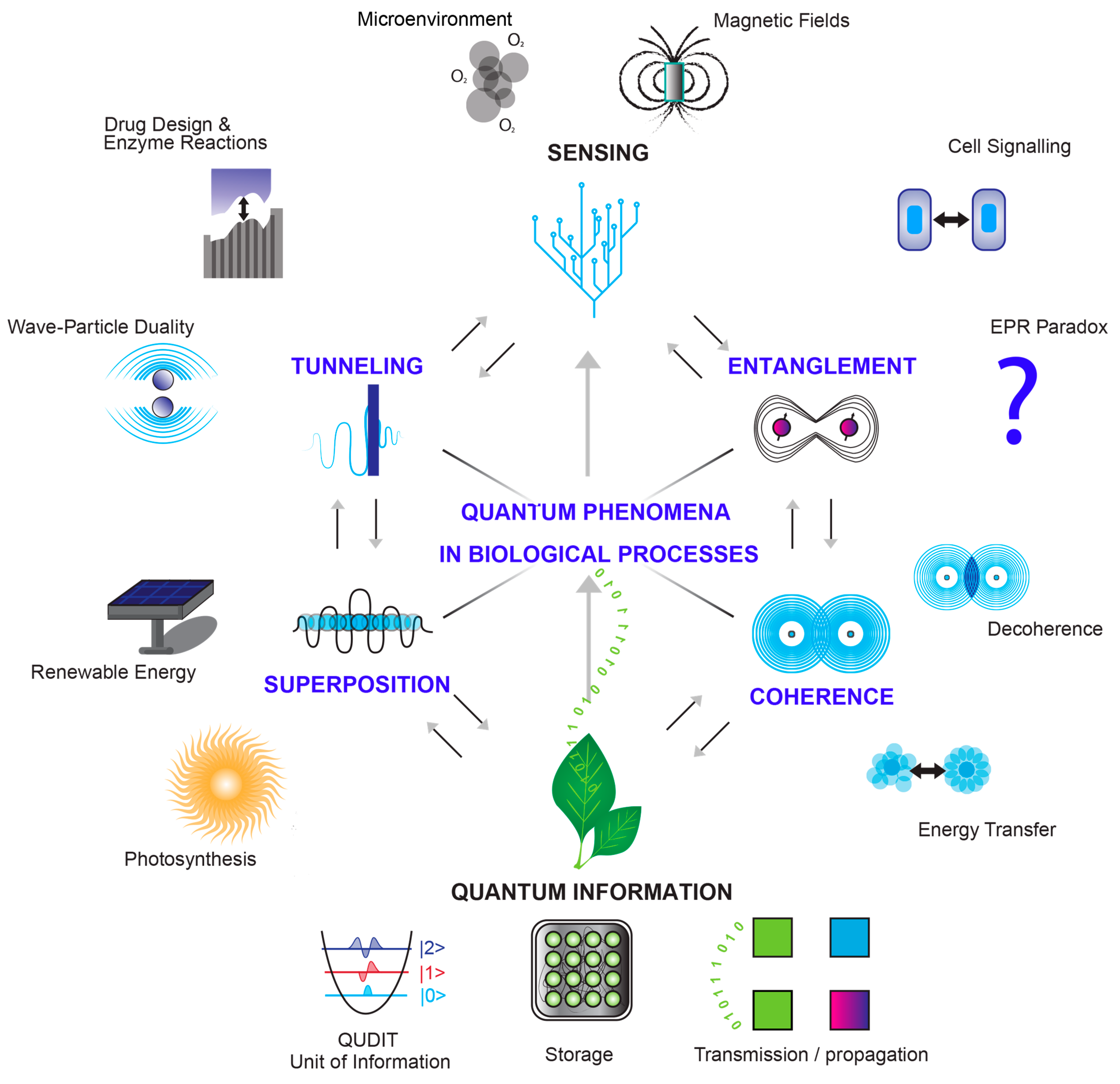
3.1. Quantum Physics Meets Biology
- Quantum superposition
- Quantum Entanglement
- Quantum tunneling
- Quantum coherence
- Quantum sensing
- Quantum information in quantum biology and radiation effects
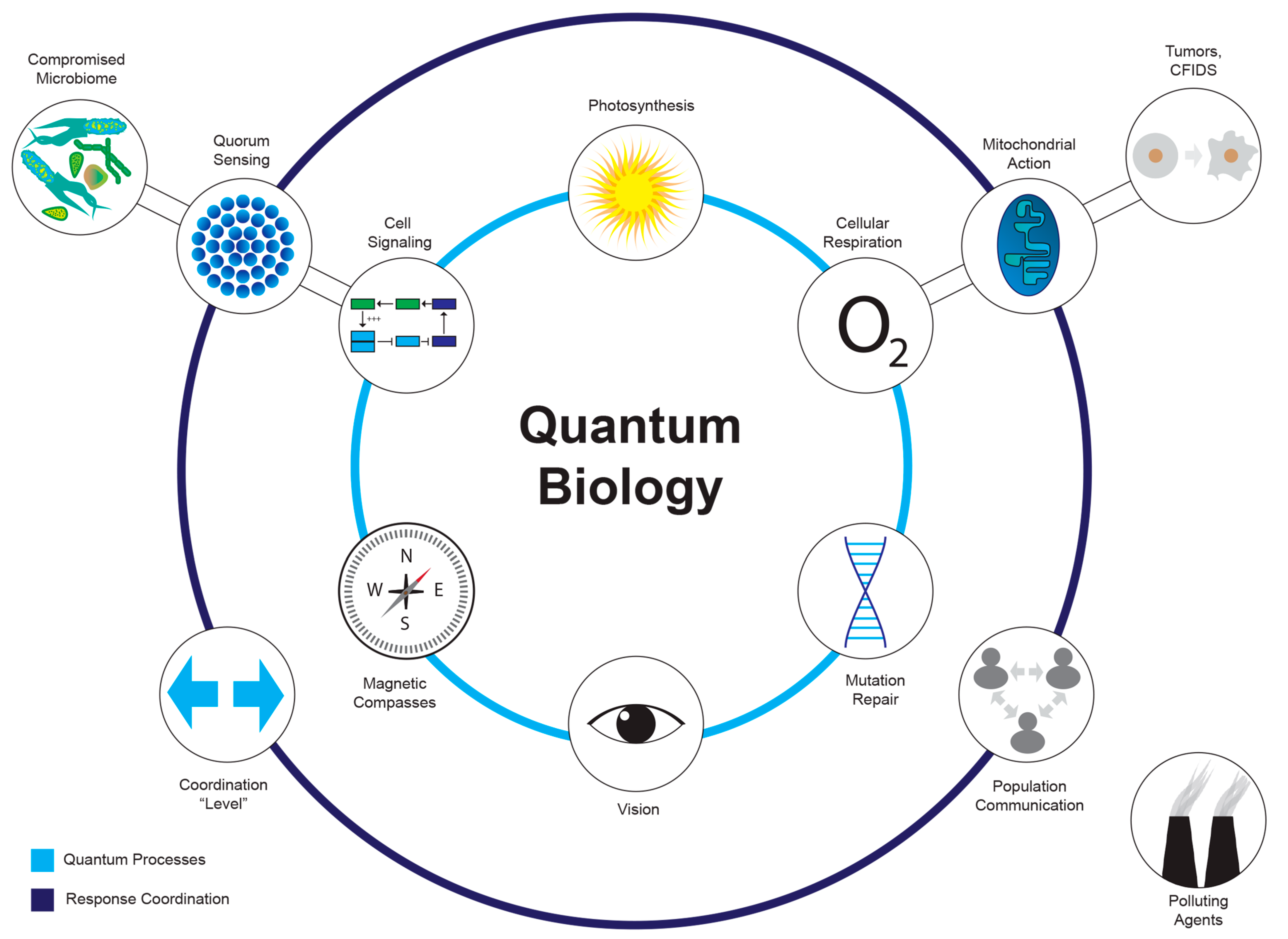
3.2. Areas Where Quantum Effects May Occur
- Quantum Effects on Morphogenetic and Bioelectric Fields
- 2.
- Quantum Effects on Mitogenic Radiation
- Photosynthesis, vision, and magnetic compasses
- Cellular Respiration
- Mutation/repair in DNA
- Cell signaling after energy capture/deposition
3.3. Anywhere Response Is Coordinated
- Mitochondrial action
- Tissue/organ/organism-level response
- Quorum sensing
- Population-level communication (bacteria/social animals)
3.4. When Quantum Behavior Goes Wrong
3.4.1. Influences on Quantum Behavior: A Glimpse into Dynamic Responses
- Temperature Variations
- Electromagnetic Field Fluctuations
- Chemical Interactions
- Radiation Exposure
3.4.2. Low-Dose Mechanisms
3.4.3. Quantum Effects in Radiation
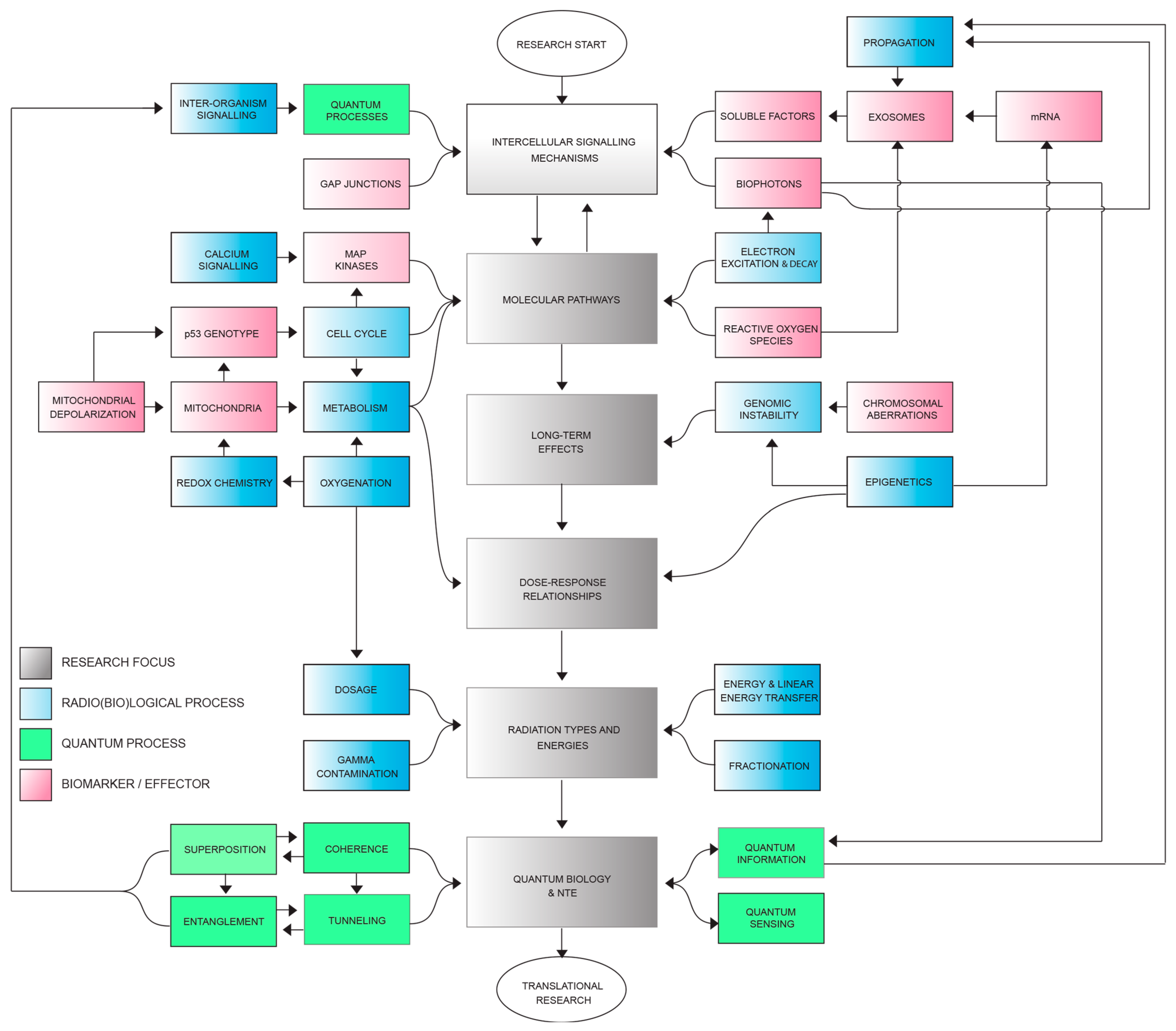
4. Proposed Model and Discussion
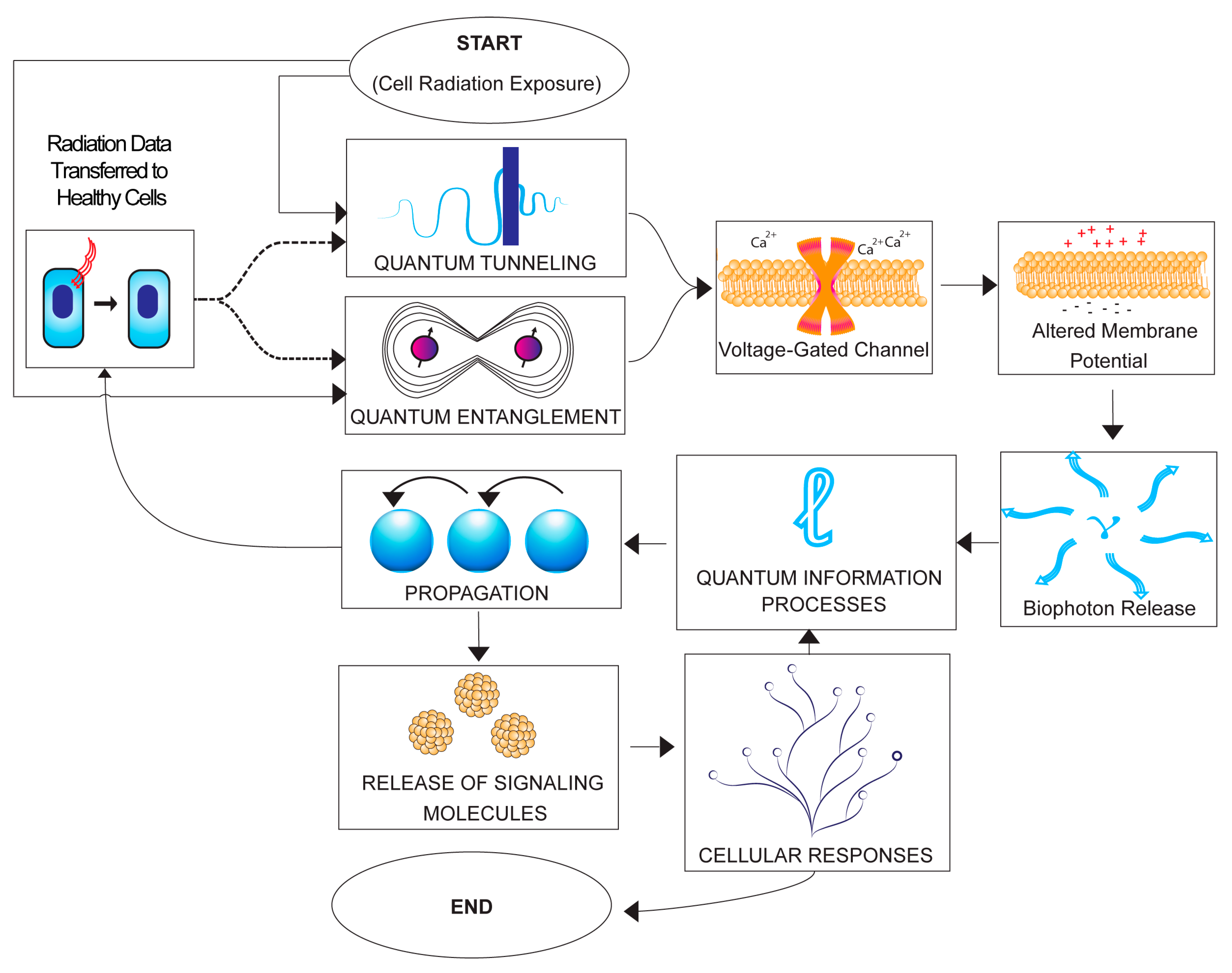
4.1. Hypothetical Sequence of Quantum Events in Radiobiology
- Ionizing Radiation Exposure: Ionizing radiation triggers quantum phenomena, including quantum tunneling and entanglement, within irradiated cells, which involve electron transfer and energy state transitions (e.g., reactive oxygen species (ROS); chirality and CISS effect; bioelectric field alteration).
- Cellular Impact: Quantum processes influenced by radiation-induced quantum phenomena affect the activation of voltage-gated ion channels within irradiated cells, which leads to changes in cellular membrane potentials, creating an altered electrochemical environment.
- Photon Emission: Altered membrane potentials in irradiated cells can result in the emission of photons, potentially carrying encoded quantum information that propagates through the cellular microenvironment.
- Quantum Information Transmission: The quantum information encoded in emitted photons is transmitted to neighboring non-irradiated cells through non-targeted effects, facilitated by signaling molecules or physical interactions between cells.
- Quantum Events in Non-Targeted Cells: Non-targeted cells that receive quantum-encoded information may undergo quantum events similar to those in irradiated cells. These events can influence the opening and closing of voltage-gated ion channels, further impacting cellular membrane potentials [318,326]. The precision and specificity of the quantum-encoded information within emitted photons, particularly those involved in CISS, can enhance these events [327].
- Signaling Molecule Release: The alteration of membrane potentials in non-targeted cells leads to the release of signaling molecules contained within exosomes. These signaling molecules, enriched by the quantum properties of ROS and the quantum-encoded information, activate various cellular responses in neighboring non-irradiated cells. These responses encompass the activation of cellular pathways, changes in gene expression, and modifications in cellular behavior, contributing to the observed non-targeted effects (NTE) in radiobiology.
- Cellular Responses: Released signaling molecules induce cellular responses in neighboring non-irradiated cells, activating cellular pathways, altering gene expression, or modifying cellular behavior, which contribute to the observed non-targeted effects (NTE) in radiobiology.
4.2. Quantum Tunneling in the Activation of Voltage-Gated Ion Channels
4.3. Quantum Entanglement in the Release of Signaling Molecules
4.4. Quantum Information Transmission and Cellular Impact
4.5. Quantum Coherence in Cellular Signaling
4.6. Supporting Evidence
4.7. Expanding the Model to Explore Factors Shaping Non-Thermal Effects of Radiation
4.8. Quantum Concepts in Radiobiology and Quantum Computing
4.9. Future Research and Experimental Directions
5. Conclusions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Mothersill, C.; Rusin, A.; Seymour, C. The Development of Bio-Assays Based on Non-Targeted Effects of Radiation; a Potential Worm-Hole into Ecosystem Level Biomarkers. In NATO Science for Peace and Security Series A: Chemistry and Biology; Springer: Berlin/Heidelberg, Germany, 2022. [Google Scholar]
- Seiler, A.; Fagundes, C.P.; Christian, L.M. The Impact of Everyday Stressors on the Immune System and Health. In Stress Challenges and Immunity in Space: From Mechanisms to Monitoring and Preventive Strategies; Springer: Berlin/Heidelberg, Germany, 2019. [Google Scholar]
- Godoy, L.D.; Rossignoli, M.T.; Delfino-Pereira, P.; Garcia-Cairasco, N.; de Lima Umeoka, E.H. A Comprehensive Overview on Stress Neurobiology: Basic Concepts and Clinical Implications. Front. Behav. Neurosci. 2018, 12, 127. [Google Scholar]
- Kadhim, M.A.; Hill, M.A. Non-Targeted Effects of Radiation Exposure: Recent Advances and Implications. Radiat. Prot. Dosimetry 2015, 166, 118–124. [Google Scholar] [CrossRef] [PubMed]
- Mothersill, C.; Rusin, A.; Seymour, C. Relevance of Non-Targeted Effects for Radiotherapy and Diagnostic Radiology; A Historical and Conceptual Analysis of Key Players. Cancers 2019, 11, 1236. [Google Scholar] [CrossRef] [PubMed]
- Han, X.; Chen, Y.; Zhang, N.; Huang, C.; He, G.; Li, T.; Wei, M.; Song, Q.; Mo, S.; Lv, Y. Single-Cell Mechanistic Studies of Radiation-Mediated Bystander Effects. Front. Immunol. 2022, 13, 849341. [Google Scholar] [CrossRef] [PubMed]
- Tang, H.; Cai, L.; He, X.; Niu, Z.; Huang, H.; Hu, W.; Bian, H.; Huang, H. Radiation-Induced Bystander Effect and Its Clinical Implications. Front. Oncol. 2023, 13, 1124412. [Google Scholar] [CrossRef] [PubMed]
- Calabrese, E.J.; Kozumbo, W.J. The Hormetic Dose-Response Mechanism: Nrf2 Activation. Pharmacol. Res. 2021, 167, 105526. [Google Scholar]
- Hauptmann, M.; Daniels, R.D.; Cardis, E.; Cullings, H.M.; Kendall, G.; Laurier, D.; Linet, M.S.; Little, M.P.; Lubin, J.H.; Preston, D.L.; et al. Epidemiological Studies of Low-Dose Ionizing Radiation and Cancer: Summary Bias Assessment and Meta-Analysis. J. Natl. Cancer Inst. Monogr. 2020, 2020, 188–200. [Google Scholar] [CrossRef]
- Hall, W.A.; Paulson, E.; Li, X.A.; Erickson, B.; Schultz, C.; Tree, A.; Awan, M.; Low, D.A.; McDonald, B.A.; Salzillo, T.; et al. Magnetic Resonance Linear Accelerator Technology and Adaptive Radiation Therapy: An Overview for Clinicians. CA Cancer J. Clin. 2022, 72, 34–56. [Google Scholar] [CrossRef] [PubMed]
- Connell, P.P.; Hellman, S. Advances in Radiotherapy and Implications for the next Century: A Historical Perspective. Cancer Res. 2009, 69, 383–392. [Google Scholar] [PubMed]
- Matarèse, B.F.E.; Lad, J.; Seymour, C.; Schofield, P.N.; Mothersill, C. Bio-Acoustic Signaling; Exploring the Potential of Sound as a Mediator of Low-Dose Radiation and Stress Responses in the Environment. Int. J. Radiat. Biol. 2020, 98, 1083–1097. [Google Scholar] [CrossRef]
- Kadhim, M.; Salomaa, S.; Wright, E.; Hildebrandt, G.; Belyakov, O.V.; Prise, K.M.; Little, M.P. Non-Targeted Effects of Ionising Radiation-Implications for Low Dose Risk. Mutat. Res. Rev. Mutat. Res. 2013, 752, 84–98. [Google Scholar]
- Yahyapour, R.; Salajegheh, A.; Safari, A.; Amini, P.; Rezaey-An, A.; Amraee, A.; Najafi, M. Radiation-Induced Non-Targeted Effect and Carcinogenesis; Implications in Clinical Radiotherapy. J. Biomed. Phys. Eng. 2018, 8, 435–446. [Google Scholar] [CrossRef]
- Laurier, D.; Rühm, W.; Paquet, F.; Applegate, K.; Cool, D.; Clement, C. Areas of Research to Support the System of Radiological Protection. Radiat. Environ. Biophys. 2021, 60, 519–530. [Google Scholar] [CrossRef] [PubMed]
- Morgan, W.F.; Sowa, M.B. Non-Targeted Effects Induced by Ionizing Radiation: Mechanisms and Potential Impact on Radiation Induced Health Effects. Cancer Lett. 2015, 356, 17–21. [Google Scholar] [CrossRef] [PubMed]
- Morgan, W.F. Non-Targeted and Delayed Effects of Exposure to Ionizing Radiation: I. Radiation-Induced Genomic Instability and Bystander Effects In Vitro. Radiat. Res. 2012, 178, AV223–AV236. [Google Scholar] [CrossRef] [PubMed]
- Morgan, W.F. Non-Targeted and Delayed Effects of Exposure to Ionizing Radiation: II. Radiation-Induced Genomic Instability and Bystander Effects in Vivo, Clastogenic Factors and Transgenerational Effects. Radiat. Res. 2003, 159, 581–596. [Google Scholar] [CrossRef]
- Liu, Y.J.; Wang, C. A Review of the Regulatory Mechanisms of Extracellular Vesicles-Mediated Intercellular Communication. Cell Commun. Signal. 2023, 21, 1–12. [Google Scholar]
- Cagatay, S.T.; Mayah, A.; Mancuso, M.; Giardullo, P.; Pazzaglia, S.; Saran, A.; Daniel, A.; Traynor, D.; Meade, A.D.; Lyng, F.; et al. Phenotypic and Functional Characteristics of Exosomes Derived from Irradiated Mouse Organs and Their Role in the Mechanisms Driving Non-Targeted Effects. Int. J. Mol. Sci. 2020, 21, 8389. [Google Scholar] [CrossRef]
- López-Díaz FJ Cross-Talk between TGF-β and P53 Regulates the Stress Response. Cancer Discov. 2013, 3, 715. [CrossRef]
- Averbeck, D. Low-Dose Non-Targeted Effects and Mitochondrial Control. Int. J. Mol. Sci. 2023, 24, 11460. [Google Scholar]
- Rouleau, N.; Karbowski, L.M.; Persinger, M.A. Experimental Evidence of Classical Conditioning and Microscopic Engrams in an Electroconductive Material. PLoS ONE 2016, 11, e0165269. [Google Scholar] [CrossRef]
- Jones, G.A.; Bradshaw, D.S. Resonance Energy Transfer: From Fundamental Theory to Recent Applications. Front. Phys. 2019, 7, 100. [Google Scholar]
- Rohli, R.V.; Li, C. Energy Transfer and Electromagnetic Radiation. In Meteorology for Coastal Scientists; Springer: Berlin/Heidelberg, Germany, 2021. [Google Scholar]
- Chelkowski, S.; Bandrauk, A.D. Control of Molecular Vibrational Excitation and Dissociation by Chirped Intense Infrared Laser Pulses. Rotational Effects. J. Chem. Phys. 1993, 99, 4279–4287. [Google Scholar] [CrossRef]
- Phelps, A.V. Rotational and Vibrational Excitation of Molecules by Low-Energy Electrons. Rev. Mod. Phys. 1968, 40, 399–410. [Google Scholar] [CrossRef]
- Krisko, A.; Radman, M. Phenotypic and Genetic Consequences of Protein Damage. PLoS Genet. 2013, 9, e1003810. [Google Scholar] [CrossRef]
- Reynaud, E. Protein Misfolding and Degenerative Diseases. Nat. Educ. 2010, 3, 28. [Google Scholar]
- Harris, M.P. Bioelectric Signaling as a Unique Regulator of Development and Regeneration. Development 2021, 148, dev180794. [Google Scholar]
- Funk, R.H.W. Endogenous Electric Fields as Guiding Cue for Cell Migration. Front Physiol. 2015, 6, 143. [Google Scholar]
- Kim, Y.; Bertagna, F.; D’souza, E.M.; Heyes, D.J.; Johannissen, L.O.; Nery, E.T.; Pantelias, A.; Jimenez, A.S.P.; Slocombe, L.; Spencer, M.G.; et al. Quantum Biology: An Update and Perspective. Quantum Rep. 2021, 3, 80–126. [Google Scholar]
- Popp, F.-A. Biophotons—Background, Experimental Results, Theoretical Approach and Applications. In Integrative Biophysics; Springer: Berlin/Heidelberg, Germany, 2003. [Google Scholar]
- Bischof, M. A Tribute to Fritz-Albert Popp on the Occasion of His 70th Birthday. Indian J. Exp. Biol. 2008, 46, 267–272. [Google Scholar]
- Popp, F.A.; Li, K.H.; Gu, Q. Recent Advances in Biophoton Research and Its Applications; World Scientific Publishing Co. Pte. Ltd.: London, UK, 1992. [Google Scholar]
- Popp, F.-A.; Gu, Q.; Li, K.-H. Biophoton Emission: Experimental Background and Theoretical Approaches. Mod. Phys. Lett. B 1994, 8, 1269–1296. [Google Scholar] [CrossRef]
- Popp, F.A.; Chang, J.J.; Herzog, A.; Yan, Z.; Yan, Y. Evidence of Non-Classical (Squeezed) Light in Biological Systems. Phys. Lett. Sect. A Gen. At. Solid State Phys. 2002, 293, 98–102. [Google Scholar] [CrossRef]
- Liu, J.; van Iersel, M.W. Photosynthetic Physiology of Blue, Green, and Red Light: Light Intensity Effects and Underlying Mechanisms. Front Plant Sci. 2021, 12, 619987. [Google Scholar] [CrossRef] [PubMed]
- Nagasawa, H.; Little, J.B. Induction of sister chromatid exchanges by extremely low doses of α-particles. Cancer Res. 1992, 52, 6394–6396. [Google Scholar] [PubMed]
- Mothersill, C.; Seymour, C. Medium from irradiated human epithelial cells but not human fibroblasts reduces the clonogenic survival of unirradiated cells. Int. J. Radiat. Biol. 1997, 71, 421–427. [Google Scholar]
- Mothersill, C.; Seymour, C.B. Cell-cell contact during gamma irradiation is not required to induce a bystander effect in normal human keratinocytes: Evidence for release during irradiation of a signal controlling survival into the medium. Radiat. Res. 1998, 149, 256–262. [Google Scholar] [CrossRef]
- Lyng, F.M.; Maguire, P.; McClean, B.; Seymour, C.; Mothersill, C. The involvement of calcium and MAP kinase signaling pathways in the production of radiation-induced bystander effects. Radiat. Res. 2006, 165, 400–409. [Google Scholar] [CrossRef] [PubMed]
- Mosse, I.; Marozik, P.; Seymour, C.; Mothersill, C. The effect of melanin on the bystander effect in human keratinocytes. Mutat. Res.-Fundam. Mol. Mech. Mutagen. 2006, 597, 133–137. [Google Scholar] [CrossRef] [PubMed]
- Le, M.; McNeill, F.E.; Seymour, C.; Rainbow, A.J.; Mothersill, C.E. An observed effect of ultraviolet radiation emitted from beta-irradiated HaCaT cells upon non-beta-irradiated bystander cells. Radiat. Res. 2015, 183, 279–290. [Google Scholar] [CrossRef]
- Le, M.; Mothersill, C.E.; Seymour, C.B.; Ahmad, S.B.; Armstrong, A.; Rainbow, A.J.; McNeill, F.E. Factors affecting ultraviolet-A photon emission from beta-irradiated human keratinocyte cells. Phys. Med. Biol. 2015, 60, 6371–6389. [Google Scholar] [CrossRef]
- Strangeway, T.S.P.; Fell, H.B. A Study of the Direct and Indirect Action of X-Rays upon the Tissues of the Embryonic Fowl. Proceedings of the Royal Society of London. Series B, Containing Papers of a Biological Character 1927, 102, 9–29. [Google Scholar]
- Parsons, W.B.; Watkins, C.H.; Pease, G.L.; Childs, D.S. Changes in sternal marrow following roentgen-ray therapy to the spleen in chronic granulocytic leukemia. Cancer 1954, 7, 179–189. [Google Scholar] [CrossRef] [PubMed]
- Morgan, G.W.; Pharm, B.; Breit, S.N. Radiation and the lung: A reevaluation of the mechanisms mediating pulmonary injury. Int. J. Radiat. Oncol.*Biol.*Phys. 1995 31, 361–369. [CrossRef]
- Seymour, C.B.; Mothersill, C.; Alper, T. High yields of lethal mutations in somatic mammalian-cells that survive ionizing-radiation. Br. J. Radiol. 1986, 59, 1138. [Google Scholar] [CrossRef]
- Pampfer, S.; Streffer, C. Increased chromosome aberration levels in cells from mouse fetuses after zygote X-irradiation. Int. J. Radiat. Biol. 1989, 55, 85–92. [Google Scholar] [CrossRef] [PubMed]
- Coen, N.; Mothersill, C.; Kadhim, M.; Wright, E.G. Heavy metals of relevance to human health induce genomic instability. J. Pathol. 2001, 195, 293–299. [Google Scholar] [CrossRef]
- Marais, A.; Adams, B.; Ringsmuth, A.K.; Ferretti, M.; Gruber, J.M.; Hendrikx, R.; Schuld, M.; Smith, S.L.; Sinayskiy, I.; Krüger, T.P.J.; et al. The Future of Quantum Biology. J. R. Soc. Interface 2018, 15, 20180640. [Google Scholar] [CrossRef] [PubMed]
- Vepsäläinen, A.P.; Karamlou, A.H.; Orrell, J.L.; Dogra, A.S.; Loer, B.; Vasconcelos, F.; Kim, D.K.; Melville, A.J.; Niedzielski, B.M.; Yoder, J.L.; et al. Impact of Ionizing Radiation on Superconducting Qubit Coherence. Nature 2020, 584, 551–556. [Google Scholar] [CrossRef] [PubMed]
- Lambert, N.; Chen, Y.-N.; Cheng, Y.-C.; Li, C.-M.; Chen, G.-Y.; Nori, F. Quantum Biology. Nat. Phys. 2013, 9, 10–18. [Google Scholar] [CrossRef]
- Cao, J.; Cogdell, R.J.; Coker, D.F.; Duan, H.G.; Hauer, J.; Kleinekathöfer, U.; Jansen, T.L.C.; Mančal, T.; Dwayne Miller, R.J.; Ogilvie, J.P.; et al. Quantum Biology Revisited. Sci. Adv. 2020, 6, eaaz4888. [Google Scholar] [CrossRef]
- Streltsov, A.; Singh, U.; Dhar, H.S.; Bera, M.N.; Adesso, G. Measuring Quantum Coherence with Entanglement. Phys. Rev. Lett. 2015, 115, 020403. [Google Scholar] [CrossRef]
- Mazzoccoli, G. Chronobiology Meets Quantum Biology: A New Paradigm Overlooking the Horizon? Front Physiol. 2022, 13, 892582. [Google Scholar]
- Sharifi-Rad, M.; Anil Kumar, N.V.; Zucca, P.; Varoni, E.M.; Dini, L.; Panzarini, E.; Rajkovic, J.; Tsouh Fokou, P.V.; Azzini, E.; Peluso, I.; et al. Lifestyle, Oxidative Stress, and Antioxidants: Back and Forth in the Pathophysiology of Chronic Diseases. Front Physiol. 2020, 11, 694. [Google Scholar]
- Kolb, H.; Kempf, K.; Röhling, M.; Martin, S. Insulin: Too Much of a Good Thing Is Bad. BMC Med. 2020, 18, 1–12. [Google Scholar]
- Sarkar, A.; Yoo, J.Y.; Dutra, S.V.O.; Morgan, K.H.; Groer, M. The Association between Early-Life Gut Microbiota and Long-Term Health and Diseases. J. Clin. Med. 2021, 10, 459. [Google Scholar] [CrossRef] [PubMed]
- Belyaev, I.; Blackman, C.; Chamberlin, K.; DeSalles, A.; Dasdag, S.; Fernández, C.; Hardell, L.; Héroux, P.; Kelley, E.; Kesari, K.; et al. Scientific Evidence Invalidates Health Assumptions Underlying the FCC and ICNIRP Exposure Limit Determinations for Radiofrequency Radiation: Implications for 5G. Environ. Health 2022, 21, 92. [Google Scholar] [CrossRef]
- Giuranno, L.; Ient, J.; De Ruysscher, D.; Vooijs, M.A. Radiation-Induced Lung Injury (RILI). Front Oncol. 2019, 9, 877. [Google Scholar] [PubMed]
- Li, L.; Li, D. Inter-Individual Variability and Non-Linear Dose-Response Relationship in Assessing Human Health Impact from Chemicals in LCA: Addressing Uncertainties in Exposure and Toxicological Susceptibility. Front. Sustain. 2021, 2, 648138. [Google Scholar] [CrossRef]
- Orsini, N.; Li, R.; Wolk, A.; Khudyakov, P.; Spiegelman, D. Meta-Analysis for Linear and Nonlinear Dose-Response Relations: Examples, an Evaluation of Approximations, and Software. Am. J. Epidemiol. 2012, 175, 66–73. [Google Scholar] [CrossRef]
- Haussmann, J.; Corradini, S.; Nestle-Kraemling, C.; Bölke, E.; Njanang, F.J.D.; Tamaskovics, B.; Orth, K.; Ruckhaeberle, E.; Fehm, T.; Mohrmann, S.; et al. Recent Advances in Radiotherapy of Breast Cancer. Radiat. Oncol. 2020, 15, 71. [Google Scholar] [CrossRef]
- Fukunaga, H.; Prise, K.M. Non-Uniform Radiation-Induced Biological Responses at the Tissue Level Involved in the Health Risk of Environmental Radiation: A Radiobiological Hypothesis. Environ. Health 2018, 17, 93. [Google Scholar] [CrossRef] [PubMed]
- Di Maggio, F.M.; Minafra, L.; Forte, G.I.; Cammarata, F.P.; Lio, D.; Messa, C.; Gilardi, M.C.; Bravatà, V. Portrait of Inflammatory Response to Ionizing Radiation Treatment. J. Inflamm. 2015, 12, 14. [Google Scholar] [CrossRef]
- Gan, B.; Liu, S.; He, Z.; Chen, F.; Niu, H.; Cheng, J.; Tan, B.; Yu, B. Research Progress of Metal-Based Shielding Materials for Neutron and Gamma Rays. Acta Metall. Sin. (Engl. Lett.) 2021, 34, 1609–1617. [Google Scholar] [CrossRef]
- Seth, I.; Schwartz, J.L.; Stewart, R.D.; Emery, R.; Joiner, M.C.; Tucker, J.D. Neutron Exposures in Human Cells: Bystander Effect and Relative Biological Effectiveness. PLoS ONE 2014, 9, e98947. [Google Scholar] [CrossRef] [PubMed]
- Khan, A.J.; Li, X.; Haines, D.K.; Hoffman, T.J.; Semkow, T.M. Investigation of Neutron Shielding Materials for Low-Background Gamma Spectrometry. J. Radioanal. Nucl. Chem. 2021, 328, 941–950. [Google Scholar] [CrossRef]
- Elbakrawy, E.M.; Mayah, A.; Hill, M.A.; Kadhim, M. Induction of Genomic Instability in a Primary Human Fibroblast Cell Line Following Low-Dose Alpha-Particle Exposure and the Potential Role of Exosomes. Biology 2021, 10, 11. [Google Scholar] [CrossRef]
- Lobo, I. Biological Complexity and Integrative Levels of Organization. Nat. Educ. 2008, 1, 141. [Google Scholar]
- Baghban, R.; Roshangar, L.; Jahanban-Esfahlan, R.; Seidi, K.; Ebrahimi-Kalan, A.; Jaymand, M.; Kolahian, S.; Javaheri, T.; Zare, P. Tumor Microenvironment Complexity and Therapeutic Implications at a Glance. Cell Commun. Signal. 2020, 18, 59. [Google Scholar] [CrossRef]
- Moysidou, C.M.; Barberio, C.; Owens, R.M. Advances in Engineering Human Tissue Models. Front Bioeng. Biotechnol. 2021, 8, 620962. [Google Scholar] [CrossRef]
- Jalali, M.S.; Botticelli, M.; Hwang, R.C.; Koh, H.K.; McHugh, R.K. The Opioid Crisis: A Contextual, Social-Ecological Framework. Health Res. Policy Syst. 2020, 18, 87. [Google Scholar] [CrossRef]
- Tang, R.; Murray, C.W.; Linde, I.L.; Kramer, N.J.; Lyu, Z.; Tsai, M.K.; Chen, L.C.; Cai, H.; Gitler, A.D.; Engleman, E.; et al. A Versatile System to Record Cell-Cell Interactions. Elife 2020, 9, e61080. [Google Scholar] [CrossRef]
- Sachs, P.C.; Mollica, P.A.; Bruno, R.D. Tissue Specific Microenvironments: A Key Tool for Tissue Engineering and Regenerative Medicine. J. Biol. Eng. 2017, 11, 34. [Google Scholar] [CrossRef]
- Mao, Y.; Wang, X.; Huang, P.; Tian, R. Spatial Proteomics for Understanding the Tissue Microenvironment. Analyst 2021, 146, 3777–3798. [Google Scholar] [CrossRef] [PubMed]
- Fleming, G.R.; Scholes, G.D. Quantum Biology: Introduction. In Quantum Effects in Biology; Cambridge University Press: Cambridge, UK, 2014. [Google Scholar]
- Sarovar, M.; Ishizaki, A.; Fleming, G.R.; Whaley, K.B. Quantum Entanglement in Photosynthetic Light-Harvesting Complexes. Nat. Phys. 2010, 6, 462–467. [Google Scholar] [CrossRef]
- Bothma, J.P.; Gilmore, J.B.; McKenzie, R.H. The Role of Quantum Effects in Proton Transfer Reactions in Enzymes: Quantum Tunneling in a Noisy Environment? New J. Phys. 2010, 12, 055002. [Google Scholar] [CrossRef]
- Ritz, T. Quantum Effects in Biology: Bird Navigation. Proc. Procedia Chem. 2011, 3, 262–275. [Google Scholar] [CrossRef]
- Ishizaki, A.; Calhoun, T.R.; Schlau-Cohen, G.S.; Fleming, G.R. Quantum Coherence and Its Interplay with Protein Environments in Photosynthetic Electronic Energy Transfer. Phys. Chem. Chem. Phys. 2010, 12, 7319–7337. [Google Scholar] [CrossRef] [PubMed]
- Alenquer, M.; Amorim, M.J. Exosome Biogenesis, Regulation, and Function in Viral Infection. Viruses 2015, 7, 5066–5083. [Google Scholar] [CrossRef]
- Minciacchi, V.R.; Freeman, M.R.; Di Vizio, D. Extracellular Vesicles in Cancer: Exosomes, Microvesicles and the Emerging Role of Large Oncosomes. Semin. Cell Dev. Biol. 2015, 40, 41–51. [Google Scholar] [CrossRef] [PubMed]
- Zaborowski, M.P.; Balaj, L.; Breakefield, X.O.; Lai, C.P. Extracellular Vesicles: Composition, Biological Relevance, and Methods of Study. Bioscience 2015, 65, 783–797. [Google Scholar] [CrossRef]
- Valadi, H.; Ekström, K.; Bossios, A.; Sjöstrand, M.; Lee, J.J.; Lötvall, J.O. Exosome-Mediated Transfer of MRNAs and MicroRNAs Is a Novel Mechanism of Genetic Exchange between Cells. Nat. Cell Biol. 2007, 9, 654–659. [Google Scholar] [CrossRef]
- Kourembanas, S. Exosomes: Vehicles of Intercellular Signaling, Biomarkers, and Vectors of Cell Therapy. Annu. Rev. Physiol. 2015, 77, 13–27. [Google Scholar] [CrossRef] [PubMed]
- Raposo, G.; Stoorvogel, W. Extracellular Vesicles: Exosomes, Microvesicles, and Friends. J. Cell Biol. 2013, 200, 373–383. [Google Scholar] [CrossRef]
- Manousakis, E. Quantum mechanics and path integrals. In Practical Quantum Mechanics; Springer: Berlin/Heidelberg, Germany, 2015. [Google Scholar]
- Styer, D.F. The Strange World of Quantum Mechanics; Cambridge University Press: Cambridge, UK, 2014. [Google Scholar]
- Zettili, N.; Zahed, I. Quantum Mechanics: Concepts and Applications. Am. J. Phys. 2003, 71, 93. [Google Scholar] [CrossRef]
- Klein, M.J. Max Planck and the Beginnings of the Quantum Theory. Arch. Hist. Exact. Sci. 1975, 1, 459–479. [Google Scholar] [CrossRef]
- Feit, M.D.; Fleck, J.A.; Steiger, A. Solution of the Schrödinger Equation by a Spectral Method. J. Comput. Phys. 1982, 47, 412–433. [Google Scholar] [CrossRef]
- Shirley, J.H. Solution of the Schrödinger Equation with a Hamiltonian Periodic in Time. Phys. Rev. 1965, 138, B979–B987. [Google Scholar] [CrossRef]
- Ettlinger, H.J.; Dirac, P.A.B. The Principles of Quantum Mechanics. Am. Math. Mon. 1931, 38, 524. [Google Scholar] [CrossRef]
- Ananthaswamy, A. Particle, Wave, Both or Neither? The Experiment That Challenges All We Know about Reality. Nature 2023, 618, 454–456. [Google Scholar] [CrossRef] [PubMed]
- Duffy, J.; Loch-Temzelides, T. A Double-Slit Experiment with Human Subjects. PLoS ONE 2021, 16, e0246526. [Google Scholar] [CrossRef]
- Parker, S. A Single-Photon Double-Slit Interference Experiment. Am. J. Phys. 1971, 39, 420–424. [Google Scholar] [CrossRef]
- Tuszynski, J.A. From Quantum Chemistry to Quantum Biology: A Path toward Consciousness. J. Integr. Neurosci. 2020, 19, 687–700. [Google Scholar] [CrossRef] [PubMed]
- Cheng, H.P.; Deumens, E.; Freericks, J.K.; Li, C.; Sanders, B.A. Application of Quantum Computing to Biochemical Systems: A Look to the Future. Front Chem. 2020, 8, 587143. [Google Scholar] [CrossRef]
- Holland, R.A. True Navigation in Birds: From Quantum Physics to Global Migration. J. Zool. 2014, 293, 1–15. [Google Scholar] [CrossRef]
- Frederiksen, A.; Teusch, T.; Solov’yov, I.A. Quantum Effects in Biological Systems. In Dynamics of Systems on the Nanoscale; Springer International Publishing: Cham, Switzerland, 2022. [Google Scholar]
- Calvillo, L.; Redaelli, V.; Ludwig, N.; Qaswal, A.B.; Ghidoni, A.; Faini, A.; Rosa, D.; Lombardi, C.; Pengo, M.; Bossolasco, P.; et al. Quantum Biology Research Meets Pathophysiology and Therapeutic Mechanisms: A Biomedical Perspective. Quantum Rep. 2022, 4, 148–172. [Google Scholar] [CrossRef]
- Nishiura, N.; Kaneko, K. Evolution of Phenotypic Fluctuation under Host-Parasite Interactions. PLoS Comput. Biol. 2021, 17, e1008694. [Google Scholar] [CrossRef]
- Aristarhov, S. Heisenberg’s Uncertainty Principle and Particle Trajectories. Found Phys. 2023, 53, 1–12. [Google Scholar] [CrossRef]
- Busch, P.; Heinonen, T.; Lahti, P. Heisenberg’s Uncertainty Principle. Phys. Rep. 2007, 452, 155–176. [Google Scholar] [CrossRef]
- Cai, J. Quantum Biology: Explore Quantum Dynamics in Biological Systems. Sci. China Inf. Sci. 2016, 59, 1–7. [Google Scholar] [CrossRef]
- Hammerschlag, R.; Levin, M.; McCraty, R.; Bat, N.; Ives, J.A.; Lutgendorf, S.K.; Oschman, J.L. Biofield Physiology: A Framework for an Emerging Discipline. Glob. Adv. Health Med. 2015, 4, 35–41. [Google Scholar] [CrossRef]
- Fels, D. Cellular Communication through Light. PLoS ONE 2009, 4, e5086. [Google Scholar] [CrossRef]
- Takeda, M.; Kobayashi, M.; Takayama, M.; Suzuki, S.; Ishida, T.; Ohnuki, K.; Moriya, T.; Ohuchi, N. Biophoton Detection as a Novel Technique for Cancer Imaging. Cancer Sci. 2004, 95, 656–661. [Google Scholar] [CrossRef] [PubMed]
- Sjostedt, S.; Bezak, E. Non-Targeted Effects of Ionising Radiation and Radiotherapy. Australas Phys. Eng. Sci. Med. 2010, 33, 219–231. [Google Scholar] [CrossRef] [PubMed]
- Zhou, H.; Ivanov, V.N.; Gillespie, J.; Geard, C.R.; Amundson, S.A.; Brenner, D.J.; Yu, Z.; Lieberman, H.B.; Hei, T.K. Mechanism of Radiation-Induced Bystander Effect: Role of the Cyclooxygenase-2 Signaling Pathway. Proc. Natl. Acad. Sci. USA 2005, 102, 14641–14646. [Google Scholar] [CrossRef]
- Benfatto, M.; Pace, E.; Curceanu, C.; Scordo, A.; Clozza, A.; Davoli, I.; Lucci, M.; Francini, R.; De Matteis, F.; Grandi, M.; et al. Biophotons and Emergence of Quantum Coherence—A Diffusion Entropy Analysis. Entropy 2021, 23, 554. [Google Scholar] [CrossRef]
- Wang, S.; Qiu, L.; Liu, X.; Xu, G.; Siegert, M.; Lu, Q.; Juneau, P.; Yu, L.; Liang, D.; He, Z.; et al. Electron Transport Chains in Organohalide-Respiring Bacteria and Bioremediation Implications. Biotechnol. Adv. 2018, 36, 1194–1206. [Google Scholar] [CrossRef]
- Peters, J.W.; Fisher, K.; Newton, W.E.; Dean, D.R. Involvement of the P Cluster in Intramolecular Electron Transfer within the Nitrogenase MoFe Protein. J. Biol. Chem. 1995, 270, 27007–27013. [Google Scholar] [CrossRef]
- Gray, H.B.; Winkler, J.R. Electron Flow through Metalloproteins. Biochim. Biophys. Acta Bioenerg. 2010, 1797, 1563–1572. [Google Scholar] [CrossRef]
- Zhang, P.; Yuly, J.L.; Lubner, C.E.; Mulder, D.W.; King, P.W.; Peters, J.W.; Beratan, D.N. Electron Bifurcation: Thermodynamics and Kinetics of Two-Electron Brokering in Biological Redox Chemistry. Acc. Chem. Res. 2017, 50, 2410–2417. [Google Scholar] [CrossRef]
- Reece, S.Y.; Nocera, D.G. Proton-Coupled Electron Transfer in Biology: Results from Synergistic Studies in Natural and Model Systems. Annu. Rev. Biochem. 2009, 78, 673–699. [Google Scholar] [CrossRef]
- Huynh, M.H.V.; Meyer, T.J. Proton-Coupled Electron Transfer. Chem. Rev. 2007, 107, 5004–5064. [Google Scholar] [CrossRef] [PubMed]
- Paganetti, H. Range Uncertainties in Proton Therapy and the Role of Monte Carlo Simulations. Phys. Med. Biol. 2012, 57, R99–R117. [Google Scholar] [CrossRef] [PubMed]
- Mohan, R. A Review of Proton Therapy–Current Status and Future Directions. Precis. Radiat. Oncol. 2022, 6, 164–176. [Google Scholar] [CrossRef] [PubMed]
- Ooms, M.D.; Dinh, C.T.; Sargent, E.H.; Sinton, D. Photon Management for Augmented Photosynthesis. Nat. Commun. 2016, 7, 12699. [Google Scholar] [CrossRef]
- Applebury, M.L. Dynamic Processes of Visual Transduction. Vision Res. 1984, 24, 1445–1454. [Google Scholar] [CrossRef]
- Pathania, N.; Qureshi, T. Quantifying Entanglement with Coherence. Int. J. Theor. Phys. 2022, 61. [Google Scholar] [CrossRef]
- Schrödinger, E. Die Gegenwärtige Situation in Der Quantenmechanik. Naturwissenschaften 1935, 23, 807–812. [Google Scholar] [CrossRef]
- Li, T.; Yin, Z.Q. Quantum Superposition, Entanglement, and State Teleportation of a Microorganism on an Electromechanical Oscillator. Sci. Bull. 2016, 61, 163–171. [Google Scholar] [CrossRef]
- Friedman, J.R.; Patel, V.; Chen, W.; Tolpygo, S.K.; Lukens, J.E. Quantum Superposition of Distinct Macroscopic States. Nature 2000, 406, 43–46. [Google Scholar] [CrossRef]
- Fein, Y.Y.; Geyer, P.; Zwick, P.; Kiałka, F.; Pedalino, S.; Mayor, M.; Gerlich, S.; Arndt, M. Quantum Superposition of Molecules beyond 25 KDa. Nat. Phys. 2019, 15, 1242–1245. [Google Scholar] [CrossRef]
- Romero, E.; Augulis, R.; Novoderezhkin, V.I.; Ferretti, M.; Thieme, J.; Zigmantas, D.; Van Grondelle, R. Quantum Coherence in Photosynthesis for Efficient Solar-Energy Conversion. Nat. Phys. 2014, 10, 676–682. [Google Scholar] [CrossRef]
- Yuan, J.; Hazarika, A.; Zhao, Q.; Ling, X.; Moot, T.; Ma, W.; Luther, J.M. Metal Halide Perovskites in Quantum Dot Solar Cells: Progress and Prospects. Joule 2020, 4, 1160–1185. [Google Scholar]
- Sharma, D.; Jha, R.; Kumar, S. Quantum Dot Sensitized Solar Cell: Recent Advances and Future Perspectives in Photoanode. Sol. Energy Mater. Sol. Cells 2016, 155, 294–322. [Google Scholar] [CrossRef]
- Ollivier, H.; Zurek, W.H. Quantum Discord: A Measure of the Quantumness of Correlations. Phys. Rev. Lett. 2002, 88, 017901. [Google Scholar] [CrossRef] [PubMed]
- Garrison, J.; Chiao, R. Quantum Optics; Oxford University Press: Oxford, UK, 2008; ISBN 9780198508861. [Google Scholar]
- Lvovsky, A.I.; Raymer, M.G. Continuous-Variable Optical Quantum-State Tomography. Rev. Mod. Phys. 2009, 81, 299–332. [Google Scholar] [CrossRef]
- Bae, J.; Kwek, L.C. Quantum State Discrimination and Its Applications. J. Phys. A Math Theor. 2015, 48, 083001. [Google Scholar]
- Niaz, M.; Fernández, R. Understanding Quantum Numbers in General Chemistry Textbooks. Int. J. Sci. Educ. 2008, 30, 869–901. [Google Scholar] [CrossRef]
- Wolf, S.A.; Awschalom, D.D.; Buhrman, R.A.; Daughton, J.M.; Von Molnár, S.; Roukes, M.L.; Chtchelkanova, A.Y.; Treger, D.M. Spintronics: A Spin-Based Electronics Vision for the Future. Science 2001, 294, 1488–1495. [Google Scholar]
- Žutić, I.; Fabian, J.; Sarma, S. Das Spintronics: Fundamentals and Applications. Rev. Mod. Phys. 2004, 76, 1–9. [Google Scholar]
- Chilton, N.F. Molecular Magnetism. Annu. Rev. Mater Res. 2022, 52, 79–101. [Google Scholar]
- Pauli, W. The Connection between Spin and Statistics. Phys. Rev. 1940, 58, 716–722. [Google Scholar] [CrossRef]
- Born, M. Statistical Interpretation of Quantum Mechanics. Science 1955, 122, 675–679. [Google Scholar] [CrossRef] [PubMed]
- Dirac, P. The Quantum Theory of the Electron. Proc. R. Soc. Lond. Ser. A Contain. Pap. A Math. Phys. Character 1928, 117, 610–624. [Google Scholar] [CrossRef]
- Varshalovich, D.A.; Moskalev, A.N.; Khersonskii, V.K. Quantum Theory of Angular Momentum; World Scientific Publishing Co. Pte. Ltd.: London, UK, 1988. [Google Scholar]
- Luo, S. Quantum Discord for Two-Qubit Systems. Phys. Rev. A 2008, 77. [Google Scholar] [CrossRef]
- Plenio, M.B.; Virmani, S. An Introduction to Entanglement Measures. Quantum Inf. Comput. 2007, 7, 1–51. [Google Scholar]
- Horodecki, R.; Horodecki, P.; Horodecki, M.; Horodecki, K. Quantum Entanglement. Rev. Mod. Phys. 2009, 81, 865–942. [Google Scholar] [CrossRef]
- Griffiths, D.J.; Schroeter, D.F. Introduction to Quantum Mechanics, 1st ed.; Cambridge University Press: Cambridge, UK, 2018; ISBN 1108100341. [Google Scholar]
- Einstein, A.; Podolsky, B.; Rosen, N. Can Quantum-Mechanical Description of Physical Reality Be Considered Complete? Phys. Rev. 1935, 47, 777. [Google Scholar] [CrossRef]
- Karimi, E.; Boyd, R.W. Classical Entanglement? Science 2015, 350, 1172–1173. [Google Scholar] [CrossRef]
- Galanis, C.; Vlachos, A. Hebbian and Homeostatic Synaptic Plasticity—Do Alterations of One Reflect Enhancement of the Other? Front Cell Neurosci. 2020, 14, 50. [Google Scholar] [CrossRef]
- Cleve, R.; Buhrman, H. Substituting Quantum Entanglement for Communication. Phys. Rev. A 1997, 56, 1201. [Google Scholar]
- Zeilinger, A. Experiment and the Foundations of Quantum Physics. Rev. Mod. Phys. 1999, 71, S288–S297. [Google Scholar] [CrossRef]
- Ursin, R.; Tiefenbacher, F.; Schmitt-Manderbach, T.; Weier, H.; Scheidl, T.; Lindenthal, M.; Blauensteiner, B.; Jennewein, T.; Perdigues, J.; Trojek, P. Entanglement-Based Quantum Communication over 144 Km. Nat. Phys. 2007, 3, 481–486. [Google Scholar] [CrossRef]
- Yang, X.; Wei, K.; Ma, H.; Sun, S.; Liu, H.; Yin, Z.; Li, Z.; Lian, S.; Du, Y.; Wu, L. Measurement-Device-Independent Entanglement-Based Quantum Key Distribution. Phys. Rev. A 2016, 93, 052303. [Google Scholar] [CrossRef]
- Rochlin, G.I. Tunneling Phenomena in Solids. J. Franklin Inst. 1971, 292, 66. [Google Scholar] [CrossRef]
- Dimitrova, T.L.; Weis, A. The Wave-Particle Duality of Light: A Demonstration Experiment. Am. J. Phys. 2008, 76, 137–142. [Google Scholar] [CrossRef]
- Kunitski, M.; Eicke, N.; Huber, P.; Köhler, J.; Zeller, S.; Voigtsberger, J.; Schlott, N.; Henrichs, K.; Sann, H.; Trinter, F.; et al. Double-Slit Photoelectron Interference in Strong-Field Ionization of the Neon Dimer. Nat. Commun. 2019, 10, 1. [Google Scholar] [CrossRef]
- Bach, R.; Pope, D.; Liou, S.H.; Batelaan, H. Controlled Double-Slit Electron Diffraction. New J. Phys. 2013, 15, 033018. [Google Scholar] [CrossRef]
- Wemer, H. The Physical Principles of the Quantum Theory; Dover Publication: Mineola, NY, USA, 1949; ISBN 0486601137. [Google Scholar]
- Kennard, E.H. On the Quantum Mechanics of a System of Particles. Phys. Rev. 1928, 31, 876–890. [Google Scholar] [CrossRef]
- Klinman, J.P.; Kohen, A. Hydrogen Tunneling Links Protein Dynamics to Enzyme Catalysis. Annu. Rev. Biochem. 2013, 82, 471–496. [Google Scholar] [CrossRef]
- Truhlar, D.G.; Gao, J.; Alhambra, C.; Garcia-Viloca, M.; Corchado, J.; Sánchez, M.L.; Villà, J. The Incorporation of Quantum Effects in Enzyme Kinetics Modeling. Acc. Chem. Res. 2002, 35, 341–349. [Google Scholar] [CrossRef]
- Nagel, Z.D.; Klinman, J.P. Tunneling and Dynamics in Enzymatic Hydride Transfer. Chem. Rev. 2006, 106, 3095–3118. [Google Scholar] [CrossRef] [PubMed]
- Kržan, M.; Vianello, R.; Maršavelski, A.; Repič, M.; Zakšek, M.; Kotnik, K.; Fijan, E.; Mavri, J. The Quantum Nature of Drug-Receptor Interactions: Deuteration Changes Binding Affinities for Histamine Receptor Ligands. PLoS ONE 2016, 11, e0154002. [Google Scholar] [CrossRef] [PubMed]
- Jaeger, G. Entanglement, Information, and the Interpretation of Quantum Mechanics. Choice Rev. Online 2010, 47, 3870. [Google Scholar] [CrossRef]
- Andersen, M.L.; Stobbe, S.; Sørensen, A.S.; Lodahl, P. Strongly Modified Plasmon-Matter Interaction with Mesoscopic Quantum Emitters. Nat. Phys. 2011, 7, 215–218. [Google Scholar] [CrossRef]
- Wolf, F.A. Mind into Matter: A New Alchemy of Science and Spirit; Red Wheel/Weiser: Newburyport, MA, USA, 2001. [Google Scholar]
- Eibenberger, S.; Gerlich, S.; Arndt, M.; Mayor, M.; Tüxen, J. Matter–Wave Interference of Particles Selected from a Molecular Library with Masses Exceeding 10000 Amu. Phys. Chem. Chem. Phys. 2013, 15, 14696–14700. [Google Scholar] [CrossRef] [PubMed]
- Roger, P. The Road to Reality: A Complete Guide to the Laws of the Universe, 1st ed.; Jonathan Cape: London, UK, 2004. [Google Scholar]
- Jozsa, R.; Linden, N. On the Role of Entanglement in Quantum-Computational Speed-Up. Proc. R. Soc. A Math. Phys. Eng. Sci. 2003, 459, 2011–2032. [Google Scholar] [CrossRef]
- Gröblacher, S.; Paterek, T.; Kaltenbaek, R.; Brukner, Č.; Zukowski, M.; Aspelmeyer, M.; Zeilinger, A. An Experimental Test of Non-Local Realism. Nature 2007, 446, 871–875. [Google Scholar] [CrossRef]
- Fan, Y.; Guo, X.; Yang, X. Quantifying Coherence of Quantum Channels via Trace Distance. Quantum Inf. Process 2022, 21, 1–16. [Google Scholar] [CrossRef]
- Li, C.M.; Lambert, N.; Chen, Y.N.; Chen, G.Y.; Nori, F. Witnessing Quantum Coherence: From Solid-State to Biological Systems. Sci. Rep. 2012, 2, srep00885. [Google Scholar] [CrossRef]
- Roztocki, N.; Soja, P.; Weistroffer, H.R. The Role of Information and Communication Technologies in Socioeconomic Development: Towards a Multi-Dimensional Framework. Inf. Technol. Dev. 2019, 25, 171–183. [Google Scholar] [CrossRef]
- Flöther, F.F.; Griffin, P.F. How Can Quantum Technologies Be Applied in Healthcare, Medicine and the Life Sciences? Res. Dir. Quantum Technol. 2023, 1. [Google Scholar] [CrossRef]
- Shimazoe, K.; Tomita, H.; Watts, D.; Moskal, P.; Kagawa, A.; Thirolf, P.G.; Budker, D.; Levin, C.S. Quantum Sensing for Biomedical Applications. In Proceedings of the 2021 IEEE Nuclear Science Symposium and Medical Imaging Conference Record, NSS/MIC 2021, Yokohama, Japan, 16–23 October 2021. [Google Scholar]
- Um, M.; Ro, D.; Chang, I.J.; Lee, H.M. A Radiation-Hardened Readout Integrated Circuits for Sensor Systems. In Proceedings of the 2020 IEEE International Conference on Consumer Electronics-Asia, ICCE-Asia 2020, Seoul, Republic of Korea, 1–3 November 2020. [Google Scholar]
- Jezzard, P.; Clare, S. Principles of nuclear magnetic resonance and MRI. In Functional Magnetic Resonance Imaging: An Introduction to Methods; Oxford Academic: Oxford, UK, 2012. [Google Scholar]
- Li, M.; Jiang, F.; Xue, L.; Peng, C.; Shi, Z.; Zhang, Z.; Li, J.; Pan, Y.; Wang, X.; Feng, C.; et al. Recent Progress in Biosensors for Detection of Tumor Biomarkers. Molecules 2022, 27, 7327. [Google Scholar] [CrossRef]
- Casacio, C.A.; Madsen, L.S.; Terrasson, A.; Waleed, M.; Barnscheidt, K.; Hage, B.; Taylor, M.A.; Bowen, W.P. Quantum-Enhanced Nonlinear Microscopy. Nature 2021, 594, 201–206. [Google Scholar] [CrossRef] [PubMed]
- Datta, A. Quantum-Enhanced Stimulated Emission Microscopy. In Proceedings of the Emerging Imaging and Sensing Technologies for Security and Defence V; and Advanced Manufacturing Technologies for Micro- and Nanosystems in Security and Defence III, Online Only, UK, 21–25 September 2020; SPIE: Bellingham, WA, USA, 2020. [Google Scholar]
- Wendin, G. Quantum Information Processing with Superconducting Circuits: A Review. Rep. Prog. Phys. 2017, 80, 106001. [Google Scholar] [CrossRef]
- Weedbrook, C.; Pirandola, S.; García-Patrón, R.; Cerf, N.J.; Ralph, T.C.; Shapiro, J.H.; Lloyd, S. Gaussian Quantum Information. Rev. Mod. Phys. 2012, 84, 621–669. [Google Scholar] [CrossRef]
- Braunstein, L.S.; Van Loock, P. Quantum Information with Continuous Variables. Rev. Mod. Phys. 2005, 77, 513–577. [Google Scholar] [CrossRef]
- Pauls, J.A.; Zhang, Y.; Berman, G.P.; Kais, S. Quantum Coherence and Entanglement in the Avian Compass. Phys. Rev. E Stat. Nonlin. Soft Matter Phys. 2013, 87, 062704. [Google Scholar] [CrossRef]
- Sun, Y.; Wang, C.; Dai, J. Biophotons as Neural Communication Signals Demonstrated by in Situ Biophoton Autography. Photochem. Photobiol. Sci. 2010, 9, 315–322. [Google Scholar] [CrossRef]
- Popp, F.A.; Nagl, W.; Li, K.H.; Scholz, W.; Weingärtner, O.; Wolf, R. Biophoton Emission-New Evidence for Coherence and DNA as Source. Cell Biophys. 1984, 6, 33–52. [Google Scholar] [CrossRef]
- Sawant, S.G.; Randers-Pehrson, G.; Geard, C.R.; Brenner, D.J.; Hall, E.J. The Bystander Effect in Radiation Oncogenesis: I. Transformation in C3H 10T1/2 Cells in Vitro Can Be Initiated in the Unirradiated Neighbors of Irradiated Cells. Radiat. Res. 2001, 155, 397–401. [Google Scholar] [CrossRef]
- Brenner, D.J.; Little, J.B.; Sachs, R.K. The Bystander Effect in Radiation Oncogenesis: II. A Quantitative Model. Radiat. Res. 2001, 155, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Giovannetti, V.; Lloyd, S.; Maccone, L. Quantum-Enhanced Measurements: Beating the Standard Quantum Limit. Science 2004, 306, 1330–1336. [Google Scholar] [CrossRef] [PubMed]
- Joo, J.; Munro, W.J.; Spiller, T.P. Quantum Metrology with Entangled Coherent States. Phys. Rev. Lett. 2011, 107, 083601. [Google Scholar] [CrossRef] [PubMed]
- Piccolino, M.; Bresadola, M.; Wade, N. The controversy between Galvani and Volta over animal electricity. In Shocking Frogs; Oxford University Press: Oxford, UK, 2014. [Google Scholar]
- Tyler, S.E.B. The Work Surfaces of Morphogenesis: The Role of the Morphogenetic Field. Biol. Theory 2014, 9, 194–208. [Google Scholar] [CrossRef]
- Kim, H.J.; Lee, S. Relation between Quantum Coherence and Quantum Entanglement in Quantum Measurements. Phys. Rev. A 2022, 106, 022401. [Google Scholar] [CrossRef]
- Ren, J.G.; Xu, P.; Yong, H.L.; Zhang, L.; Liao, S.K.; Yin, J.; Liu, W.Y.; Cai, W.Q.; Yang, M.; Li, L.; et al. Ground-to-Satellite Quantum Teleportation. Nature 2017, 549, 70–73. [Google Scholar] [CrossRef]
- Debnath, P.; Huirem, R.S.; Dutta, P.; Palchaudhuri, S. Epithelial-Mesenchymal Transition and Its Transcription Factors. Biosci. Rep. 2022, 42, BSR20211754. [Google Scholar] [CrossRef]
- Bhavsar, M.B.; Leppik, L.; Costa Oliveira, K.M.; Barker, J.H. Role of Bioelectricity During Cell Proliferation in Different Cell Types. Front Bioeng. Biotechnol. 2020, 8, 603. [Google Scholar] [CrossRef]
- George, L.F.; Bates, E.A. Mechanisms Underlying Influence of Bioelectricity in Development. Front. Cell Dev. Biol. 2022, 10, 772230. [Google Scholar]
- Camelo, L.G. Pathophysiology of Biophoton: Vibratory Impact Syndrome Leading to Physical Effects and Metabolic Changes—Part 1. Neurosci. Med. 2021, 12, 126–162. [Google Scholar] [CrossRef]
- Zou, N. Quantum Entanglement and Its Application in Quantum Communication. Proc. J. Phys. Conf. Ser. 2021, 1827, 012120. [Google Scholar]
- Belousov, L.V. “Our Standpoint Different from Common...” (Scientific Heritage of Alexander Gurwitsch). Russ. J. Dev. Biol. 2008, 39, 307–315. [Google Scholar] [CrossRef]
- Gurwitsch, A.A. A Historical Review of the Problem of Mitogenetic Radiation. Experientia 1988, 44, 545–550. [Google Scholar] [PubMed]
- Jung, S.K.; Lee, K.W.; Byun, S.; Lee, E.J.; Kim, J.E.; Bode, A.M.; Dong, Z.; Lee, H.J. Myricetin Inhibits UVB-Induced Angiogenesis by Regulating PI-3 Kinase in Vivo. Carcinogenesis 2009, 31, 911–917. [Google Scholar] [CrossRef] [PubMed]
- Kaiser, S.; Scheuring, D. To Lead or to Follow: Contribution of the Plant Vacuole to Cell Growth. Front Plant Sci. 2020, 11, 553. [Google Scholar] [PubMed]
- Syljuåsen, R.G. Cell Cycle Effects in Radiation Oncology. In Radiation Oncology; Springer: Berlin/Heidelberg, Germany, 2019. [Google Scholar]
- Fraikin, G.Y.; Belenikina, N.S. Photochemistry and Signaling Activity of Plant Cryptochromes: A Review. Biol. Bull. 2023, 50, 266–275. [Google Scholar] [CrossRef]
- Fraikin, G.Y. Photosensory and Signaling Properties of Cryptochromes. Moscow Univ. Biol. Sci. Bull. 2022, 77, 54–63. [Google Scholar] [CrossRef]
- Chadwick, G.L.; Skennerton, C.T.; Laso-Pérez, R.; Leu, A.O.; Speth, D.R.; Yu, H.; Morgan-Lang, C.; Hatzenpichler, R.; Goudeau, D.; Malmstrom, R.; et al. Comparative Genomics Reveals Electron Transfer and Syntrophic Mechanisms Differentiating Methanotrophic and Methanogenic Archaea. PLoS Biol. 2022, 20, e3001508. [Google Scholar] [CrossRef]
- Lanting, T.; Przybysz, A.J.; Smirnov, A.Y.; Spedalieri, F.M.; Amin, M.H.; Berkley, A.J.; Harris, R.; Altomare, F.; Boixo, S.; Bunyk, P.; et al. Entanglement in a Quantum Annealing Processor. Phys. Rev. X 2014, 4, 021041. [Google Scholar] [CrossRef]
- Devault, D. Quantum Mechanical Tunnelling in Biological Systems. Q Rev. Biophys. 1980, 13, 387–564. [Google Scholar] [CrossRef]
- Ball, P. Is Photosynthesis Quantum-Ish? Phys. World 2018, 31, 249–251. [Google Scholar] [CrossRef]
- Xu, C.; Zhang, J.; Mihai, D.M.; Washington, I. Light-Harvesting Chlorophyll Pigments Enable Mammalian Mitochondria to Capture Photonic Energy and Produce ATP. J. Cell Sci. 2014, 127, 388–399. [Google Scholar] [CrossRef]
- Yeles, C.; Vlachavas, E.I.; Papadodima, O.; Pilalis, E.; Vorgias, C.E.; Georgakilas, A.G.; Chatziioannou, A. Integrative Bioinformatic Analysis of Transcriptomic Data Identifies Conserved Molecular Pathways Underlying Ionizing Radiation-Induced Bystander Effects (RIBE). Cancers 2017, 9, 160. [Google Scholar] [CrossRef] [PubMed]
- Sia, P.I.; Luiten, A.N.; Stace, T.M.; Wood, J.P.M.; Casson, R.J. Quantum Biology of the Retina. Clin. Exp. Ophthalmol. 2014, 42, 582–589. [Google Scholar] [PubMed]
- Sun, L.; Zhao, L.; Peng, R.Y. Research Progress in the Effects of Terahertz Waves on Biomacromolecules. Mil. Med. Res. 2021, 8, 1–8. [Google Scholar]
- Lindinger, M.I. Structured Water: Effects on Animals. J. Anim. Sci. 2021, 99, skab063. [Google Scholar] [CrossRef]
- Danopoulos, E.; Twiddy, M.; Rotchell, J.M. Microplastic Contamination of Drinking Water: A Systematic Review. PLoS ONE 2020, 15, e0236838. [Google Scholar]
- Pal, S.; Batra, P.; Krisnanda, T.; Paterek, T.; Mahesh, T.S. Experimental Localisation of Quantum Entanglement through Monitored Classical Mediator. Quantum 2021, 5, 478. [Google Scholar] [CrossRef]
- Khrennikov, A. Quantum Versus Classical Entanglement: Eliminating the Issue of Quantum Nonlocality. Found Phys. 2020, 50, 1762–1780. [Google Scholar] [CrossRef]
- Slocombe, L.; Al-Khalili, J.S.; Sacchi, M. Quantum and Classical Effects in DNA Point Mutations: Watson-Crick Tautomerism in at and GC Base Pairs. Phys. Chem. Chem. Phys. 2021, 23, 4141–4150. [Google Scholar] [CrossRef]
- Slocombe, L.; Sacchi, M.; Al-Khalili, J. An Open Quantum Systems Approach to Proton Tunnelling in DNA. Commun. Phys. 2022, 5, 1–9. [Google Scholar] [CrossRef]
- Löwdin, P.O. Quantum Genetics and the Aperiodic Solid. Some Aspects on the Biological Problems of Heredity, Mutations, Aging, and Tumors in View of the Quantum Theory of the DNA Molecule. In Advances in Quantum Chemistry; Academic Press: Cambridge, MA, USA, 1966; pp. 213–360. [Google Scholar] [CrossRef]
- Guallar, V.; Douhal, A.; Moreno, M.; Lluch, J.M. DNA Mutations Induced by Proton and Charge Transfer in the Low-Lying Excited Singlet Electronic States of the DNA Base Pairs: A Theoretical Insight. J. Phys. Chem. A 1999, 103, 6251–6256. [Google Scholar] [CrossRef]
- Yang, M. A Quantum Mechanical Approach to Understanding DNA Mutations. Berkeley Sci. J. 2019, 24, 37–40. [Google Scholar] [CrossRef]
- Yin, M.; O’Neill, L.A.J. The Role of the Electron Transport Chain in Immunity. FASEB J. 2021, 35, e21974. [Google Scholar] [PubMed]
- Zhao, R.Z.; Jiang, S.; Zhang, L.; Yu, Z. Bin Mitochondrial Electron Transport Chain, ROS Generation and Uncoupling (Review). Int. J. Mol. Med. 2019, 44, 3–15. [Google Scholar] [PubMed]
- Cogliati, S.; Cabrera-Alarcón, J.L.; Enriquez, J.A. Regulation and Functional Role of the Electron Transport Chain Supercomplexes. Biochem. Soc. Trans. 2021, 49, 2655–2668. [Google Scholar]
- Colombo, M.; Raposo, G.; Théry, C. Biogenesis, Secretion, and Intercellular Interactions of Exosomes and Other Extracellular Vesicles. Annu. Rev. Cell Dev. Biol. 2014, 30, 255–289. [Google Scholar]
- Van Niel, G.; D’Angelo, G.; Raposo, G. Shedding Light on the Cell Biology of Extracellular Vesicles. Nat. Rev. Mol. Cell Biol. 2018, 19, 213–228. [Google Scholar]
- Fedunov, R.G.; Ivanov, A.I. Effect of the Excitation Pulse Frequency on the Ultrafast Photoinduced Electron Transfer Dynamics. J. Chem. Phys. 2005, 122, 064501. [Google Scholar] [CrossRef]
- Parlak, C.; Ramasami, P. Theoretical and Experimental Study of Infrared Spectral Data of 2-Bromo-4-Chlorobenzaldehyde. SN Appl. Sci. 2020, 2, 1–9. [Google Scholar] [CrossRef]
- Vatansever, F.; Hamblin, M.R. Far Infrared Radiation (FIR): Its Biological Effects and Medical Applications. Photonics Lasers Med. 2012, 1, 255–266. [Google Scholar] [CrossRef]
- Sengupta, B.; Stemmler, M.; Laughlin, S.B.; Niven, J.E. Action Potential Energy Efficiency Varies among Neuron Types in Vertebrates and Invertebrates. PLoS Comput. Biol. 2010, 6, e1000840. [Google Scholar] [CrossRef]
- Aubrun, G.; Lami, L.; Palazuelos, C.; Plávala, M. Entanglement and Superposition Are Equivalent Concepts in Any Physical Theory. Phys. Rev. Lett. 2022, 128, 160402. [Google Scholar] [CrossRef] [PubMed]
- Gidon, A.; Aru, J.; Larkum, M.E. Does Brain Activity Cause Consciousness? A Thought Experiment. PLoS Biol. 2022, 20, e3001651. [Google Scholar] [CrossRef]
- Kim, J.H.; Chae, J.W.; Jeong, Y.C.; Kim, Y.H. Long-Range Distribution of High-Quality Time-Bin Entangled Photons for Quantum Communication. J. Korean Phys. Soc. 2022, 80, 203–213. [Google Scholar] [CrossRef]
- Xu, G.; Xiao, K.; Li, Z.; Niu, X.X.; Ryan, M. Controlled Secure Direct Communication Protocol via the Three-Qubit Partially Entangled Set of States. Comput. Mater. Contin. 2019, 58, 809–827. [Google Scholar] [CrossRef]
- Sommer, A.P.; Haddad, M.K.; Fecht, H.J. Light Effect on Water Viscosity: Implication for ATP Biosynthesis. Sci. Rep. 2015, 5, 12029. [Google Scholar] [CrossRef]
- Nesterov, S.V.; Smirnova, E.G.; Yaguzhinsky, L.S. Mechanism of Energy Storage and Transformation in the Mitochondria at the Water–Membrane Interface. Biochemistry 2022, 87, 179–190. [Google Scholar] [CrossRef]
- Kühlbrandt, W. Structure and Function of Mitochondrial Membrane Protein Complexes. BMC Biol. 2015, 13, 89. [Google Scholar] [CrossRef]
- Dröse, S.; Krack, S.; Sokolova, L.; Zwicker, K.; Barth, H.D.; Morgner, N.; Heide, H.; Steger, M.; Nübel, E.; Zickermann, V.; et al. Functional Dissection of the Proton Pumping Modules of Mitochondrial Complex I. PLoS Biol. 2011, 9, e1001128. [Google Scholar] [CrossRef]
- How Mitochondria Evolved from Bacteria. Nat. India 2022. [CrossRef]
- Carvalho, D.S.; Andrade, R.F.S.; Pinho, S.T.R.; Góes-Neto, A.; Lobão, T.C.P.; Bomfim, G.C.; El-Hani, C.N. What Are the Evolutionary Origins of Mitochondria? A Complex Network Approach. PLoS ONE 2015, 10, e0134988. [Google Scholar] [CrossRef] [PubMed]
- Damulewicz, M.; Mazzotta, G.M. One Actor, Multiple Roles: The Performances of Cryptochrome in Drosophila. Front Physiol. 2020, 11, 99. [Google Scholar] [PubMed]
- Ponnu, J.; Hoecker, U. Signaling Mechanisms by Arabidopsis Cryptochromes. Front Plant Sci. 2022, 13, 844714. [Google Scholar] [PubMed]
- Sullivan, C.M.; Nienhaus, L. Generating Spin-Triplet States at the Bulk Perovskite/Organic Interface for Photon Upconversion. Nanoscale 2022, 15, 998–1013. [Google Scholar]
- Nebgen, B.; Emmert, F.L.; Slipchenko, L.V. Vibronic Coupling in Asymmetric Bichromophores: Theory and Application to Diphenylmethane. J. Chem. Phys. 2012, 137, 084112. [Google Scholar] [CrossRef]
- Jansen, T.L.C.; Saito, S.; Jeon, J.; Cho, M. Theory of Coherent Two-Dimensional Vibrational Spectroscopy. J. Chem. Phys. 2019, 150, 100901. [Google Scholar] [CrossRef]
- Nishiyama, A.; Tanaka, S.; Tuszynski, J.A. Non-Equilibrium Quantum Brain Dynamics: Super-Radiance and Equilibration in 2 + 1 Dimensions. Entropy 2019, 21, 1066. [Google Scholar] [CrossRef]
- Yan, W.; Diao, S.; Fan, Z. The Role and Mechanism of Mitochondrial Functions and Energy Metabolism in the Function Regulation of the Mesenchymal Stem Cells. Stem Cell Res. Ther. 2021, 12, 1–17. [Google Scholar] [CrossRef]
- Schweber, S. Herbert Fröhlich: A Physicist Ahead of His Time. Phys. Today 2016, 69, 52. [Google Scholar] [CrossRef]
- Fröhlich, H. Coherent Excitation in Active Biological Systems. In Modern Bioelectrochemistry; Springer: Berlin/Heidelberg, Germany, 1986. [Google Scholar]
- Frohlich, H. Coherent Electric Vibrations in Biological Systems and the Cancer Problem. IEEE Trans. Microw. Theory Tech. 1978, 26, 613–618. [Google Scholar] [CrossRef]
- Pokorný, J. Coherent Vibration Interaction among Cells in Biological Systems. Czechoslov. J. Phys. 1980, 30, 1339–1342. [Google Scholar] [CrossRef]
- Cho, K.H.; Rhee, Y.M. Computational Elucidations on the Role of Vibrations in Energy Transfer Processes of Photosynthetic Complexes. Phys. Chem. Chem. Phys. 2021, 23, 26623–26639. [Google Scholar] [CrossRef] [PubMed]
- Daniel, C. Ultrafast Processes: Coordination Chemistry and Quantum Theory. Phys. Chem. Chem. Phys. 2021, 23, 43–58. [Google Scholar] [CrossRef]
- Feinberg, T.E.; Mallatt, J. Phenomenal Consciousness and Emergence: Eliminating the Explanatory Gap. Front Psychol. 2020, 11, 1041. [Google Scholar] [CrossRef]
- Hameroff, S. How Quantum Brain Biology Can Rescue Conscious Free Will. Front Integr. Neurosci. 2012, 6, 93. [Google Scholar] [CrossRef]
- York, A. Biofilms: Shocking Biofilms. Nat. Rev. Microbiol. 2017, 15, 132–133. [Google Scholar]
- Lopez, L.; Fasano, C.; Perrella, G.; Facella, P. Cryptochromes and the Circadian Clock: The Story of a Very Complex Relationship in a Spinningworld. Genes 2021, 12, 672. [Google Scholar] [CrossRef]
- Wu, L.; Luo, Y. Bacterial Quorum-Sensing Systems and Their Role in Intestinal Bacteria-Host Crosstalk. Front Microbiol. 2021, 12, 611413. [Google Scholar] [CrossRef]
- Fleitas Martínez, O.; Rigueiras, P.O.; da Pires, Á.S.; Porto, W.F.; Silva, O.N.; de la Fuente-Nunez, C.; Franco, O.L. Interference with Quorum-Sensing Signal Biosynthesis as a Promising Therapeutic Strategy Against Multidrug-Resistant Pathogens. Front Cell Infect. Microbiol. 2019, 8, 444. [Google Scholar] [CrossRef]
- Majumdar, S.; Pal, S. Cross-Species Communication in Bacterial World. J. Cell Commun. Signal 2017, 11, 187–190. [Google Scholar] [CrossRef] [PubMed]
- Matarèse, B.F.E.; Rahmoune, H.; Vo, N.T.K.; Seymour, C.B.; Schofield, P.N.; Mothersill, C. X-Ray-Induced Bio-Acoustic Emissions from Cultured Cells. Int. J. Radiat. Biol 2022, 99, 1285–1290. [Google Scholar] [CrossRef] [PubMed]
- Mothersill, C.; Smith, R.; Wang, J.; Rusin, A.; Fernandez-Palomo, C.; Fazzari, J.; Seymour, C. Biological Entanglement–Like Effect After Communication of Fish Prior to X-Ray Exposure. Dose-Response 2018, 16, 1559325817750067. [Google Scholar] [CrossRef]
- Smith, R.W.; Moccia, R.D.; Seymour, C.B.; Mothersill, C.E. Irradiation of Rainbow Trout at Early Life Stages Results in a Proteomic Legacy in Adult Gills. Part A; Proteomic Responses in the Irradiated Fish and in Non-Irradiated Bystander Fish. Environ. Res. 2018, 163, 297–306. [Google Scholar] [CrossRef] [PubMed]
- Van der Oost, R.; Beyer, J.; Vermeulen, N.P.E. Fish Bioaccumulation and Biomarkers in Environmental Risk Assessment: A Review. Environ. Toxicol. Pharmacol. 2003, 13, 57–149. [Google Scholar] [CrossRef] [PubMed]
- Schwarzenbach, R.P.; Escher, B.I.; Fenner, K.; Hofstetter, T.B.; Johnson, C.A.; Von Gunten, U.; Wehrli, B. The Challenge of Micropollutants in Aquatic Systems. Science 2006, 313, 1072–1077. [Google Scholar] [CrossRef]
- Zait, R.; Fighir, D.; Sluser, B.; Plavan, O.; Teodosiu, C. Priority Pollutants Effects on Aquatic Ecosystems Evaluated through Ecotoxicity, Impact, and Risk Assessments. Water 2022, 14, 3237. [Google Scholar] [CrossRef]
- Singh, S.; Sharma, P.; Pal, N.; Kumawat, M.; Shubham, S.; Sarma, D.K.; Tiwari, R.R.; Kumar, M.; Nagpal, R. Impact of Environmental Pollutants on Gut Microbiome and Mental Health via the Gut–Brain Axis. Microorganisms 2022, 10, 1457. [Google Scholar] [CrossRef]
- García-Peñarrubia, P.; Ruiz-Alcaraz, A.J.; Martínez-Esparza, M.; Marín, P.; Machado-Linde, F. Hypothetical Roadmap towards Endometriosis: Prenatal Endocrine-Disrupting Chemical Pollutant Exposure, Anogenital Distance, Gut-Genital Microbiota and Subclinical Infections. Hum. Reprod. Update 2020, 26, 246. [Google Scholar] [CrossRef]
- Toporova, L.; Balaguer, P. Nuclear Receptors Are the Major Targets of Endocrine Disrupting Chemicals. Mol. Cell Endocrinol. 2020, 502, 110665. [Google Scholar] [CrossRef]
- Latune, C.L.; Sinayskiy, I.; Petruccione, F. Apparent Temperature: Demystifying the Relation between Quantum Coherence, Correlations, and Heat Flows. Quantum Sci. Technol. 2019, 4, 025005. [Google Scholar] [CrossRef]
- Ishizaki, A.; Fleming, G.R. Theoretical Examination of Quantum Coherence in a Photosynthetic System at Physiological Temperature. Proc. Natl. Acad. Sci. USA 2009, 106, 17255–17260. [Google Scholar] [CrossRef] [PubMed]
- Lostaglio, M.; Korzekwa, K.; Jennings, D.; Rudolph, T. Quantum Coherence, Time-Translation Symmetry, and Thermodynamics. Phys. Rev. X 2015, 5, 021001. [Google Scholar] [CrossRef]
- Lipka-Bartosik, P.; Perarnau-Llobet, M.; Brunner, N. Operational Definition of the Temperature of a Quantum State. Phys. Rev. Lett. 2023, 130, 040401. [Google Scholar] [CrossRef]
- Ding, Y.S.; Yu, K.X.; Reta, D.; Ortu, F.; Winpenny, R.E.P.; Zheng, Y.Z.; Chilton, N.F. Field- and Temperature-Dependent Quantum Tunnelling of the Magnetisation in a Large Barrier Single-Molecule Magnet. Nat. Commun. 2018, 9, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Kamenetska, M.; Widawsky, J.R.; Dell’Angela, M.; Frei, M.; Venkataraman, L. Temperature Dependent Tunneling Conductance of Single Molecule Junctions. J. Chem. Phys. 2017, 146, 092311. [Google Scholar] [CrossRef]
- Weiss, U.; Grabert, H.; Linkwitz, S. Influence of Friction and Temperature on Coherent Quantum Tunneling. J. Low Temp. Phys. 1987, 68, 213–244. [Google Scholar] [CrossRef]
- Carvalho-Silva, V.H.; Coutinho, N.D.; Aquilanti, V. Temperature Dependence of Rate Processes beyond Arrhenius and Eyring: Activation and Transitivity. Front Chem. 2019, 7, 380. [Google Scholar] [CrossRef]
- Huang, Z. Quantum Coherence for an Atom Interacting with an Electromagnetic Field in the Background of Cosmic String Spacetime. Quantum Inf. Process 2020, 19, 1–11. [Google Scholar] [CrossRef]
- Wang, Y.T.; Tang, J.S.; Wei, Z.Y.; Yu, S.; Ke, Z.J.; Xu, X.Y.; Li, C.F.; Guo, G.C. Directly Measuring the Degree of Quantum Coherence Using Interference Fringes. Phys. Rev. Lett. 2017, 118, 020403. [Google Scholar] [CrossRef]
- Aleksandrov, I.A.; Di Piazza, A.; Plunien, G.; Shabaev, V.M. Stimulated Vacuum Emission and Photon Absorption in Strong Electromagnetic Fields. Phys. Rev. D 2022, 105, 116005. [Google Scholar] [CrossRef]
- Brodie, G. Energy Transfer from Electromagnetic Fields to Materials. In Electromagnetic Fields and Waves; Springer: Berlin/Heidelberg, Germany, 2019. [Google Scholar]
- Evlyukhin, A.B.; Tuz, V.R. Electromagnetic Scattering by Arbitrary-Shaped Magnetic Particles and Multipole Decomposition: Analytical and Numerical Approaches. Phys. Rev. B 2023, 107, 155425. [Google Scholar] [CrossRef]
- Mansuripur, M. A Tutorial on the Classical Theories of Electromagnetic Scattering and Diffraction. Nanophotonics 2020, 10, 315–342. [Google Scholar] [CrossRef]
- Truong, K.; Bradley, S.; Baginski, B.; Wilson, J.R.; Medlin, D.; Zheng, L.; Wilson, R.K.; Rusin, M.; Takacs, E.; Dean, D. The Effect of Well-Characterized, Very Low-Dose X-ray Radiation on Fibroblasts. PLoS ONE 2018, 13, e0190330. [Google Scholar] [CrossRef] [PubMed]
- Levitt, B.B.; Lai, H.C.; Manville, A.M. Low-Level EMF Effects on Wildlife and Plants: What Research Tells Us about an Ecosystem Approach. Front. Public Health 2022, 10, 1000840. [Google Scholar] [CrossRef] [PubMed]
- Bruneval, F.; Dattani, N.; van Setten, M.J. The GW Miracle in Many-Body Perturbation Theory for the Ionization Potential of Molecules. Front. Chem. 2021, 9, 749779. [Google Scholar] [CrossRef] [PubMed]
- Lin, J.C. Electromagnetic Fields in Biological Systems; Taylor & Francis Ltd: London, UK, 2016. [Google Scholar]
- Feyen, P.L.C.; Matarèse, B.F.E.; Urbano, L.; Abelha, T.F.; Rahmoune, H.; Green, M.; Dailey, L.A.; de Mello, J.C.; Benfenati, F. Photosensitized and Photothermal Stimulation of Cellular Membranes by Organic Thin Films and Nanoparticles. Front Bioeng. Biotechnol. 2022, 10, 932877. [Google Scholar] [CrossRef]
- Alizadeh, E.; Sanz, A.G.; García, G.; Sanche, L. Radiation Damage to DNA: The Indirect Effect of Low-Energy Electrons. J. Phys. Chem. Lett. 2013, 4, 820–825. [Google Scholar] [CrossRef]
- Kumar, A.; Becker, D.; Adhikary, A.; Sevilla, M.D. Reaction of Electrons with Dna: Radiation Damage to Radiosensitization. Int. J. Mol. Sci. 2019, 20, 3998. [Google Scholar] [CrossRef]
- Baiocco, G.; Bartzsch, S.; Conte, V.; Friedrich, T.; Jakob, B.; Tartas, A.; Villagrasa, C.; Prise, K.M. A Matter of Space: How the Spatial Heterogeneity in Energy Deposition Determines the Biological Outcome of Radiation Exposure. Radiat. Environ. Biophys. 2022, 61, 545–559. [Google Scholar] [CrossRef]
- Mothersill, C.; Seymour, C. Low Dose Radiation Mechanisms: The Certainty of Uncertainty. Mutat. Res. Genet. Toxicol. Environ. Mutagen. 2022, 876, 503451. [Google Scholar] [CrossRef]
- Joiner, M.C.; Marples, B.; Lambin, P.; Short, S.C.; Turesson, I. Low-Dose Hypersensitivity: Current Status and Possible Mechanisms. Proc. Int. J. Radiat. Oncol. Biol. Phys. 2001, 49, 379–389. [Google Scholar] [CrossRef] [PubMed]
- Fernandez-Palomo, C.; Seymour, C.; Mothersill, C. Inter-Relationship between Low-Dose Hyper-Radiosensitivity and Radiation-Induced Bystander Effects in the Human T98G Glioma and the Epithelial HaCaT Cell Line. Radiat. Res. 2016, 185, 124–133. [Google Scholar] [CrossRef] [PubMed]
- Hanahan, D.; Weinberg, R.A. Hallmarks of Cancer: The next Generation. Cell 2011, 144, 646–674. [Google Scholar] [CrossRef]
- Clutton, S.M.; Townsend, K.M.S.; Walker, C.; Ansell, J.D.; Wright, E.G. Radiation-Induced Genomic Instability and Persisting Oxidative Stress in Primary Bone Marrow Cultures. Carcinogenesis 1996, 17, 1633–1639. [Google Scholar] [CrossRef] [PubMed]
- Neriishi, K.; Nakashima, E.; Delongchamps, R.R. Persistent Subclinical Inflammation among A-Bomb Survivors. Int. J. Radiat. Biol. 2001, 77, 475–482. [Google Scholar] [CrossRef] [PubMed]
- Hayashi, T.; Kusunoki, Y.; Hakoda, M.; Morishita, Y.; Kubo, Y.; Maki, M.; Kasagi, F.; Kodama, K.; MacPhee, D.G.; Kyoizumi, S. Radiation Dose-Dependent Increases in Inflammatory Response Markers in A-Bomb Survivors. Int. J. Radiat. Biol. 2003, 79, 129–136. [Google Scholar] [CrossRef]
- Lorimore, S.A.; Wright, E.G. Radiation-Induced Genomic Instability and Bystander Effects: Related Inflammatory-Type Responses to Radiation-Induced Stress and Injury? A Review. Int. J. Radiat. Biol. 2003, 79, 15–25. [Google Scholar] [CrossRef]
- Lorimore, S.A.; Mukherjee, D.; Robinson, J.I.; Chrystal, J.A.; Wright, E.G. Long-Lived Inflammatory Signaling in Irradiated Bone Marrow Is Genome Dependent. Cancer Res. 2011, 71, 6485–6491. [Google Scholar] [CrossRef]
- Mukherjee, D.; Coates, P.J.; Lorimore, S.A.; Wright, E.G. Responses to Ionizing Radiation Mediated by Inflammatory Mechanisms. J. Pathol. 2014, 232, 289–299. [Google Scholar] [CrossRef]
- Frey, B.; Hehlgans, S.; Rödel, F.; Gaipl, U.S. Modulation of Inflammation by Low and High Doses of Ionizing Radiation: Implications for Benign and Malign Diseases. Cancer Lett. 2015, 368, 230–237. [Google Scholar] [CrossRef] [PubMed]
- Dawood, A.; Mothersill, C.; Seymour, C. Low Dose Ionizing Radiation and the Immune Response: What Is the Role of Non-Targeted Effects? Int. J. Radiat. Biol. 2021, 97, 1368–1382. [Google Scholar] [CrossRef] [PubMed]
- Widel, M.; Krzywon, A.; Gajda, K.; Skonieczna, M.; Rzeszowska-Wolny, J. Induction of Bystander Effects by UVA, UVB, and UVC Radiation in Human Fibroblasts and the Implication of Reactive Oxygen Species. Free Radic. Biol. Med. 2014, 68, 278–287. [Google Scholar] [CrossRef] [PubMed]
- Whiteside, J.R.; Allinson, S.L.; McMillan, T.J. Timeframes of UVA-Induced Bystander Effects in Human Keratinocytes. Photochem. Photobiol. 2011, 87, 435–440. [Google Scholar] [CrossRef]
- Zhang, J.; Wang, X.; Vikash, V.; Ye, Q.; Wu, D.; Liu, Y.; Dong, W. ROS and ROS-Mediated Cellular Signaling. Oxid. Med. Cell Longev. 2016, 2016, 4350965. [Google Scholar] [CrossRef]
- Sinenko, S.A.; Starkova, T.Y.; Kuzmin, A.A.; Tomilin, A.N. Physiological Signaling Functions of Reactive Oxygen Species in Stem Cells: From Flies to Man. Front Cell Dev. Biol. 2021, 9, 714370. [Google Scholar] [CrossRef]
- Tsubata, T. Involvement of Reactive Oxygen Species (ROS) in BCR Signaling as a Second Messenger. In Advances in Experimental Medicine and Biology; Springer: Berlin/Heidelberg, Germany, 2020; Volume 1254. [Google Scholar]
- Levy, H.M.; Schneider, A.; Tiwari, S.; Zer, H.; Yochelis, S.; Goloubinoff, P.; Keren, N.; Paltiel, Y. The Effect of Spin Exchange Interaction on Protein Structural Stability. Phys. Chem. Chem. Phys. 2022, 24, 29176–29185. [Google Scholar] [CrossRef]
- Naaman, R.; Paltiel, Y.; Waldeck, D.H. Chiral Molecules and the Electron Spin. Nat. Rev. Chem. 2019, 3, 250–260. [Google Scholar] [CrossRef]
- Cataldo, F. Radiation-Induced Racemization and Amplification of Chirality: Implications for Comets and Meteorites. Int. J. Astrobiol. 2007, 6, 1–10. [Google Scholar] [CrossRef]
- Xu, Y.; Mi, W. Chiral-Induced Spin Selectivity in Biomolecules, Hybrid Organic-Inorganic Perovskites and Inorganic Materials: A Comprehensive Review on Recent Progress. Mater. Horiz. 2023, 10, 1924–1955. [Google Scholar] [CrossRef]
- Michaeli, K.; Kantor-Uriel, N.; Naaman, R.; Waldeck, D.H. The Electron’s Spin and Molecular Chirality-How Are They Related and How Do They Affect Life Processes? Chem. Soc. Rev. 2016, 45, 6478–6487. [Google Scholar] [CrossRef] [PubMed]
- Silver, B.B.; Nelson, C.M. The Bioelectric Code: Reprogramming Cancer and Aging from the Interface of Mechanical and Chemical Microenvironments. Front. Cell Dev. Biol. 2018, 6, 21. [Google Scholar] [CrossRef] [PubMed]
- Naaman, R.; Waldeck, D.H.; Paltiel, Y. Chiral Molecules-Ferromagnetic Interfaces, an Approach towards Spin Controlled Interactions. Appl. Phys. Lett. 2019, 115, 133701. [Google Scholar] [CrossRef]
- Slussarenko, S.; Pryde, G.J. Photonic Quantum Information Processing: A Concise Review. Appl. Phys. Rev. 2019, 6, 041303. [Google Scholar] [CrossRef]
- Rezaie, J.; Feghhi, M.; Etemadi, T. A Review on Exosomes Application in Clinical Trials: Perspective, Questions, and Challenges. Cell Commun. Signal. 2022, 20, 1–13. [Google Scholar] [CrossRef]
- Gurung, S.; Perocheau, D.; Touramanidou, L.; Baruteau, J. The Exosome Journey: From Biogenesis to Uptake and Intracellular Signalling. Cell Commun. Signal. 2021, 19, 1–19. [Google Scholar] [CrossRef]
- De Almeida, A.J.P.O.; De Oliveira, J.C.P.L.; Da Silva Pontes, L.V.; De Souza Júnior, J.F.; Gonçalves, T.A.F.; Dantas, S.H.; De Almeida Feitosa, M.S.; Silva, A.O.; De Medeiros, I.A. ROS: Basic Concepts, Sources, Cellular Signaling, and Its Implications in Aging Pathways. Oxid. Med. Cell Longev. 2022, 2022, 1–23. [Google Scholar] [CrossRef]
- D’Autréaux, B.; Toledano, M.B. ROS as Signalling Molecules: Mechanisms That Generate Specificity in ROS Homeostasis. Nat. Rev. Mol. Cell Biol. 2007, 8, 813–824. [Google Scholar] [CrossRef] [PubMed]
- Kumar, R.R.; Arora, K.; Goswami, S.; Sakhare, A.; Singh, B.; Chinnusamy, V.; Praveen, S. MAPK Enzymes: A ROS Activated Signaling Sensors Involved in Modulating Heat Stress Response, Tolerance and Grain Stability of Wheat under Heat Stress. 3 Biotech. 2020, 10, 1–11. [Google Scholar] [CrossRef]
- Qaswal, A.B. Quantum Tunneling of Ions through the Closed Voltage-Gated Channels of the Biological Membrane: A Mathematical Model and Implications. Quantum Rep. 2019, 1, 219–225. [Google Scholar] [CrossRef]
- Ferraro, A. Promising Ways to Encode and Manipulate Quantum Information. Nature 2019, 566, 460–462. [Google Scholar] [CrossRef]
- Hübner, C.A.; Jentsch, T.J. Ion Channel Diseases. Hum. Mol. Genet. 2002, 11, 2435–2445. [Google Scholar] [CrossRef] [PubMed]
- Joukar, S. A Comparative Review on Heart Ion Channels, Action Potentials and Electrocardiogram in Rodents and Human: Extrapolation of Experimental Insights to Clinic. Lab. Anim. Res. 2021, 37, 1–15. [Google Scholar] [CrossRef]
- Sakmann, B. Sparks in the Brain: The Story of Ion Channels and Nerve Cells. Front. Young Minds 2022, 10, 858193. [Google Scholar] [CrossRef]
- Chen, L.; Hassani Nia, F.; Stauber, T. Ion Channels and Transporters in Muscle Cell Differentiation. Int. J. Mol. Sci. 2021, 22, 13615. [Google Scholar] [CrossRef] [PubMed]
- Seifi, M.; Soltanmanesh, A.; Shafiee, A. Quantum Coherence on Selectivity and Transport of Ion Channels. Sci. Rep. 2022, 12, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Al-Rawashdeh, B.M.; Qaswal, A.B.; Suleiman, A.; Zayed, F.M.; Al-Rawashdeh, S.M.; Tawalbeh, M.; Khreesha, L.; Alzubaidi, A.; Al-Zubidi, E.; Ghala, Z.; et al. The Quantum Tunneling of Ions Model Can Explain the Pathophysiology of Tinnitus. Brain Sci. 2022, 12, 426. [Google Scholar] [CrossRef] [PubMed]
- Lambert, C.; Zappia, J.; Sanchez, C.; Florin, A.; Dubuc, J.E.; Henrotin, Y. The Damage-Associated Molecular Patterns (DAMPs) as Potential Targets to Treat Osteoarthritis: Perspectives from a Review of the Literature. Front. Med. 2021, 7, 607186. [Google Scholar] [CrossRef]
- Murao, A.; Aziz, M.; Wang, H.; Brenner, M.; Wang, P. Release Mechanisms of Major DAMPs. Apoptosis 2021, 26, 152–162. [Google Scholar] [CrossRef]
- Arakawa, Y.; Holmes, M.J. Progress in Quantum-Dot Single Photon Sources for Quantum Information Technologies: A Broad Spectrum Overview. Appl. Phys. Rev. 2020, 7, 021309. [Google Scholar] [CrossRef]
- Li, X.; Jones, A.C.; Choi, J.; Zhao, H.; Chandrasekaran, V.; Pettes, M.T.; Piryatinski, A.; Tschudin, M.A.; Reiser, P.; Broadway, D.A.; et al. Proximity-Induced Chiral Quantum Light Generation in Strain-Engineered WSe2/NiPS3 Heterostructures. Nat. Mater. 2023, 1–6. [Google Scholar] [CrossRef]
- Kumar, A.; Capua, E.; Kesharwani, M.K.; Martin, J.M.L.; Sitbon, E.; Waldeck, D.H.; Naaman, R. Chirality-Induced Spin Polarization Places Symmetry Constraints on Biomolecular Interactions. Proc. Natl. Acad. Sci. USA 2017, 114, 2474–2478. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.; Yang, Q.; Chen, Y.; Liu, W. Extremize Optical Chiralities through Polarization Singularities. Phys. Rev. Lett. 2021, 126, 253901. [Google Scholar] [CrossRef] [PubMed]
- Benfatto, M.; Pace, E.; Curceanu, C.; Scordo, A.; Clozza, A.; Davoli, I.; Lucci, M.; Francini, R.; De Matteis, F.; Grandi, M.; et al. Biophotons: Low Signal/Noise Ratio Reveals Crucial Events. BioRxiv 2019, 558353. [Google Scholar] [CrossRef]
- Mothersill, C.; Le, M.; Rusin, A.; Seymour, C. Biophotons in Radiobiology: Inhibitors, Communicators and Reactors. Radiat. Prot. Dosimetry 2019, 183, 136–141. [Google Scholar] [CrossRef]
- Ma, Z.; Zhang, Z.; Dai, Y.; Dong, Y.; Zhang, C. Detecting and Estimating Coherence Based on Coherence Witnesses. Phys. Rev. A 2021, 103, 012409. [Google Scholar] [CrossRef]
- Wu, K.; Da Streltsov, A.; Regula, B.; Xiang, G.Y.; Li, C.F.; Guo, G.C. Experimental Progress on Quantum Coherence: Detection, Quantification, and Manipulation. Adv. Quantum Technol. 2021, 4, 2100040. [Google Scholar] [CrossRef]
- Basiri-Esfahani, S.; Shahandeh, F. Locally Incoherent Witnessing of Quantum Coherence. Phys. Rev. A 2022, 106, 022410. [Google Scholar] [CrossRef]
- Marcus, M.; Knee, G.C.; Datta, A. Towards a Spectroscopic Protocol for Unambiguous Detection of Quantum Coherence in Excitonic Energy Transport. Faraday Discuss 2019, 221, 110–132. [Google Scholar] [CrossRef]
- Yi, Z.; Begzjav, T.K.; Ariunbold, G.O.; Zheltikov, A.M.; Sokolov, A.V.; Scully, M.O. Multiple Pathway Quantum Beat Spectroscopy. Front. Phys. 2022, 10, 921499. [Google Scholar] [CrossRef]
- Summhammer, J.; Salari, V.; Bernroider, G. A Quantum-Mechanical Description of Ion Motion within the Confining Potentials of Voltage-Gated Ion Channels. J. Integr. Neurosci. 2012, 11, 123–135. [Google Scholar] [CrossRef] [PubMed]
- Cao, Q.Y.; Yang, P.C.; Gong, M.S.; Yu, M.; Retzker, A.; Plenio, M.B.; Müller, C.; Tomek, N.; Naydenov, B.; McGuinness, L.P.; et al. Protecting Quantum Spin Coherence of Nanodiamonds in Living Cells. Phys. Rev. Appl. 2020, 13, 024021. [Google Scholar] [CrossRef]
- Maqbool, M. Interaction of Gamma Rays and X-Rays with Matter. In An Introduction to Medical Physics; Springer: Cham, Switzerland, 2017; pp. 43–61. [Google Scholar]
- Vítor, A.C.; Huertas, P.; Legube, G.; de Almeida, S.F. Studying DNA Double-Strand Break Repair: An Ever-Growing Toolbox. Front Mol. Biosci. 2020, 7, 24. [Google Scholar] [CrossRef]
- Zhang, X.; Ye, C.; Sun, F.; Wei, W.; Hu, B.; Wang, J. Both Complexity and Location of DNA Damage Contribute to Cellular Senescence Induced by Ionizing Radiation. PLoS ONE 2016, 11, e0155725. [Google Scholar] [CrossRef] [PubMed]
- Smith, T.A.; Kirkpatrick, D.R.; Smith, S.; Smith, T.K.; Pearson, T.; Kailasam, A.; Herrmann, K.Z.; Schubert, J.; Agrawal, D.K. Radioprotective Agents to Prevent Cellular Damage Due to Ionizing Radiation. J. Transl. Med. 2017, 15, 1–18. [Google Scholar] [CrossRef]
- Hu, C.; Zuo, H.; Li, Y. Effects of Radiofrequency Electromagnetic Radiation on Neurotransmitters in the Brain. Front Public Health 2021, 9, 1139. [Google Scholar] [CrossRef]
- Little, M.P.; Azizova, T.V.; Richardson, D.B.; Tapio, S.; Bernier, M.O.; Kreuzer, M.; Cucinotta, F.A.; Bazyka, D.; Chumak, V.; Ivanov, V.K.; et al. Ionising Radiation and Cardiovascular Disease: Systematic Review and Meta-Analysis. BMJ 2023, 380, e072924. [Google Scholar] [CrossRef]
- Belli, M.; Tabocchini, M.A. Ionizing Radiation-Induced Epigenetic Modifications and Their Relevance to Radiation Protection. Int. J. Mol. Sci. 2020, 21, 5993. [Google Scholar] [CrossRef]
- Shuryak, I. Quantitative Modeling of Responses to Chronic Ionizing Radiation Exposure Using Targeted and Non-Targeted Effects. PLoS ONE 2017, 12, e0176476. [Google Scholar] [CrossRef]
- Modic, D.; Anderson, R.; Palomäki, J. We Will Make You like Our Research: The Development of a Susceptibility-to-Persuasion Scale. PLoS ONE 2018, 13, e0194119. [Google Scholar] [CrossRef]
- Doniach, S. Biological Physics: Energy, Information, Life. Phys Today 2004, 57, 63–64. [Google Scholar] [CrossRef]
- Adams, B.; Petruccione, F. Quantum Effects in the Brain: A Review. AVS Quantum Sci. 2020, 2, 022901. [Google Scholar] [CrossRef]
- Frixione, E. Consciousness and Neuronal Microtubules: The Penrose-Hameroff Quantum Model in Retrospect. In History, Philosophy and Theory of the Life Sciences; Springer: Berlin/Heidelberg, Germany, 2014; Volume 6. [Google Scholar]
- Hameroff, S.; Penrose, R. Orchestrated Reduction of Quantum Coherence in Brain Microtubules: A Model for Consciousness. Math. Comput. Simul. 1996, 40, 453–480. [Google Scholar] [CrossRef]
- Hameroff, S.; Nip, A.; Porter, M.; Tuszynski, J. Conduction Pathways in Microtubules, Biological Quantum Computation, and Consciousness. BioSystems 2002, 64, 149–168. [Google Scholar] [CrossRef] [PubMed]
- Hagan, S.; Hameroff, S.R.; Tuszyński, J.A. Quantum Computation in Brain Microtubules: Decoherence and Biological Feasibility. Phys. Rev. E Stat. Phys. Plasmas Fluids Relat. Interdiscip. Top. 2002, 65, 061901. [Google Scholar] [CrossRef] [PubMed]
- Dresp-Langley, B. Consciousness Beyond Neural Fields: Expanding the Possibilities of What Has Not Yet Happened. Front. Psychol. 2022, 12, 762349. [Google Scholar] [CrossRef] [PubMed]
- Baars, B.J.; Edelman, D.B. Consciousness, Biology and Quantum Hypotheses. Phys. Life Rev. 2012, 9, 285–294. [Google Scholar] [CrossRef] [PubMed]
- Jiang, C.; Yu, Z.W.; Hu, X.L.; Wang, X.B. Robust Twin-Field Quantum Key Distribution through Sending or Not Sending. Natl. Sci. Rev. 2023, 10, nwac186. [Google Scholar] [CrossRef]
- Curty, M.; Azuma, K.; Lo, H.K. Simple Security Proof of Twin-Field Type Quantum Key Distribution Protocol. npj Quantum Inf. 2019, 5, 64. [Google Scholar] [CrossRef]
- Li, H.; Zhang, C. Alternative Schemes for Twin-Field Quantum Key Distribution with Discrete-Phase-Randomized Sources. Front. Phys. 2023, 11, 132. [Google Scholar] [CrossRef]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Matarèse, B.F.E.; Rusin, A.; Seymour, C.; Mothersill, C. Quantum Biology and the Potential Role of Entanglement and Tunneling in Non-Targeted Effects of Ionizing Radiation: A Review and Proposed Model. Int. J. Mol. Sci. 2023, 24, 16464. https://doi.org/10.3390/ijms242216464
Matarèse BFE, Rusin A, Seymour C, Mothersill C. Quantum Biology and the Potential Role of Entanglement and Tunneling in Non-Targeted Effects of Ionizing Radiation: A Review and Proposed Model. International Journal of Molecular Sciences. 2023; 24(22):16464. https://doi.org/10.3390/ijms242216464
Chicago/Turabian StyleMatarèse, Bruno F. E., Andrej Rusin, Colin Seymour, and Carmel Mothersill. 2023. "Quantum Biology and the Potential Role of Entanglement and Tunneling in Non-Targeted Effects of Ionizing Radiation: A Review and Proposed Model" International Journal of Molecular Sciences 24, no. 22: 16464. https://doi.org/10.3390/ijms242216464