
Global Analysis of Lysine Lactylation of Germinated Seeds in Wheat
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Global Profiling of Proteome in Embryos of Wheat at Different Hours after Imbibition
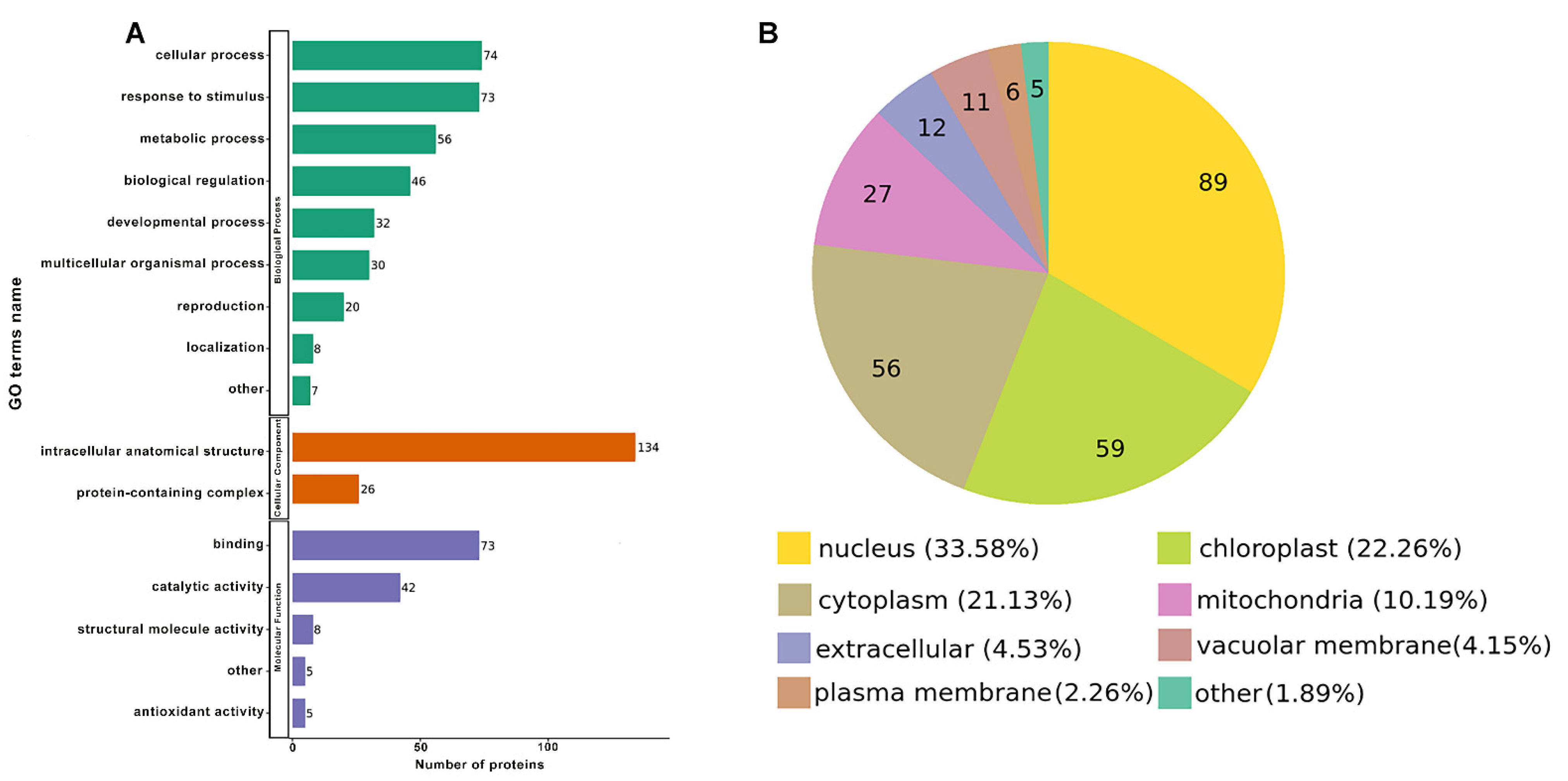
2.2. Functional Enrichment Analysis of the DEPs
2.3. Proteome-Wide Analysis of Lysine Lactylation Sites and Proteins in Germinated Seeds of Wheat
2.4. Characterization of DLPs in Wheat Germinated Seeds
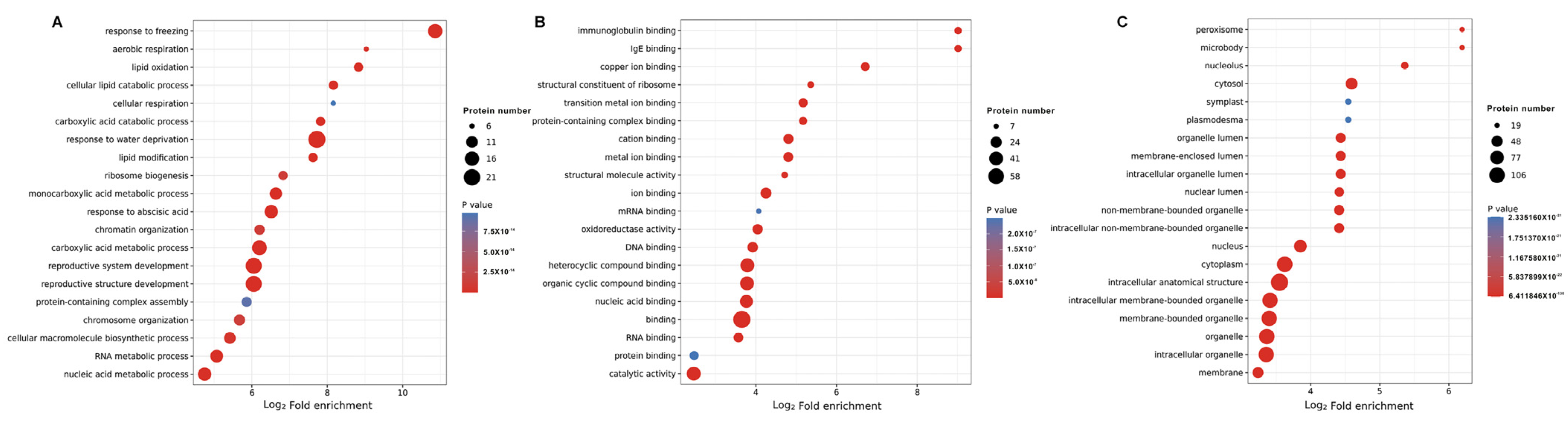
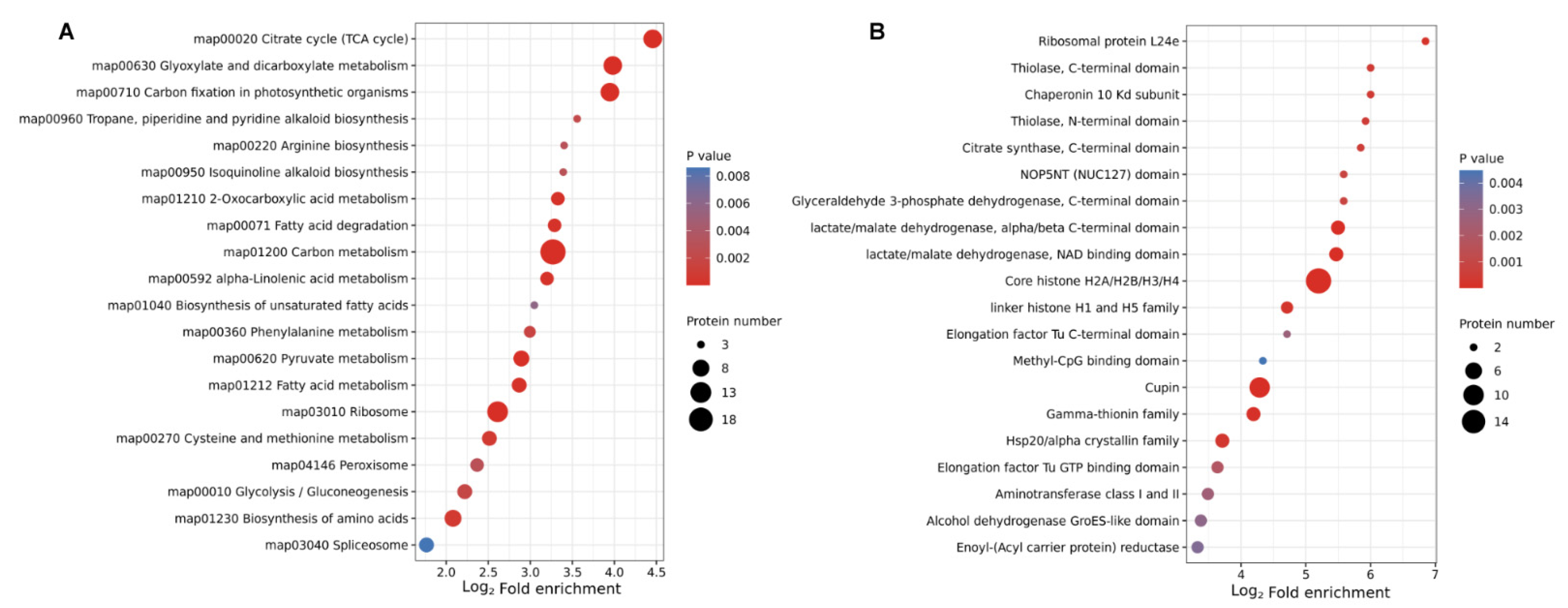
2.5. Functional Enrichment Analysis of DLPs
2.6. Motif Analysis of Lysine Lactylated Peptides
3. Discussion
4. Materials and Methods
4.1. Plant Material and Growth Conditions
4.2. Protein Extraction and Western Blotting
4.3. Trypsin Digestion
4.4. TMT Labelling
4.5. Affinity Enrichment of Lys-lactylated Peptides
4.6. HPLC-MS/MS Analysis
4.7. Database Search
4.8. Bioinformatics Analyses
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Zhang, D.; Tang, Z.; Huang, H.; Zhou, G.; Cui, C.; Weng, Y.; Liu, W.; Kim, S.; Lee, S.; Perez-Neut, M.; et al. Metabolic regulation of gene expression by histone lactylation. Nature 2019, 574, 575–580. [Google Scholar] [CrossRef] [PubMed]
- Cui, H.; Xie, N.; Banerjee, S.; Ge, J.; Jiang, D.; Dey, T.; Matthews, Q.; Liu, R.M.; Liu, G. Lung Myofibroblast Promote Macrophage Pro-fibrotic Activity through Lactate-induced Histone Lactylation. Am. J. Respir. Cell Mol. 2021, 64, 115–125. [Google Scholar] [CrossRef] [PubMed]
- Meng, X.; Baine, J.; Yan, T.; Wang, S. Comprehensive Analysis of Lysine Lactylation in Rice (Oryza sativa) Grains. J. Agric. Food Chem. 2021, 69, 8287–8297. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Ouyang, L.; Wei, L. Novel Insight of Nitrogen Deprivation Affected Lipid Accumulation by Genome-Wide Lactylation in Nannochloropsis oceanica. J. Agric. Food Chem. 2023, 71, 10107–10123. [Google Scholar] [CrossRef] [PubMed]
- Wen, J.; Qin, Z.; Sun, L.; Zhang, Y.; Wang, D.; Peng, H.; Yao, Y.; Hu, Z.; Ni, Z.; Sun, Q.; et al. Alternative splicing of TaHSFA6e modulates heat shock protein–mediated translational regulation in response to heat stress in wheat. New Phytol. 2023, 239, 2235–2247. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Song, L.; Liang, W.; Mu, P.; Wang, S.; Lin, Q. Comprehensive profiling of lysine acetylproteome analysis reveals diverse functions of lysine acetylation in common wheat. Sci. Rep. 2016, 6, 21069. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Wang, G.; Song, L.; Mu, P.; Lin, Q. Global analysis of protein lysine succinylation profiles in common wheat. BMC Genom. 2017, 18, 309. [Google Scholar] [CrossRef]
- Fu, Z.; Jin, X.; Ding, D.; Li, Y.; Fu, Z.; Tang, J. Proteomic analysis of heterosis during maize seed germination. Proteomics 2011, 11, 1462–1472. [Google Scholar] [CrossRef]
- Guo, W.; Han, L.; Li, X.; Wang, H.; Mu, P.; Lin, Q.; Liu, Q.; Zhang, Y. Proteome and lysine acetylome analysis reveals insights into the molecular mechanism of seed germination in wheat. Sci. Rep. 2020, 10, 13454. [Google Scholar] [CrossRef]
- Meng, X.; Xing, S.; Perez, L.M.; Peng, X.; Zhao, Q.; Redoña, E.D.; Wang, C.; Peng, Z. Proteome-wide analysis of lysine 2-hydroxyisobutyrylation in developing rice (Oryza sativa) seeds. Sci. Rep. 2017, 7, 17486. [Google Scholar] [CrossRef]
- Meng, X.; Lv, Y.; Mujahid, H.; Edelmann, M.J.; Zhao, H.; Peng, X.; Peng, Z. Proteome-wide lysine acetylation identification in developing rice (Oryza sativa) seeds and protein co-modification by acetylation, succinylation, ubiquitination, and phosphorylation. Biochim. Biophys. Acta Proteins Proteom. 2018, 1866, 451–463. [Google Scholar] [CrossRef] [PubMed]
- Meng, X.; Mujahid, H.; Zhang, Y.; Peng, X.; Redoña, E.D.; Wang, C.; Peng, Z. Comprehensive analysis of the lysine succinylome and protein co-modifications in developing rice seeds. Mol. Cell Proteom. 2019, 18, 2359–2372. [Google Scholar] [CrossRef]
- Mujahid, H.; Meng, X.; Xing, S.; Peng, X.; Wang, C.; Peng, Z. Malonylome analysis in developing rice (Oryza sativa) seeds suggesting that protein lysine malonylation is well-conserved and overlaps with acetylation and succinylation substantially. J. Proteom. 2018, 170, 88–98. [Google Scholar] [CrossRef] [PubMed]
- Hagihara, H.; Shoji, H.; Otabi, H.; Toyoda, A.; Katoh, K.; Namihira, M.; Miyakawa, T. Protein lactylation induced by neural excitation. Cell Rep. 2021, 37, 109820. [Google Scholar] [CrossRef] [PubMed]
- Gao, M.; Zhang, N.; Liang, W.J. Systematic Analysis of Lysine Lactylation in the Plant Fungal Pathogen Botrytis cinerea. Front. Microbiol. 2020, 11, 594743. [Google Scholar] [CrossRef] [PubMed]
- IWGSC. A chromosome-based draft sequence of the hexaploid bread wheat (Triticum aestivum L.) genome. Science 2014, 345, 1251788. [Google Scholar] [CrossRef] [PubMed]
- Nallamilli, B.; Edelmann, M.; Zhong, X.; Tan, F.; Mujahid, H.; Zhang, J.; Nanduri, B.; Peng, Z. Global analysis of lysine acetylation suggests the involvement of protein acetylation in diverse biological processes in rice (Oryza sativa). PLoS ONE 2014, 9, e89283. [Google Scholar] [CrossRef] [PubMed]
- Finnie, C.; Melchior, S.; Roepstorff, P.; Svensson, B. Proteome analysis of grain flling and seed maturation in barley. Plant Physiol. 2002, 129, 1308–1319. [Google Scholar] [CrossRef]
- Wang, W.; Vinocur, B.; Shoseyov, O.; Altman, A. Role of plant heat-shock proteins and molecular chaperones in the abiotic stress response. Trends Plant Sci. 2004, 9, 244–252. [Google Scholar] [CrossRef]
- Kaur, A.; Gupta, O.; Sharma, P. Comparative expression analysis of HSP genes in wheat and barley under heat stress. J. Wheat Res. 2016, 8, 63–64. [Google Scholar]
- Chitteti, B.; Tan, F.; Mujahid, H.; Magee, B.; Bridges, S.; Peng, Z. Comparative analysis of proteome differential regulation during cell dedifferentiation in Arabidopsis. Proteomics 2008, 8, 4303–4316. [Google Scholar] [CrossRef] [PubMed]
- Xie, L.; Liu, W.; Li, Q.; Chen, S.; Xu, M.; Huang, Q.; Zeng, J.; Zhou, M.; Xie, J. First succinyl-proteome profiling of extensively drug-resistant Mycobacterium tuberculosis revealed involvement of succinylation in cellular physiology. J Proteome Res. 2015, 14, 107–119. [Google Scholar] [CrossRef] [PubMed]
- Qi, T.; Li, J.; Wang, H.; Han, X.; Li, J.; Du, J. Global analysis of protein lysine 2-hydroxyisobutyrylation (Khib) profiles in Chinese herb rhubarb (Dahuang). BMC Genom. 2021, 22, 542. [Google Scholar] [CrossRef] [PubMed]
- He, D.; Wang, Q.; Li, M.; Damaris, R.; Yi, X.; Cheng, Z.; Yang, P. Global proteome analyses of lysine acetylation and succinylation reveal the widespread involvement of both modifcation in metabolism in the embryo of germinating rice seed. J. Proteome Res. 2016, 15, 879–890. [Google Scholar] [CrossRef] [PubMed]
- Guo, W.; Han, J.; Li, X.; He, Z.; Zhang, Y. Large-scale analysis of protein crotonylation reveals its diverse functions in Pinellia ternate. BMC Plant Biol. 2022, 22, 457. [Google Scholar] [CrossRef] [PubMed]
- Zhou, S.; Yang, Q.; Yin, C.; Liu, L.; Liang, W. Systematic analysis of the lysine acetylome in Fusarium graminearum. BMC Genom. 2016, 17, 1019. [Google Scholar] [CrossRef]








Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhu, J.; Guo, W.; Lan, Y. Global Analysis of Lysine Lactylation of Germinated Seeds in Wheat. Int. J. Mol. Sci. 2023, 24, 16195. https://doi.org/10.3390/ijms242216195
Zhu J, Guo W, Lan Y. Global Analysis of Lysine Lactylation of Germinated Seeds in Wheat. International Journal of Molecular Sciences. 2023; 24(22):16195. https://doi.org/10.3390/ijms242216195
Chicago/Turabian StyleZhu, Junke, Weiwei Guo, and Yubin Lan. 2023. "Global Analysis of Lysine Lactylation of Germinated Seeds in Wheat" International Journal of Molecular Sciences 24, no. 22: 16195. https://doi.org/10.3390/ijms242216195