
Characterization of the FLA Gene Family in Tomato (Solanum lycopersicum L.) and the Expression Analysis of SlFLAs in Response to Hormone and Abiotic Stresses

 and
and 
Abstract
:1. Introduction
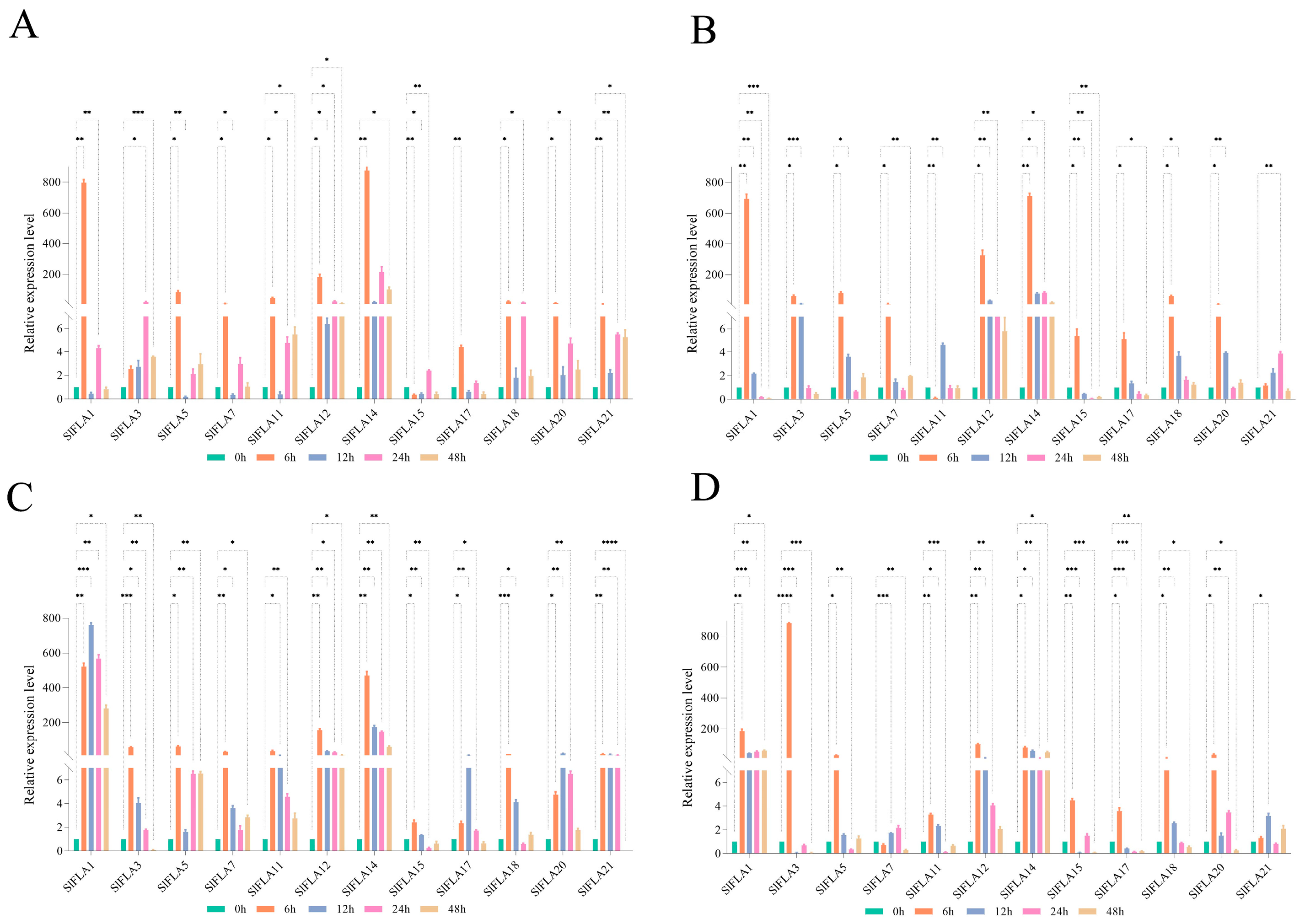
2. Results
2.1. Identification of FLA Genes in Tomato
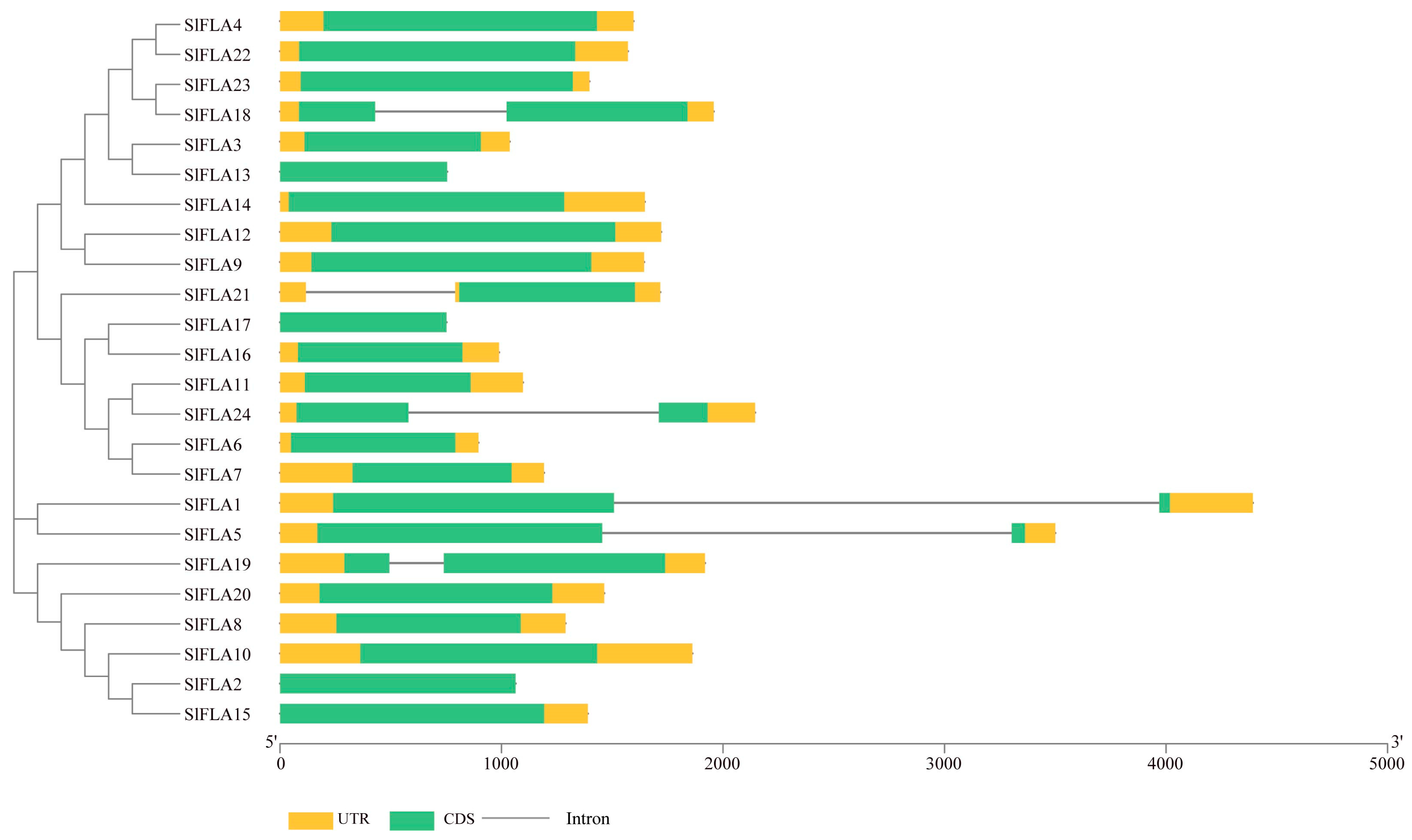
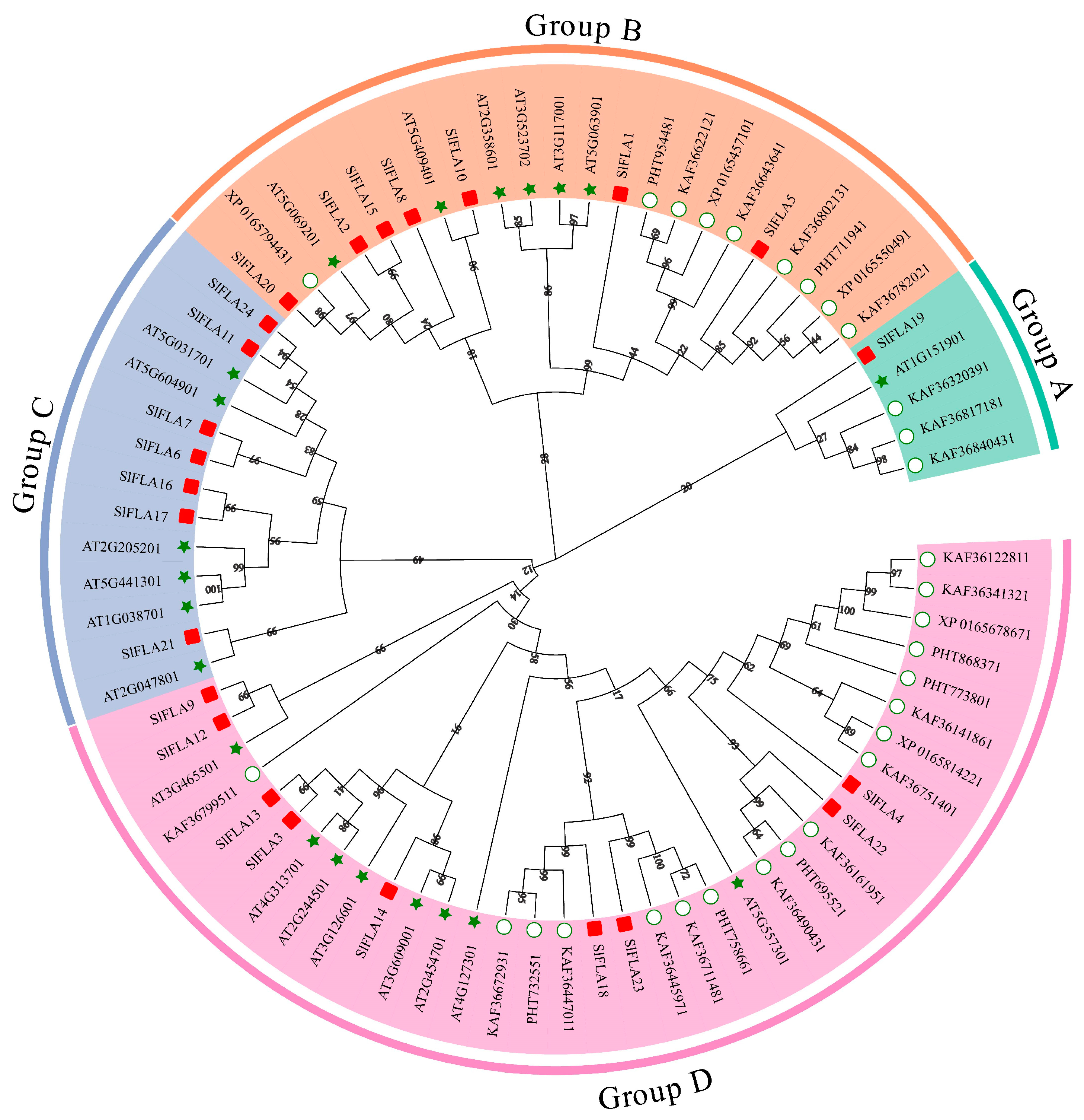
2.2. Structural Analysis and Phylogenetic Tree Analysis of the SlFLA Gene Family
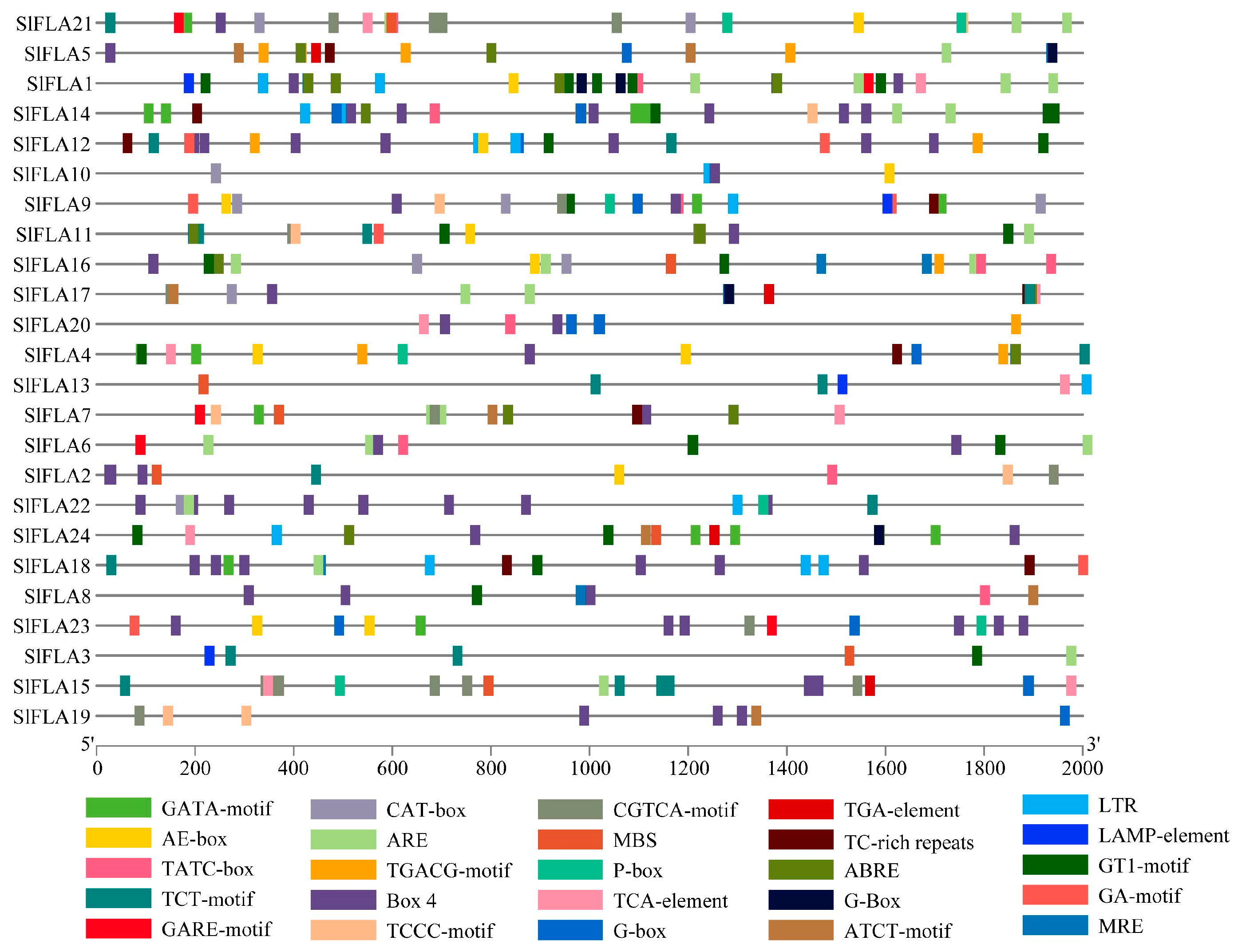
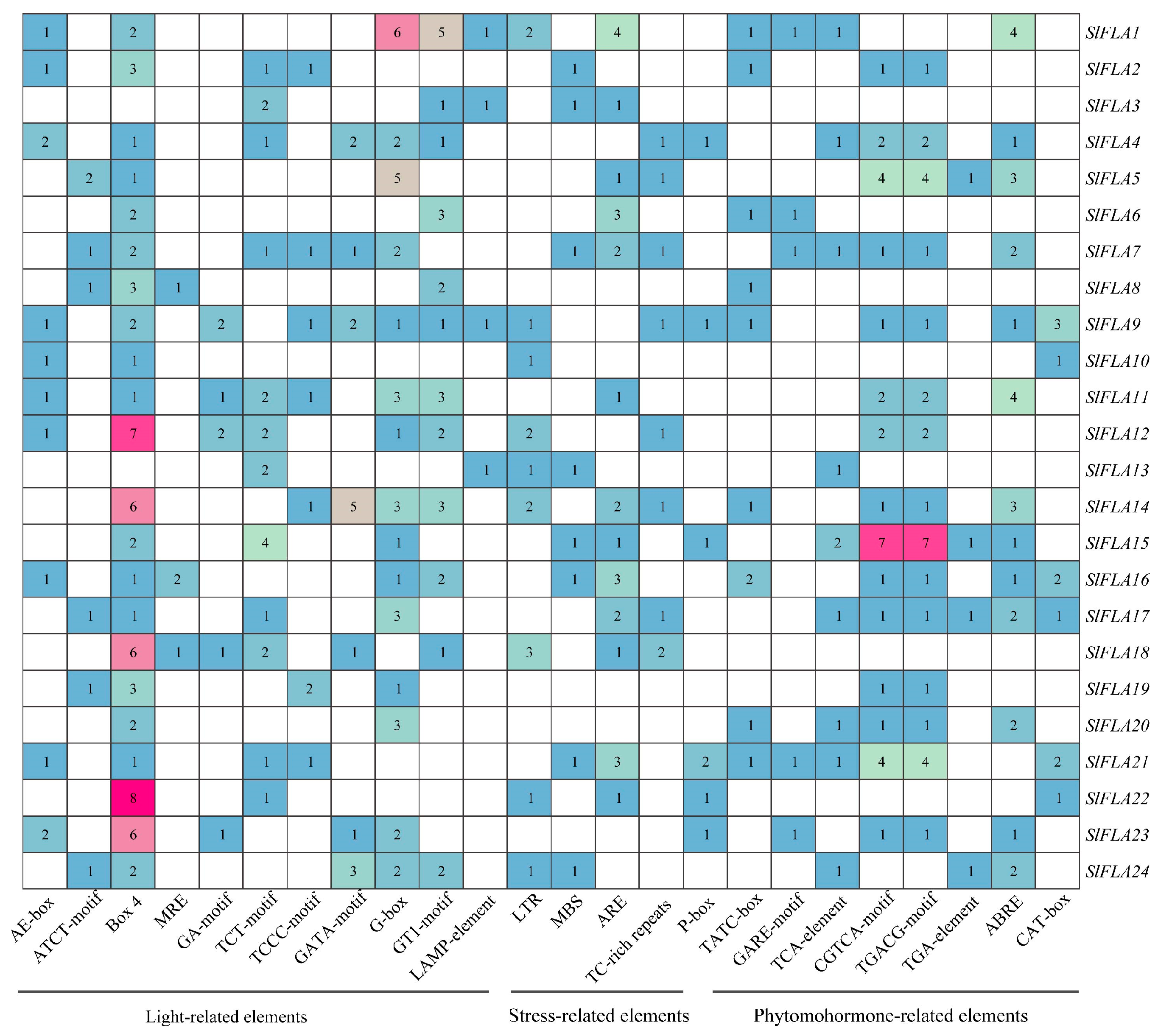
2.3. Analysis of Cis-Acting Elements of Tomato FLA Family Genes
2.4. Conserved Domain and Conserved Motifs of the Tomato FLA family
2.5. Analysis of the Protein Secondary Structure of Tomato FLA Family Genes
2.6. Tissue-Specific Expression Pattern of Tomato FLA Genes
2.7. Expression of Tomato FLA Genes in Response to MeJA and ABA Treatments
2.8. Expression Profile Analysis of FLA Genes in Tomato under NaCl, Dark, Cold, and PEG Treatments
3. Discussion
4. Materials and Methods
4.1. Identification of the FLA Family Members in Tomato
4.2. Physicochemical Properties and Signal Peptide Analysis of the FLA Gene Family in Tomato
4.3. Gene Location, Ka (Nonsynonymous)/Ks (Synonymous) Analysis, and Gene Structure Analysis
4.4. Conserved Motif and Protein Conserved Domain Analysis
4.5. Phylogenetic Tree and Cis-Acting Elements Analysis
4.6. Tissue Expression Analysis of the FLA Genes in Tomato
4.7. Transcriptional Analysis of the FLA Genes in Tomato under Different Abiotic Stresses and Hormone Treatments, Plant Materials, and Treatments
4.8. RNA Extraction and qRT-PCR Fluorescence Quantification
4.9. Data Statistics and Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Johnson, K.L.; Jones, B.J.; Bacic, A.; Schultz, C.J. The Fasciclin-Like Arabinogalactan Proteins of Arabidopsis. A multigene family of putative cell adhesion molecules. Plant Physiol. 2003, 133, 1911–1925. [Google Scholar] [CrossRef] [PubMed]
- Faik, A.; Abouzouhair, J.; Sarhan, F. Putative fasciclin-like arabinogalactan-proteins (FLA) in wheat (Triticum aestivum) and rice (Oryza sativa): Identification and bioinformatic analyses. Mol. Genet. Genom. MGG 2006, 276, 478–494. [Google Scholar] [CrossRef] [PubMed]
- De Lorenzo, G.; Ferrari, S.; Giovannoni, M.; Mattei, B.; Cervone, F. Cell wall traits that influence plant development, immunity, and bioconversion. Plant J. 2019, 97, 134–147. [Google Scholar] [CrossRef] [PubMed]
- Bacete, L.; Melida, H.; Miedes, E.; Molina, A. Plant cell wall-mediated immunity: Cell wall changes trigger disease resistance responses. Plant J. 2018, 93, 614–636. [Google Scholar] [CrossRef]
- Showalter, A.M.; Keppler, B.; Lichtenberg, J.; Gu, D.; Welch, L.R. A Bioinformatics Approach to the Identification, Classification, and Analysis of Hydroxyproline-Rich Glycoproteins. Plant Physiol. 2010, 153, 485–513. [Google Scholar] [CrossRef] [PubMed]
- Hijazi, M.; Velasquez, S.M.; Jamet, E.; Estevez, J.M.; Albenne, C. An update on post-translational modifications of hydroxyproline-rich glycoproteins: Toward a model highlighting their contribution to plant cell wall architecture. Front. Plant Sci. 2014, 5, 395. [Google Scholar] [CrossRef]
- Showalter, A.M. Arabinogalactan-proteins: Structure, expression and function. Cell. Mol. Life Sci. CMLS 2001, 58, 1399–1417. [Google Scholar] [CrossRef]
- Clout, N.J.; Tisi, D.; Hohenester, E. Novel fold revealed by the structure of a FAS1 domain pair from the insect cell adhesion molecule fasciclin I. Structure 2003, 11, 197–203. [Google Scholar] [CrossRef]
- Ma, H.; Zhao, J. Genome-wide identification, classification, and expression analysis of the arabinogalactan protein gene family in rice (Oryza sativa L.). J. Exp. Bot. 2010, 61, 2647–2668. [Google Scholar] [CrossRef]
- Zang, L.; Zheng, T.; Chu, Y.; Ding, C.; Zhang, W.; Huang, Q.; Su, X. Genome-Wide Analysis of the Fasciclin-Like Arabinogalactan Protein Gene Family Reveals Differential Expression Patterns, Localization, and Salt Stress Response in Populus. Front. Plant Sci. 2015, 6, 1140. [Google Scholar] [CrossRef] [PubMed]
- Huang, G.-Q.; Xu, W.-L.; Gong, S.-Y.; Li, B.; Wang, X.-L.; Xu, D.; Li, X.-B. Characterization of 19 novel cotton FLA genes and their expression profiling in fiber development and in response to phytohormones and salt stress. Physiol. Plant. 2008, 134, 348–359. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.; Park, J.; Yeom, S.I.; Kim, Y.M.; Seo, E.; Kim, K.T.; Kim, M.S.; Lee, J.M.; Cheong, K.; Shin, H.S.; et al. New reference genome sequences of hot pepper reveal the massive evolution of plant disease-resistance genes by retroduplication. Genome Biol. 2017, 18, 210. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Wu, X. Genome-wide identification, classification and expression analysis of genes encoding putative fasciclin-like arabinogalactan proteins in Chinese cabbage (Brassica rapa L.). Mol. Biol. Rep. 2012, 39, 10541–10555. [Google Scholar] [CrossRef]
- Wang, H.; Jin, Y.; Wang, C.; Li, B.; Jiang, C.; Sun, Z.; Zhang, Z.; Kong, F.; Zhang, H. Fasciclin-like arabinogalactan proteins, PtFLAs, play important roles in GA-mediated tension wood formation in Populus. Sci. Rep. 2017, 7, 6182. [Google Scholar] [CrossRef]
- Zhang, Z.; Xin, W.; Wang, S.; Zhang, X.; Dai, H.; Sun, R.; Frazier, T.; Zhang, B.; Wang, Q. Xylem sap in cotton contains proteins that contribute to environmental stress response and cell wall development. Funct. Integr. Genom. 2015, 15, 17–26. [Google Scholar] [CrossRef] [PubMed]
- Pereira, A.M.; Pereira, L.G.; Coimbra, S. Arabinogalactan proteins: Rising attention from plant biologists. Plant Reprod. 2015, 28, 1–15. [Google Scholar] [CrossRef]
- Shi, H.; Kim, Y.; Guo, Y.; Stevenson, B.; Zhu, J.-K. The Arabidopsis SOS5 locus encodes a putative cell surface adhesion protein and is required for normal cell expansion. Plant Cell 2003, 15, 19–32. [Google Scholar] [CrossRef]
- MacMillan, C.P.; Mansfield, S.D.; Stachurski, Z.H.; Evans, R.; Southerton, S.G. Fasciclin-like arabinogalactan proteins: Specialization for stem biomechanics and cell wall architecture in Arabidopsis and Eucalyptus. Plant J. 2010, 62, 689–703. [Google Scholar] [CrossRef]
- Liu, E.; MacMillan, C.P.; Shafee, T.; Ma, Y.; Ratcliffe, J.; van de Meene, A.; Bacic, A.; Humphries, J.; Johnson, K.L. Fasciclin-Like Arabinogalactan-Protein 16 (FLA16) Is Required for Stem Development in Arabidopsis. Front. Plant Sci. 2020, 11, 615392. [Google Scholar] [CrossRef]
- Wang, H.; Jiang, C.; Wang, C.; Yang, Y.; Yang, L.; Gao, X.; Zhang, H. Antisense expression of the fasciclin-like arabinogalactan protein FLA6 gene in Populus inhibits expression of its homologous genes and alters stem biomechanics and cell wall composition in transgenic trees. J. Exp. Bot. 2015, 66, 1291–1302. [Google Scholar] [CrossRef]
- Hsieh, T.-H.; Li, C.-W.; Su, R.-C.; Cheng, C.-P.; Sanjaya; Tsai, Y.-C.; Chan, M.-T. A tomato bZIP transcription factor, SlAREB, is involved in water deficit and salt stress response. Planta 2010, 231, 1459–1473. [Google Scholar] [CrossRef] [PubMed]
- Krishna, R.; Karkute, S.G.; Ansari, W.A.; Jaiswal, D.K.; Verma, J.P.; Singh, M. Transgenic tomatoes for abiotic stress tolerance: Status and way ahead. 3 Biotech 2019, 9, 143. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Zhou, F.; Wang, H.; Chen, Y.; Yin, T.; Wu, H. Genome-Wide Comparative Analysis of the Fasciclin-like Arabinogalactan Proteins (FLAs) in Salicacea and Identification of Secondary Tissue Development-Related Genes. Int. J. Mol. Sci. 2023, 24, 1481. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.; Lai, Y.; Lv, L.; Ji, M.; Han, K.; Yan, D.; Lu, Y.; Peng, J.; Rao, S.; Yan, F.; et al. Fasciclin-like arabinogalactan gene family inNicotiana benthamiana: Genome-wide identification, classification and expression in response to pathogens. BMC Plant Biol. 2020, 20, 305. [Google Scholar] [CrossRef] [PubMed]
- Qin, N.; Gao, Y.; Cheng, X.; Yang, Y.; Wu, J.; Wang, J.; Li, S.; Xing, G. Genome-wide identification of CLE gene family and their potential roles in bolting and fruit bearing in cucumber (Cucumis sativus L.). BMC Plant Biol. 2021, 21, 143. [Google Scholar] [CrossRef]
- Hsu, P.D.; Scott, D.A.; Weinstein, J.A.; Ran, F.A.; Konermann, S.; Agarwala, V.; Li, Y.; Fine, E.J.; Wu, X.; Shalem, O.; et al. DNA targeting specificity of RNA-guided Cas9 nucleases. Nat. Biotechnol. 2013, 31, 827–832. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Yu, M.; Geng, L.L.; Zhao, J. The fasciclin-like arabinogalactan protein gene, FLA3, is involved in microspore development of Arabidopsis. Plant J. 2010, 64, 482–497. [Google Scholar] [CrossRef]
- Lafarguette, F.; Leple, J.-C.; Dejardin, A.; Laurans, F.; Costa, G.; Lesage-Descauses, M.-C.; Pilate, G. Poplar genes encoding fasciclin-like arabinogalactan proteins are highly expressed in tension wood. New Phytol. 2004, 164, 107–121. [Google Scholar] [CrossRef] [PubMed]
- Adelana, B.O. Relationship between lodging, morphological characters and yield of tomato cultivars. Sci. Hortic. 1980, 13, 143–148. [Google Scholar] [CrossRef]
- Ge, P.; Hao, P.; Cao, M.; Guo, G.; Lv, D.; Subburaj, S.; Li, X.; Yan, X.; Xiao, J.; Ma, W.; et al. iTRAQ-based quantitative proteomic analysis reveals new metabolic pathways of wheat seedling growth under hydrogen peroxide stress. Proteomics 2013, 13, 3046–3058. [Google Scholar] [CrossRef]
- Takahashi, D.; Kawamura, Y.; Uemura, M. Cold acclimation is accompanied by complex responses of glycosylphosphatidylinositol (GPI)-anchored proteins in Arabidopsis. J. Exp. Bot. 2016, 67, 5203–5215. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Chen, H.; Zhang, Y.; Thomas, H.R.; Frank, M.H.; He, Y.; Xia, R. TBtools: An Integrative Toolkit Developed for Interactive Analyses of Big Biological Data. Mol. Plant 2020, 13, 1194–1202. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.-H.; Sato, S.; Sasaki, K.; Saburi, W.; Matsui, H.; Imai, R. COLD SHOCK DOMAIN PROTEIN 3 is involved in salt and drought stress tolerance in Arabidopsis. FEBS Open Bio 2013, 3, 438–442. [Google Scholar] [CrossRef]
- Pan, X.; Wang, C.; Liu, Z.; Gao, R.; Feng, L.; Li, A.; Yao, K.; Liao, W. Identification of ABF/AREB gene family in tomato (Solanum lycopersicum L.) and functional analysis of ABF/AREB in response to ABA and abiotic stresses. PeerJ 2023, 11, e15310. [Google Scholar] [CrossRef] [PubMed]
- Bertinetto, C.; Engel, J.; Jansen, J. ANOVA simultaneous component analysis: A tutorial review. Anal. Chim. Acta X 2020, 6, 100061. [Google Scholar] [CrossRef] [PubMed]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | Gene ID | Number of Amino Acid | Molecular Weight | Theoretical pI | Instability Index | Aliphatic Index | Grand Average of Hydropathicity | Subcellular Localization | Signal Peptide | Transmembrane Domain | GPI |
|---|---|---|---|---|---|---|---|---|---|---|---|
| SlFLA1 | 101263740 | 439 | 48,342.31 | 5.89 | 49.1 | 94.62 | −0.215 | Chloroplast | Yes | Yes | Yes |
| SlFLA2 | 101262658 | 414 | 43,073.97 | 5.6 | 43.06 | 93.55 | 0.085 | Chloroplast | Yes | Yes | Yes |
| SlFLA3 | 101262937 | 427 | 46,167.18 | 5.06 | 46.37 | 97.47 | 0.076 | Chloroplast | Yes | Yes | Yes |
| SlFLA4 | 101255812 | 356 | 39,469.92 | 4.54 | 48.68 | 94.38 | 0.114 | Extracellular | Yes | Yes | Yes |
| SlFLA5 | 101244241 | 249 | 26,003.19 | 4.99 | 36.89 | 83.09 | 0.031 | Cytoplasm | Yes | Yes | Yes |
| SlFLA6 | 101249754 | 421 | 45,266.73 | 5.91 | 52.28 | 101.16 | 0.204 | Chloroplast | Yes | Yes | Yes |
| SlFLA7 | 101249482 | 411 | 43,611.72 | 5.61 | 45.28 | 89.34 | −0.013 | Chloroplast | Yes | Yes | Yes |
| SlFLA8 | 104644532 | 250 | 27,513.02 | 4.89 | 35.76 | 93.96 | −0.238 | Mitochondrion | Yes | Yes | Yes |
| SlFLA9 | 101249179 | 350 | 38,531.73 | 8.83 | 52.89 | 109.74 | 0.249 | Extracellular | Yes | Yes | Yes |
| SlFLA10 | 101264822 | 247 | 26,425.84 | 6.08 | 39.01 | 85.26 | −0.29 | Chloroplast | Yes | Yes | Yes |
| SlFLA11 | 101261419 | 264 | 28,015.18 | 6.58 | 50.2 | 93.14 | 0.17 | Chloroplast | Yes | Yes | Yes |
| SlFLA12 | 101252377 | 409 | 44,082.41 | 6.06 | 31.15 | 88.7 | −0.068 | Extracellular | Yes | Yes | Yes |
| SlFLA13 | 101252705 | 265 | 27,776.55 | 4.83 | 52.11 | 83.66 | −0.064 | Chloroplast | Yes | Yes | Yes |
| SlFLA14 | 101257010 | 251 | 26,706.26 | 5.89 | 44.16 | 82.87 | −0.186 | Extracellular | Yes | Yes | Yes |
| SlFLA15 | 104649632 | 362 | 36,790.89 | 4.49 | 60.78 | 73.76 | −0.096 | Endoplasmic ergatoplasm | Yes | Yes | Yes |
| SlFLA16 | 101256014 | 247 | 26,542.2 | 5.6 | 46.36 | 89.6 | −0.006 | Plasma membrane | Yes | Yes | Yes |
| SlFLA17 | 101266654 | 239 | 26,134.93 | 8.56 | 47.59 | 89.46 | 0.023 | Vacuole | Yes | Yes | Yes |
| SlFLA18 | 101254871 | 354 | 38,682.99 | 7.16 | 39.12 | 101.05 | 0.289 | Vacuole | Yes | Yes | Yes |
| SlFLA19 | 104644785 | 449 | 49,434.64 | 5.67 | 46.5 | 95.75 | −0.14 | Endoplasmic ergatoplasm | No | Yes | Yes |
| SlFLA20 | 101244600 | 415 | 44,208.4 | 5.73 | 38.73 | 81.9 | −0.047 | Chloroplast | Yes | Yes | Yes |
| SlFLA21 | 101249187 | 397 | 42,609.93 | 6.29 | 49.14 | 95.52 | 0.251 | Plasma membrane | Yes | Yes | Yes |
| SlFLA22 | 101265484 | 401 | 44,767.54 | 6.3 | 44.95 | 99.68 | 0.047 | Vacuole | Yes | Yes | Yes |
| SlFLA23 | 101260248 | 242 | 25,793.38 | 5.43 | 27.98 | 92.69 | 0.058 | Vacuole | Yes | Yes | Yes |
| SlFLA24 | 101254060 | 277 | 30,716.84 | 9.34 | 36.03 | 83.86 | −0.168 | Chloroplast | Yes | Yes | Yes |
| Cis-Element | Number of Genes | Sequence of Cis-Element | Functions of Cis-Elements |
|---|---|---|---|
| ABRE | 27 | ACGTG | cis-acting element involved in abscisic acid responsiveness |
| AE-box | 12 | AGAAACTT | part of a module for light response |
| ARE | 25 | AAACCA | cis-acting regulatory element essential for anaerobic induction |
| ATCT-motif | 7 | AATCTAATCC | part of a conserved DNA module involved in light responsiveness |
| Box 4 | 63 | ATTAAT | part of a conserved DNA module involved in light responsiveness |
| CAT-box | 10 | GCCACT | cis-acting regulatory element related to meristem expression |
| CGTCA-motif | 30 | CGTCA | cis-acting regulatory element involved in MeJA responsiveness |
| GA-motif | 7 | ATAGATAA | part of a light-responsive element |
| GARE-motif | 5 | TCTGTTG | gibberellin-responsive element |
| GATA-motif | 15 | GATAGGG | part of a light-responsive element |
| G-box | 36 | CACGTC | cis-acting regulatory element involved in light responsiveness |
| GT1-motif | 26 | GGTTAA | light-responsive element |
| LAMP-element | 4 | CTTTATCA | part of a light-responsive element |
| LTR | 14 | CCGAAA | cis-acting element involved in low-temperature responsiveness |
| MBS | 8 | TTTTTACGGTTA | MYB binding site involved in drought inducibility |
| MRE | 4 | AACCTAA | MYB binding site involved in light responsiveness |
| P-box | 7 | CCTTTTG | gibberellin-responsive element |
| TATC-box | 10 | TATCCCA | cis-acting element involved in gibberellin responsiveness |
| TCA-element | 10 | CCATCTTTTT | cis-acting element involved in salicylic acid responsiveness |
| TCCC-motif | 8 | TCTCCCT | part of a light-responsive element |
| TC-rich repeats | 9 | ATTCTCTAAC | cis-acting element involved in defense and stress responsiveness |
| TCT-motif | 20 | TCTTAC | part of a light-responsive element |
| TGACG-motif | 30 | TGACG | cis-acting regulatory element involved in MeJA responsiveness |
| TGA-element | 4 | AACGAC | auxin-responsive element |
| Motif | Width (aa) | Motif Sequence |
|---|---|---|
| Motif 1 | 41 | AHNITKILEKYPZFSTFNRLLSTTQLAAEINSRLTITVLAV |
| Motif 2 | 50 | JKNVLSLHVLLDYFDAKKLHKITDGTTLVTTMFQATGKAPGNSGFVNITD |
| Motif 3 | 50 | ATNGAGKFPLNITTDGDQVNISTGIVTAKISNTIYDDNPLAIYQVDKVLL |
| Motif 4 | 29 | ZQKVQLLQYHVJPSYYSLSSLQTLSNPVR |
| Motif 5 | 48 | KGGKVGFGSADNBGHLPSTFVKSVMEIPYNISVJQISQPJVSPGAEAP |
| Motif 6 | 14 | TIFAPTDEAFSNLK |
| Motif 7 | 23 | FINGVEITDPDJYVNGRFVVHGI |
| Motif 8 | 21 | SPAPAPAPAPETAKAKTKSSK |
| Motif 9 | 29 | LMSAHGCKVFASLLLASPVEKTFEDBEEG |
| Motif 10 | 50 | DSHYTELSELVEKALLLQPLEEAVSKHNITIFAPKNEALERDLDPEFKRF |
| Protein | Alpha Helix (%) | Extended Strand (%) | Beta Turn (%) | Random Coil (%) | Distribution of Secondary Structure Elements |
|---|---|---|---|---|---|
| SlFLA1 | 35.08 | 15.26 | 6.61 | 43.05 |  |
| SlFLA2 | 22.6 | 25.99 | 6.5 | 44.92 |  |
| SlFLA3 | 37.36 | 14.72 | 3.4 | 44.53 |  |
| SlFLA4 | 36.98 | 16.55 | 7.06 | 39.42 |  |
| SlFLA5 | 35.19 | 18.04 | 5.12 | 41.65 |  |
| SlFLA6 | 28.34 | 19.84 | 4.86 | 46.96 |  |
| SlFLA7 | 37.66 | 16.74 | 5.44 | 40.17 |  |
| SlFLA8 | 30.32 | 22.38 | 5.05 | 42.24 |  |
| SlFLA9 | 32.3 | 19.48 | 3.8 | 44.42 |  |
| SlFLA10 | 25 | 19.94 | 7.87 | 47.19 |  |
| SlFLA11 | 28.92 | 15.26 | 2.41 | 53.41 |  |
| SlFLA12 | 30.68 | 19.67 | 7.26 | 42.39 |  |
| SlFLA13 | 41.43 | 13.15 | 4.78 | 40.64 |  |
| SlFLA14 | 33.33 | 18.6 | 5.31 | 42.75 |  |
| SlFLA15 | 28.97 | 15.37 | 5.54 | 50.13 |  |
| SlFLA16 | 27.13 | 18.22 | 4.05 | 50.61 |  |
| SlFLA17 | 32.8 | 20.8 | 2.8 | 43.6 |  |
| SlFLA18 | 35.4 | 19.9 | 8.01 | 36.69 |  |
| SlFLA19 | 30.92 | 21.7 | 7.23 | 40.15 |  |
| SlFLA20 | 27.43 | 18 | 6.86 | 47.71 |  |
| SlFLA21 | 35.61 | 17.42 | 6.06 | 40.91 |  |
| SlFLA22 | 41.2 | 14.46 | 6.27 | 38.07 |  |
| SlFLA23 | 34.72 | 19.56 | 6.36 | 39.36 |  |
| SlFLA24 | 28.1 | 23.97 | 4.96 | 42.98 |  |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yao, K.; Yao, Y.; Ding, Z.; Pan, X.; Zheng, Y.; Huang, Y.; Zhang, Z.; Li, A.; Wang, C.; Li, C.; et al. Characterization of the FLA Gene Family in Tomato (Solanum lycopersicum L.) and the Expression Analysis of SlFLAs in Response to Hormone and Abiotic Stresses. Int. J. Mol. Sci. 2023, 24, 16063. https://doi.org/10.3390/ijms242216063
Yao K, Yao Y, Ding Z, Pan X, Zheng Y, Huang Y, Zhang Z, Li A, Wang C, Li C, et al. Characterization of the FLA Gene Family in Tomato (Solanum lycopersicum L.) and the Expression Analysis of SlFLAs in Response to Hormone and Abiotic Stresses. International Journal of Molecular Sciences. 2023; 24(22):16063. https://doi.org/10.3390/ijms242216063
Chicago/Turabian StyleYao, Kangding, Yandong Yao, Zhiqi Ding, Xuejuan Pan, Yongqi Zheng, Yi Huang, Zhuohui Zhang, Ailing Li, Chunlei Wang, Changxia Li, and et al. 2023. "Characterization of the FLA Gene Family in Tomato (Solanum lycopersicum L.) and the Expression Analysis of SlFLAs in Response to Hormone and Abiotic Stresses" International Journal of Molecular Sciences 24, no. 22: 16063. https://doi.org/10.3390/ijms242216063