1. Introduction
Inflammatory bowel disease (IBD), including ulcerative colitis (UC), Crohn’s disease (CD), as well as IBD unclassified (IBDU), is a category of chronic or recurrent inflammatory conditions principally targeting the small and large intestine [
1]. Among them, UC is characterized by continuous inflammation mainly located in the colon, whereas CD presents as regional inflammation all over the whole gastrointestinal tract. Moreover, inflammation in UC is confined to the intestinal mucosa, whereas CD involves multiple layers of the intestinal wall [
2]. Currently, the prevalence of IBD is increasing worldwide. It is estimated that the number of people suffering from IBD in the Western world could exceed ten million in 2030 [
3]. Recent research has indicated that environmental factors, gut microbiota, genetic susceptibility, and excess immunity participate in the pathogenesis of IBD. However, the exact cause of IBD remains uncertain [
4]. Based on this, IBD is not possible to cure so far. Therefore, the primary goal of IBD treatment is to ameliorate the inflammation, resulting in the signs and symptoms of patients. In general, IBD treatment involves drug therapy and surgery. The drug treatment of IBD mainly includes antibiotics, anti-inflammatory drugs, immune system suppressors, and biologics, which have troublesome side effects when used for a long time. Moreover, surgery does not cure CD, and the benefits are usually temporary [
5]. Recently, alternative medicine has increasingly emerged as a research focus in IBD treatment. This includes the investigation of phytochemicals that offer greater tolerance and fewer adverse effects, which has garnered significant interest among medical scientists.
A recent survey showed the total daily amount of flavonoids in the average Western diet ranges from 200–1000 mg/day and drops to 144 mg/day in IBD patients. In addition, a positive correlation has been established between low dietary flavonoid consumption and severe IBD [
6]. A randomized trial indicated that adherence to the Mediterranean diet, which is rich in flavonoids, improved clinical scores and inflammatory markers in children and adolescents with mildly to moderately active IBD [
7]. The above trial suggests that the supplementation of flavonoids benefits IBD patients. Moreover, certain flavonoids have potential protective effects against IBD. For instance, Fu et al. utilized a dextran sodium sulfate (DSS)-induced colitis mouse model to evaluate the effect of apigenin on UC and found that apigenin effectively inhibited inflammation and protected the gut barrier via balancing the gut microbiome [
8]. Wu et al. found that oral epigallocatechin-3-gallate (EGCG) ameliorated colonic inflammation in a gut microbiota-dependent manner [
9]. Chaen et al. found that naringenin promoted recovery from colonic damage via suppression of epithelial tumor necrosis factor-α production and induction of M
2-type macrophages in colitic mice [
10]. In addition, quercetin was reported to be able to alleviate UC via activating aryl hydrocarbon receptors to improve intestinal barrier integrity [
11]. Recent studies suggested that a combination of various flavonoids may exhibit synergistic anti-inflammatory effects in IBD [
12]. However, only limited research has thus far focused on the synergistic anti-inflammatory properties of a flavonoid combo in IBD.
Flavo-Natin is a combined formulation, mainly containing apigenin and EGCG, with smaller amounts of quercetin and epicatechin. In recent years, Flavo-Natin has found clinical application in the management of diarrhea associated with irritable bowel syndrome (IBS) and in the amelioration of dyspeptic symptoms [
13,
14]. On the one hand, the symptoms and pathogenesis of IBS are similar to those of IBD. On the other hand, the main components in Flavo-Natin are apigenin and EGCG, which have been confirmed to have the potential to treat IBD. Therefore, we have reason to believe that Flavo-Natin is a suitable candidate for the adjunctive treatment of IBD.
In this study, to evaluate the effect of Flavo-Natin administration on UC, we used a hapten-mediated (oxazolone) experimental colitis model to induce acute and chronic colitis in mice. The histological features of acute colitis, such as epithelial damage and infiltration of neutrophils, macrophages, and lymphocytes in mucosa and submucosa, together with the inflammation of the distal colon and elevated levels of type 2 cytokines by lamina propria T cells, are similar to the features of human UC [
15]. Additionally, the oxazolone colitis model may be useful in the study of the inflammatory response of chronic UC since chronic colitis can be induced by repetitive administration of oxazolone [
15]. Furthermore, we explored whether Flavo-Natin could alleviate colitis by improving the epithelial barrier integrity, decreasing the impairment of colonic tissue, inhibiting the inflammatory level, and balancing the gut microbiome structure in oxazolone-induced colitis mice.
3. Discussion
Numerous studies have confirmed that certain flavonoids have a considerable prospect for treating IBD [
19,
20,
21,
22,
23]. To date, pathways have been found to participate in the regulation of IBD mediated by flavonoids, in which the preservation of the intestinal epithelial barrier is a key step in preventing the aggravation of IBD [
24]. Flavonoids have previously been demonstrated to improve the epithelial barrier. For instance, Watson et al. found that EGCG blocked epithelial barrier dysfunction induced by interferon-γ (
IFN-γ) by measuring TEER in T84 (human colonic epithelial cell) monolayers [
25]. In addition, quercetin and naringenin have been found to dose-dependently increase the TEER of Caco-2 cell monolayers within the concentration range of 10–100 µM [
16,
17]. Consistent with these findings, EGCG, quercetin, and naringenin significantly increased the TEER of Caco-2 cell monolayers in our study, indicating the preservation of the intestinal epithelial barrier by flavonoids. However, not all flavonoids possess the capability to enhance the epithelial barrier. In our observations, apigenin significantly decreased the TEER of Caco-2 cell monolayers, suggesting the dysfunction of the intestinal epithelial barrier. Moreover, Flavo-Natin, containing apigenin and EGCG, significantly increased the TEER of Caco-2 cell monolayers at a high concentration. However, no notable difference was observed at a low concentration. The discrepancy may be attributed to apigenin potentially counteracting the positive effect of EGCG. When Caco-2 cell monolayers were incubated with a low concentration of Flavo-Natin, apigenin, a main component of Flavo-Natin, greatly offset the increase in TEER induced by EGCG. As a result, we did not observe an enhancement in TEER values due to the low concentration of Flavo-Natin. However, at a high concentration of Flavo-Natin, the amount of quercetin significantly increased. Furthermore, quercetin demonstrated a substantial increase in TEER values (see
Figure 2). Therefore, apigenin did not counteract the augmentation of TEER values induced by both EGCG and quercetin.
It has been reported that a combination of multiple phytochemicals, including flavonoids, can lead to synergistic anti-inflammatory activity [
26]. For instance, the combined application of EGCG and genistein resulted in an improvement in EGCG uptake within the small intestine [
27]. Moreover, the combined administration of quercetin and resveratrol exhibited a synergistic anti-inflammatory effect in rats subjected to a high-fat–sucrose diet [
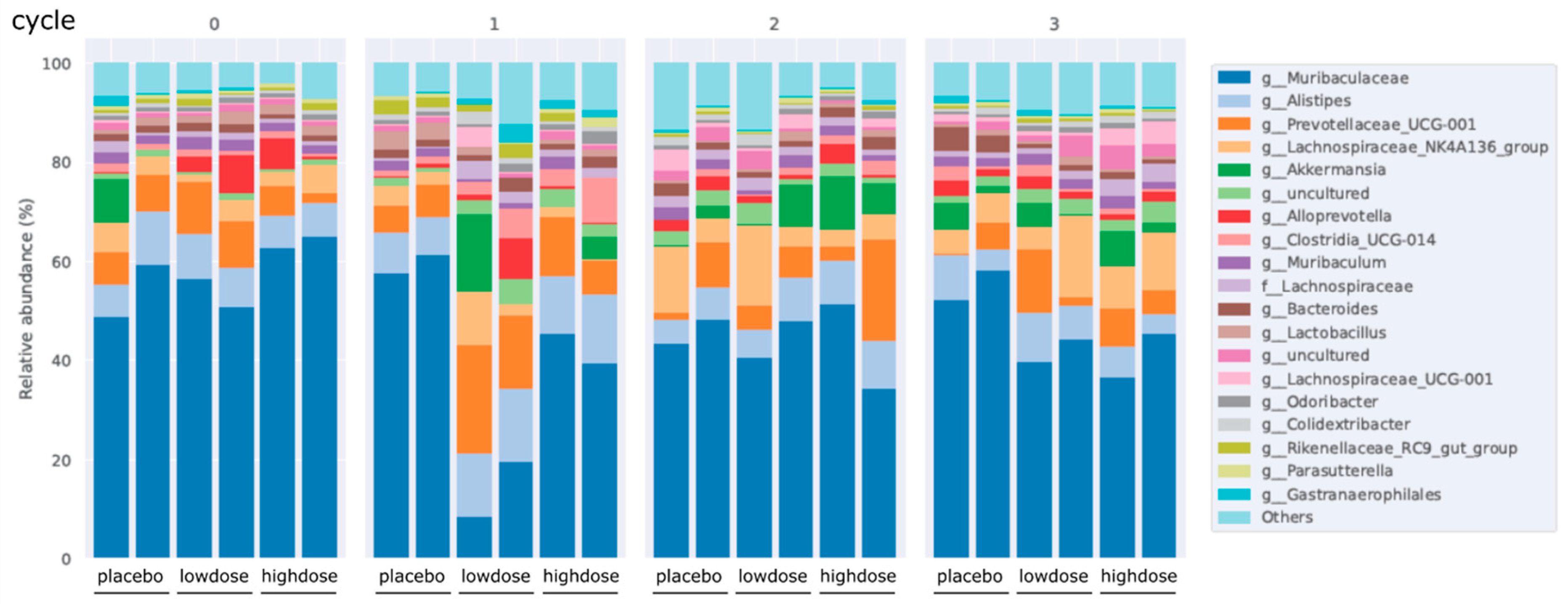
28]. Therefore, it is plausible to consider that the concurrent administration of multiple flavonoids could potentially evoke synergistic anti-inflammatory properties in IBD. In this study, Flavo-Natin as a flavonoid combo, at a dose of 0.2 mg/day and 0.8 mg/day, was used to treat the chronic colitis mice induced by oxazolone. However, the reduced inflammation measured by MEICS was not observed in the low-dose and high-dose groups after the 1st and 2nd challenges, compared with the placebo group. Only after the 3rd challenge were significant differences in inflammatory extent (MEICS, histological score, MPO activity, and cytokine levels) between the treatment and placebo groups evident, indicating the time-dependent therapeutic effect of Flavo-Natin. Moreover, we observed alterations in the intestinal microbiota in chronic colitis models, revealing an increase in the proportion of
Alloprevotella and
Akkermansia following oral administration of Flavo-Natin. Recent studies have found the crucial role of
Alloprevotella in maintaining the intestinal epithelial barrier and improving intestinal immune function [
29], while
Akkermansia is able to regulate the thickness of the intestinal mucosa layer and strengthen therapeutical outcomes in chronic diseases associated with leaky gut and inflammation [
30]. Furthermore, in alignment with our results, Wu et al. observed that oral administration of EGCG significantly increased
Akkermansia abundance in mice, leading to the attenuation of colitis. The potential protective mechanism may involve the substantial enhancement of short-chain fatty acids (SCFAs) production by
Akkermansia. These SCFAs contribute to an anti-oxidative and anti-inflammatory state, thus providing further protection against colon damage [
9]. In addition, apigenin also significantly enhanced the abundance of
Akkermansia and
Faecalibaculum. These beneficial bacteria produced a large amount of SCFAs, mainly including acetic acid, propionic acid, and butyric acid. Subsequently, microbiota-derived SCFAs upregulated IL-22 production, which protects intestines from inflammation to maintain intestinal homeostasis [
8]. In our study, the protective pathway of alteration in intestinal microbiota has not been investigated. However, considering Flavo-Natin as a combination of apigenin and EGCG, we have grounds to believe that an increase in the proportion of
Alloprevotella and
Akkermansia could stimulate the production of SCFAs, which is likely associated with the mitigation of colitis.
The results in chronic colitis models were not as favorable as anticipated. We hypothesized that the failure of Flavo-Natin to alleviate inflammation after the 1st and 2nd challenges may be attributed to an insufficient dosage. Conversely, the success observed after the 3rd challenge can be attributed to the cumulative effect of Flavo-Natin over time. Additionally, Fu et al. demonstrated significant improvement in ulcerative colitis in chronic colitis mice induced by DSS through the administration of apigenin at doses of 3 mg/day and 5 mg/day [
8]. Wu et al. effectively alleviated experimental colitis by orally administering 1 mg/day of EGCG to acute DSS-induced colitis mice [
9]. It should be noted that the doses of apigenin and EGCG utilized in the above studies were considerably higher than those employed in our study. To validate this hypothesis, higher doses of Flavo-Natin were administered to mice with acute oxazolone-induced colitis. Significant disparities in inflammation were observed between the treatment groups and placebo group in acute colitis models when having a gavage of Flavo-Natin at a dose of 3 mg/day, as assessed by MEICS, histological score, MPO activity, and cytokine levels. These findings suggest a dose-dependent correlation between the administration of Flavo-Natin and its therapeutic efficacy in IBD. Besides, Fu et al. employed oral administration of apigenin to treat chronic colitis in mice induced by DSS and discovered that a daily dosage of 5 mg exhibited a more robust anti-inflammatory effect than a dosage of 3 mg [
8]. Xu et al. investigated the therapeutic effect of EGCG on acute colitis in mice induced by DSS, revealing that a dosage of 2 mg/day produced a better therapeutic outcome compared with a dosage of 1 mg/day [
31]. These observations align with our findings, suggesting that both Flavo-Natin and its primary components, apigenin, and EGCG, indicate a dose-dependent correlation between their oral administration and the therapeutic effect in IBD, despite variances in experimental setups.
In conclusion, the time- and dose-dependent protective effects of Flavo-Natin on IBD were demonstrated via both chronic and acute colitis models induced by oxazolone, supporting the importance of the protective effect of Flavo-Natin in the inflammatory bowel disease model. However, there are some limitations in our study. First, we only employed an oxazolone-induced colitis model to assess the protective effects of Flavo-Natin on IBD, which can partially mimic IBD to a certain extent but not replicate it fully. In the next animal experiment, we intend to utilize a DSS-induced colitis model to address this limitation comprehensively. Second, the sample size of the collected stool specimens in this study was relatively small. Therefore, the reliability of our intestinal microbiome analysis may focus on the small samples but can be increased in the future. Lastly, we did not conduct a comparative analysis of Flavo-Natin with apigenin and EGCG regarding their therapeutic efficacy in IBD. We plan to address these limitations in our future study.
4. Materials and Methods
4.1. Cell Culture
Caco-2 cells were obtained from the European Collection of Authenticated Cell Cultures (ECACC) and cultured in complete RPMI 1640 Medium supplemented with 10% Fetal Calf Serum (FCS), 1% GlutaMAX (100×), 1% Sodium Pyruvate (100×), Penicillin (100 U/mL), and Streptomycin (100 µg/mL) at 37 °C in a 5% CO2 incubator. In our study, Caco-2 cells were from passage number 20–50.
4.2. Transepithelial Electrical Resistance (TEER)
TEER often serves as a rough indicator of tight junction permeability; here, we performed a TEER measurement to evaluate intestinal epithelial barrier integrity in Caco-2 cell monolayer incubated by different drugs. Briefly, Caco-2 cells were seeded at a density of 50,000 cells/mL on Transwell inserts (24-well, 0.4 µm pore size, 0.336 cm
2 area, transparent, Greiner Bio-One Co., Kremsmünster, Austria) and cultured in complete RPMI 1640 medium for 21 days, which was replaced twice a week. Based on the method of Yamashita et al. [
32], TEER values were measured at 20 °C using a Millicell-ERS resistance monitoring apparatus (Millipore, Burlington, MA, USA). The chopstick electrodes, connected to the epithelial volt-ohmmeter, had to be calibrated to 0 ohm (Ω) before each measurement. Subsequently, they were placed into two chambers of each Transwell insert to measure TEER values. The TEER value (in Ω × cm
2) was calculated by subtracting the background value (see the following equation).
After culturing for 21 days, only Caco-2 cell monolayers with TEER values above 400 Ω × cm2 were used for further experiments. Whereafter, the qualified Caco-2 cell monolayers were incubated with different concentrations of drugs for 72 h, which were dissolved in a complete RPMI 1640 medium. At regular intervals, the TEER value of each insert was measured in triplicated and recorded. Over 72 h of drug incubation, the cell medium was not changed.
4.3. Immunofluorescence Staining of Caco-2 Cell Monolayer
A total volume of 100 μL of 4.5% Paraformaldehyde (PFA) was used to fix Caco-2 cell monolayers for 10 min. Subsequently, PFA was removed, and PBS (200 μL) was used to wash the residual PFA. The borders of Caco-2 cells were stained with 100 μL of rhodamine-phalloidin (1:50) in buffered PBS + 0.2% Triton-X 100 for 15 min. PBS (200 μL) was used to wash the residual dye after removing the solution, and 100 μL of DAPI (1:5000) was used to stain the cell nucleus for 5 min. Finally, the inserts were cut and mounted on glass slides with Mowiol after removing the DAPI solution. The slides were stored at 4 °C, and the above steps were all performed in the dark. Images were captured using a confocal microscope.
4.4. Scratch Assay
Silicone chambers, purchased from the Ibidi company, were utilized for culturing Caco-2 cells. First, 350,000 Caco-2 cells in 70 µL medium were evenly added into each of the two silicone chambers via pipette. These chambers were subsequently transferred into an incubator (37 °C and 5% CO
2) for 72 h until the gap between the two silicone chambers was colonized by Caco-2 cells. The silicone chambers were then removed, and 2 mL of medium was added to the chambers. This medium contained different concentrations of Flavo-Natin (0 μg/mL, 10 μg/mL, 100 μg/mL). The chambers were analyzed after 24, 48, and 72 h. At different time points, the images were captured under a microscope, and the migrated distance was measured using Leica Application Suite (LAS) software SP5. The migration rate was calculated using the following formula.
In the above formula, D0 h represents the distance of the gap at 0 h, and D h represents the distance at 24, 48, and 72 h.
4.5. Mice
For our experiments, 8–10-week-old female C57BL/6 mice (weighing 18–20 g) were obtained from Jackson Laboratory. All mice were maintained on a 12-h light-dark cycle at 23 ± 1 °C with a relatively constant humidity of 50 ± 5% under specific pathogen-free (SPF) conditions and had free access to water and standard laboratory pellets. The health and potential infestation with microbes and parasites were regularly analyzed by our veterinarian. Before the experiment, the mice were given 5 days to adapt to our animal facility. The animal study was performed in accordance with institutional guidelines and under the permission of the government of Middle Franconia in Germany.
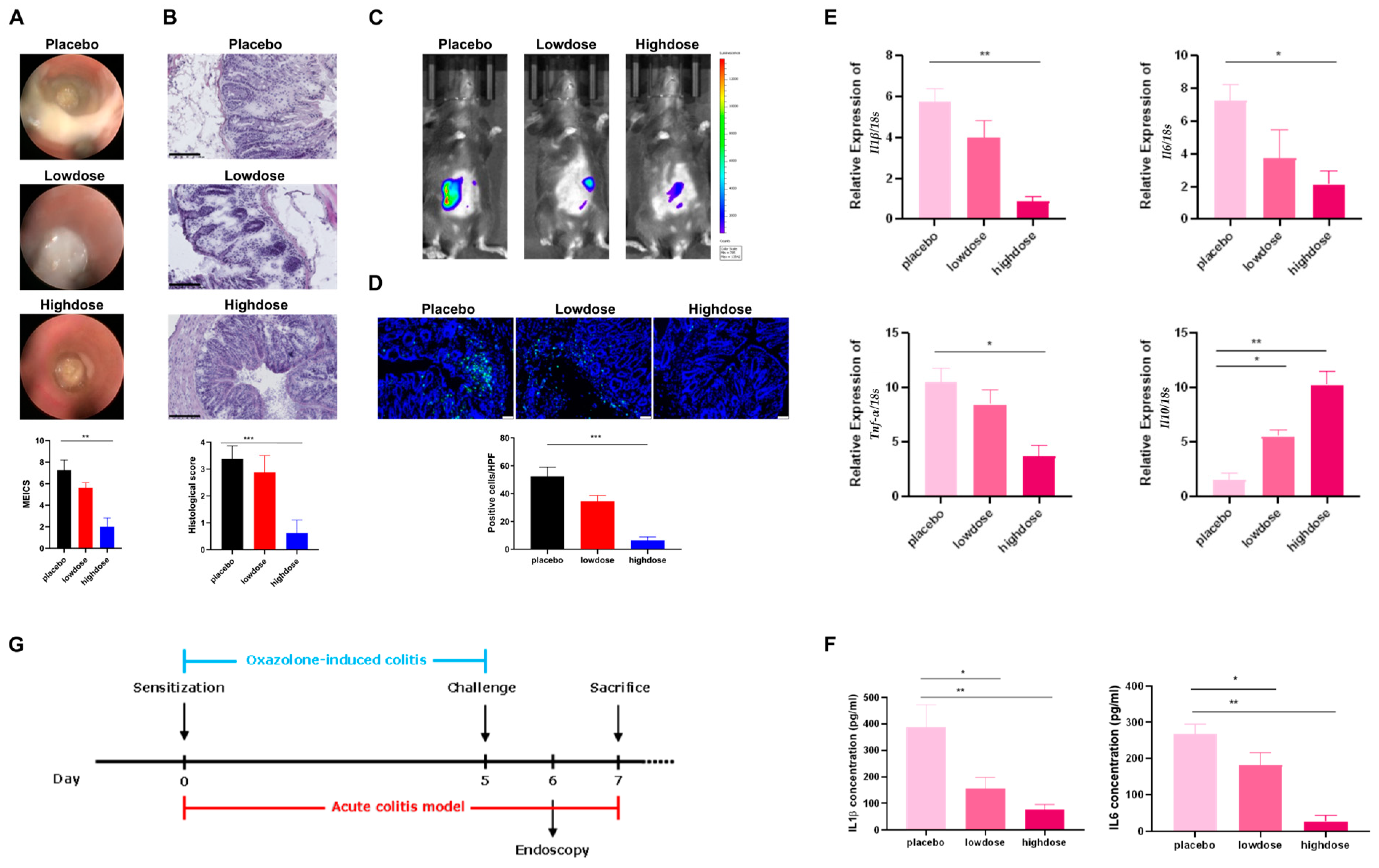
4.6. Acute and Chronic Oxazolone-Induced Colitis Models
For acute oxazolone-induced colitis, the first step was sensitization. In this process, the abdomens of the mice were shaved, followed by a topical administration of 100 μL of 3% oxazolone solution. They were then challenged using oxazolone solution 5 days after sensitization, and the operation was as follows: 1% oxazolone solution (100 μL) was administered into the colon of the mice after they were anesthetized. We used a mini endoscopy to evaluate the severity of colitis 24 h after the challenge. Finally, all mice were euthanized 48 h after the challenge, and their organ tissues were removed to be further analyzed. Compared with acute oxazolone-induced colitis, chronic oxazolone-induced colitis can mimic the characteristic symptoms of UC better. Therefore, we also set up chronic oxazolone-induced colitis models. First, sensitization was conducted. Afterward, the challenges were repeated for 3 cycles at a 0.5% oxazolone concentration, followed by cycles of recovery. The extent of colitis was analyzed with a mini endoscopy 24 h after each challenge. Finally, all mice were euthanized 48 h after the last challenge [
15].
4.7. Administering Medications to Mice in Colitis Models
Flavo-Natin is a combined formulation, mainly containing apigenin and EGCG, with smaller amounts of quercetin and epicatechin. For the composition of Flavo-Natin, see
Table 1. First, different amounts of Flavo-Natin were diluted in 2% carboxymethylcellulose sodium (CMC-Na) solutions. To observe its protective effect on IBD, 100 μL of Flavo-Natin suspension was orally administered in acute or chronic oxazolone-induced colitis mice as treatment groups. Contrastingly, the mice accepted 100 μL of 2% CMC-Na solution were considered as a placebo group. The mice were orally administered Flavo-Natin from the start to the end in acute or chronic oxazolone-induced colitis models.
In regard to the acute oxazolone-induced colitis models, all the mice were randomly distributed over the cages and divided into 3 groups: placebo group, low-dose group, and high-dose group (
n ≥ 5 per group). We orally administered 100 μL of 2% CMC-Na solutions to the mice in the placebo group each day. In addition, 100 μL of Flavo-Natin suspensions, in amounts of 1 mg and 3 mg, were orally administered in the low-dose and high-dose groups each day, respectively. All mice were induced with colitis with oxazolone (
Figure 5G).
As for the chronic oxazolone-induced colitis models, we randomly distributed all mice over the cages and divided them into 3 groups: placebo group, low-dose group, and high-dose group (
n ≥ 4 per group). For the placebo group, 100 μL of 2% CMC-Na solution was orally administered in mice. Contrastingly, 100 μL of Flavo-Natin suspension, at a concentration of 2 mg/mL or 8 mg/mL, was orally administered in the low-dose group or high-dose group. Three groups were then induced with colitis with oxazolone (
Figure 3G).
To minimize observational bias, one researcher was responsible for the randomization process of allocating mice to different groups and oral administration of drugs each day. Contrastingly, another researcher, blinded to the treatment groups, was in charge of establishing acute and chronic oxazolone-induced colitis models and subsequently conducted endoscopic and tissue analyses.
4.8. Endoscopic Analysis
A mini endoscopy analysis was conducted to assess the severity of colonic inflammation 24 h after each challenge by oxazolone during the colitis course. First, the mice were anesthetized with 2% isoflurane by placing them in a chamber. The mini endoscope was then introduced rectally, and photographs were captured. The indicators outlined in
Table 2 were assessed and graded on a scale of 0–3 for each indicator. These scores were aggregated, and the MEICS was determined accordingly [
33]. The MEICS spans from 0 (the absence of inflammation) to 15 (severe inflammation).
4.9. Comparison of MPO Activity Using IVIS
In vivo staining to detect MPO activity was performed 24 h after challenge during the oxazolone-induced colitis course. MPO is a key marker for inflammation and is predominantly expressed by neutrophils and macrophages. It mediates the conversion of H
2O
2/chloride ions into hypochlorous acid. Luminol interacts with hypochlorous acid, then emitting luminescent light that is detectable. This luminescence emission can offer insights into MPO activity [
34]. To measure MPO activity, mice were intraperitoneally injected with 20 nM luminol L-012 (100 µL) and anesthetized using 2% isoflurane in a chamber. With their shaved bellies facing upwards, the mice were placed in the IVIS chamber. Luminescence was then measured for 1 min.
4.10. Stool Microbiome Test
Recent studies found that IBD is associated with intestinal dysbiosis [
35]. In addition, some flavonoids can modulate gut microbiota to prevent and treat IBD [
8]. To explore the correlation between flavonoids and the alteration of intestinal flora, we collected the stool derived from the mice in chronic colitis models. The stools were collected 2 days after the starting point and the following 1st, 2nd, and 3rd challenges. Afterward, total microbial genomic DNA was isolated from each fecal sample. The concentration of DNA was measured by a NanoDrop 1000 device. DNA was stored at −20 °C for further studies. Finally, all DNA samples were tested through 16s rRNA gene sequencing, and microbiota data were analyzed.
4.11. Hematoxylin and Eosin (H and E) Staining
The colon tissue was fixed on the slides and stained with hematoxylin and eosin. Afterward, bright field microscopy was used to capture images. All H and E staining slides were analyzed by a pathologist, in which the extent of colitis was scored in
Table 3 [
15].
4.12. MPO Staining of Tissue
The colon tissue on the slides was fixed by 4.5% PFA for 15 min. Subsequently, the slides underwent a triple wash with PBS, and the tissue was subjected to permeabilization using 1× Permeabilization buffer in PBS for 10 min. To minimize nonspecific antibody binding, a blocking solution was applied and incubated for 30 min. Following this, the colon tissue on the slide was intracellularly stained for MPO using a goat antibody (dilution 1:50). The antibody was then appropriately diluted in antibody dilution buffer, and the tissue sections were incubated with this antibody solution overnight at 4 °C. Following overnight incubation, the slides were once again washed thrice with PBS-Tween, and a rabbit-anti-goat antibody (dilution 1:200) was applied for 2 h at room temperature. After another round of PBS washing (3 times), the cell nuclei were stained with DAPI (1:5000 in PBS) for 5 min. Finally, a cover slip was affixed using Mowiol. The immunofluorescence images were captured using a confocal microscope. During the process of MPO staining, positive and negative controls were established to demonstrate the reliability of the results. Negative control slides were incubated with PBS instead of primary and secondary antibodies, whereas positive control slides were incubated with PBS and secondary antibodies.
4.13. RNA Isolation, Synthesis of cDNA, and Quantitative PCR (qPCR)
Total RNA was isolated from the colonic tissues of mice using the Nucleo Spin RNA Kit. After measuring the concentration of RNA, cDNA was synthesized from 500 ng RNA using the SCRIPT cDNA Synthesis Kit. Synthesized cDNA was stored at −20 °C and diluted with nuclease-free water in a 1:10 ratio before conducting a qPCR assay. The housekeeping gene
18S rRNA, exhibiting constitutive expression in the inflamed colon tissue, served as the reference gene for the target genes.
Table 4 lists the primers of target genes, including
Il1β,
Il6,
Tnf-α, and
Il10. The mRNA levels of these target genes were calculated by the 2
−△△Ct method. All assays were repeated independently in triplicate to enhance the robustness of the results.
4.14. Enzyme-Linked Immunosorbent Assay (ELISA)
After the death of the mice, the plasma was immediately separated from blood samples via centrifugation at 12,500 rpm and subsequently stored at −20 °C for further study. The concentrations of IL-1β, IL-6, IL-10, and TNF-α in the plasma were determined using ELISA. In brief, 96-well plates were coated with the respective capture antibody (50 µL) in duplicate, followed by incubation overnight at 4 °C. On the second day, they were washed 3 times using 200 µL of PBS-Tween (0.05%). A total volume of 100 µL of assay diluent was then used to incubate the wells for 1 h at room temperature. Subsequently, samples (50 µL) or the respective standards were incubated in the wells overnight at 4 °C after a rinse with PBS-Tween. It was worth noting that all samples were diluted 1:2 with assay diluent. On the third day, the wells were washed again, and a detection antibody (50 µL) was added to the wells, which were slightly shaken for 1 h at room temperature. Avidin-HRP (50 µL) reagent was added into the wells after a rinse with PBS-Tween. The plates were incubated for 30 min in the dark. Thereafter, 50 µL of substrate solution was added into each well after washing again. The plates were placed in the dark for 15–30 min. Finally, 50 µL of 2N H2SO4 was added to each well to stop the chemical conversion. The absorbance was measured at 450 nm, simultaneously with a reference wavelength (570 nm), using an ELISA reader instrument.
4.15. Statistics
Statistical differences between the two groups were assessed using paired Student’s t-tests. When statistical differences between more than two groups were assessed, one-way ANOVA with an additional Tukey post-test was used. Statistical analysis was performed using GraphPad Prism 9.0. p values < 0.05 were considered statistically significant and marked with asterisks (* < 0.05, ** < 0.01, *** < 0.001, **** < 0.0001). In the t-test for independent samples, the effect size was determined according to Cohen’s by calculating the mean difference between the two groups and dividing the result by the pooled standard deviation. Data were presented as means +/− standard error of the mean (SEM). D-value in this study was qualified.








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}