
Auraptene Enhances AMP-Activated Protein Kinase Phosphorylation and Thereby Inhibits the Proliferation, Migration and Expression of Androgen Receptors and Prostate-Specific Antigens in Prostate Cancer Cells

 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Auraptene Suppresses Proliferation and Migration of Prostate Cancer LNCaP Cells
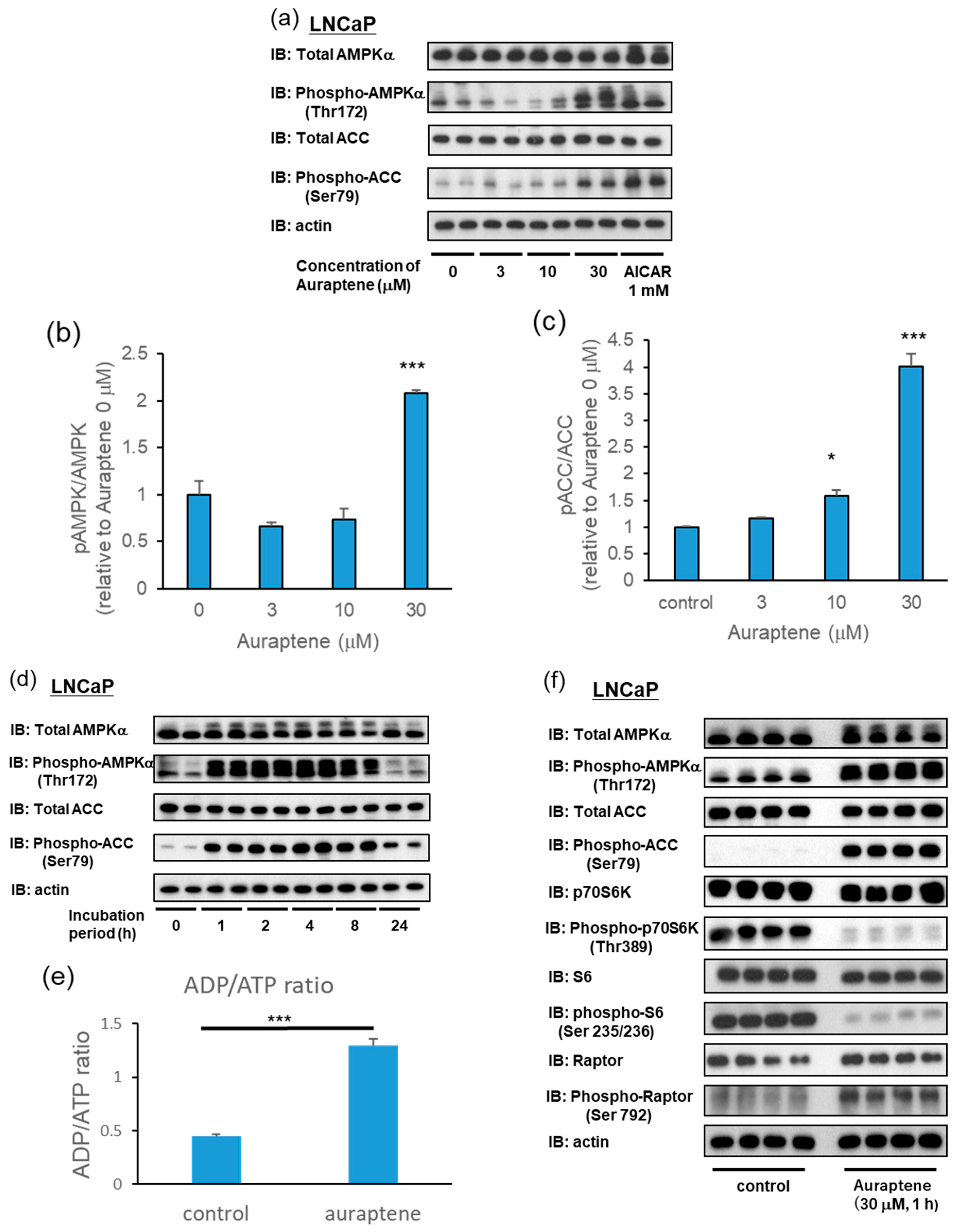
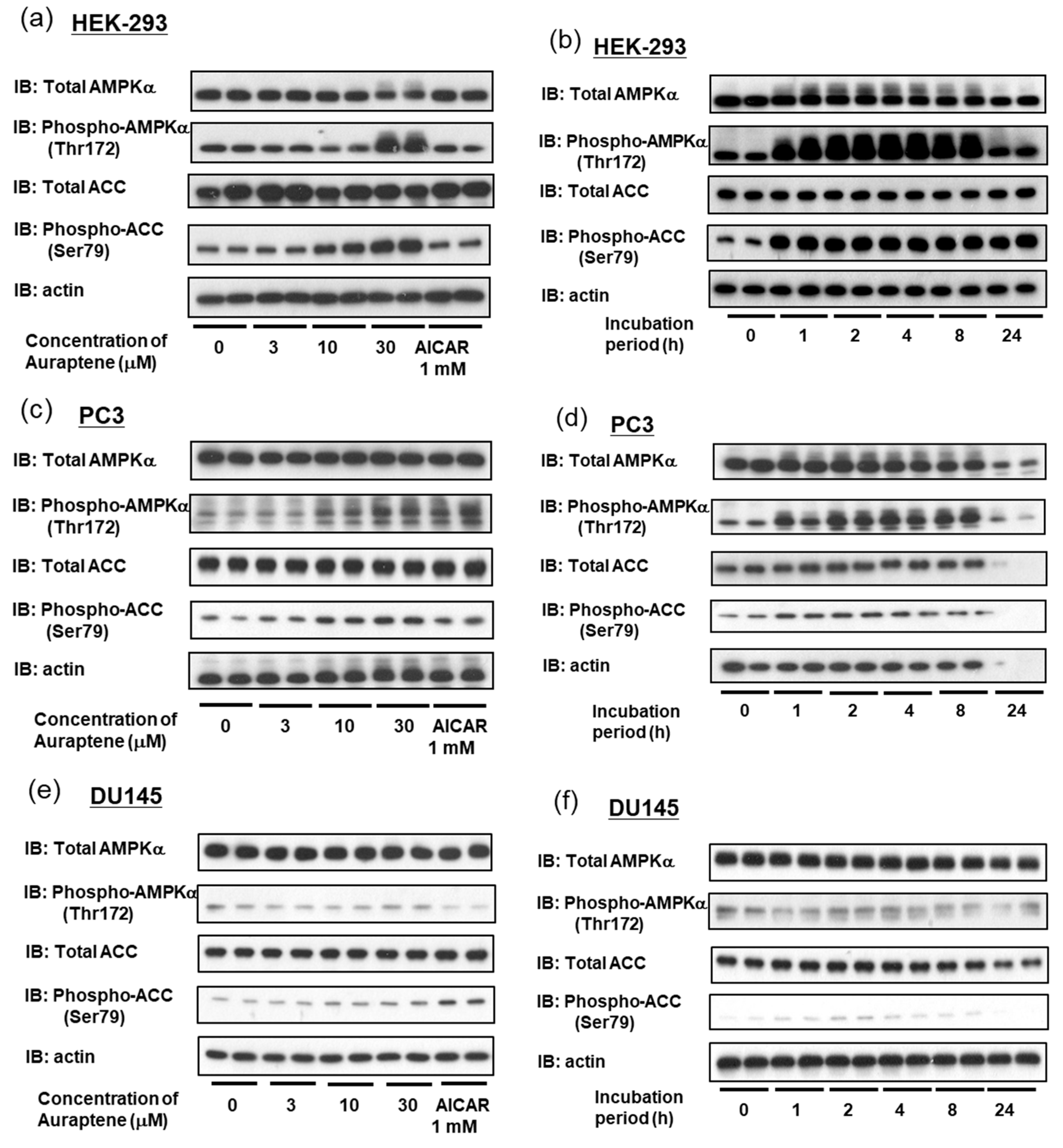
2.2. Auraptene Induces AMPK Activation
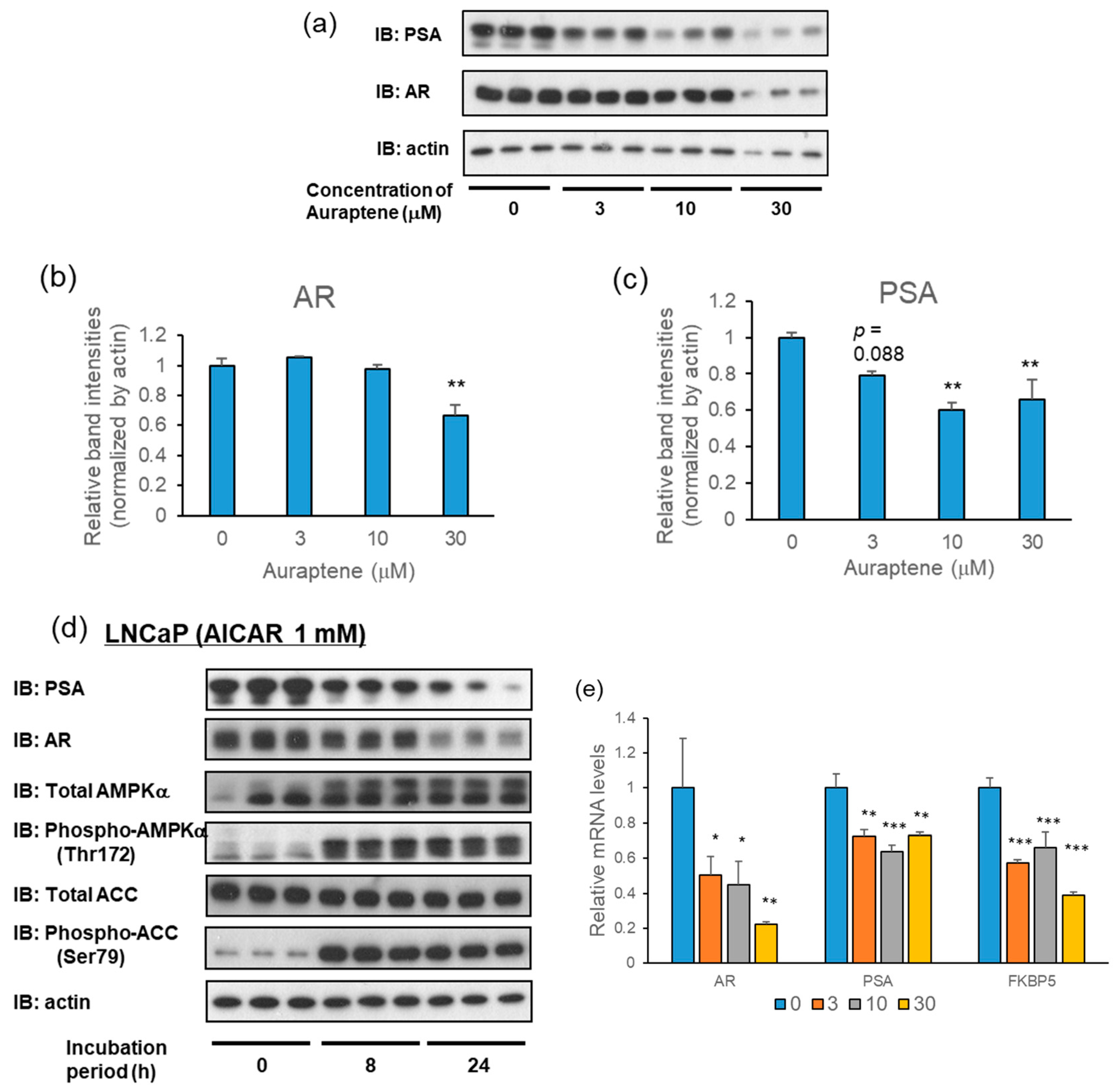
2.3. Auraptene Attenuates the Expressions of AR and PSA in LNCaP Cells
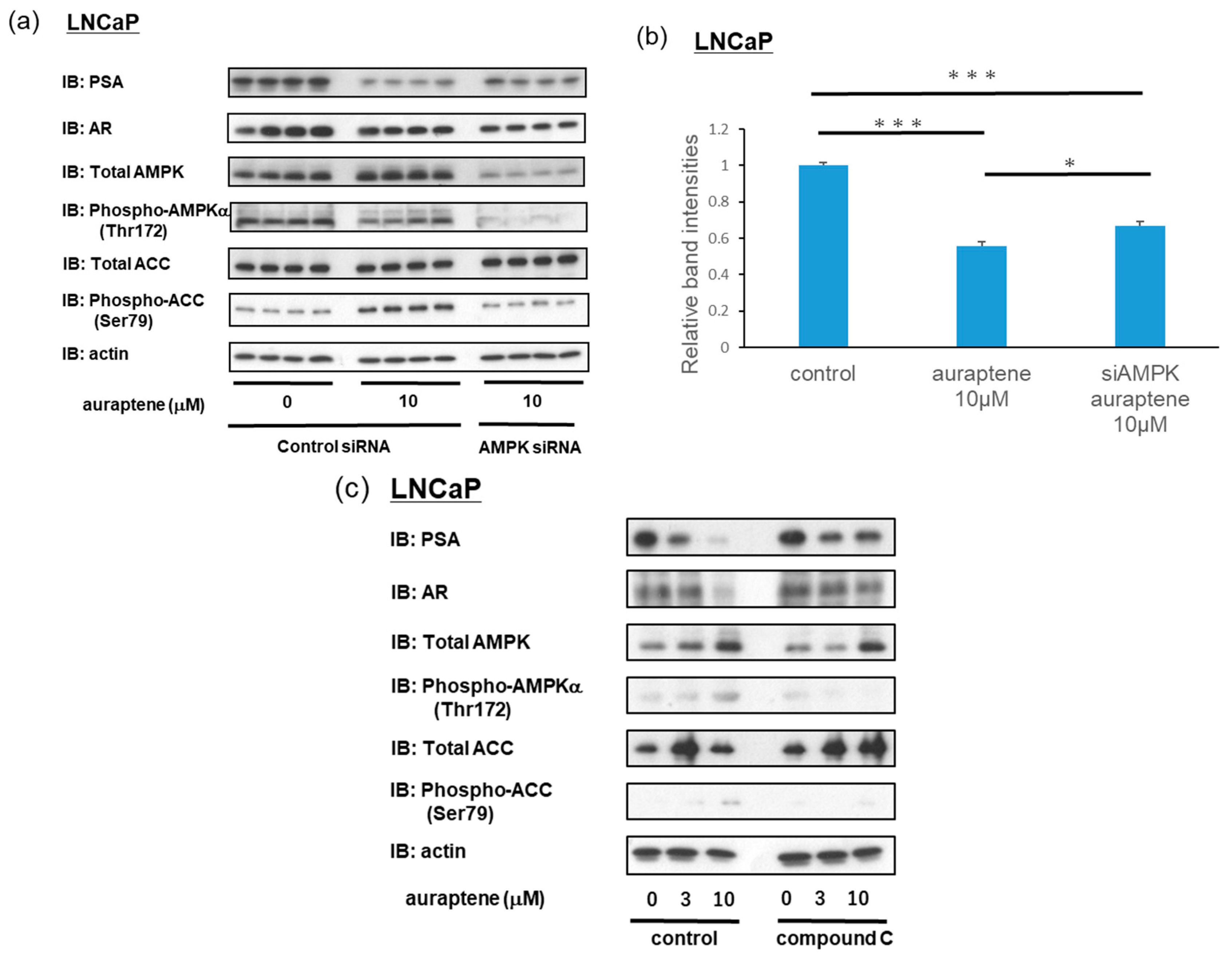
2.4. Suppression of AR Target Gene PSA by Auraptene in LNCaP Cells Is at Least Partially Mediated via AMPK Activation
2.5. Auraptene Induces AMPK Activation through the LKB1 Pathway
3. Discussion
4. Materials and Methods
4.1. Cell Culture
4.2. Reagents
4.3. ADP/ATP Assay
4.4. CCK-8 Assay
4.5. Wound Healing Assay
4.6. RNAi Interference
4.7. Real-Time Quantitative PCR
- Primers for human GAPDH: forward, 5′-GGC CTC CAA GGA GTA AGA CC-3′, reverse, 5′-AGG GGT CTA CAT GGC AAC TG-3′.
- Primers for human AR: forward, 5′-GGT GAG CAG AGT GCC CTA TC-3′, reverse, 5′-GAA GAC CTT GCA GCT TCC AC-3′.
- Primers for human PSA: forward, 5′-TCA CAG CTG CCC ACT GCA TCA-3′, reverse, 5′-AGG TCG TGG CTG GAG TCA TC-3′.
- Primers for human FKBP5: forward, 5′-AGG CTG CAA GAC TGC AGA TC-3′, reverse, 5′-CTT GCC CAT TGC TTT ATT GG-3′.
4.8. Immunoblot Analysis
4.9. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- A Website of World Health Organization. Available online: https://www.who.int/news-room/fact-sheets/detail/cancer) (accessed on 1 August 2023).
- Litwin, M.S.; Tan, H.J. The Diagnosis and Treatment of Prostate Cancer: A Review. JAMA 2017, 317, 2532–2542. [Google Scholar] [CrossRef] [PubMed]
- Sekhoacha, M.; Riet, K.; Motloung, P.; Gumenku, L.; Adegoke, A.; Mashele, S. Prostate Cancer Review: Genetics, Diagnosis, Treatment Options, and Alternative Approaches. Molecules 2022, 27, 5730. [Google Scholar] [CrossRef] [PubMed]
- Perera, S.; McDonald, J.; Williams, I.; O’Brien, J.; Murphy, D.; Lawrentschuk, N. Active surveillance versus nonradical treatment for low-risk men with prostate cancer: A review. Prostate Int. 2022, 10, 117–122. [Google Scholar] [CrossRef] [PubMed]
- Russell, F.M.; Hardie, D.G. AMP-Activated Protein Kinase: Do We Need Activators or Inhibitors to Treat or Prevent Cancer? Int. J. Mol. Sci. 2020, 22, 186. [Google Scholar] [CrossRef] [PubMed]
- Mihaylova, M.M.; Shaw, R.J. The AMPK signalling pathway coordinates cell growth, autophagy and metabolism. Nat. Cell Biol. 2011, 13, 1016–1023. [Google Scholar] [CrossRef]
- Inoki, K.; Zhu, T.; Guan, K.L. TSC2 mediates cellular energy response to control cell growth and survival. Cell 2003, 115, 577–590. [Google Scholar] [CrossRef]
- Gwinn, D.M.; Shackelford, D.B.; Egan, D.F.; Mihaylova, M.M.; Mery, A.; Vasquez, D.S.; Turk, B.E.; Shaw, R.J. AMPK phosphorylation of raptor mediates a metabolic checkpoint. Mol. Cell 2008, 30, 214–226. [Google Scholar] [CrossRef]
- Herzig, S.; Shaw, R.J. AMPK: Guardian of metabolism and mitochondrial homeostasis. Nat. Rev. Mol. Cell Biol. 2018, 19, 121–135. [Google Scholar] [CrossRef]
- Sohretoglu, D.; Zhang, C.; Luo, J.; Huang, S. ReishiMax inhibits mTORC1/2 by activating AMPK and inhibiting IGFR/PI3K/Rheb in tumor cells. Signal Transduct. Target. Ther. 2019, 4, 21. [Google Scholar] [CrossRef]
- Van Nostrand, J.L.; Hellberg, K.; Luo, E.C.; Van Nostrand, E.L.; Dayn, A.; Yu, J.; Shokhirev, M.N.; Dayn, Y.; Yeo, G.W.; Shaw, R.J. AMPK regulation of Raptor and TSC2 mediate metformin effects on transcriptional control of anabolism and inflammation. Genes Dev. 2020, 34, 1330–1344. [Google Scholar] [CrossRef]
- Imamura, K.; Ogura, T.; Kishimoto, A.; Kaminishi, M.; Esumi, H. Cell cycle regulation via p53 phosphorylation by a 5’-AMP activated protein kinase activator, 5-aminoimidazole-4-carboxamide-1-beta-D-ribofuranoside, in a human hepatocellular carcinoma cell line. Biochem. Biophys. Res. Commun. 2001, 287, 562–567. [Google Scholar] [CrossRef] [PubMed]
- He, P.; Li, Z.; Xu, F.; Ru, G.; Huang, Y.; Lin, E.; Peng, S. AMPK Activity Contributes to G2 Arrest and DNA Damage Decrease via p53/p21 Pathways in Oxidatively Damaged Mouse Zygotes. Front. Cell Dev. Biol. 2020, 8, 539485. [Google Scholar] [CrossRef]
- Bibak, B.; Shakeri, F.; Barreto, G.E.; Keshavarzi, Z.; Sathyapalan, T.; Sahebkar, A. A review of the pharmacological and therapeutic effects of auraptene. Biofactors 2019, 45, 867–879. [Google Scholar] [CrossRef] [PubMed]
- Tayarani-Najaran, Z.; Tayarani-Najaran, N.; Eghbali, S. A Review of Auraptene as an Anticancer Agent. Front. Pharmacol. 2021, 12, 698352. [Google Scholar] [CrossRef] [PubMed]
- Afshari, A.R.; Jalili-Nik, M.; Soukhtanloo, M.; Ghorbani, A.; Sadeghnia, H.R.; Mollazadeh, H.; Karimi Roshan, M.; Rahmani, F.; Sabri, H.; Vahedi, M.M.; et al. Auraptene-induced cytotoxicity mechanisms in human malignant glioblastoma (U87) cells: Role of reactive oxygen species (ROS). EXCLI J. 2019, 18, 576–590. [Google Scholar] [CrossRef]
- Jamialahmadi, K.; Salari, S.; Alamolhodaei, N.S.; Avan, A.; Gholami, L.; Karimi, G. Auraptene Inhibits Migration and Invasion of Cervical and Ovarian Cancer Cells by Repression of Matrix Metalloproteinasas 2 and 9 Activity. J. Pharmacopunct. 2018, 21, 177–184. [Google Scholar] [CrossRef]
- Tang, M.; Ogawa, K.; Asamoto, M.; Hokaiwado, N.; Seeni, A.; Suzuki, S.; Takahashi, S.; Tanaka, T.; Ichikawa, K.; Shirai, T. Protective effects of citrus nobiletin and auraptene in transgenic rats developing adenocarcinoma of the prostate (TRAP) and human prostate carcinoma cells. Cancer Sci. 2007, 98, 471–477. [Google Scholar] [CrossRef]
- Lee, J.C.; Shin, E.A.; Kim, B.; Kim, B.I.; Chitsazian-Yazdi, M.; Iranshahi, M.; Kim, S.H. Auraptene Induces Apoptosis via Myeloid Cell Leukemia 1-Mediated Activation of Caspases in PC3 and DU145 Prostate Cancer Cells. Phytother. Res. 2017, 31, 891–898. [Google Scholar] [CrossRef]
- Akashi, S.; Morita, A.; Mochizuki, Y.; Shibuya, F.; Kamei, Y.; Miura, S. Citrus hassaku Extract Powder Increases Mitochondrial Content and Oxidative Muscle Fibers by Upregulation of PGC-1α in Skeletal Muscle. Nutrients 2021, 13, 497. [Google Scholar] [CrossRef]
- Hasei, S.; Yamamotoya, T.; Nakatsu, Y.; Ohata, Y.; Itoga, S.; Nonaka, Y.; Matsunaga, Y.; Sakoda, H.; Fujishiro, M.; Kushiyama, A.; et al. Carnosic Acid and Carnosol Activate AMPK, Suppress Expressions of Gluconeogenic and Lipogenic Genes, and Inhibit Proliferation of HepG2 Cells. Int. J. Mol. Sci. 2021, 22, 4040. [Google Scholar] [CrossRef]
- Hardie, D.G.; Ross, F.A.; Hawley, S.A. AMPK: A nutrient and energy sensor that maintains energy homeostasis. Nat. Rev. Mol. Cell Biol. 2012, 13, 251–262. [Google Scholar] [CrossRef]
- Dengler, F. Activation of AMPK under Hypoxia: Many Roads Leading to Rome. Int. J. Mol. Sci. 2020, 21, 2428. [Google Scholar] [CrossRef] [PubMed]
- Shen, M.; Zhang, Z.; Ratnam, M.; Dou, Q.P. The interplay of AMP-activated protein kinase and androgen receptor in prostate cancer cells. J. Cell Physiol. 2014, 229, 688–695. [Google Scholar] [CrossRef] [PubMed]
- Zadra, G.; Photopoulos, C.; Tyekucheva, S.; Heidari, P.; Weng, Q.P.; Fedele, G.; Liu, H.; Scaglia, N.; Priolo, C.; Sicinska, E.; et al. A novel direct activator of AMPK inhibits prostate cancer growth by blocking lipogenesis. EMBO Mol. Med. 2014, 6, 519–538. [Google Scholar] [CrossRef] [PubMed]
- Jurmeister, S.; Ramos-Montoya, A.; Neal, D.E.; Fryer, L.G. Transcriptomic analysis reveals inhibition of androgen receptor activity by AMPK in prostate cancer cells. Oncotarget 2014, 5, 3785–3799. [Google Scholar] [CrossRef] [PubMed]
- Karacosta, L.G.; Foster, B.A.; Azabdaftari, G.; Feliciano, D.M.; Edelman, A.M. A regulatory feedback loop between Ca2+/calmodulin-dependent protein kinase kinase 2 (CaMKK2) and the androgen receptor in prostate cancer progression. J. Biol. Chem. 2012, 287, 24832–24843. [Google Scholar] [CrossRef]
- Pulliam, T.L.; Goli, P.; Awad, D.; Lin, C.; Wilkenfeld, S.R.; Frigo, D.E. Regulation and role of CAMKK2 in prostate cancer. Nat. Rev. Urol. 2022, 19, 367–380. [Google Scholar] [CrossRef]
- Coutinho, I.; Day, T.K.; Tilley, W.D.; Selth, L.A. Androgen receptor signaling in castration-resistant prostate cancer: A lesson in persistence. Endocr. Relat. Cancer 2016, 23, T179–T197. [Google Scholar] [CrossRef]
- Vakili, T.; Iranshahi, M.; Arab, H.; Riahi, B.; Roshan, N.M.; Karimi, G. Safety evaluation of auraptene in rats in acute and subacute toxicity studies. Regul. Toxicol. Pharmacol. 2017, 91, 159–164. [Google Scholar] [CrossRef]
- Li, Y.; Xu, S.; Mihaylova, M.M.; Zheng, B.; Hou, X.; Jiang, B.; Park, O.; Luo, Z.; Lefai, E.; Shyy, J.Y.; et al. AMPK phosphorylates and inhibits SREBP activity to attenuate hepatic steatosis and atherosclerosis in diet-induced insulin-resistant mice. Cell Metab. 2011, 13, 376–388. [Google Scholar] [CrossRef]
- Shimano, H.; Sato, R. SREBP-regulated lipid metabolism: Convergent physiology-divergent pathophysiology. Nat. Rev. Endocrinol. 2017, 13, 710–730. [Google Scholar] [CrossRef]
- Han, Y.; Hu, Z.; Cui, A.; Liu, Z.; Ma, F.; Xue, Y.; Liu, Y.; Zhang, F.; Zhao, Z.; Yu, Y.; et al. Post-translational regulation of lipogenesis via AMPK-dependent phosphorylation of insulin-induced gene. Nat. Commun. 2019, 10, 623. [Google Scholar] [CrossRef] [PubMed]
- Fang, C.; Pan, J.; Qu, N.; Lei, Y.; Han, J.; Zhang, J.; Han, D. The AMPK pathway in fatty liver disease. Front. Physiol. 2022, 13, 970292. [Google Scholar] [CrossRef] [PubMed]
- Swinnen, J.V.; Ulrix, W.; Heyns, W.; Verhoeven, G. Coordinate regulation of lipogenic gene expression by androgens: Evidence for a cascade mechanism involving sterol regulatory element binding proteins. Proc. Natl. Acad. Sci. USA 1997, 94, 12975–12980. [Google Scholar] [CrossRef] [PubMed]
- Swinnen, J.V.; Heemers, H.; van de Sande, T.; de Schrijver, E.; Brusselmans, K.; Heyns, W.; Verhoeven, G. Androgens, lipogenesis and prostate cancer. J. Steroid Biochem. Mol. Biol. 2004, 92, 273–279. [Google Scholar] [CrossRef] [PubMed]
- Pizer, E.S.; Pflug, B.R.; Bova, G.S.; Han, W.F.; Udan, M.S.; Nelson, J.B. Increased fatty acid synthase as a therapeutic target in androgen-independent prostate cancer progression. Prostate 2001, 47, 102–110. [Google Scholar] [CrossRef] [PubMed]
- Swinnen, J.V.; Roskams, T.; Joniau, S.; Van Poppel, H.; Oyen, R.; Baert, L.; Heyns, W.; Verhoeven, G. Overexpression of fatty acid synthase is an early and common event in the development of prostate cancer. Int. J. Cancer 2002, 98, 19–22. [Google Scholar] [CrossRef]
- Vara-Ciruelos, D.; Russell, F.M.; Hardie, D.G. The strange case of AMPK and cancer: Dr Jekyll or Mr Hyde? (†). Open Biol. 2019, 9, 190099. [Google Scholar] [CrossRef]
- Khan, A.S.; Frigo, D.E. A spatiotemporal hypothesis for the regulation, role, and targeting of AMPK in prostate cancer. Nat. Rev. Urol. 2017, 14, 164–180. [Google Scholar] [CrossRef]
- Penfold, L.; Woods, A.; Pollard, A.E.; Arizanova, J.; Pascual-Navarro, E.; Muckett, P.J.; Dore, M.H.; Montoya, A.; Whilding, C.; Fets, L.; et al. AMPK activation protects against prostate cancer by inducing a catabolic cellular state. Cell Rep. 2023, 42, 112396. [Google Scholar] [CrossRef]
- Shiran, M.R.; Mahmoudian, E.; Ajami, A.; Hosseini, S.M.; Khojasteh, A.; Rashidi, M.; Maroufi, N.F. Effect of Auraptene on angiogenesis in Xenograft model of breast cancer. Horm. Mol. Biol. Clin. Investig. 2021, 43, 7–14. [Google Scholar] [CrossRef] [PubMed]
- Krishnan, P.; Kleiner-Hancock, H. Effects of Auraptene on IGF-1 Stimulated Cell Cycle Progression in the Human Breast Cancer Cell Line, MCF-7. Int. J. Breast Cancer 2012, 2012, 502092. [Google Scholar] [CrossRef] [PubMed]
- Kanda, Y. Investigation of the freely available easy-to-use software ‘EZR’ for medical statistics. Bone Marrow Transplant. 2013, 48, 452–458. [Google Scholar] [CrossRef] [PubMed]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Akasaka, Y.; Hasei, S.; Ohata, Y.; Kanna, M.; Nakatsu, Y.; Sakoda, H.; Fujishiro, M.; Kushiyama, A.; Ono, H.; Matsubara, A.; et al. Auraptene Enhances AMP-Activated Protein Kinase Phosphorylation and Thereby Inhibits the Proliferation, Migration and Expression of Androgen Receptors and Prostate-Specific Antigens in Prostate Cancer Cells. Int. J. Mol. Sci. 2023, 24, 16011. https://doi.org/10.3390/ijms242116011
Akasaka Y, Hasei S, Ohata Y, Kanna M, Nakatsu Y, Sakoda H, Fujishiro M, Kushiyama A, Ono H, Matsubara A, et al. Auraptene Enhances AMP-Activated Protein Kinase Phosphorylation and Thereby Inhibits the Proliferation, Migration and Expression of Androgen Receptors and Prostate-Specific Antigens in Prostate Cancer Cells. International Journal of Molecular Sciences. 2023; 24(21):16011. https://doi.org/10.3390/ijms242116011
Chicago/Turabian StyleAkasaka, Yasuyuki, Shun Hasei, Yukino Ohata, Machi Kanna, Yusuke Nakatsu, Hideyuki Sakoda, Midori Fujishiro, Akifumi Kushiyama, Hiraku Ono, Akio Matsubara, and et al. 2023. "Auraptene Enhances AMP-Activated Protein Kinase Phosphorylation and Thereby Inhibits the Proliferation, Migration and Expression of Androgen Receptors and Prostate-Specific Antigens in Prostate Cancer Cells" International Journal of Molecular Sciences 24, no. 21: 16011. https://doi.org/10.3390/ijms242116011