
Investigating the Metabolism of Plants Germinated in Heavy Water, D2O, and H218O-Enriched Media Using High-Resolution Mass Spectrometry
 , , , , and
, , , , and
Abstract
:1. Introduction
2. Results
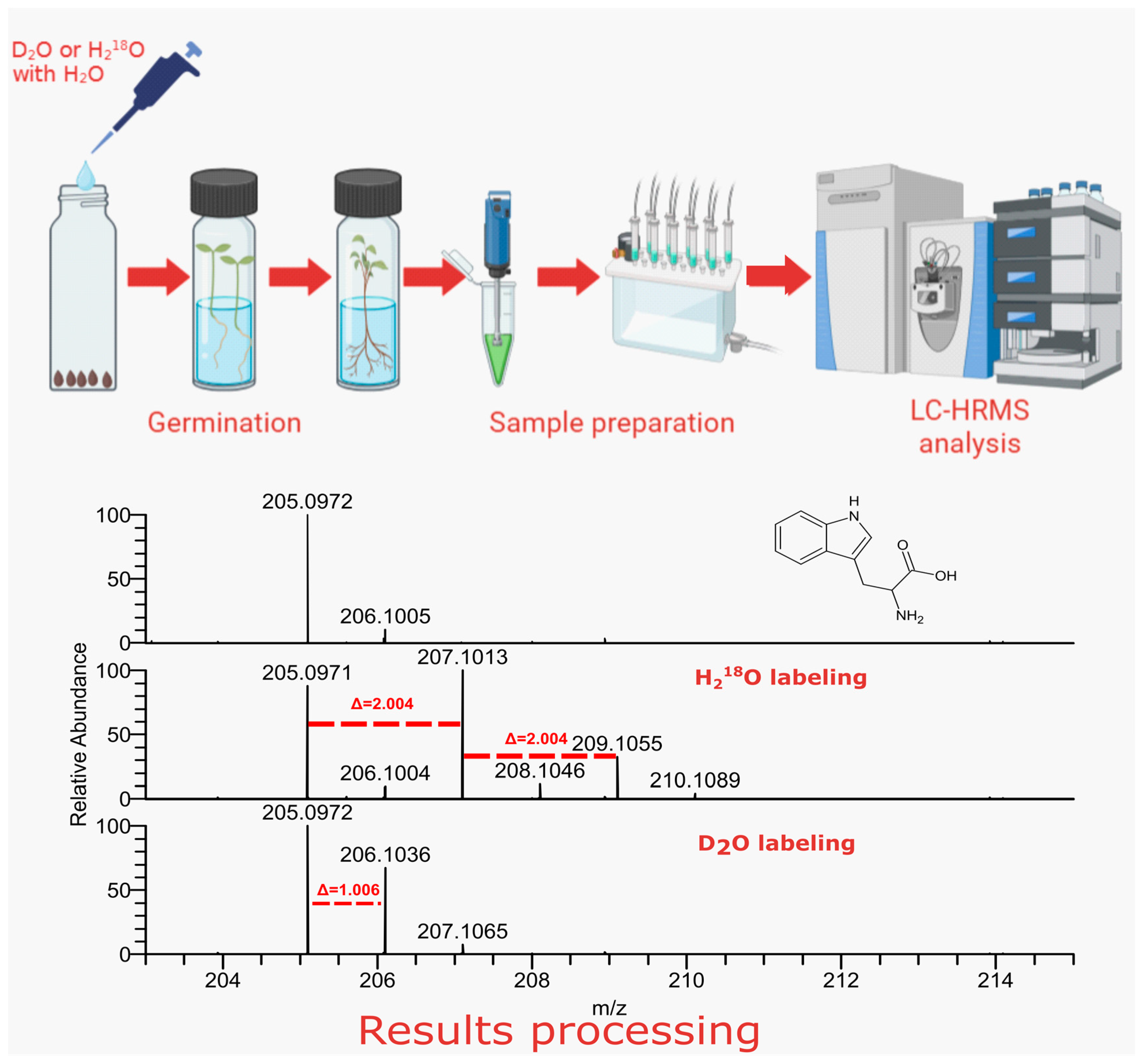
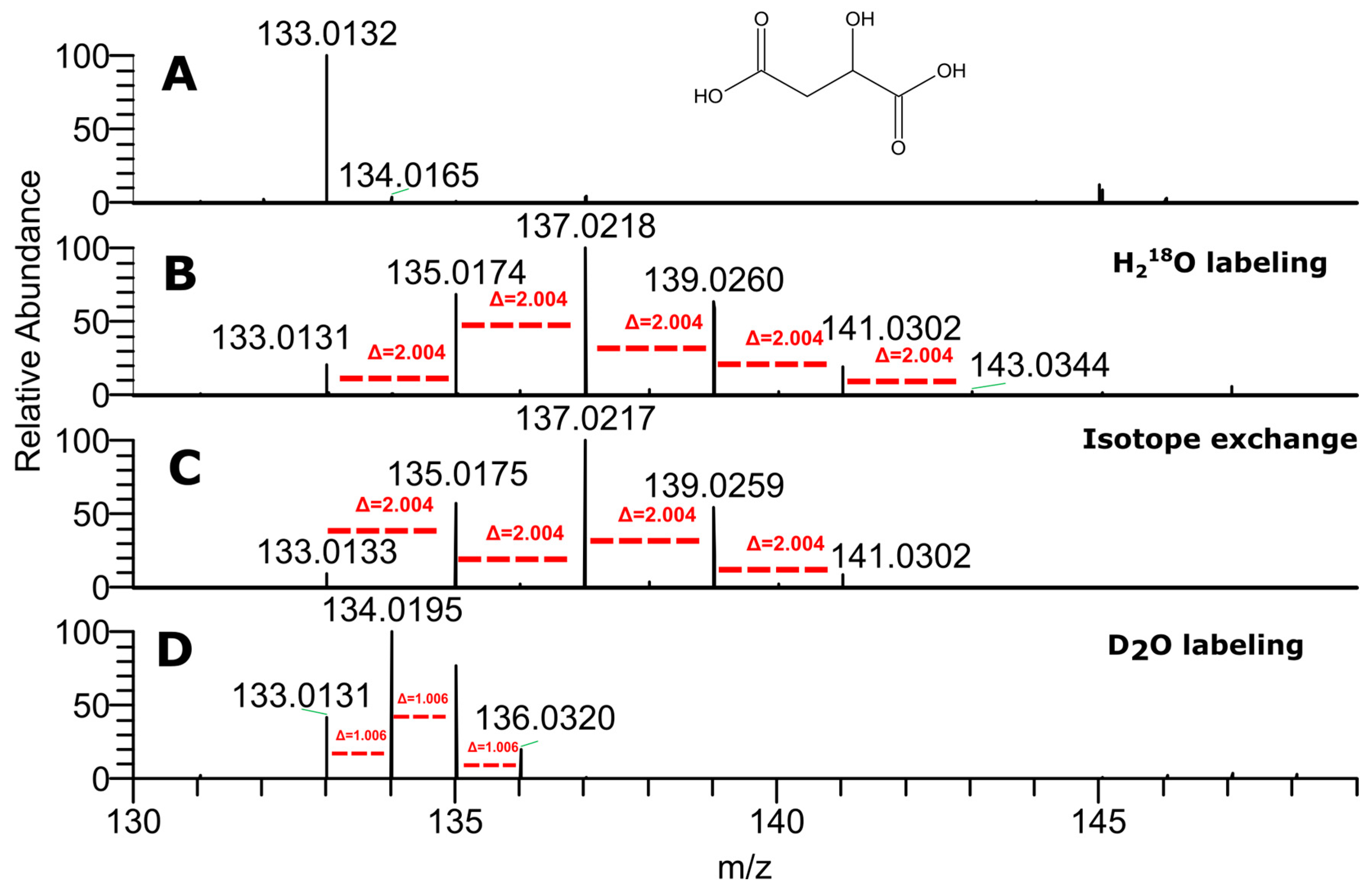
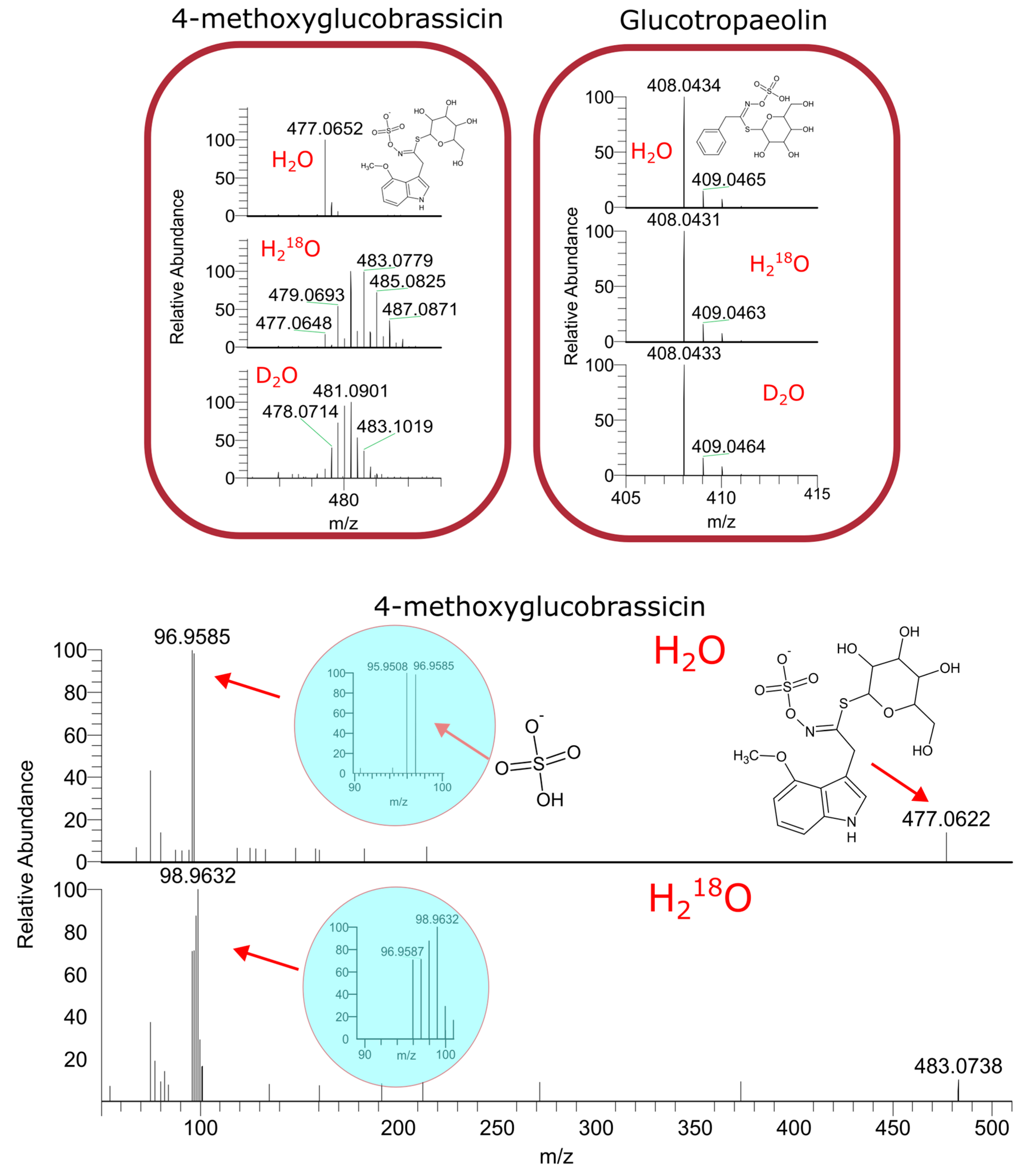
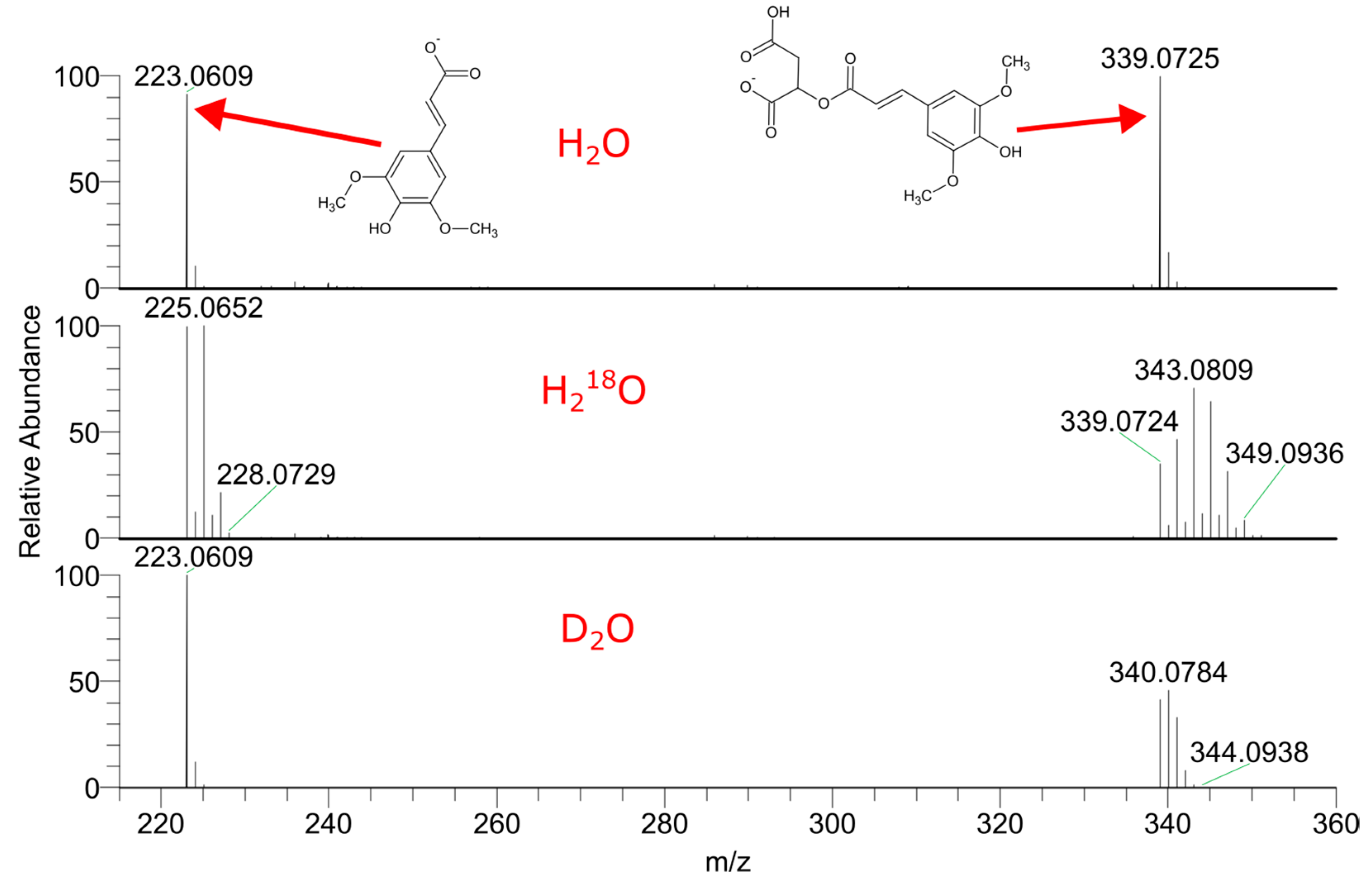
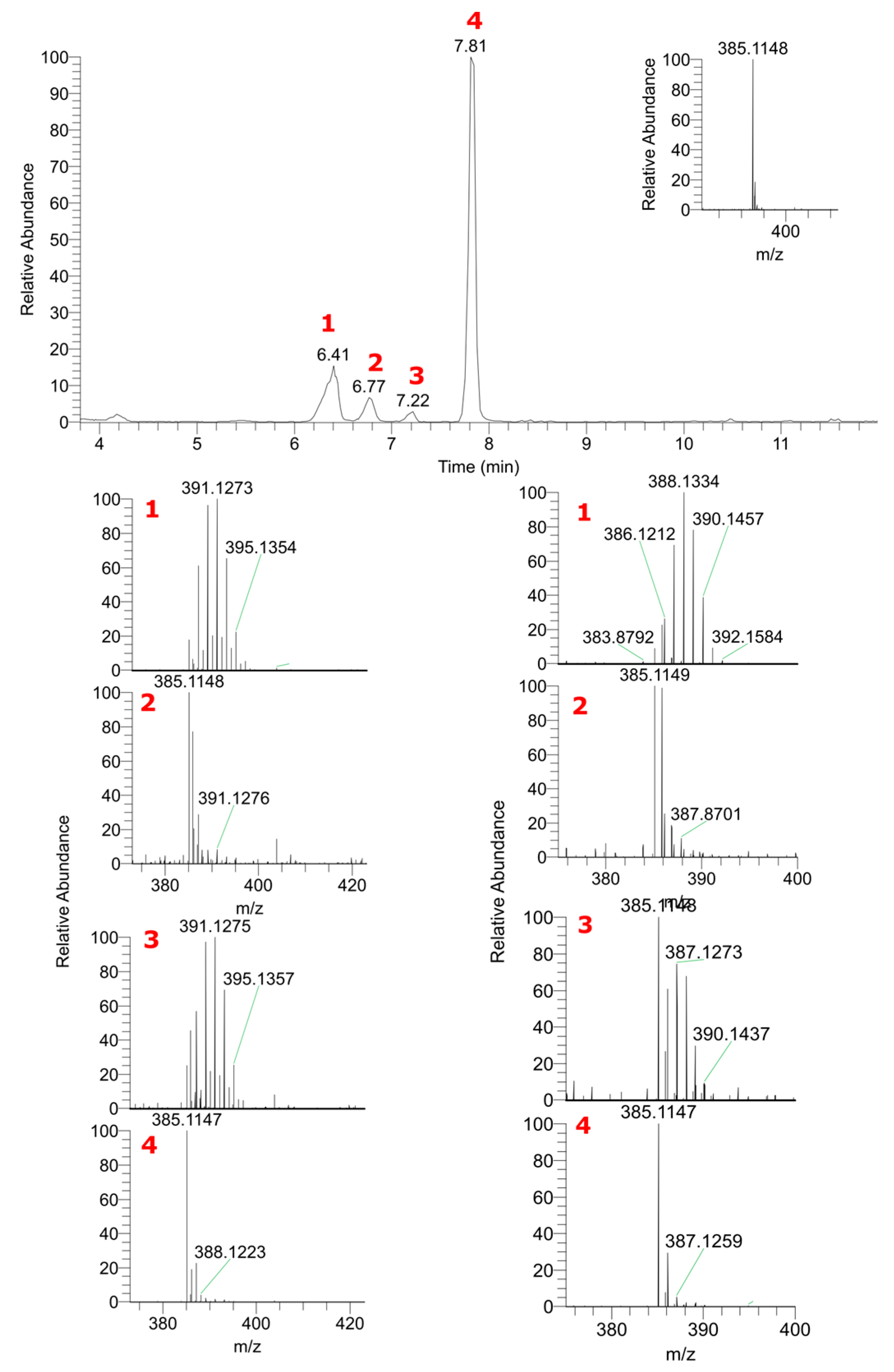
2.1. LC–HRMS Analysis of Labeled Plant Extracts
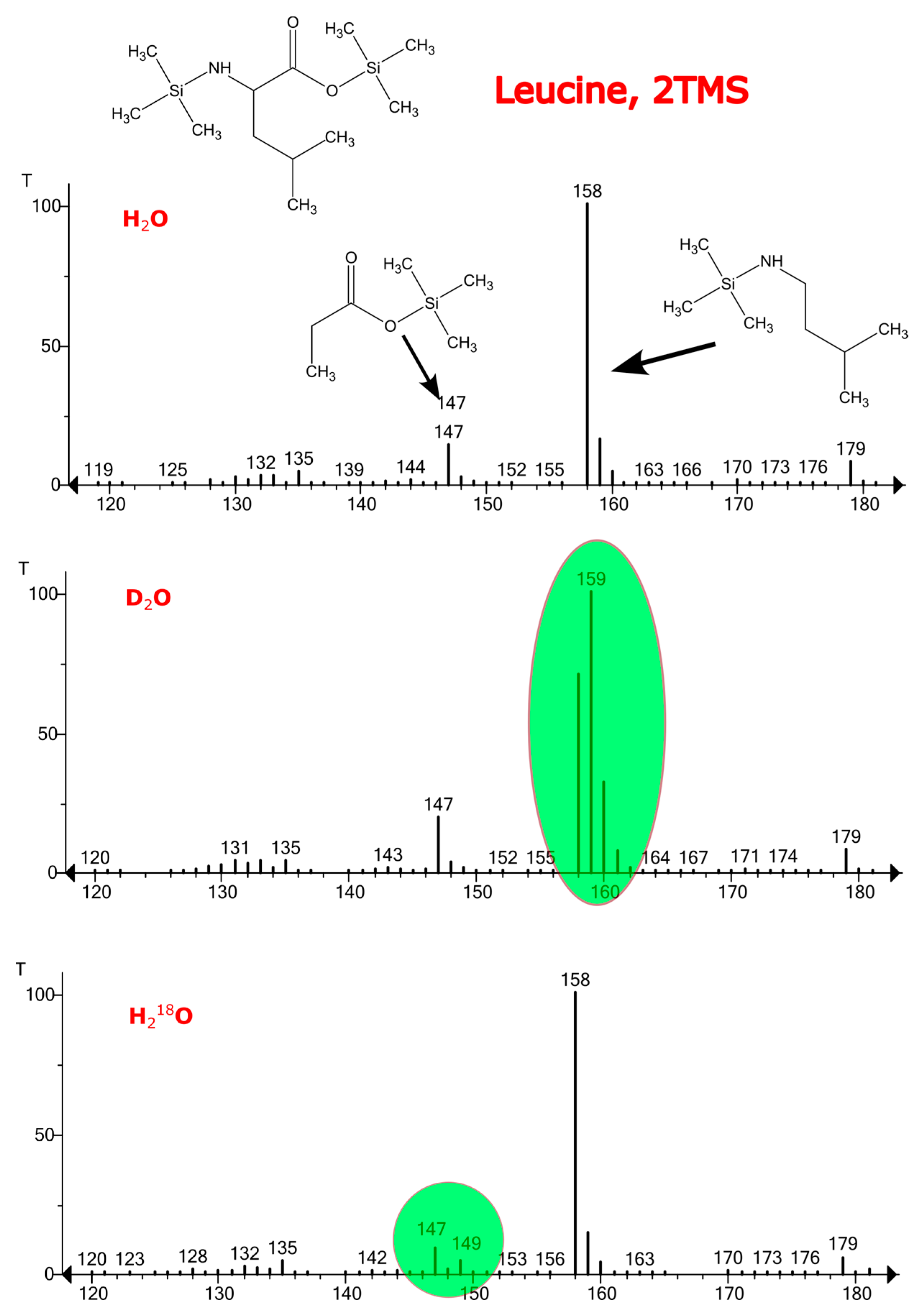
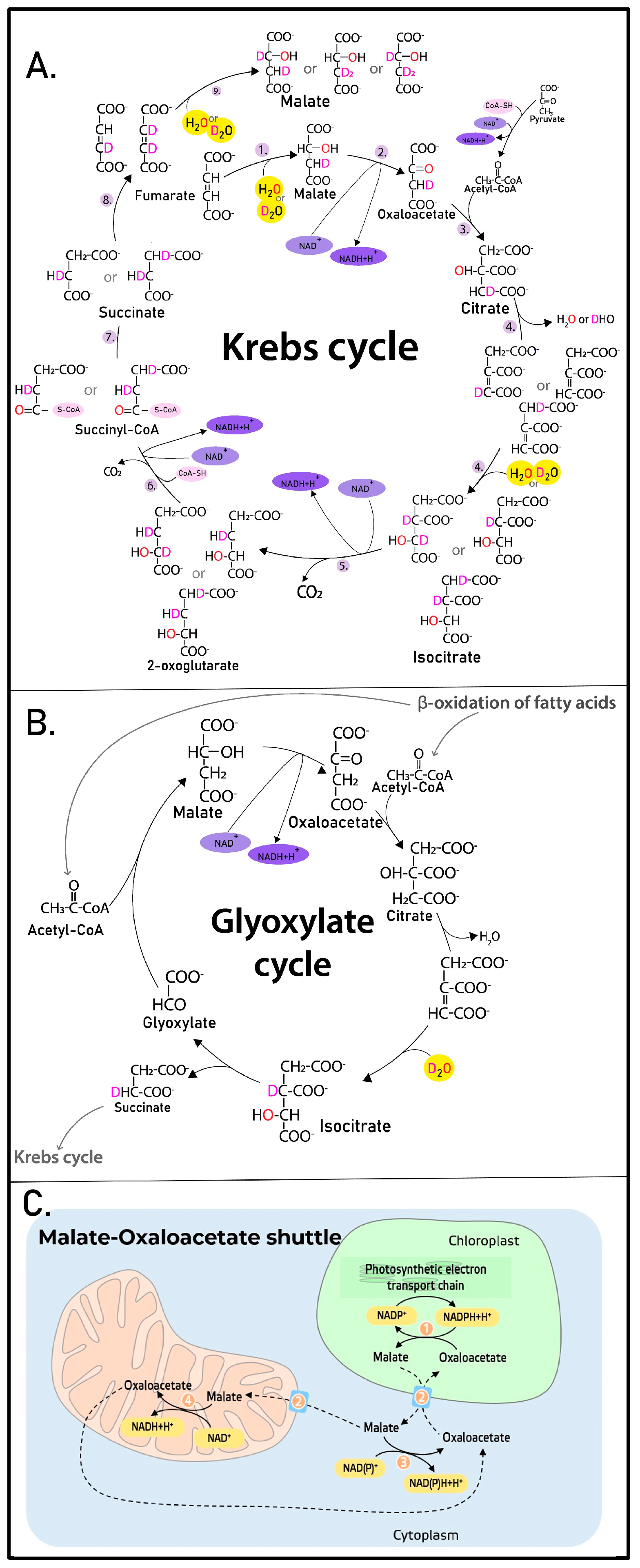
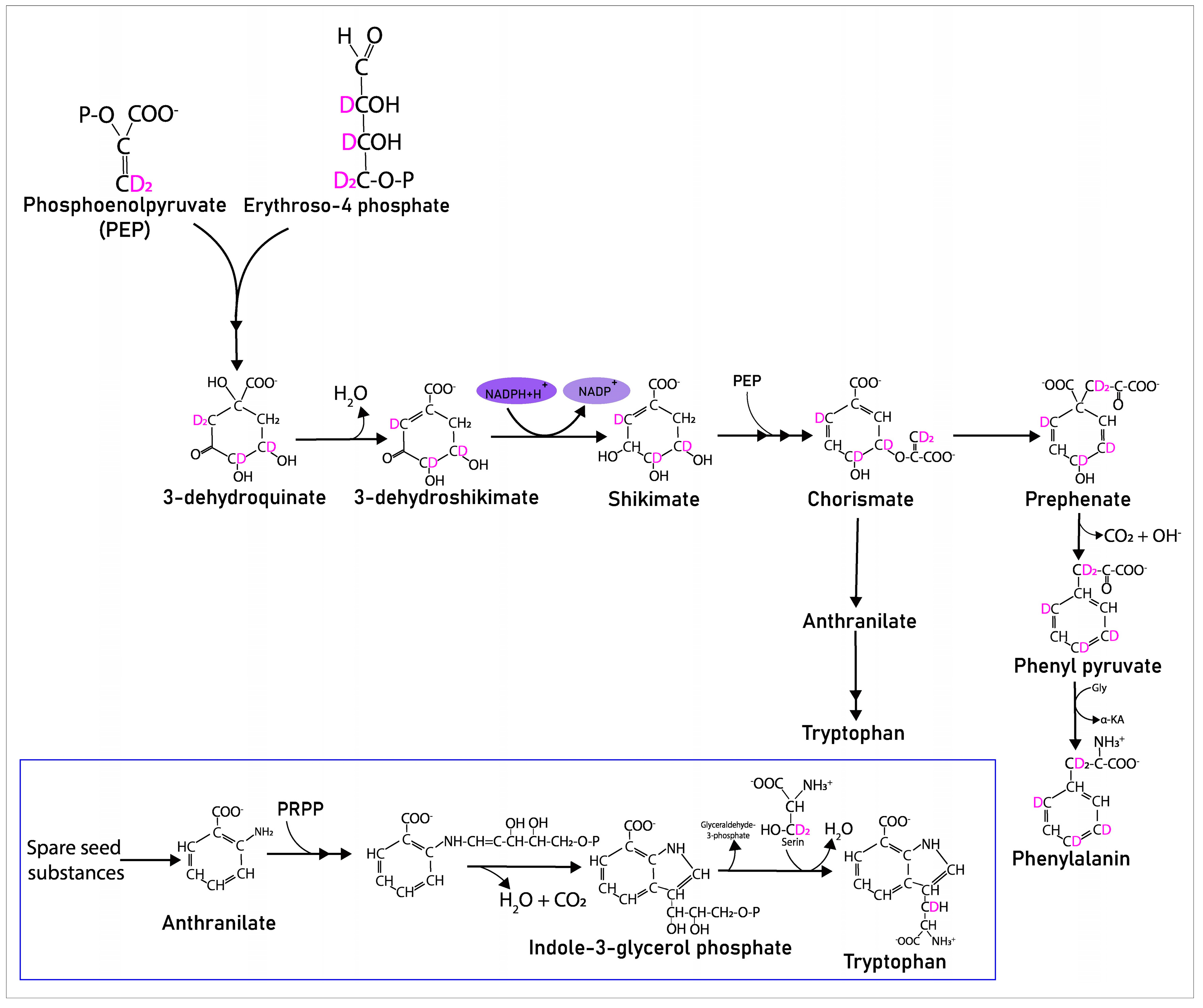
2.2. The Contribution of the Isotope Exchange Reaction
3. Discussion
4. Materials and Methods
4.1. Plant Germination and Sample Preparation
4.2. LC–MS Analysis
4.3. GC–MS Analysis
4.4. Data Processing
4.5. Data Availability
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Kisiel, A.; Krzemińska, A.; Cembrowska-Lech, D.; Miller, T. Data Science and Plant Metabolomics. Metabolites 2023, 13, 454. [Google Scholar] [CrossRef]
- Zhu, M.Z.; Chen, G.L.; Wu, J.L.; Li, N.; Liu, Z.H.; Guo, M.Q. Recent development in mass spectrometry and its hyphenated techniques for the analysis of medicinal plants. Phytochem. Anal. 2018, 29, 365–374. [Google Scholar] [CrossRef]
- Ruttkies, C.; Schymanski, E.L.; Strehmel, N.; Hollender, J.; Neumann, S.; Williams, A.J.; Krauss, M. Supporting non-target identification by adding hydrogen deuterium exchange MS/MS capabilities to MetFrag. Anal. Bioanal. Chem. 2019, 411, 4683–4700. [Google Scholar] [CrossRef] [PubMed]
- Kostyukevich, Y.; Sosnin, S.; Osipenko, S.; Kovaleva, O.; Rumiantseva, L.; Kireev, A.; Zherebker, A.; Fedorov, M.; Nikolaev, E.N. PyFragMS—A Web Tool for the Investigation of the Collision-Induced Fragmentation Pathways. ACS Omega 2022, 7, 9710–9719. [Google Scholar] [CrossRef] [PubMed]
- Osipenko, S.; Bashkirova, I.; Sosnin, S.; Kovaleva, O.; Fedorov, M.; Nikolaev, E.; Kostyukevich, Y. Machine learning to predict retention time of small molecules in nano-HPLC. Anal. Bioanal. Chem. 2020, 412, 7767–7776. [Google Scholar] [CrossRef] [PubMed]
- Osipenko, S.; Botashev, K.; Nikolaev, E.; Kostyukevich, Y. Transfer learning for small molecule retention predictions. J. Chromatogr. A 2021, 1644, 462119. [Google Scholar] [CrossRef]
- Osipenko, S.; Nikolaev, E.; Kostyukevich, Y. Retention Time Prediction with Message-Passing Neural Networks. Separations 2022, 9, 291. [Google Scholar] [CrossRef]
- Capellades, J.; Navarro, M.; Samino, S.; Garcia-Ramirez, M.; Hernandez, C.; Simo, R.; Vinaixa, M.; Yanes, O. geoRge: A computational tool to detect the presence of stable isotope labeling in LC/MS-based untargeted metabolomics. Anal. Chem. 2016, 88, 621–628. [Google Scholar] [CrossRef]
- Klein, S.; Heinzle, E. Isotope labeling experiments in metabolomics and fluxomics. Wiley Interdiscip. Rev. Syst. Biol. Med. 2012, 4, 261–272. [Google Scholar] [CrossRef]
- Bueschl, C.; Krska, R.; Kluger, B.; Schuhmacher, R. Isotopic labeling-assisted metabolomics using LC–MS. Anal. Bioanal. Chem. 2013, 405, 27–33. [Google Scholar] [CrossRef]
- Pang, Q.; Zhang, T.; Wang, Y.; Kong, W.; Guan, Q.; Yan, X.; Chen, S. Metabolomics of early stage plant cell–microbe interaction using stable isotope labeling. Front. Plant Sci. 2018, 9, 760. [Google Scholar] [CrossRef]
- Huang, X.; Regnier, F.E. Differential metabolomics using stable isotope labeling and two-dimensional gas chromatography with time-of-flight mass spectrometry. Anal. Chem. 2008, 80, 107–114. [Google Scholar] [CrossRef] [PubMed]
- Pinnick, K.E.; Gunn, P.J.; Hodson, L. Measuring human lipid metabolism using deuterium labeling: In vivo and in vitro protocols. In Metabolic Signaling Methods and Protocols; Springer: Berlin/Heidelberg, Germany, 2019; pp. 83–96. [Google Scholar]
- Dufner, D.; Previs, S.F. Measuring in vivo metabolism using heavy water. Curr. Opin. Clin. Nutr. Metab. Care 2003, 6, 511–517. [Google Scholar] [CrossRef]
- Diraison, F.; Pachiaudi, C.; Beylot, M. In vivo measurement of plasma cholesterol and fatty acid synthesis with deuterated water: Determination of the average number of deuterium atoms incorporated. Metabolism 1996, 45, 817–821. [Google Scholar] [CrossRef] [PubMed]
- Castro-Perez, J.; Previs, S.F.; McLaren, D.G.; Shah, V.; Herath, K.; Bhat, G.; Johns, D.G.; Wang, S.-P.; Mitnaul, L.; Jensen, K. In vivo D2O labeling to quantify static and dynamic changes in cholesterol and cholesterol esters by high resolution LC/MS. J. Lipid Res. 2011, 52, 159–169. [Google Scholar] [CrossRef] [PubMed]
- Åstot, C.; Dolezal, K.; Moritz, T.; Sandberg, G. Deuterium in vivo labelling of cytokinins in Arabidopsis thaliana analysed by capillary liquid chromatography/frit-fast atom bombardment mass spectrometry. J. Mass Spectrom. 2000, 35, 13–22. [Google Scholar] [CrossRef]
- Tarkowski, P.; Floková, K.; Václavíková, K.; Jaworek, P.; Raus, M.; Nordström, A.; Novák, O.; Doležal, K.; Šebela, M.; Frébortová, J. An Improved in Vivo Deuterium Labeling Method for Measuring the Biosynthetic Rate of Cytokinins. Molecules 2010, 15, 9214–9229. [Google Scholar] [CrossRef]
- Pengelly, W.L.; Bandurski, R.S. Analysis of Indole-3-acetic Acid Metabolism in Zea mays Using Deuterium Oxide as a Tracer 1. Plant Physiol. 1983, 73, 445–449. [Google Scholar] [CrossRef]
- Lerma, C.; Hanson, A.D.; Rhodes, D. Oxygen-18 and Deuterium Labeling Studies of Choline Oxidation by Spinach and Sugar Beet 1. Plant Physiol. 1988, 88, 695–702. [Google Scholar] [CrossRef]
- Ippel, J.H.; Pouvreau, L.; Kroef, T.; Gruppen, H.; Versteeg, G.; van den Putten, P.; Struik, P.C.; van Mierlo, C.P.M. In vivo uniform 15N-isotope labelling of plants: Using the greenhouse for structural proteomics. Proteomics 2004, 4, 226–234. [Google Scholar] [CrossRef]
- Cegelski, L.; Schaefer, J. Glycine metabolism in intact leaves by in vivo 13C and 15N labeling. J. Biol. Chem. 2005, 280, 39238–39245. [Google Scholar] [CrossRef]
- Hasunuma, T.; Harada, K.; Miyazawa, S.-I.; Kondo, A.; Fukusaki, E.; Miyake, C. Metabolic turnover analysis by a combination of in vivo 13C-labelling from 13CO2 and metabolic profiling with CE-MS/MS reveals rate-limiting steps of the C3 photosynthetic pathway in Nicotiana tabacum leaves. J. Exp. Bot. 2010, 61, 1041–1051. [Google Scholar] [CrossRef]
- Cegelski, L.; Schaefer, J. NMR determination of photorespiration in intact leaves using in vivo 13CO2 labeling. J. Magn. Reson. 2006, 178, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Huege, J.; Sulpice, R.; Gibon, Y.; Lisec, J.; Koehl, K.; Kopka, J. GC-EI-TOF-MS analysis of in vivo carbon-partitioning into soluble metabolite pools of higher plants by monitoring isotope dilution after 13CO2 labelling. Phytochemistry 2007, 68, 2258–2272. [Google Scholar] [CrossRef] [PubMed]
- Kutzner, E.; Manukyan, A.; Eisenreich, W. Profiling of terpene metabolism in 13CO2-labelled Thymus transcaucasicus. Metabolomics 2014, 4, 2153–07691000129. [Google Scholar]
- Kikuchi, J.; Shinozaki, K.; Hirayama, T. Stable isotope labeling of Arabidopsis thaliana for an NMR-based metabolomics approach. Plant Cell Physiol. 2004, 45, 1099–1104. [Google Scholar] [CrossRef] [PubMed]
- Damont, A.; Legrand, A.; Cao, C.; Fenaille, F.; Tabet, J.-C. Hydrogen/deuterium exchange mass spectrometry in the world of small molecules. Mass Spectrom. Rev. 2023, 42, 1300–1331. [Google Scholar] [CrossRef]
- Kipping, M.; Schierhorn, A. Improving hydrogen/deuterium exchange mass spectrometry by reduction of the back-exchange effect. J. Mass Spectrom. 2003, 38, 271–276. [Google Scholar] [CrossRef]
- Liu, D.Q.; Hop, C.E.C.A.; Beconi, M.G.; Mao, A.; Chiu, S.H.L. Use of on-line hydrogen/deuterium exchange to facilitate metabolite identification. Rapid Commun. Mass Spectrom. 2001, 15, 1832–1839. [Google Scholar] [CrossRef]
- Liu, T.; Du, F.; Wan, Y.; Zhu, F.; Xing, J. Rapid identification of phase I and II metabolites of artemisinin antimalarials using LTQ-Orbitrap hybrid mass spectrometer in combination with online hydrogen/deuterium exchange technique. J. Mass Spectrom. 2011, 46, 725–733. [Google Scholar] [CrossRef]
- Kostyukevich, Y.; Acter, T.; Zherebker, A.; Ahmed, A.; Kim, S.; Nikolaev, E. Hydrogen/deuterium exchange in mass spectrometry. Mass Spectrom. Rev. 2018, 37, 811–853. [Google Scholar] [CrossRef]
- Osipenko, S.; Nikolaev, E.; Kostyukevich, Y. Amine additives for improved in-ESI H/D exchange. Analyst 2022, 147, 3180–3185. [Google Scholar] [CrossRef]
- Lam, W.; Ramanathan, R. In electrospray ionization source hydrogen/deuterium exchange LC-MS and LC-MS/MS for characterization of metabolites. J. Am. Soc. Mass Spectrom. 2002, 13, 345–353. [Google Scholar] [CrossRef]
- Shi, C.; Jia, H.; Chen, S.; Huang, J.; Peng, Y.e.; Guo, W. Hydrogen/deuterium exchange aiding metabolite identification in single-cell nanospray high-resolution mass spectrometry analysis. Anal. Chem. 2021, 94, 650–657. [Google Scholar] [CrossRef]
- Osipenko, S.; Zherebker, A.; Rumiantseva, L.; Kovaleva, O.; Nikolaev, E.N.; Kostyukevich, Y. Oxygen Isotope Exchange Reaction for Untargeted LC–MS Analysis. J. Am. Soc. Mass Spectrom. 2022, 33, 390–398. [Google Scholar] [CrossRef]
- Rumiantseva, L.; Osipenko, S.; Zharikov, A.; Kireev, A.; Nikolaev, E.N.; Kostyukevich, Y. Analysis of 16O/18O and H/D Exchange Reactions between Carbohydrates and Heavy Water Using High-Resolution Mass Spectrometry. Int. J. Mol. Sci. 2022, 23, 3585. [Google Scholar] [CrossRef] [PubMed]
- Rumiantseva, L.; Osipenko, S.; Podolskiy, I.I.; Burmykin, D.A.; Kovaleva, O.; Nikolaev, E.N.; Kostyukevich, Y. Increasing the reliability of compound identification in biological samples using 16O/18O-exchange mass spectrometry. Anal. Bioanal. Chem. 2022, 414, 2537–2543. [Google Scholar] [CrossRef] [PubMed]
- Tupertsev, B.; Osipenko, S.; Kireev, A.; Nikolaev, E.; Kostyukevich, Y. Simple In Vitro 18O Labeling for Improved Mass Spectrometry-Based Drug Metabolites Identification: Deep Drug Metabolism Study. Int. J. Mol. Sci. 2023, 24, 4569. [Google Scholar] [CrossRef]
- Ling-Fei, K.; Yu-Nan, C.; Pan, Y.; Tuo, Q.; Xin-Tang, W.; Rui-Qi, L.; Xiao-Juan, R.; Cai, T. 16O/18O-exchange internal standard preparation enhancing reliability of bio-sample natural bioactive compounds absolute quantitation. J. Chromatogr. B 2023, 1219, 123651. [Google Scholar] [CrossRef]
- Hegeman, A.D.; Schulte, C.F.; Cui, Q.; Lewis, I.A.; Huttlin, E.L.; Eghbalnia, H.; Harms, A.C.; Ulrich, E.L.; Markley, J.L.; Sussman, M.R. Stable Isotope Assisted Assignment of Elemental Compositions for Metabolomics. Anal. Chem. 2007, 79, 6912–6921. [Google Scholar] [CrossRef] [PubMed]
- Kera, K.; Fine, D.D.; Wherritt, D.J.; Nagashima, Y.; Shimada, N.; Ara, T.; Ogata, Y.; Sumner, L.W.; Suzuki, H. Pathway-specific metabolome analysis with 18O2-labeled Medicago truncatula via a mass spectrometry-based approach. Metabolomics 2018, 14, 71. [Google Scholar] [CrossRef]
- Ando, S.; Tanaka, Y.; Toyoda, Y.; Kon, K. Turnover of myelin lipids in aging brain. Neurochem. Res. 2003, 28, 5–13. [Google Scholar] [PubMed]
- Schoenheimer, R.; Rittenberg, D. The application of isotopes to the study of intermediary metabolism. Science 1938, 87, 221–226. [Google Scholar]
- Duarte, J.A.G.; Carvalho, F.; Pearson, M.; Horton, J.D.; Browning, J.D.; Jones, J.G.; Burgess, S.C. A high-fat diet suppresses de novo lipogenesis and desaturation but not elongation and triglyceride synthesis in mice. J. Lipid Res. 2014, 55, 2541–2553. [Google Scholar] [PubMed]
- Kostyukevich, Y.; Stekolshikova, E.; Levashova, A.; Kovalenko, A.; Vishnevskaya, A.; Bashilov, A.; Kireev, A.; Tupertsev, B.; Rumiantseva, L.; Khaitovich, P.; et al. Untargeted Lipidomics after D2O Administration Reveals the Turnover Rate of Individual Lipids in Various Organs of Living Organisms. Int. J. Mol. Sci. 2023, 24, 11725. [Google Scholar] [CrossRef] [PubMed]
- Rochfort, S.J.; Trenerry, V.C.; Imsic, M.; Panozzo, J.; Jones, R. Class targeted metabolomics: ESI ion trap screening methods for glucosinolates based on MSn fragmentation. Phytochemistry 2008, 69, 1671–1679. [Google Scholar] [CrossRef] [PubMed]
- Mohn, T.; Cutting, B.; Ernst, B.; Hamburger, M. Extraction and analysis of intact glucosinolates—A validated pressurized liquid extraction/liquid chromatography–mass spectrometry protocol for Isatis tinctoria, and qualitative analysis of other cruciferous plants. J. Chromatogr. A 2007, 1166, 142–151. [Google Scholar]
- Gil, V.; MacLeod, A.J. Studies on glucosinolate degradation in Lepidium sativum seed extracts. Phytochemistry 1980, 19, 1369–1374. [Google Scholar]
- Jain, T.; Grover, K. A comprehensive review on the nutritional and nutraceutical aspects of garden cress (Lepidium sativum Linn.). Proc. Natl. Acad. Sci. India Sect. B Biol. Sci. 2018, 88, 829–836. [Google Scholar]
- Painuli, S.; Quispe, C.; Herrera-Bravo, J.; Semwal, P.; Martorell, M.; Almarhoon, Z.M.; Seilkhan, A.; Ydyrys, A.; Rad, J.S.; Alshehri, M.M. Nutraceutical profiling, bioactive composition, and biological applications of Lepidium sativum L. Oxidative Med. Cell. Longev. 2022, 2022, 2910411. [Google Scholar]
- Lijina, P.; Manjunatha, J.R.; Kumar, B.S.G. Characterization of free oligosaccharides from garden cress seed aqueous exudate using PGC LC-MS/MS and NMR spectroscopy. Carbohydr. Res. 2023, 532, 108914. [Google Scholar] [PubMed]
- Alqahtani, F.Y.; Aleanizy, F.S.; Mahmoud, A.Z.; Farshori, N.N.; Alfaraj, R.; Al-Sheddi, E.S.; Alsarra, I.A. Chemical composition and antimicrobial, antioxidant, and anti-inflammatory activities of Lepidium sativum seed oil. Saudi J. Biol. Sci. 2019, 26, 1089–1092. [Google Scholar]
- Kaiyrkulova, A.; Li, J.; Aisa, H. Chemical constituents of Lepidium sativum seeds. Chem. Nat. Compd. 2019, 55, 736–737. [Google Scholar]
- Singh, B.; Jain, D.; Joshi, A.; Namrata; Dodiya, N.S.; Chauhan, S.; Mittal, J.; Singh, A. Molecular diversity analysis and metabolic profiling of seed oil in Lepidium sativum L. Genotypes. Plant Mol. Biol. Rep. 2020, 38, 641–654. [Google Scholar]
- Macleod, A.J.; Islam, R. Volatile flavour components of garden cress. J. Sci. Food Agric. 1976, 27, 909–912. [Google Scholar]
- Oszmiański, J.; Kolniak-Ostek, J.; Wojdyło, A. Application of ultra performance liquid chromatography-photodiode detector-quadrupole/time of flight-mass spectrometry (UPLC-PDA-Q/TOF-MS) method for the characterization of phenolic compounds of Lepidium sativum L. sprouts. Eur. Food Res. Technol. 2013, 236, 699–706. [Google Scholar] [CrossRef]
- Radwan, H.; El-Missiry, M.; Al-Said, W.; Ismail, A.; Abdel Shafeek, K.; Seif-El-Nasr, M. Investigation of the glucosinolates of Lepidium sativum growing in Egypt and their biological activity. Res. J. Med. Med. Sci. 2007, 2, 127–132. [Google Scholar]
- Sakran, M.; Selim, Y.; Zidan, N. A new isoflavonoid from seeds of Lepidium sativum L. and its protective effect on hepatotoxicity induced by paracetamol in male rats. Molecules 2014, 19, 15440–15451. [Google Scholar]
- Akash, S.S.; Singh, S.K. Phytoconstituents estimation of Lepidium sativum L. seed extract using GC-MS spectroscopy. World J. Pharm. Res. 2017, 7, 1360–1367. [Google Scholar]
- Al-Snafi, A. Chemical constituents and pharmacological effects of Lepidium sativum. Int. J. Curr. Pharm. Res. 2019, 11, 1–10. [Google Scholar]
- El-Haggar, M.; El-Hosseiny, L.; Ghazy, N.M.; El-Fiky, F.K.; El-Hawiet, A. Phytochemical investigation, antimicrobial and cytotoxic activities of suspension cultures of Lepidium sativum L. S. Afr. J. Bot. 2021, 138, 500–505. [Google Scholar]
- Nazir, S.; El-Sherif, A.A.; Abdel-Ghani, N.T.; Ibrahim, M.A.; Hegazy, M.-E.F.; Atia, M.A. Lepidium sativum secondary metabolites (Essential Oils): In vitro and in silico studies on human hepatocellular carcinoma cell lines. Plants 2021, 10, 1863. [Google Scholar]
- Reichl, B.; Himmelsbach, M.; Emhofer, L.; Klampfl, C.W.; Buchberger, W. Uptake and metabolism of the antidepressants sertraline, clomipramine, and trazodone in a garden cress (Lepidium sativum) model. Electrophoresis 2018, 39, 1301–1308. [Google Scholar] [CrossRef]
- Emhofer, L.; Himmelsbach, M.; Buchberger, W.; Klampfl, C.W. High-performance liquid chromatography drift-tube ion-mobility quadrupole time-of-flight/mass spectrometry for the identity confirmation and characterization of metabolites from three statins (lipid-lowering drugs) in the model plant cress (Lepidium sativum) after uptake from water. J. Chromatogr. A 2019, 1592, 122–132. [Google Scholar]
- Sarvin, B.; Himmelsbach, M.; Baygildiev, T.; Shpigun, O.; Rodin, I.; Stavrianidi, A.; Buchberger, W. Nerve agent markers screening after accumulation in garden cress (Lepidium sativum) used as a model plant object. J. Chromatogr. A 2019, 1597, 214–219. [Google Scholar] [CrossRef] [PubMed]
- Mlynek, F.; Himmelsbach, M.; Buchberger, W.; Klampfl, C.W. Time study on the uptake of four different beta-blockers in garden cress (Lepidium sativum) as a model plant. Environ. Sci. Pollut. Res. 2021, 28, 59382–59390. [Google Scholar]
- Maeda, H.; Dudareva, N. The Shikimate Pathway and Aromatic Amino Acid Biosynthesis in Plants. Annu. Rev. Plant Biol. 2012, 63, 73–105. [Google Scholar] [CrossRef]
- Bentley, R.; Haslam, E. The shikimate pathway—A metabolic tree with many branche. Crit. Rev. Biochem. Mol. Biol. 1990, 25, 307–384. [Google Scholar]
- Herrmann, K.M.; Weaver, L.M. The shikimate pathway. Annu. Rev. Plant Biol. 1999, 50, 473–503. [Google Scholar]
- Maier, U.H.; Gundlach, H.; Zenk, M.H. Seven imidazole alkaloids from Lepidium sativum. Phytochemistry 1998, 49, 1791–1795. [Google Scholar] [CrossRef]
- Tsugawa, H.; Nakabayashi, R.; Mori, T.; Yamada, Y.; Takahashi, M.; Rai, A.; Sugiyama, R.; Yamamoto, H.; Nakaya, T.; Yamazaki, M.; et al. A cheminformatics approach to characterize metabolomes in stable-isotope-labeled organisms. Nat. Methods 2019, 16, 295–298. [Google Scholar] [CrossRef] [PubMed]
- Lai, Z.; Tsugawa, H.; Wohlgemuth, G.; Mehta, S.; Mueller, M.; Zheng, Y.; Ogiwara, A.; Meissen, J.; Showalter, M.; Takeuchi, K.; et al. Identifying metabolites by integrating metabolome databases with mass spectrometry cheminformatics. Nat. Methods 2018, 15, 53–56. [Google Scholar] [CrossRef] [PubMed]
- Horai, H.; Arita, M.; Kanaya, S.; Nihei, Y.; Ikeda, T.; Suwa, K.; Ojima, Y.; Tanaka, K.; Tanaka, S.; Aoshima, K. MassBank: A public repository for sharing mass spectral data for life sciences. J. Mass Spectrom. 2010, 45, 703–714. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound | Formula | Number of Detected D | Number of Detected 18O |
|---|---|---|---|
| Arginine | C6H14N4O2 | 0 | 2 |
| Asparagine | C4H8N2O3 | 3 | 3 |
| Aspartate | C4H7NO4 | 3 | 4 |
| Glutamine | C5H10N2O3 | 5 | 3 |
| Histidine | C6H9N3O2 | 3 | 2 |
| Isoleucine | C6H13NO2 | 6 | 2 |
| Leucine | C6H13NO2 | 6 | 2 |
| Lysine | C6H14N2O2 | 1 | 2 |
| Phenylalanine | C9H11NO2 | 5 | 2 |
| Threonine | C4H9NO3 | 2 | 3 |
| Tryptophan | C11H12N2O2 | 1 | 2 |
| Tyrosine | C9H11NO3 | 4 | 3 |
| Valine | C5H11NO2 | 5 | 2 |
| Proline | C5H9NO2 | 1 | 2 |
| Methionine | C5H11NO2S | 3 | 2 |
| Pyroglutamic acid | C5H7NO3 | 5 | 2 |
| Malic acid | C4H6O5 | 3 | 2 |
| Fumaric acid | C4H4O4 | 2 | 2 |
| Succinic acid | C4H6O4 | 4 | 2 |
| Benzoic acid | C7H6O2 | 0 | 2 |
| Glucose-6-phosphate | C6H13O9P | 5 | 2 |
| Betaine | C5H11NO2 | 6 | 2 |
| Phosphocholine | C5H15NO4P+ | 0 | 2 |
| p-Coumaric acid | C9H8O3 | 2 | 2 |
| Acetyl-L-Carnitine | C9H17NO4 | ND | 2 |
| Glucotropaeolin | C14H19NO9S2 | 0 | 2 |
| 4-methoxyglucobrassicin | C17H22N2O10S2 | 9 | 2 |
| Sinapoyl malate | C15H16O9 | 3 | 2 |
| Sinapine | C16H24NO5 | 0 | 2 |
| Kaempferol rhamnose-hexoside | C33H40O20 | 0 | 2 |
| Semilepidinoside A | C16H20N2O6 | 6 | 2 |
| Semilepidinoside B | C17H22N2O7 | 8 | 2 |
| Lepidine A/C | C21H20N4O2 | 0 | 0 |
| Lepidine B/D/E/F | C20H18N4O2 | 0 | 0 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Osipenko, S.; Bashilov, A.; Vishnevskaya, A.; Rumiantseva, L.; Levashova, A.; Kovalenko, A.; Tupertsev, B.; Kireev, A.; Nikolaev, E.; Kostyukevich, Y. Investigating the Metabolism of Plants Germinated in Heavy Water, D2O, and H218O-Enriched Media Using High-Resolution Mass Spectrometry. Int. J. Mol. Sci. 2023, 24, 15396. https://doi.org/10.3390/ijms242015396
Osipenko S, Bashilov A, Vishnevskaya A, Rumiantseva L, Levashova A, Kovalenko A, Tupertsev B, Kireev A, Nikolaev E, Kostyukevich Y. Investigating the Metabolism of Plants Germinated in Heavy Water, D2O, and H218O-Enriched Media Using High-Resolution Mass Spectrometry. International Journal of Molecular Sciences. 2023; 24(20):15396. https://doi.org/10.3390/ijms242015396
Chicago/Turabian StyleOsipenko, Sergey, Anton Bashilov, Anna Vishnevskaya, Lidiia Rumiantseva, Anna Levashova, Anna Kovalenko, Boris Tupertsev, Albert Kireev, Eugene Nikolaev, and Yury Kostyukevich. 2023. "Investigating the Metabolism of Plants Germinated in Heavy Water, D2O, and H218O-Enriched Media Using High-Resolution Mass Spectrometry" International Journal of Molecular Sciences 24, no. 20: 15396. https://doi.org/10.3390/ijms242015396