
Vaping-Induced Proteolysis Causes Airway Surface Dehydration

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Smokers’ and Vapers’ BALF Alter ENaC Subunit Composition
2.2. Vapers and Smokers BALF Increase ENaC Activity in Cultured Airway Epithelia
2.3. Proteases in Vapers’ and Smokers’ BALF Cause ASL Dehydration
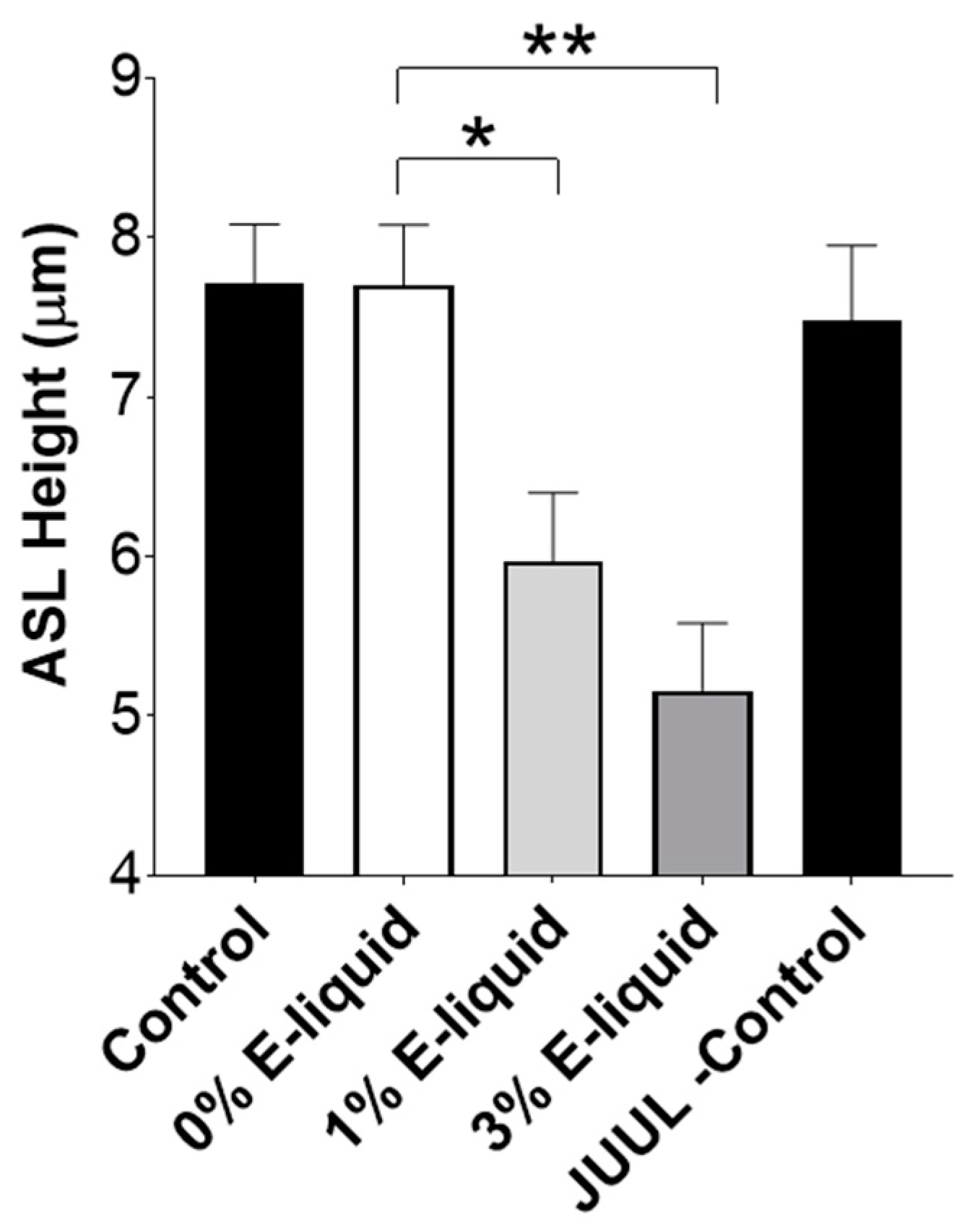
2.4. E-Liquids Increase Protease Release and Can Indirectly Cause ASL Dehydration
3. Discussion
4. Materials and Methods
4.1. Subject Recruitment and Sample Collection
4.2. Cell Culture
4.3. ASL Height Measurements
4.4. Electrophysiological Measurements
4.5. Transfection of HEK293T Cells with ENaC Subunits and Western Blotting
4.6. Isolation of Neutrophils from Blood and Exposure to Menthol-Flavored E-Liquid
4.7. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Talih, S.; Salman, R.; El-Hage, R.; Karam, E.; Salam, S.; Karaoghlanian, N.; El-Hellani, A.; Saliba, N.; Shihadeh, A. A comparison of the electrical characteristics, liquid composition, and toxicant emissions of JUUL USA and JUUL UK e-cigarettes. Sci. Rep. 2020, 10, 7322. [Google Scholar] [CrossRef] [PubMed]
- Pulvers, K.; Rice, M.; Ahluwalia, J.S.; Arnold, M.J.; Marez, C.; Nollen, N.L. “It is the One Thing that has Worked”: Facilitators and barriers to switching to nicotine salt pod system e-cigarettes among African American and Latinx people who smoke: A content analysis. Harm Reduct. J. 2021, 18, 98. [Google Scholar] [CrossRef]
- Leone, F.T.; Carlsen, K.H.; Chooljian, D.; Crotty Alexander, L.E.; Detterbeck, F.C.; Eakin, M.N.; Evers-Casey, S.; Farber, H.J.; Folan, P.; Kathuria, H.; et al. Recommendations for the Appropriate Structure, Communication, and Investigation of Tobacco Harm Reduction Claims. An Official American Thoracic Society Policy Statement. Am. J. Respir. Crit. Care Med. 2018, 198, e90–e105. [Google Scholar] [CrossRef] [PubMed]
- Bandi, P.; Cahn, Z.; Goding Sauer, A.; Douglas, C.E.; Drope, J.; Jemal, A.; Fedewa, S.A. Trends in E-Cigarette Use by Age Group and Combustible Cigarette Smoking Histories, U.S. Adults, 2014–2018. Am. J. Prev. Med. 2021, 60, 151–158. [Google Scholar] [CrossRef] [PubMed]
- Park-Lee, E.; Ren, C.; Sawdey, M.D.; Gentzke, A.S.; Cornelius, M.; Jamal, A.; Cullen, K.A. Notes from the Field: E-Cigarette Use Among Middle and High School Students—National Youth Tobacco Survey, United States, 2021. MMWR Morb. Mortal. Wkly. Rep. 2021, 70, 1387–1389. [Google Scholar] [CrossRef]
- Bhatta, D.N.; Glantz, S.A. Association of E-Cigarette Use with Respiratory Disease Among Adults: A Longitudinal Analysis. Am. J. Prev. Med. 2020, 58, 182–190. [Google Scholar] [CrossRef]
- Berlowitz, J.B.; Xie, W.; Harlow, A.F.; Blaha, M.J.; Bhatnagar, A.; Benjamin, E.J.; Stokes, A.C. Cigarette–E-cigarette Transitions and Respiratory Symptom Development. Am. J. Prev. Med. 2023, 64, 556–560. [Google Scholar] [CrossRef]
- Tackett, A.P.; Urman, R.; Barrington-Trimis, J.; Liu, F.; Hong, H.; Pentz, M.A.; Islam, T.S.; Eckel, S.P.; Rebuli, M.; Leventhal, A.; et al. Prospective study of e-cigarette use and respiratory symptoms in adolescents and young adults. Thorax 2023. [Google Scholar] [CrossRef]
- Bowler, R.P.; Hansel, N.N.; Jacobson, S.; Graham Barr, R.; Make, B.J.; Han, M.K.; O’Neal, W.K.; Oelsner, E.C.; Casaburi, R.; Barjaktarevic, I.; et al. Electronic Cigarette Use in US Adults at Risk for or with COPD: Analysis from Two Observational Cohorts. J. Gen. Intern. Med. 2017, 32, 1315–1322. [Google Scholar] [CrossRef]
- Mei, F.; Dalmartello, M.; Bonifazi, M.; Bertuccio, P.; Levi, F.; Boffetta, P.; Negri, E.; La Vecchia, C.; Malvezzi, M. Chronic obstructive pulmonary disease (COPD) mortality trends worldwide: An update to 2019. Respirology 2022, 27, 941–950. [Google Scholar] [CrossRef]
- Ghosh, A.; Boucher, R.C.; Tarran, R. Airway hydration and COPD. Cell. Mol. Life Sci. 2015, 72, 3637–3652. [Google Scholar] [CrossRef] [PubMed]
- Anderson, W.H.; Coakley, R.D.; Button, B.; Henderson, A.G.; Zeman, K.L.; Alexis, N.E.; Peden, D.B.; Lazarowski, E.R.; Davis, C.W.; Bailey, S.; et al. The Relationship of Mucus Concentration (Hydration) to Mucus Osmotic Pressure and Transport in Chronic Bronchitis. Am. J. Respir. Crit. Care Med. 2015, 192, 182–190. [Google Scholar] [CrossRef] [PubMed]
- Hogg, J.C.; Chu, F.; Utokaparch, S.; Woods, R.; Elliott, W.M.; Buzatu, L.; Cherniack, R.M.; Rogers, R.M.; Sciurba, F.C.; Coxson, H.O.; et al. The nature of small-airway obstruction in chronic obstructive pulmonary disease. N. Engl. J. Med. 2004, 350, 2645–2653. [Google Scholar] [CrossRef] [PubMed]
- Collawn, J.F.; Matalon, S. CFTR and lung homeostasis. Am. J. Physiol. Lung Cell. Mol. Physiol. 2014, 307, L917–L923. [Google Scholar] [CrossRef]
- Zhao, R.; Liang, X.; Zhao, M.; Liu, S.L.; Huang, Y.; Idell, S.; Li, X.; Ji, H.L. Correlation of apical fluid-regulating channel proteins with lung function in human COPD lungs. PLoS ONE 2014, 9, e109725. [Google Scholar] [CrossRef]
- Mall, M.; Grubb, B.R.; Harkema, J.R.; O’Neal, W.K.; Boucher, R.C. Increased airway epithelial Na+ absorption produces cystic fibrosis-like lung disease in mice. Nat. Med. 2004, 10, 487–493. [Google Scholar] [CrossRef]
- Mall, M.A.; Harkema, J.R.; Trojanek, J.B.; Treis, D.; Livraghi, A.; Schubert, S.; Zhou, Z.; Kreda, S.M.; Tilley, S.L.; Hudson, E.J.; et al. Development of chronic bronchitis and emphysema in beta-epithelial Na+ channel-overexpressing mice. Am. J. Respir. Crit. Care Med. 2008, 177, 730–742. [Google Scholar] [CrossRef]
- Churg, A.; Wright, J.L. Proteases and emphysema. Curr. Opin. Pulm. Med. 2005, 11, 153–159. [Google Scholar] [CrossRef]
- Rossier, B.C.; Stutts, M.J. Activation of the epithelial sodium channel (ENaC) by serine proteases. Annu. Rev. Physiol. 2009, 71, 361–379. [Google Scholar] [CrossRef]
- Thibodeau, P.H.; Butterworth, M.B. Proteases, cystic fibrosis and the epithelial sodium channel (ENaC). Cell Tissue Res. 2013, 351, 309–323. [Google Scholar] [CrossRef]
- Ghosh, A.; Coakley, R.D.; Ghio, A.J.; Muhlebach, M.S.; Esther, C.R.; Alexis, N.E., Jr.; Tarran, R. Chronic E-Cigarette Use Increases Neutrophil Elastase and Matrix Metalloprotease Levels in the Lung. Am. J. Respir. Crit. Care Med. 2019, 200, 1392–1401. [Google Scholar] [CrossRef] [PubMed]
- Reidel, B.; Radicioni, G.; Clapp, P.; Ford, A.A.; Abdelwahab, S.; Rebuli, M.E.; Haridass, P.; Alexis, N.E.; Jaspers, I.; Kesimer, M. E-Cigarette Use Causes a Unique Innate Immune Response in the Lung Involving Increased Neutrophilic Activation and Altered Mucin Secretion. Am. J. Respir. Crit. Care Med. 2017, 197, 492–501. [Google Scholar] [CrossRef] [PubMed]
- Tarran, R.; Sabater, J.R.; Clarke, T.C.; Tan, C.D.; Davies, C.M.; Liu, J.; Yeung, A.; Garland, A.L.; Stutts, M.J.; Abraham, W.M.; et al. Nonantibiotic macrolides prevent human neutrophil elastase-induced mucus stasis and airway surface liquid volume depletion. Am. J. Physiol. Lung Cell. Mol. Physiol. 2013, 304, L746–L756. [Google Scholar] [CrossRef] [PubMed]
- Althaus, M.; Lawong, R.Y. Proteolytic ENaC activation in health and disease—A complicated puzzle. Pflug. Arch. 2022, 474, 177–179. [Google Scholar] [CrossRef]
- Ghosh, A.; Girish, V.; Yuan, M.L.; Coakley, R.D.; Wrennall, J.A.; Alexis, N.E.; Sausville, E.L.; Vasudevan, A.; Chait, A.R.; Sheltzer, J.M.; et al. Combustible and Electronic Cigarette Exposures Increase ACE2 Activity and SARS-CoV-2 Spike Binding. Am. J. Respir. Crit. Care Med. 2021, 205, 129–133. [Google Scholar] [CrossRef] [PubMed]
- Tarran, R.; Button, B.; Picher, M.; Paradiso, A.M.; Ribeiro, C.M.; Lazarowski, E.R.; Zhang, L.; Collins, P.L.; Pickles, R.J.; Fredberg, J.J.; et al. Normal and cystic fibrosis airway surface liquid homeostasis. The effects of phasic shear stress and viral infections. J. Biol. Chem. 2005, 280, 35751–35759. [Google Scholar] [CrossRef]
- Anand, D.; Hummler, E.; Rickman, O.J. ENaC activation by proteases. Acta Physiol. 2022, 235, e13811. [Google Scholar] [CrossRef]
- Zhou-Suckow, Z.; Duerr, J.; Hagner, M.; Agrawal, R.; Mall, M.A. Airway mucus, inflammation and remodeling: Emerging links in the pathogenesis of chronic lung diseases. Cell Tissue Res. 2017, 367, 537–550. [Google Scholar] [CrossRef]
- Caldwell, R.A.; Boucher, R.C.; Stutts, M.J. Neutrophil elastase activates near-silent epithelial Na+ channels and increases airway epithelial Na+ transport. Am. J. Physiol. Lung Cell. Mol. Physiol. 2005, 288, L813–L819. [Google Scholar] [CrossRef]
- Kleyman, T.R.; Carattino, M.D.; Hughey, R.P. ENaC at the cutting edge: Regulation of epithelial sodium channels by proteases. J. Biol. Chem. 2009, 284, 20447–20451. [Google Scholar] [CrossRef]
- Hanrahan, J.W.; Abu-Arish, A.; Wong, F.H.; Turner, M.J.; Carlile, G.W.; Thomas, D.Y.; Cantin, A.M. Chronic obstructive pulmonary disease and the modulation of CFTR by acute exposure to cigarette smoke. Am. J. Physiol. Cell Physiol. 2022, 323, C1374–C1392. [Google Scholar] [CrossRef] [PubMed]
- Clunes, L.A.; Davies, C.M.; Coakley, R.D.; Aleksandrov, A.A.; Henderson, A.G.; Zeman, K.L.; Worthington, E.N.; Gentzsch, M.; Kreda, S.M.; Cholon, D.; et al. Cigarette smoke exposure induces CFTR internalization and insolubility, leading to airway surface liquid dehydration. FASEB J. 2012, 26, 533–545. [Google Scholar] [CrossRef]
- Rasmussen, L.W.; Stanford, D.; LaFontaine, J.; Allen, A.D.; Raju, S.V. Nicotine aerosols diminish airway CFTR function and mucociliary clearance. Am. J. Physiol. Lung Cell. Mol. Physiol. 2023, 324, L557–L570. [Google Scholar] [CrossRef]
- Lin, V.Y.; Fain, M.D.; Jackson, P.L.; Berryhill, T.F.; Wilson, L.S.; Mazur, M.; Barnes, S.J.; Blalock, J.E.; Raju, S.V.; Rowe, S.M. Vaporized E-Cigarette Liquids Induce Ion Transport Dysfunction in Airway Epithelia. Am. J. Respir. Cell Mol. Biol. 2019, 61, 162–173. [Google Scholar] [CrossRef]
- Kim, M.D.; Chung, S.; Baumlin, N.; Sun, L.; Silswal, N.; Dennis, J.S.; Yoshida, M.; Sabater, J.; Horrigan, F.T.; Salathe, M. E-cigarette aerosols of propylene glycol impair BK channel activity and parameters of mucociliary function. Am. J. Physiol. Lung Cell. Mol. Physiol. 2023, 324, L468–L479. [Google Scholar] [CrossRef] [PubMed]
- Tarran, R.; Grubb, B.R.; Parsons, D.; Picher, M.; Hirsh, A.J.; Davis, C.W.; Boucher, R.C. The CF salt controversy: In vivo observations and therapeutic approaches. Mol. Cell 2001, 8, 149–158. [Google Scholar] [CrossRef] [PubMed]
- Donaldson, S.H.; Bennett, W.D.; Zeman, K.L.; Knowles, M.R.; Tarran, R.; Boucher, R.C. Mucus clearance and lung function in cystic fibrosis with hypertonic saline. N. Engl. J. Med. 2006, 354, 241–250. [Google Scholar] [CrossRef]
- Boucher, R.C. Human airway ion transport. Part one. Am. J. Respir. Crit. Care Med. 1994, 150, 271–281. [Google Scholar] [CrossRef]
- Tarran, R.; Trout, L.; Donaldson, S.H.; Boucher, R.C. Soluble mediators, not cilia, determine airway surface liquid volume in normal and cystic fibrosis superficial airway epithelia. J. Gen. Physiol. 2006, 127, 591–604. [Google Scholar] [CrossRef]
- Myerburg, M.M.; Butterworth, M.B.; McKenna, E.E.; Peters, K.W.; Frizzell, R.A.; Kleyman, T.R.; Pilewski, J.M. Airway surface liquid volume regulates ENaC by altering the serine protease-protease inhibitor balance: A mechanism for sodium hypersabsorption in cystic fibrosis. J. Biol. Chem. 2006, 281, 27942–27949. [Google Scholar] [CrossRef]
- Rasmussen, J.E.; Sheridan, J.T.; Polk, W.; Davies, C.M.; Tarran, R. Cigarette smoke-induced Ca2+ release leads to cystic fibrosis transmembrane conductance regulator (CFTR) dysfunction. J. Biol. Chem. 2014, 289, 7671–7681. [Google Scholar] [CrossRef] [PubMed]
- Webster, M.J.; Reidel, B.; Tan, C.D.; Ghosh, A.; Alexis, N.E.; Donaldson, S.H.; Kesimer, M.; Ribeiro, C.M.P.; Tarran, R. SPLUNC1 degradation by the cystic fibrosis mucosal environment drives airway surface liquid dehydration. Eur. Respir. J. 2018, 52, 1800668. [Google Scholar] [CrossRef] [PubMed]
- Maouche, K.; Medjber, K.; Zahm, J.M.; Delavoie, F.; Terryn, C.; Coraux, C.; Pons, S.; Cloez-Tayarani, I.; Maskos, U.; Birembaut, P.; et al. Contribution of alpha7 nicotinic receptor to airway epithelium dysfunction under nicotine exposure. Proc. Natl. Acad. Sci. USA 2013, 110, 4099–4104. [Google Scholar] [CrossRef]
- Mincham, K.T.; Bruno, N.; Singanayagam, A.; Snelgrove, R.J. Our evolving view of neutrophils in defining the pathology of chronic lung disease. Immunology 2021, 164, 701–721. [Google Scholar] [CrossRef] [PubMed]
- Alexander, L.E.C.; Bellinghausen, A.L.; Eakin, M.N. What are the mechanisms underlying vaping-induced lung injury? J. Clin. Investig. 2020, 130, 2754–2756. [Google Scholar] [CrossRef]
- Hunninghake, G.W.; Crystal, R.G. Cigarette smoking and lung destruction. Accumulation of neutrophils in the lungs of cigarette smokers. Am. Rev. Respir. Dis. 1983, 128, 833–838. [Google Scholar]
- Fulcher, M.L.; Gabriel, S.; Burns, K.A.; Yankaskas, J.R.; Randell, S.H. Well-differentiated human airway epithelial cell cultures. Methods Mol. Med. 2005, 107, 183–206. [Google Scholar]
- Ghosh, A.; Abdelwahab, S.H.; Reeber, S.L.; Reidel, B.; Marklew, A.J.; Garrison, A.J.; Lee, S.; Dang, H.; Herring, A.H.; Glish, G.L.; et al. Little Cigars are More Toxic than Cigarettes and Uniquely Change the Airway Gene and Protein Expression. Sci. Rep. 2017, 7, 46239. [Google Scholar] [CrossRef]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ghosh, A.; Coakley, R.D.; Alexis, N.E.; Tarran, R. Vaping-Induced Proteolysis Causes Airway Surface Dehydration. Int. J. Mol. Sci. 2023, 24, 15348. https://doi.org/10.3390/ijms242015348
Ghosh A, Coakley RD, Alexis NE, Tarran R. Vaping-Induced Proteolysis Causes Airway Surface Dehydration. International Journal of Molecular Sciences. 2023; 24(20):15348. https://doi.org/10.3390/ijms242015348
Chicago/Turabian StyleGhosh, Arunava, Raymond D. Coakley, Neil E. Alexis, and Robert Tarran. 2023. "Vaping-Induced Proteolysis Causes Airway Surface Dehydration" International Journal of Molecular Sciences 24, no. 20: 15348. https://doi.org/10.3390/ijms242015348