

Genome-Wide Association Study and Genomic Prediction of Fusarium Wilt Resistance in Common Bean Core Collection

 , ,
, , 
Abstract
:1. Introduction
2. Results
2.1. Phenotypic Variation
2.2. Population Structure and Phylogenetic Analysis
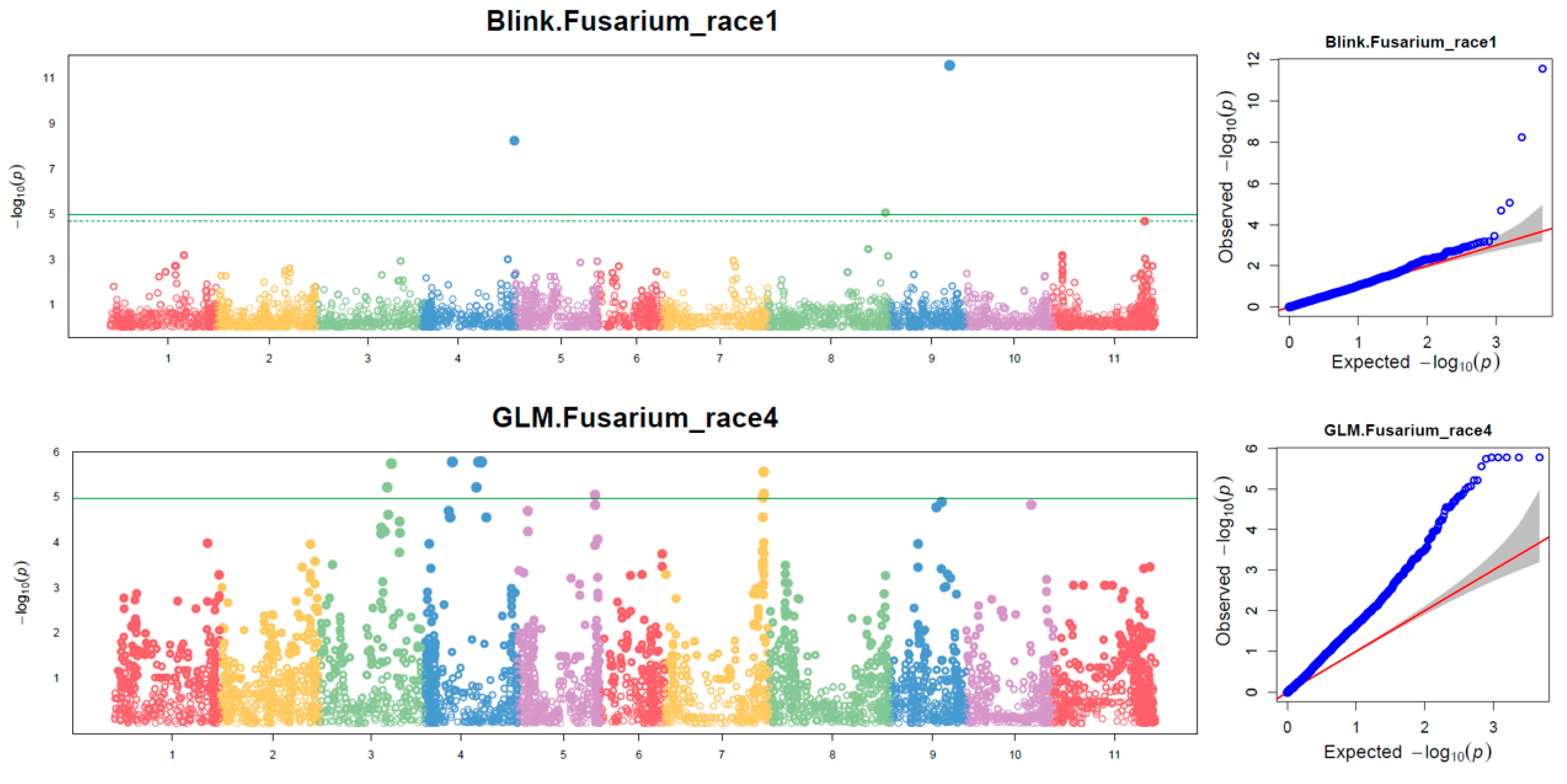
2.3. Association Analysis and SNP Marker Identification
2.3.1. The Associated SNPs for Race 1 Resistance
2.3.2. The Associated SNPs for Race 4 Resistance
2.4. Candidate Genes for Fusarium Wilt Resistance
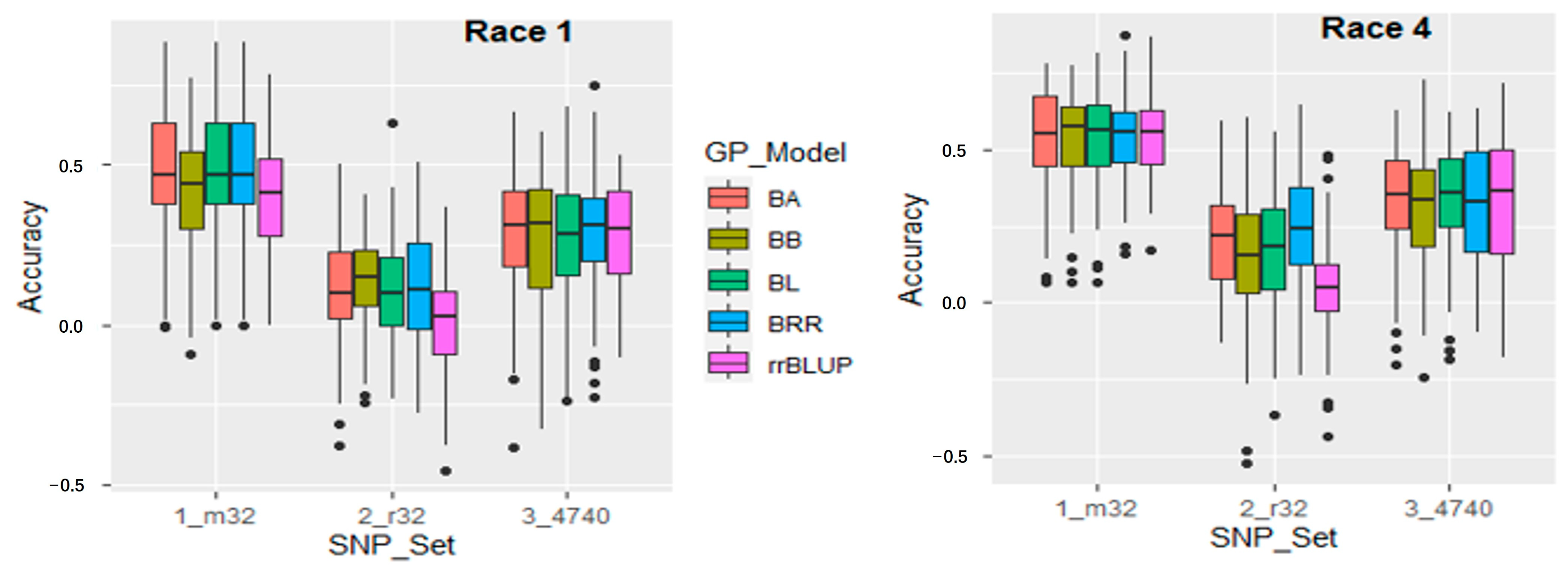
2.5. Genomic Prediction
3. Discussion
3.1. Genetic Diversity and Population Structure for the Common Bean Germplasm
3.2. Fusarium Wilt Phenotyping
3.3. Genome-Wide Association Study and SNP Identification
3.4. Candidate Gene for Fusarium Wilt Resistance
3.5. Genomic Prediction for Genomic Selection of Fusarium Wilt Resistance
4. Materials and Method
4.1. Plant Material and Phenotyping
4.2. Genotyping and SNP Selection
4.3. Principal Component Analysis (PCA) and Genetic Diversity
4.4. Genome-Wide Association Study and SNP Marker Identification
4.5. Candidate Gene Estimation
4.6. Genomic Prediction for Genomic Selection of Fusarium Wilt Resistance
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Rendón-Anaya, M.; Montero-Vargas, J.M.; Saburido-Álvarez, S.; Vlasova, A.; Capella-Gutierrez, S.; Ordaz-Ortiz, J.J.; Aguilar, O.M.; Vianello-Brondani, R.P.; Santalla, M.; Delaye, L.; et al. Genomic History of the Origin and Domestication of Common Bean Unveils Its Closest Sister Species. Genome Biol. 2017, 18, 60. [Google Scholar] [CrossRef]
- Schmutz, J.; McClean, P.E.; Mamidi, S.; Wu, G.A.; Cannon, S.B.; Grimwood, J.; Jenkins, J.; Shu, S.; Song, Q.; Chavarro, C.; et al. A Reference Genome for Common Bean and Genome-Wide Analysis of Dual Domestications. Nat. Genet. 2014, 46, 707–713. [Google Scholar] [CrossRef]
- Pathania, A.; Sharma, S.K.; Sharma, P.N. Common Bean. In Broadening the Genetic Base of Grain Legumes; Springer: New Delhi, India, 2014; pp. 11–50. [Google Scholar]
- Blair, M.W. Mineral Biofortification Strategies for Food Staples: The Example of Common Bean. J. Agric. Food Chem. 2013, 61, 8287–8294. [Google Scholar] [CrossRef]
- Reinprecht, Y.; Schram, L.; Marsolais, F.; Smith, T.H.; Hill, B.; Pauls, K.P. Effects of Nitrogen Application on Nitrogen Fixation in Common Bean Production. Front. Plant Sci. 2020, 11, 534817. [Google Scholar] [CrossRef]
- Gurel, F.B.; Kabir, N. Comparative Performance of Fungicides and Biocontrol Products in Suppression of Rhizoctonia Root Rot in Viburnum Integration of Sanitation Practice and Fungicide Application for Assuring Better Post-Harvest Shelflife of Cut Flowers and Greenery View Project. J. Plant Pathol. Microbiol. 2018, 9, 451. [Google Scholar] [CrossRef]
- Mihajlović, M.; Rekanović, E.; Hrustić, J.; Grahovac, M.; Tanović, B. In Vitro and in Vivo Toxicity of Fungicides and Biofungicides for the Control of Verticillium and Fusarium Wilt of Pepper. Pestic. Fitomedicina 2021, 36, 23–34. [Google Scholar] [CrossRef]
- Panth, M.; Hassler, S.C.; Baysal-Gurel, F. Methods for Management of Soilborne Diseases in Crop Production. Agriculture 2020, 10, 16. [Google Scholar] [CrossRef]
- Galdino, A.C.S.; de Freitas, M.B.; de Borba, M.C.; Stadnik, M.J. Phenolic and Flavonoid Content in Roots and Hypocotyls of Resistant and Susceptible Bean Plants (Phaseolus vulgaris L.) during Early Stage of Colonization by Fusarium oxysporum f. sp. phaseoli. Trop. Plant Pathol. 2021, 46, 553–558. [Google Scholar] [CrossRef]
- de Toledo-Souza, E.D.; da Silveira, P.M.; Café-Filho, A.C.; Lobo Junior, M. Fusarium Wilt Incidence and Common Bean Yield According to the Preceding Crop and the Soil Tillage System. Pesqui. Agropecuária Bras. 2012, 47, 1031–1037. [Google Scholar] [CrossRef]
- Fall, A.L.; Byrne, P.F.; Jung, G.; Coyne, D.P.; Brick, M.A.; Schwartz, H.F. Detection and Mapping of a Major Locus for Fusarium Wilt Resistance in Common Bean. Crop Sci. 2001, 41, 1494–1498. [Google Scholar] [CrossRef]
- Dita, M.; Barquero, M.; Heck, D.; Mizubuti, E.S.G.; Staver, C.P. Fusarium Wilt of Banana: Current Knowledge on Epidemiology and Research Needs toward Sustainable Disease Management. Front. Plant Sci. 2018, 871, 398832. [Google Scholar] [CrossRef]
- Jha, U.C.; Bohra, A.; Pandey, S.; Parida, S.K. Breeding, Genetics, and Genomics Approaches for Improving Fusarium Wilt Resistance in Major Grain Legumes. Front. Genet. 2020, 11, 560972. [Google Scholar] [CrossRef]
- Alves-Santos, F.M.; Cordeiro-Rodrigues, L.; Sayagués, J.M.; Martín-Domínguez, R.; García-Benavides, P.; Crespo, M.C.; Díaz-Mínguez, J.M.; Eslava, A.P. Pathogenicity and Race Characterization of Fusarium oxysporum f. sp. phaseoli Isolates from Spain and Greece. Plant Pathol. 2002, 51, 605–611. [Google Scholar] [CrossRef]
- Batista, R.O.; Silva, L.C.; Moura, L.M.; Souza, M.H.; Carneiro, P.C.S.; Filho, J.L.S.C.; de Souza Carneiro, J.E. Inheritance of Resistance to Fusarium Wilt in Common Bean. Euphytica 2017, 213, 133. [Google Scholar] [CrossRef]
- Nay, M.M.; Souza, T.L.P.O.; Raatz, B.; Mukankusi, C.M.; Pastor-Corrales, M.A.; Abreu, A.F.B.; Melo, L.C. A Review of Angular Leaf Spot Resistance in Common Bean. Crop Sci. 2019, 59, 1376–1391. [Google Scholar] [CrossRef]
- Brick, M.A.; Byrne, P.F.; Schwartz, H.F.; Ogg, J.B.; Otto, K.; Fall, A.L.; Gilbert, J. Reaction to Three Races of Fusarium Wilt in the Phaseolus vulgaris Core Collection. Crop Sci. 2006, 46, 1245–1252. [Google Scholar] [CrossRef]
- Paulino, J.F.D.C.; de Almeida, C.P.; Bueno, C.J.; Song, Q.; Fritsche-Neto, R.; Carbonell, S.A.M.; Chiorato, A.F.; Benchimol-Reis, L.L. Genome-Wide Association Study Reveals Genomic Regions Associated with Fusarium Wilt Resistance in Common Bean. Genes 2021, 12, 765. [Google Scholar] [CrossRef]
- Arie, T. Fusarium Diseases of Cultivated Plants, Control, Diagnosis, and Molecular and Genetic Studies. J. Pestic. Sci. 2019, 44, 275–281. [Google Scholar] [CrossRef]
- Dryad|Data—Is the USDA Core Collection of Common Bean Representative of Genetic Diversity of the Species, as Assessed by SNP Diversity? Available online: https://datadryad.org/stash/dataset/doi:10.25338/B8KP45 (accessed on 27 September 2023).
- Song, Q.; Hyten, D.L.; Jia, G.; Quigley, C.V.; Fickus, E.W.; Nelson, R.L.; Cregan, P.B. Development and Evaluation of SoySNP50K, a High-Density Genotyping Array for Soybean. PLoS ONE 2013, 8, e54985. [Google Scholar] [CrossRef]
- Lombardi, M.; Materne, M.; Cogan, N.O.I.; Rodda, M.; Daetwyler, H.D.; Slater, A.T.; Forster, J.W.; Kaur, S. Assessment of Genetic Variation within a Global Collection of Lentil (Lens culinaris Medik.) Cultivars and Landraces Using SNP Markers. BMC Genet. 2014, 15, 150. [Google Scholar] [CrossRef]
- Leitao, S.T.; Malosetti, M.; Song, Q.; Van Eeuwijk, F.; Rubiales, D.; Vaz Patto, M.C. Natural Variation in Portuguese Common Bean Germplasm Reveals New Sources of Resistance against Fusarium oxysporum f. sp. phaseoli and Resistance-Associated Candidate Genes. Phytopathology 2020, 110, 633–647. [Google Scholar] [CrossRef]
- Zia, B.; Shi, A.; Olaoye, D.; Xiong, H.; Ravelombola, W.; Gepts, P.; Schwartz, H.F.; Brick, M.A.; Otto, K.; Ogg, B.; et al. Genome-Wide Association Study and Genomic Prediction for Bacterial Wilt Resistance in Common Bean (Phaseolus vulgaris) Core Collection. Front. Genet. 2022, 13, 853114. [Google Scholar] [CrossRef] [PubMed]
- Shi, A.; Gepts, P.; Song, Q.; Xiong, H.; Michaels, T.E.; Chen, S. Genome-Wide Association Study and Genomic Prediction for Soybean Cyst Nematode Resistance in USDA Common Bean (Phaseolus vulgaris) Core Collection. Front. Plant Sci. 2021, 12, 624156. [Google Scholar] [CrossRef] [PubMed]
- Crossa, J.; Pérez-Rodríguez, P.; Cuevas, J.; Montesinos-López, O.; Jarquín, D.; de los Campos, G.; Burgueño, J.; González-Camacho, J.M.; Pérez-Elizalde, S.; Beyene, Y.; et al. Genomic Selection in Plant Breeding: Methods, Models, and Perspectives. Trends Plant Sci. 2017, 22, 961–975. [Google Scholar] [CrossRef]
- Zhang, Q.; Zhang, Q.; Jensen, J. Association Studies and Genomic Prediction for Genetic Improvements in Agriculture. Front. Plant Sci. 2022, 13, 904230. [Google Scholar] [CrossRef] [PubMed]
- Xavier, A.; Muir, W.M.; Rainey, K.M. Assessing Predictive Properties of Genome-Wide Selection in Soybeans. G3 Genes Genomes Genet. 2016, 6, 2611–2616. [Google Scholar] [CrossRef] [PubMed]
- Bao, Y.; Vuong, T.; Meinhardt, C.; Tiffin, P.; Denny, R.; Chen, S.; Nguyen, H.T.; Orf, J.H.; Young, N.D. Potential of Association Mapping and Genomic Selection to Explore PI 88788 Derived Soybean Cyst Nematode Resistance. Plant Genome 2014, 7, 3. [Google Scholar] [CrossRef]
- Diaz, L.M.; Arredondo, V.; Ariza-Suarez, D.; Aparicio, J.; Buendia, H.F.; Cajiao, C.; Mosquera, G.; Beebe, S.E.; Mukankusi, C.M.; Raatz, B. Genetic Analyses and Genomic Predictions of Root Rot Resistance in Common Bean Across Trials and Populations. Front. Plant Sci. 2021, 12, 629221. [Google Scholar] [CrossRef]
- Wen, L.; Chang, H.X.; Brown, P.J.; Domier, L.L.; Hartman, G.L. Genome-Wide Association and Genomic Prediction Identifies Soybean Cyst Nematode Resistance in Common Bean Including a Syntenic Region to Soybean Rhg1 Locus. Hortic. Res. 2019, 6, 9. [Google Scholar] [CrossRef]
- Meza, N.; Rosas, J.C.; Martín, J.P.; Ortiz, J.M. Biodiversity of Common Bean (Phaseolus vulgaris L.) in Honduras, Evidenced by Morphological Characterization. Genet. Resour. Crop Evol. 2013, 60, 1329–1336. [Google Scholar] [CrossRef]
- Blair, M.W.; Hurtado, N.; Chavarro, C.M.; Muñoz-Torres, M.C.; Giraldo, M.C.; Pedraza, F.; Tomkins, J.; Wing, R. Gene-Based SSR Markers for Common Bean (Phaseolus vulgaris L.) Derived from Root and Leaf Tissue ESTs: An Integration of the BMc Series. BMC Plant Biol. 2011, 11, 50. [Google Scholar] [CrossRef] [PubMed]
- Cortés, A.J.; Chavarro, M.C.; Blair, M.W. SNP Marker Diversity in Common Bean (Phaseolus vulgaris L.). Theor. Appl. Genet. 2011, 123, 827–845. [Google Scholar] [CrossRef]
- Song, Q.; Jia, G.; Hyten, D.L.; Jenkins, J.; Hwang, E.Y.; Schroeder, S.G.; Osorno, J.M.; Schmutz, J.; Jackson, S.A.; McClean, P.E.; et al. SNP Assay Development for Linkage Map Construction, Anchoring Whole-Genome Sequence, and Other Genetic and Genomic Applications in Common Bean. G3 Genes Genomes Genet. 2015, 5, 2285–2290. [Google Scholar] [CrossRef]
- Brothers, M.E.; Kelly, J.D. Allozyme Evaluation of Upright Common Bean Genotypes. Euphytica 1993, 67, 65–70. [Google Scholar] [CrossRef]
- McClean, P.E.; Terpstra, J.; McConnell, M.; White, C.; Lee, R.; Mamidi, S. Population Structure and Genetic Differentiation among the USDA Common Bean (Phaseolus vulgaris L.) Core Collection. Genet. Resour. Crop Evol. 2012, 59, 499–515. [Google Scholar] [CrossRef]
- Zhou, D.; Jing, T.; Chen, Y.; Wang, F.; Qi, D.; Feng, R.; Xie, J.; Li, H. Deciphering Microbial Diversity Associated with Fusarium Wilt-Diseased and Disease-Free Banana Rhizosphere Soil. BMC Microbiol. 2019, 19, 161. [Google Scholar] [CrossRef]
- Chitwood-Brown, J.; Vallad, G.E.; Lee, T.G.; Hutton, S.F. Breeding for Resistance to Fusarium Wilt of Tomato: A Review. Genes 2021, 12, 1673. [Google Scholar] [CrossRef]
- Zhang, C.; Wang, W.; Xue, M.; Liu, Z.; Zhang, Q.; Hou, J.; Xing, M.; Wang, R.; Liu, T. The Combination of a Biocontrol Agent Trichoderma Asperellum SC012 and Hymexazol Reduces the Effective Fungicide Dose to Control Fusarium Wilt in Cowpea. J. Fungi 2021, 7, 685. [Google Scholar] [CrossRef]
- Jendoubi, W.; Bouhadida, M.; Boukteb, A.; Béji, M.; Kharrat, M. Fusarium Wilt Affecting Chickpea Crop. Agriculture 2017, 7, 23. [Google Scholar] [CrossRef]
- Holland, J.B.; Nyquist, W.E.; Cervantes-Martínez, C.T.; Janick, J. Estimating and Interpreting Heritability for Plant Breeding: An Update. Plant Breed. Rev. 2003, 22, 9–112. [Google Scholar] [CrossRef]
- Deng, Y.; Ning, Y.; Yang, D.L.; Zhai, K.; Wang, G.L.; He, Z. Molecular Basis of Disease Resistance and Perspectives on Breeding Strategies for Resistance Improvement in Crops. Mol. Plant 2020, 13, 1402–1419. [Google Scholar] [CrossRef]
- Sharma, M.; Ghosh, R.; Telangre, R.; Rathore, A.; Saifulla, M.; Mahalinga, D.M.; Saxena, D.R.; Jain, Y.K. Environmental Influences on Pigeonpea-Fusarium Udum Interactions and Stability of Genotypes to Fusarium Wilt. Front. Plant Sci. 2016, 7, 172780. [Google Scholar] [CrossRef] [PubMed]
- Ndeve, A.D.; Roberts, P.A. Quantitative Phenotyping of Vascular Damage Caused by Fusarium Wilt Disease in Cowpea. bioRxiv 2019. [Google Scholar] [CrossRef]
- Jorben, J.; Rao, A.; Nagappa Chowluru, S.; Tomar, S.; Kumar, N.; Bharadwaj, C.; Siddana-gowda Patil, B.; Ram Soren, K.; Siddana, B.; Jorben, J.; et al. Identification of Multi-Race Fusarium Wilt Resistance in Chickpea (Cicer arietinum L.) Using Rapid Hydroponic Phenotyping. Phytopathol. Mediterr. 2023, 62, 3–15. [Google Scholar] [CrossRef]
- Perseguini, J.M.K.C.; Oblessuc, P.R.; Rosa, J.R.B.F.; Gomes, K.A.; Chiorato, A.F.; Carbonell, S.A.M.; Garcia, A.A.F.; Vianello, R.P.; Benchimol-Reis, L.L. Genome-Wide Association Studies of Anthracnose and Angular Leaf Spot Resistance in Common Bean (Phaseolus vulgaris L.). PLoS ONE 2016, 11, e0150506. [Google Scholar] [CrossRef]
- Monteiro, A.L.R.; Pantaleão, A.S.L.; Badel, J.L.; Soares, P.H.M.; Carneiro, V.Q.; Carneiro, P.C.S.; Carneiro, J.E.S. Genome-Wide Association Study (GWAS) of Phaseolus vulgaris Resistance to Xanthomonas citri pv. fuscans. Plant Pathol. 2021, 70, 1733–1744. [Google Scholar] [CrossRef]
- Delfini, J.; Moda-Cirino, V.; dos Santos Neto, J.; Zeffa, D.M.; Nogueira, A.F.; Ribeiro, L.A.B.; Ruas, P.M.; Gepts, P.; Gonçalves, L.S.A. Genome-Wide Association Study Identifies Genomic Regions for Important Morpho-Agronomic Traits in Mesoamerican Common Bean. Front. Plant Sci. 2021, 12, 748829. [Google Scholar] [CrossRef]
- Evangelou, E.; Ioannidis, J.P.A. Meta-Analysis Methods for Genome-Wide Association Studies and Beyond. Nat. Rev. Genet. 2013, 14, 379–389. [Google Scholar] [CrossRef] [PubMed]
- Van Ooijen, G.; Mayr, G.; Kasiem, M.M.A.; Albrecht, M.; Cornelissen, B.J.C.; Takken, F.L.W. Structure–Function Analysis of the NB-ARC Domain of Plant Disease Resistance Proteins. J. Exp. Bot. 2008, 59, 1383–1397. [Google Scholar] [CrossRef]
- Gupta, S.K.; Rai, A.K.; Kanwar, S.S.; Sharma, T.R. Comparative Analysis of Zinc Finger Proteins Involved in Plant Disease Resistance. PLoS ONE 2012, 7, e42578. [Google Scholar] [CrossRef]
- Jones, D.A.; Jones, J.D.G. The Role of Leucine-Rich Repeat Proteins in Plant Defences. Adv. Bot. Res. 1997, 24, 89–167. [Google Scholar] [CrossRef]
- Xiong, H.; Chen, Y.; Pan, Y.B.; Wang, J.; Lu, W.; Shi, A. A Genome-Wide Association Study and Genomic Prediction for Phakopsora Pachyrhizi Resistance in Soybean. Front. Plant Sci. 2023, 14, 1179357. [Google Scholar] [CrossRef] [PubMed]
- Pottorff, M.; Wanamaker, S.; Ma, Y.Q.; Ehlers, J.D.; Roberts, P.A.; Close, T.J. Genetic and Physical Mapping of Candidate Genes for Resistance to Fusarium oxysporum f. sp. tracheiphilum Race 3 in Cowpea [Vigna unguiculata (L.) Walp]. PLoS ONE 2012, 7, e41600. [Google Scholar] [CrossRef] [PubMed]
- Channale, S.; Thompson, J.P.; Varshney, R.K.; Thudi, M.; Zwart, R.S. Multi-Locus Genome-Wide Association Study of Chickpea Reference Set Identifies Genetic Determinants of Pratylenchus Thornei Resistance. Front. Plant Sci. 2023, 14, 1139574. [Google Scholar] [CrossRef]
- Pal, A.; Chakrabarti, A.; Basak, J. New Motifs within the NB-ARC Domain of R Proteins: Probable Mechanisms of Integration of Geminiviral Signatures within the Host Species of Fabaceae Family and Implications in Conferring Disease Resistance. J. Theor. Biol. 2007, 246, 564–573. [Google Scholar] [CrossRef]
- Schmidt, E.D.L.; Guzzo, F.; Toonen, M.A.J.; De Vries, S.C. A Leucine-Rich Repeat Containing Receptor-like Kinase Marks Somatic Plant Cells Competent to Form Embryos. Development 1997, 124, 2049–2062. [Google Scholar] [CrossRef]
- Li, X.; Han, M.; Zhang, H.; Liu, F.; Pan, Y.; Zhu, J.; Liao, Z.; Chen, X.; Zhang, B. Structures and Biological Functions of Zinc Finger Proteins and Their Roles in Hepatocellular Carcinoma. Biomark. Res. 2022, 10, 2. [Google Scholar] [CrossRef]
- Leipe, D.D.; Koonin, E.V.; Aravind, L. STAND, a Class of P-Loop NTPases Including Animal and Plant Regulators of Programmed Cell Death: Multiple, Complex Domain Architectures, Unusual Phyletic Patterns, and Evolution by Horizontal Gene Transfer. J. Mol. Biol. 2004, 343, 1–28. [Google Scholar] [CrossRef]
- Ojiewo, C.; Monyo, E.; Desmae, H.; Boukar, O.; Mukankusi-Mugisha, C.; Thudi, M.; Pandey, M.K.; Saxena, R.K.; Gaur, P.M.; Chaturvedi, S.K.; et al. Genomics, Genetics and Breeding of Tropical Legumes for Better Livelihoods of Smallholder Farmers. Plant Breed. 2019, 138, 487–499. [Google Scholar] [CrossRef]
- Keller, B.; Ariza-Suarez, D.; de la Hoz, J.; Aparicio, J.S.; Portilla-Benavides, A.E.; Buendia, H.F.; Mayor, V.M.; Studer, B.; Raatz, B. Genomic Prediction of Agronomic Traits in Common Bean (Phaseolus vulgaris L.) Under Environmental Stress. Front. Plant Sci. 2020, 11, 543352. [Google Scholar] [CrossRef]
- Ravelombola, W.S.; Qin, J.; Shi, A.; Nice, L.; Bao, Y.; Lorenz, A.; Orf, J.H.; Young, N.D.; Chen, S. Genome-Wide Association Study and Genomic Selection for Soybean Chlorophyll Content Associated with Soybean Cyst Nematode Tolerance. BMC Genom. 2019, 20, 904. [Google Scholar] [CrossRef]
- Shi, A.; Bhattarai, G.; Xiong, H.; Avila, C.A.; Feng, C.; Liu, B.; Joshi, V.; Stein, L.; Mou, B.; Du Toit, L.J.; et al. Genome-Wide Association Study and Genomic Prediction of White Rust Resistance in USDA GRIN Spinach Germplasm. Hortic. Res. 2022, 9, uhac069. [Google Scholar] [CrossRef]
- Lopez-Cruz, M.; Crossa, J.; Bonnett, D.; Dreisigacker, S.; Poland, J.; Jannink, J.L.; Singh, R.P.; Autrique, E.; de los Campos, G. Increased Prediction Accuracy in Wheat Breeding Trials Using a Marker × Environment Interaction Genomic Selection Model. G3 Genes Genomes Genet. 2015, 5, 569–582. [Google Scholar] [CrossRef] [PubMed]
- Rice, B.R.; Lipka, A.E. Diversifying Maize Genomic Selection Models. Mol. Breed. 2021, 41, 33. [Google Scholar] [CrossRef]
- Cross, H.; Brick, M.A.; Schwartz, H.F.; Panella, L.W.; Byrne, P.F. Inheritance of Resistance to Fusarium Wilt in Two Common Bean Races. Crop Sci. 2000, 40, 954–958. [Google Scholar] [CrossRef]
- Lipka, A.E.; Tian, F.; Wang, Q.; Peiffer, J.; Li, M.; Bradbury, P.J.; Gore, M.A.; Buckler, E.S.; Zhang, Z. GAPIT: Genome Association and Prediction Integrated Tool. Bioinformatics 2012, 28, 2397–2399. [Google Scholar] [CrossRef]
- Tamura, K.; Stecher, G.; Kumar, S. MEGA11: Molecular Evolutionary Genetics Analysis Version 11. Mol. Biol. Evol. 2021, 38, 3022–3027. [Google Scholar] [CrossRef]
- Huang, M.; Liu, X.; Zhou, Y.; Summers, R.M.; Zhang, Z. BLINK: A Package for the next Level of Genome-Wide Association Studies with Both Individuals and Markers in the Millions. Gigascience 2019, 8, giy154. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Ersoz, E.; Lai, C.Q.; Todhunter, R.J.; Tiwari, H.K.; Gore, M.A.; Bradbury, P.J.; Yu, J.; Arnett, D.K.; Ordovas, J.M.; et al. Mixed Linear Model Approach Adapted for Genome-Wide Association Studies. Nat. Genet. 2010, 42, 355–360. [Google Scholar] [CrossRef]
- Jiang, J. Linear and Generalized Linear Mixed Models and Their Applications; Springer: New York, NY, USA, 2007. [Google Scholar] [CrossRef]
- Liu, X.; Huang, M.; Fan, B.; Buckler, E.S.; Zhang, Z. Iterative Usage of Fixed and Random Effect Models for Powerful and Efficient Genome-Wide Association Studies. PLoS Genet. 2016, 12, e1005767. [Google Scholar] [CrossRef]
- Yi, H.; Breheny, P.; Imam, N.; Liu, Y.; Hoeschele, I.; Yi, H. Penalized Multimarker vs. Single-Marker Regression Methods for Genome-Wide Association Studies of Quantitative Traits. Genetics 2015, 199, 205–222. [Google Scholar] [CrossRef] [PubMed]
- Bradbury, P.J.; Zhang, Z.; Kroon, D.E.; Casstevens, T.M.; Ramdoss, Y.; Buckler, E.S. TASSEL: Software for Association Mapping of Complex Traits in Diverse Samples. Bioinformatics 2007, 23, 2633–2635. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Lourenco, D.; Aguilar, I.; Legarra, A.; Misztal, I. Weighting Strategies for Single-Step Genomic BLUP: An Iterative Approach for Accurate Calculation of GEBV and GWAS. Front. Genet. 2016, 7, 210065. [Google Scholar] [CrossRef] [PubMed]
- Waldmann, P. Approximate Bayesian Neural Networks in Genomic Prediction. Genet. Sel. Evol. 2018, 50, 70. [Google Scholar] [CrossRef] [PubMed]
- Shikha, M.; Kanika, A.; Rao, A.R.; Mallikarjuna, M.G.; Gupta, H.S.; Nepolean, T. Genomic Selection for Drought Tolerance Using Genome-Wide SNPs in Maize. Front. Plant Sci. 2017, 8, 550. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| PI_Accession | PI_Name | Origin.Country | Race1_2006_ Disease_Rate | Race4_2006_ Disease_Rate | Rate ≤ 3 Either Race 1 or Race 4 or Both |
|---|---|---|---|---|---|
| PI260418 | PV-3 | Bolivia | 3 | 9 | race 1 ≤ 3 |
| PI387865 | W-941d | Bolivia | 2.7 | 5 | race 1 ≤ 3 |
| PI207207 | Z-#4 | Colombia | 6.8 | 2.7 | race 4 ≤ 3 |
| PI207279 | Chiapas 36-3 | Colombia | 3 | 7 | race 1 ≤ 3 |
| PI207336 | Jalisco 31-1 | Colombia | 2.3 | 3.7 | race 1 ≤ 3 |
| PI313572 | Antioquia 12 | Colombia | 3 | 4 | race 1 ≤ 3 |
| PI313598 | Cauca 38 | Colombia | 2.7 | 4.3 | race 1 ≤ 3 |
| PI209482 | G16837 | Costa Rica | 2 | 1 | race 1 and 4 ≤ 3 |
| PI209498 | G1363 | Costa Rica | 1.7 | 6.3 | race 1 ≤ 3 |
| PI308898 | Line 7 | Costa Rica | 3 | 3.3 | race 1 ≤ 3 |
| PI308908 | Criollo blanco No. 2 | Costa Rica | 3 | 1 | race 1 and 4 ≤ 3 |
| PI309877 | Col. No. 20670, lot #33 | Costa Rica | 1.4 | 2.3 | race 1 and 4 ≤ 3 |
| PI313693 | Col. No. | Ecuador | 6.3 | 3 | race 4 ≤ 3 |
| PI307788 | S-219-R | El Salvador | 7 | 2.5 | race 4 ≤ 3 |
| PI310761 | G2022 | Guatemala | 2.5 | 9 | race 1 ≤ 3 |
| PI310778 | G2031 | Guatemala | 5 | 1 | race 4 ≤ 3 |
| PI311843 | Frijol de gato | Guatemala | 1.7 | race 1 ≤ 3 | |
| PI311853 | Colorado del suelo | Guatemala | 1 | 8.7 | race 1 ≤ 3 |
| PI451917 | Guatemala | 6 | 3 | race 4 ≤ 3 | |
| PI288016 | Negro Nicaraguense | Nicaragua | 1.3 | 1.7 | race 1 and 4 ≤ 3 |
| PI310842 | G2084 | Nicaragua | 3 | 2.3 | race 1 and 4 ≤ 3 |
| SNP | Chr | Position (bp) | LOD [−log(p-Value)] | Associated FW Race | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| GAPIT 3 | TASSEL 5 | t-Test | |||||||||
| Blink | FarmCPU | GLM | MLM | SMR | GLM | MLM | |||||
| ss715650990_Chr04_26314820 | 4 | 26,314,820 | 0.47 | 2.92 | 4.87 | 4.63 | 4.87 | 4.63 | 1.30 | 3.92 | Race 1 |
| ss715647361_Chr04_45301836 | 4 | 45,301,836 | 8.24 | 0.79 | 1.66 | 1.26 | 1.66 | 1.26 | 2.54 | 1.97 | |
| ss715647824_Chr05_275140 | 5 | 275,140 | 2.40 | 3.22 | 5.04 | 4.77 | 5.04 | 4.77 | 2.13 | 3.76 | |
| ss715645682_Chr07_517953 | 7 | 517,953 | 1.07 | 3.04 | 5.07 | 4.83 | 5.07 | 4.83 | 2.09 | 10.08 | |
| ss715646092_Chr08_57870335 | 8 | 57,870,335 | 5.06 | 2.15 | 3.13 | 2.74 | 3.13 | 2.74 | 1.40 | 3.21 | |
| ss715646367_Chr09_29788600 | 9 | 29,788,600 | 11.56 | 3.30 | 5.52 | 5.23 | 5.52 | 5.23 | 2.46 | 3.42 | |
| ss715649363_Chr03_35509497 | 3 | 35,509,497 | 2.78 | 2.85 | 5.73 | 2.34 | 5.59 | 6.11 | 2.10 | 4.16 | Race 4 |
| ss715650990_Chr04_26314820 | 4 | 26,314,820 | 3.55 | 1.60 | 5.77 | 2.81 | 6.42 | 6.69 | 2.81 | 7.26 | |
| ss715645397_Chr05_37965834 | 5 | 37,965,834 | 1.64 | 4.77 | 5.04 | 2.62 | 3.58 | 4.90 | 2.02 | 4.45 | |
| ss715646025_Chr07_48806850 | 7 | 48,806,850 | 2.05 | 0.89 | 5.55 | 2.14 | 5.76 | 5.87 | 1.94 | 4.59 | |
| ss715645623_Chr09_32650091 | 9 | 32,650,091 | 5.05 | 4.18 | 2.85 | 2.75 | 1.93 | 2.70 | 2.25 | 5.50 | |
| ss715647542_Chr11_44755455 | 11 | 44,755,455 | 2.72 | 6.01 | 2.39 | 2.81 | 0.01 | 2.50 | 2.84 | 0.31 | |
| Gene | Chr | Gene Size (bp) | Gene Annotation | Comment | Closest SNP Marker | Fusarium Wilt Race | Distance (kb) |
|---|---|---|---|---|---|---|---|
| Phvul.004G016532 | 4 | 4457 | NB-ARC domain-containing disease resistance protein | R gene | ss715647806_Chr04_1827663 | race1 | <30 kb |
| Phvul.004G016800 | 4 | 5948 | HD domain-containing metal-dependent phosphohydrolase family protein | ss715647808_Chr04_1845589 | <1 kb | ||
| Phvul.004G151100 | 4 | 6906 | Zinc finger (Ran-binding) family protein | R gene | ss715647361_Chr04_45301836 | 0 | |
| Phvul.005G003400 | 5 | 2088 | SNARE-like superfamily protein | ss715647824_Chr05_275140 | <2 kb | ||
| Phvul.007G007100 | 7 | 3311 | Nitrilase/cyanide hydratase and apolipoprotein N-acyltransferase family protein | ss715645682_Chr07_517953 | <2 kb | ||
| Phvul.007G008400 | 7 | 2702 | Peroxidase superfamily protein | ss715645685_Chr07_606814 | <1 kb | ||
| Phvul.007G008300 | 7 | 3835 | Pre-mRNA-splicing factor 3 | <2 kb | |||
| Phvul.008G228500 | 8 | 4138 | Protein of Unknown Function (DUF239) | ss715646092_Chr08_57870335 | 0 | ||
| Phvul.009G153600 | 9 | 7435 | Amino acid permease 2 | ss715648883_Chr09_22785976 | 0 | ||
| Phvul.009G195900 | 9 | 2567 | Leucine-rich repeat protein kinase family protein | R gene | ss715646367_Chr09_29788600 | <10 kb | |
| Phvul.003G129400 | 3 | 2678 | Leucine-rich repeat protein kinase family protein | R gene | ss715650616_Chr03_32369163 | race4 | <5 kb |
| Phvul.004G073900 | 4 | 13,305 | Polynucleotidyl transferase, ribonuclease H fold protein with HRDC domain | ss715648302_Chr04_12157925 | <2 kb | ||
| Phvul.005G045400 | 5 | 2233 | P-loop-containing nucleoside triphosphate hydrolase superfamily protein | R gene | ss715650411_Chr05_4808704 | <3 kb | |
| Phvul.005G137400 | 5 | 4765 | Carbamoyl phosphate synthetase B | ss715645397_Chr05_37965834 | <2 kb | ||
| Phvul.009G153600 | 9 | 7435 | Amino acid permease 2 | ss715648883_Chr09_22785976 | 0 | ||
| Phvul.009G216500 | 9 | 18,256 | RNA binding | ss715645623_Chr09_32650091 | <1 kb | ||
| Phvul.010G071766 | 10 | 6447 | Nucleotidyltransferase family protein | ss715650855_Chr10_32133091 | 0 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chiwina, K.; Xiong, H.; Bhattarai, G.; Dickson, R.W.; Phiri, T.M.; Chen, Y.; Alatawi, I.; Dean, D.; Joshi, N.K.; Chen, Y.; et al. Genome-Wide Association Study and Genomic Prediction of Fusarium Wilt Resistance in Common Bean Core Collection. Int. J. Mol. Sci. 2023, 24, 15300. https://doi.org/10.3390/ijms242015300
Chiwina K, Xiong H, Bhattarai G, Dickson RW, Phiri TM, Chen Y, Alatawi I, Dean D, Joshi NK, Chen Y, et al. Genome-Wide Association Study and Genomic Prediction of Fusarium Wilt Resistance in Common Bean Core Collection. International Journal of Molecular Sciences. 2023; 24(20):15300. https://doi.org/10.3390/ijms242015300
Chicago/Turabian StyleChiwina, Kenani, Haizheng Xiong, Gehendra Bhattarai, Ryan William Dickson, Theresa Makawa Phiri, Yilin Chen, Ibtisam Alatawi, Derek Dean, Neelendra K. Joshi, Yuyan Chen, and et al. 2023. "Genome-Wide Association Study and Genomic Prediction of Fusarium Wilt Resistance in Common Bean Core Collection" International Journal of Molecular Sciences 24, no. 20: 15300. https://doi.org/10.3390/ijms242015300