
Notopterol Suppresses IL-17-Induced Proliferation and Invasion of A549 Lung Adenocarcinoma Cells via Modulation of STAT3, NF-κB, and AP-1 Activation


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Effect of Notopterol on the Cytotoxicity of A549 Lung Adenocarcinoma Cells
2.2. Notopterol Inhibits IL-17 Stimulation of A549 Cell Proliferation
2.3. Notopterol-Induced Cell Cycle Arrest
2.4. Notopterol Suppressed IL-17-Induced A549 Cell Invasion and Migration
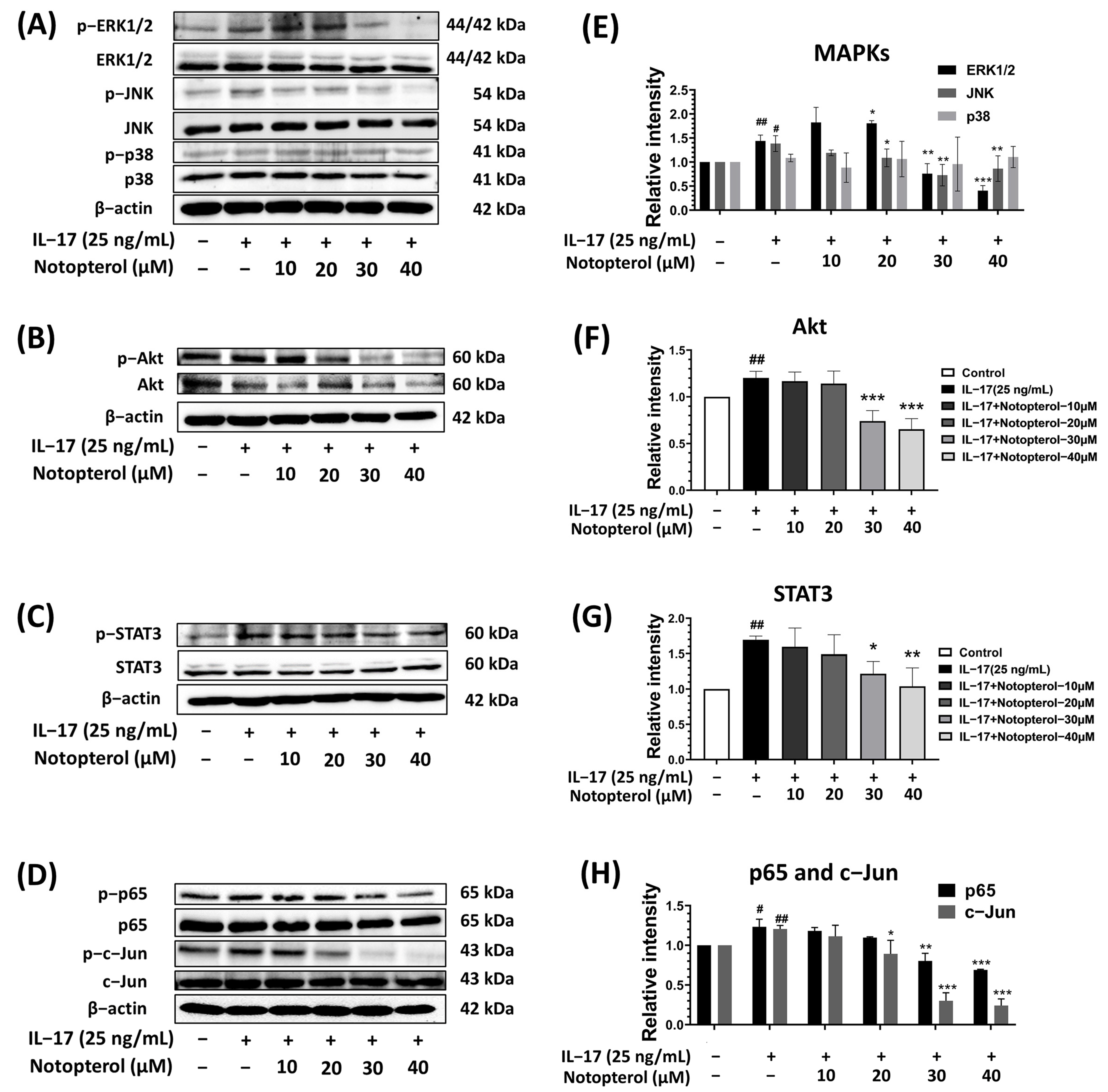
2.5. Effect of Notopterol on IL-17-Induced Oncogenic Signaling Pathways
3. Discussion
4. Materials and Methods
4.1. Chemicals and Reagents
4.2. Cells and Cell Cultures
4.3. Cell Viability Assay
4.4. Colony Formation
4.5. Cell Cycle Analysis
4.6. Apoptosis Assay
4.7. Cell Migration Assay
4.8. Cell Invasion Assay
4.9. Gelatin Zymography Assay
4.10. Western Blot Analysis
4.11. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| NSCLC | Non-small-cell lung cancer |
| IL-17 | Interleukin-17 |
| STAT3 | Signal transducer and activator of transcription 3 |
| MAPKs | Mitogen-activated protein kinases |
| Akt | AKT serine/threonine kinase 1 |
| NF-κB | Nuclear factor kappa B |
| EMT | Epithelial–mesenchymal transition |
| Ki-67 | Antigen Kiel 67 |
| MCM2 | Mini-chromosome maintenance protein 2 |
| PCNA | Proliferating cell nuclear antigen |
| CDKs | Cyclin-dependent kinases |
| ERK | Extracellular regulated kinase |
| p38 | p38 mitogen-activated protein kinases |
| JNK | c-Jun N-terminal kinase |
| AP-1 | Activator protein 1 |
| ECM | Extracellular matrix |
References
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global cancer statistics 2020: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA A Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef] [PubMed]
- Herbst, R.S.; Morgensztern, D.; Boshoff, C. The biology and management of non-small cell lung cancer. Nature 2018, 553, 446–454. [Google Scholar] [CrossRef] [PubMed]
- Bade, B.C.; Cruz, C.S.D. Lung cancer 2020: Epidemiology, etiology, and prevention. Clin. Chest Med. 2020, 41, 1–24. [Google Scholar] [CrossRef] [PubMed]
- Sui, H.; Ma, N.; Wang, Y.; Li, H.; Liu, X.; Su, Y.; Yang, J. Anti-PD-1/PD-L1 therapy for non-small-cell lung cancer: Toward personalized medicine and combination strategies. J. Immunol. Res. 2018, 2018, 6984948. [Google Scholar] [CrossRef]
- Zappa, C.; Mousa, S.A. Non-small cell lung cancer: Current treatment and future advances. Transl. Lung Cancer Res. 2016, 5, 288. [Google Scholar] [CrossRef]
- Chen, X.; Wan, J.; Liu, J.; Xie, W.; Diao, X.; Xu, J.; Zhu, B.; Chen, Z. Increased IL-17-producing cells correlate with poor survival and lymphangiogenesis in NSCLC patients. Lung Cancer 2010, 69, 348–354. [Google Scholar] [CrossRef]
- Li, Q.; Han, Y.; Fei, G.; Guo, Z.; Ren, T.; Liu, Z. IL-17 promoted metastasis of non-small-cell lung cancer cells. Immunol. Lett. 2012, 148, 144–150. [Google Scholar] [CrossRef]
- Murugaiyan, G.; Saha, B. Protumor vs antitumor functions of IL-17. J. Immunol. 2009, 183, 4169–4175. [Google Scholar] [CrossRef]
- Wu, F.; Xu, J.; Huang, Q.; Han, J.; Duan, L.; Fan, J.; Lv, Z.; Guo, M.; Hu, G.; Chen, L.; et al. The role of interleukin-17 in lung cancer. Mediat. Inflamm. 2016, 2016, 8494079. [Google Scholar] [CrossRef]
- Xu, C.; Hao, K.; Yu, L.; Zhang, X. Serum interleukin-17 as a diagnostic and prognostic marker for non-small cell lung cancer. Biomarkers 2014, 19, 287–290. [Google Scholar] [CrossRef]
- Xu, C.; Yu, L.; Zhan, P.; Zhang, Y. Elevated pleural effusion IL-17 is a diagnostic marker and outcome predictor in lung cancer patients. Eur. J. Med. Res. 2014, 19, 23. [Google Scholar] [CrossRef]
- Wu, L.; Awaji, M.; Saxena, S.; Varney, M.L.; Sharma, B.; Singh, R.K. IL-17–CXC chemokine receptor 2 axis facilitates breast cancer progression by up-regulating neutrophil recruitment. Am. J. Pathol. 2020, 190, 222–233. [Google Scholar] [CrossRef] [PubMed]
- Zhao, C.; Li, Y.; Zhang, W.; Zhao, D.; Ma, L.; Ma, P.; Yang, F.; Wang, Y.; Shu, Y.; Qiu, W. IL-17 induces NSCLC A549 cell proliferation via the upregulation of HMGA1, resulting in an increased cyclin D1 expression. Int. J. Oncol. 2018, 52, 1579–1592. [Google Scholar] [CrossRef] [PubMed]
- Xu, B.; Guenther, J.F.; Pociask, D.A.; Wang, Y.; Kolls, J.K.; You, Z.; Chandrasekar, B.; Shan, B.; Sullivan, D.E.; Morris, G.F. Promotion of lung tumor growth by interleukin-17. Am. J. Physiol.-Lung Cell. Mol. Physiol. 2014, 307, L497–L508. [Google Scholar] [CrossRef]
- Huang, Q.; Du, J.; Fan, J.; Lv, Z.; Qian, X.; Zhang, X.; Han, J.; Chen, C.; Wu, F.; Jun, Y. The effect of proinflammatory cytokines on IL-17RA expression in NSCLC. Med. Oncol. 2014, 31, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Gu, K.; Li, M.-M.; Shen, J.; Liu, F.; Cao, J.-Y.; Jin, S.; Yu, Y. Interleukin-17-induced EMT promotes lung cancer cell migration and invasion via NF-κB/ZEB1 signal pathway. Am. J. Cancer Res. 2015, 5, 1169. [Google Scholar]
- Lin, Z.; Huang, Q.; Liu, J.; Wang, H.; Zhang, X.; Zhu, Z.; Zhang, W.; Wei, Y.; Liu, Z.; Du, W. Interleukin-17D promotes lung cancer progression by inducing tumor-associated macrophage infiltration via the p38 MAPK signaling pathway. Aging 2022, 14, 6149. [Google Scholar] [CrossRef]
- Yang, Y.-F.; Lee, Y.-C.; Lo, S.; Chung, Y.-N.; Hsieh, Y.-C.; Chiu, W.-C.; Yuan, S.-S. A positive feedback loop of IL-17B-IL-17RB activates ERK/β-catenin to promote lung cancer metastasis. Cancer Lett. 2018, 422, 44–55. [Google Scholar] [CrossRef]
- Azietaku, J.T.; Ma, H.; Yu, X.; Li, J.; Oppong, M.B.; Cao, J.; An, M.; Chang, Y. A review of the ethnopharmacology, phytochemistry and pharmacology of Notopterygium incisum. J. Ethnopharmacol. 2017, 202, 241–255. [Google Scholar] [CrossRef]
- Huang, L.; Li, H.; Huang, S.; Wang, S.; Liu, Q.; Luo, L.; Gan, S.; Fu, G.; Zou, P.; Chen, G.; et al. Notopterol attenuates monocrotaline-induced pulmonary arterial hypertension in rat. Front. Cardiovasc. Med. 2022, 9, 859422. [Google Scholar] [CrossRef]
- Wu, S.-B.; Pang, F.; Wen, Y.; Zhang, H.-F.; Zhao, Z.; Hu, J.-F. Antiproliferative and apoptotic activities of linear furocoumarins from Notopterygium incisum on cancer cell lines. Planta Medica 2010, 76, 82–85. [Google Scholar] [CrossRef]
- Huang, Q.; Wang, L.; Ran, Q.; Wang, J.; Wang, C.; He, H.; Li, L.; Qi, H. Notopterol-induced apoptosis and differentiation in human acute myeloid leukemia HL-60 cells. Drug Des. Dev. Ther. 2019, 13, 1927–1940. [Google Scholar] [CrossRef]
- Huang, T.-Y.; Yang, C.-K.; Chen, M.-Y.; Yadav, V.K.; Fong, I.-H.; Yeh, C.-T.; Cherng, Y.-G. Furanocoumarin Notopterol: Inhibition of Hepatocellular Carcinogenesis through Suppression of Cancer Stemness Signaling and Induction of Oxidative Stress-Associated Cell Death. Nutrients 2023, 15, 2447. [Google Scholar] [CrossRef]
- Wang, Q.; Zhou, X.; Yang, L.; Zhao, Y.; Chew, Z.; Xiao, J.; Liu, C.; Zheng, X.; Zheng, Y.; Shi, Q.; et al. The natural compound notopterol binds and targets JAK2/3 to ameliorate inflammation and arthritis. Cell Rep. 2020, 32, 108158. [Google Scholar] [CrossRef] [PubMed]
- Wu, P.; Wu, D.; Ni, C.; Ye, J.; Chen, W.; Hu, G.; Wang, Z.; Wang, C.; Zhang, Z.; Xia, W.; et al. γδT17 cells promote the accumulation and expansion of myeloid-derived suppressor cells in human colorectal cancer. Immunity 2014, 40, 785–800. [Google Scholar] [CrossRef] [PubMed]
- Yamada, Y.; Saito, H.; Ikeguchi, M. Prevalence and clinical relevance of Th17 cells in patients with gastric cancer. J. Surg. Res. 2012, 178, 685–691. [Google Scholar] [CrossRef] [PubMed]
- Coffelt, S.B.; Kersten, K.; Doornebal, C.W.; Weiden, J.; Vrijland, K.; Hau, C.-S.; Verstegen, N.; Ciampricotti, M.; Hawinkels, L.; Jonkers, J.; et al. IL-17-producing γδ T cells and neutrophils conspire to promote breast cancer metastasis. Nature 2015, 522, 345–348. [Google Scholar] [CrossRef]
- Phillips, J.D.; Knab, L.M.; Blatner, N.R.; Haghi, L.; DeCamp, M.M.; Meyerson, S.L.; Heiferman, M.J.; Heiferman, J.R.; Gounari, F.; Bentrem, D.J.; et al. Preferential expansion of pro-inflammatory Tregs in human non-small cell lung cancer. Cancer Immunol. Immunother. 2015, 64, 1185–1191. [Google Scholar] [CrossRef]
- Punt, S.; Langenhoff, J.M.; Putter, H.; Fleuren, G.J.; Gorter, A.; Jordanova, E.S. The correlations between IL-17 vs. Th17 cells and cancer patient survival: A systematic review. Oncoimmunology 2015, 4, e984547. [Google Scholar] [CrossRef]
- Sakaue-Sawano, A.; Kobayashi, T.; Ohtawa, K.; Miyawaki, A. Drug-induced cell cycle modulation leading to cell-cycle arrest, nuclear mis-segregation, or endoreplication. BMC Cell Biol. 2011, 12, 2. [Google Scholar] [CrossRef]
- Bai, J.; Li, Y.; Zhang, G. Cell cycle regulation and anticancer drug discovery. Cancer Biol. Med. 2017, 14, 348. [Google Scholar]
- Xie, D.; Pei, Q.; Li, J.; Wan, X.; Ye, T. Emerging role of E2F family in cancer stem cells. Front. Oncol. 2021, 11, 723137. [Google Scholar] [CrossRef] [PubMed]
- Ren, B.; Cam, H.; Takahashi, Y.; Volkert, T.; Terragni, J.; Young, R.A.; Dynlacht, B.D. E2F integrates cell cycle progression with DNA repair, replication, and G2/M checkpoints. Genes Dev. 2002, 16, 245–256. [Google Scholar] [CrossRef] [PubMed]
- Kent, L.N.; Bae, S.; Tsai, S.-Y.; Tang, X.; Srivastava, A.; Koivisto, C.; Martin, C.K.; Ridolfi, E.; Miller, G.C.; Zorko, S.M.; et al. Dosage-dependent copy number gains in E2f1 and E2f3 drive hepatocellular carcinoma. J. Clin. Investig. 2017, 127, 830–842. [Google Scholar] [CrossRef] [PubMed]
- Bryant, C.S.; Kumar, S.; Chamala, S.; Shah, J.; Pal, J.; Haider, M.; Seward, S.; Qazi, A.M.; Morris, R.; Semaan, A.; et al. Sulforaphane induces cell cycle arrest by protecting RB-E2F-1 complex in epithelial ovarian cancer cells. Mol. Cancer 2010, 9, 1–9. [Google Scholar] [CrossRef]
- Oliveira, A.R.; Beyer, G.; Chugh, R.; Skube, S.J.; Majumder, K.; Banerjee, S.; Sangwan, V.; Dawra, R.K.; Subramanian, S.; Saluja, A.K. Triptolide abrogates growth of colon cancer and induces cell cycle arrest by inhibiting transcriptional activation of E2F. Lab. Investig. 2015, 95, 648–659. [Google Scholar] [CrossRef]
- Lin, J.-T.; Li, H.-Y.; Chang, N.-S.; Lin, C.-H.; Chen, Y.-C.; Lu, P.-J. WWOX suppresses prostate cancer cell progression through cyclin D1-mediated cell cycle arrest in the G1 phase. Cell Cycle 2015, 14, 408–416. [Google Scholar] [CrossRef]
- Seyfried, T.N.; Huysentruyt, L.C. On the origin of cancer metastasis. Crit. Rev. Oncog. 2013, 18, 43–73. [Google Scholar] [CrossRef]
- Tarin, D. Cell and tissue interactions in carcinogenesis and metastasis and their clinical significance. In Seminars in Cancer Biology; Elsevier: Amsterdam, The Netherlands, 2011. [Google Scholar]
- Bogaczewicz, J.; Sysa-Jedrzejowska, A.; Woźniacka, A. Role of matrix metalloproteinases in primary systemic vasculitis. Pol. Merkur. Lek. Organ Pol. Tow. Lek. 2008, 24, 85–89. [Google Scholar]
- Jabłońska-Trypuć, A.; Matejczyk, M.; Rosochacki, S. Matrix metalloproteinases (MMPs), the main extracellular matrix (ECM) enzymes in collagen degradation, as a target for anticancer drugs. J. Enzym. Inhib. Med. Chem. 2016, 31 (Suppl. S1), 177–183. [Google Scholar] [CrossRef]
- Chae, Y.K.; Chang, S.; Ko, T.; Anker, J.; Agte, S.; Iams, W.; Choi, W.M.; Lee, K.; Cruz, M. Epithelial-mesenchymal transition (EMT) signature is inversely associated with T-cell infiltration in non-small cell lung cancer (NSCLC). Sci. Rep. 2018, 8, 2918. [Google Scholar] [CrossRef] [PubMed]
- Pastushenko, I.; Blanpain, C. EMT transition states during tumor progression and metastasis. Trends Cell Biol. 2019, 29, 212–226. [Google Scholar] [CrossRef] [PubMed]
- Huang, Q.; Han, J.; Fan, J.; Duan, L.; Guo, M.; Lv, Z.; Hu, G.; Chen, L.; Wu, F.; Tao, X.; et al. IL-17 induces EMT via Stat3 in lung adenocarcinoma. Am. J. Cancer Res. 2016, 6, 440. [Google Scholar]
- Guo, Y.J.; Pan, W.W.; Liu, S.B.; Shen, Z.F.; Xu, Y.; Hu, L.L. ERK/MAPK signalling pathway and tumorigenesis. Exp. Ther. Med. 2020, 19, 1997–2007. [Google Scholar] [CrossRef]
- Liang, J.; Li, H.; Han, J.; Jiang, J.; Wang, J.; Li, Y.; Feng, Z.; Zhao, R.; Sun, S.; Lv, B.; et al. Mex3a interacts with LAMA2 to promote lung adenocarcinoma metastasis via PI3K/AKT pathway. Cell Death Dis. 2020, 11, 614. [Google Scholar] [CrossRef] [PubMed]
- Hwang, S.-Y.; Kim, J.-Y.; Kim, K.-W.; Park, M.-K.; Moon, Y.; Kim, W.-U.; Kim, H.-Y. IL-17 induces production of IL-6 and IL-8 in rheumatoid arthritis synovial fibroblasts via NF-κB-and PI3-kinase/Akt-dependent pathways. Arthritis Res. Ther. 2004, 6, 1–9. [Google Scholar] [CrossRef]
- Chen, D.; Wang, Q.; Li, Y.; Sun, P.; Kuek, V.; Yuan, J.; Yang, J.; Wen, L.; Wang, H.; Xu, J.; et al. Notopterol attenuates estrogen deficiency-induced osteoporosis via repressing RANKL signaling and reactive oxygen species. Front. Pharmacol. 2021, 12, 664836. [Google Scholar] [CrossRef]
- Bai, Y.; Li, H.; Lv, R. Interleukin-17 activates JAK2/STAT3, PI3K/Akt and nuclear factor-κB signaling pathway to promote the tumorigenesis of cervical cancer. Exp. Ther. Med. 2021, 22, 1–11. [Google Scholar] [CrossRef]
- Chen, X.; Xie, Q.; Cheng, X.; Diao, X.; Cheng, Y.; Liu, J.; Xie, W.; Chen, Z.; Zhu, B. Role of interleukin-17 in lymphangiogenesis in non-small-cell lung cancer: Enhanced production of vascular endothelial growth factor C in non-small-cell lung carcinoma cells. Cancer Sci. 2010, 101, 2384–2390. [Google Scholar] [CrossRef]
- Yuan, S.; Zhang, S.; Zhuang, Y.; Zhang, H.; Bai, J.; Hou, Q. Interleukin-17 stimulates STAT3-mediated endothelial cell activation for neutrophil recruitment. Cell. Physiol. Biochem. 2015, 36, 2340–2356. [Google Scholar] [CrossRef]
- Liu, T.; Zong, S.; Jiang, Y.; Zhao, R.; Wang, J.; Hua, Q. Neutrophils promote larynx squamous cell carcinoma progression via activating the IL-17/JAK/STAT3 pathway. J. Immunol. Res. 2021, 2021, 8078646. [Google Scholar] [CrossRef] [PubMed]
- Xu, N.-W.; Chen, Y.; Chen, Y.-J.; Fan, Z.-M.; Liu, M.; Li, L.-J. Inhibition of JAK2/STAT3 signaling pathway suppresses proliferation of Burkitt’s lymphoma Raji cells via cell cycle progression, apoptosis, and oxidative stress by modulating HSP70. Med. Sci. Monit. Int. Med. J. Exp. Clin. Res. 2018, 24, 6255. [Google Scholar] [CrossRef]
- Shin, S.S.; Won, S.Y.; Noh, D.H.; Hwang, B.; Kim, W.J.; Moon, S.K. Morin inhibits proliferation, migration, and invasion of bladder cancer EJ cells via modulation of signaling pathways, cell cycle regulators, and transcription factor-mediated MMP-9 expression. Drug Dev. Res. 2017, 78, 81–90. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Tian, S.; Liu, M.; Jian, L.; Zhao, L. Wogonin inhibits the proliferation and invasion, and induces the apoptosis of HepG2 and Bel7402 HCC cells through NF-κB/Bcl-2, EGFR and EGFR downstream ERK/AKT signaling. Int. J. Mol. Med. 2016, 38, 1250–1256. [Google Scholar] [CrossRef] [PubMed]
- Ooppachai, C.; Limtrakul, P.; Yodkeeree, S. Dicentrine potentiates TNF-α-induced apoptosis and suppresses invasion of A549 lung adenocarcinoma cells via modulation of NF-κB and AP-1 activation. Molecules 2019, 24, 4100. [Google Scholar] [CrossRef]
- Chen, J.; Liao, M.-y.; Gao, X.-l.; Zhong, Q.; Tang, T.-t.; Yu, X.; Liao, Y.-h.; Cheng, X. IL-17A induces pro-inflammatory cytokines production in macrophages via MAPKinases, NF-κB and AP-1. Cell Physiol. Biochem. 2013, 32, 1265–1274. [Google Scholar] [CrossRef]
- Wang, B.; Zhao, C.-H.; Sun, G.; Zhang, Z.-W.; Qian, B.-M.; Zhu, Y.-F.; Cai, M.-Y.; Pandey, S.; Zhao, D.; Wang, Y.-W.; et al. IL-17 induces the proliferation and migration of glioma cells through the activation of PI3K/Akt1/NF-κB-p65. Cancer Lett. 2019, 447, 93–104. [Google Scholar] [CrossRef]
- Mohankumar, K.; Francis, A.P.; Pajaniradje, S.; Rajagopalan, R. Synthetic curcumin analog: Inhibiting the invasion, angiogenesis, and metastasis in human laryngeal carcinoma cells via NF-kB pathway. Mol. Biol. Rep. 2021, 48, 6065–6074. [Google Scholar] [CrossRef]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Inthanon, S.; Dejkriengkraikul, P.; Yodkeeree, S. Notopterol Suppresses IL-17-Induced Proliferation and Invasion of A549 Lung Adenocarcinoma Cells via Modulation of STAT3, NF-κB, and AP-1 Activation. Int. J. Mol. Sci. 2023, 24, 15057. https://doi.org/10.3390/ijms242015057
Inthanon S, Dejkriengkraikul P, Yodkeeree S. Notopterol Suppresses IL-17-Induced Proliferation and Invasion of A549 Lung Adenocarcinoma Cells via Modulation of STAT3, NF-κB, and AP-1 Activation. International Journal of Molecular Sciences. 2023; 24(20):15057. https://doi.org/10.3390/ijms242015057
Chicago/Turabian StyleInthanon, Sirinada, Pornngarm Dejkriengkraikul, and Supachai Yodkeeree. 2023. "Notopterol Suppresses IL-17-Induced Proliferation and Invasion of A549 Lung Adenocarcinoma Cells via Modulation of STAT3, NF-κB, and AP-1 Activation" International Journal of Molecular Sciences 24, no. 20: 15057. https://doi.org/10.3390/ijms242015057