
The KDET Motif in the Intracellular Domain of the Cell Adhesion Molecule L1 Interacts with Several Nuclear, Cytoplasmic, and Mitochondrial Proteins Essential for Neuronal Functions
Abstract
:1. Introduction
2. Results
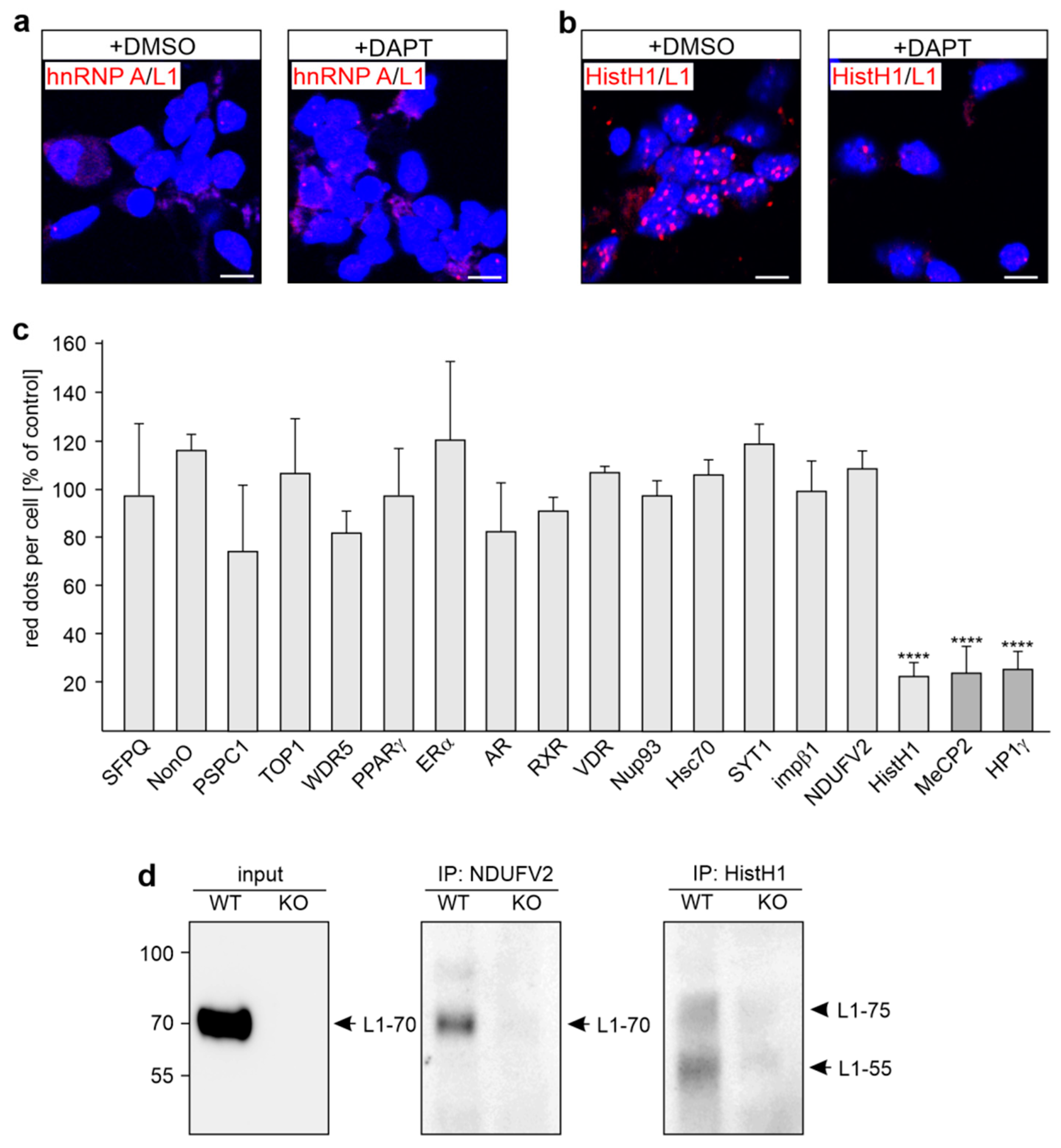
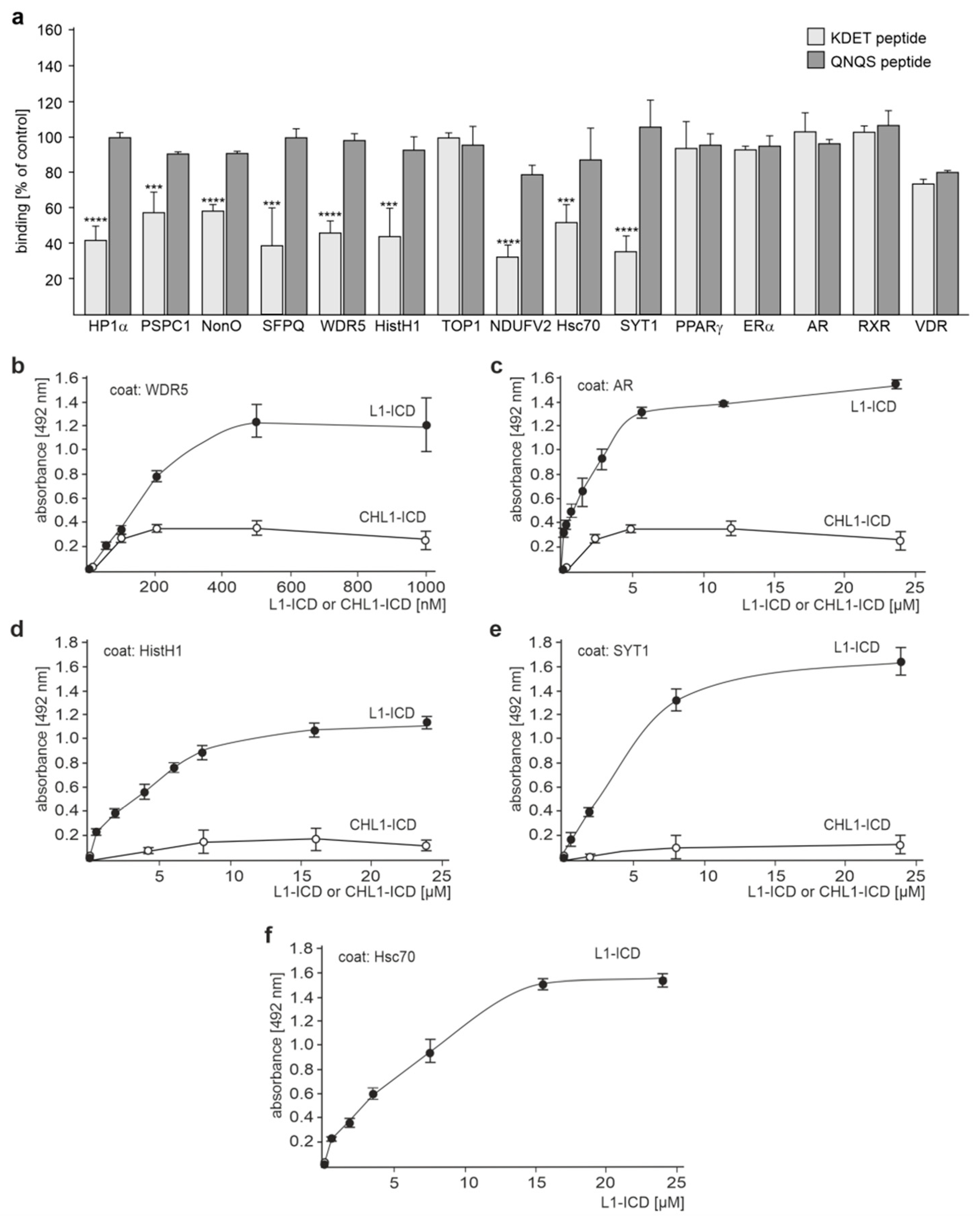
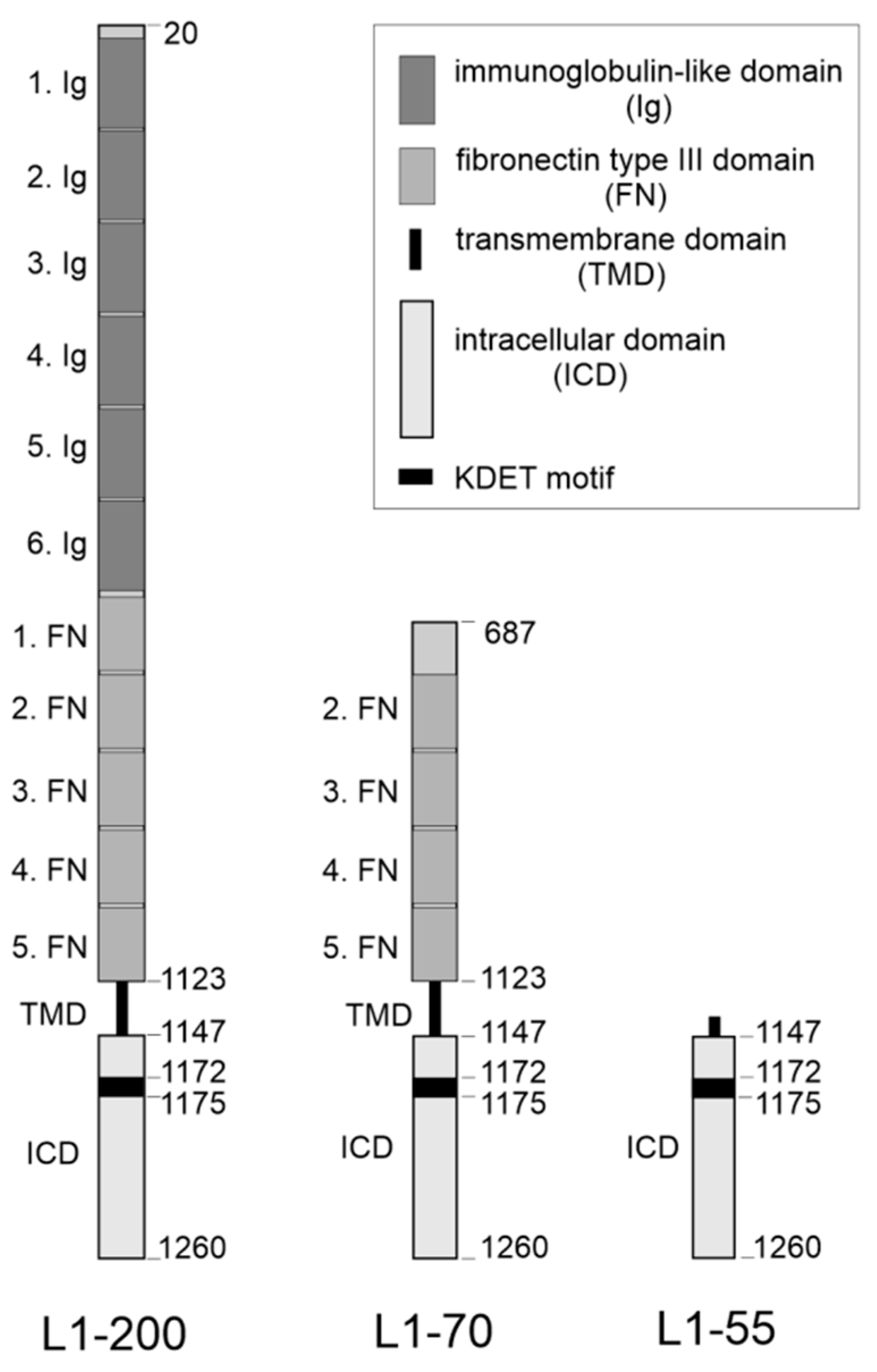
2.1. Several Binding Partners Interact with L1 via the KDET Motif
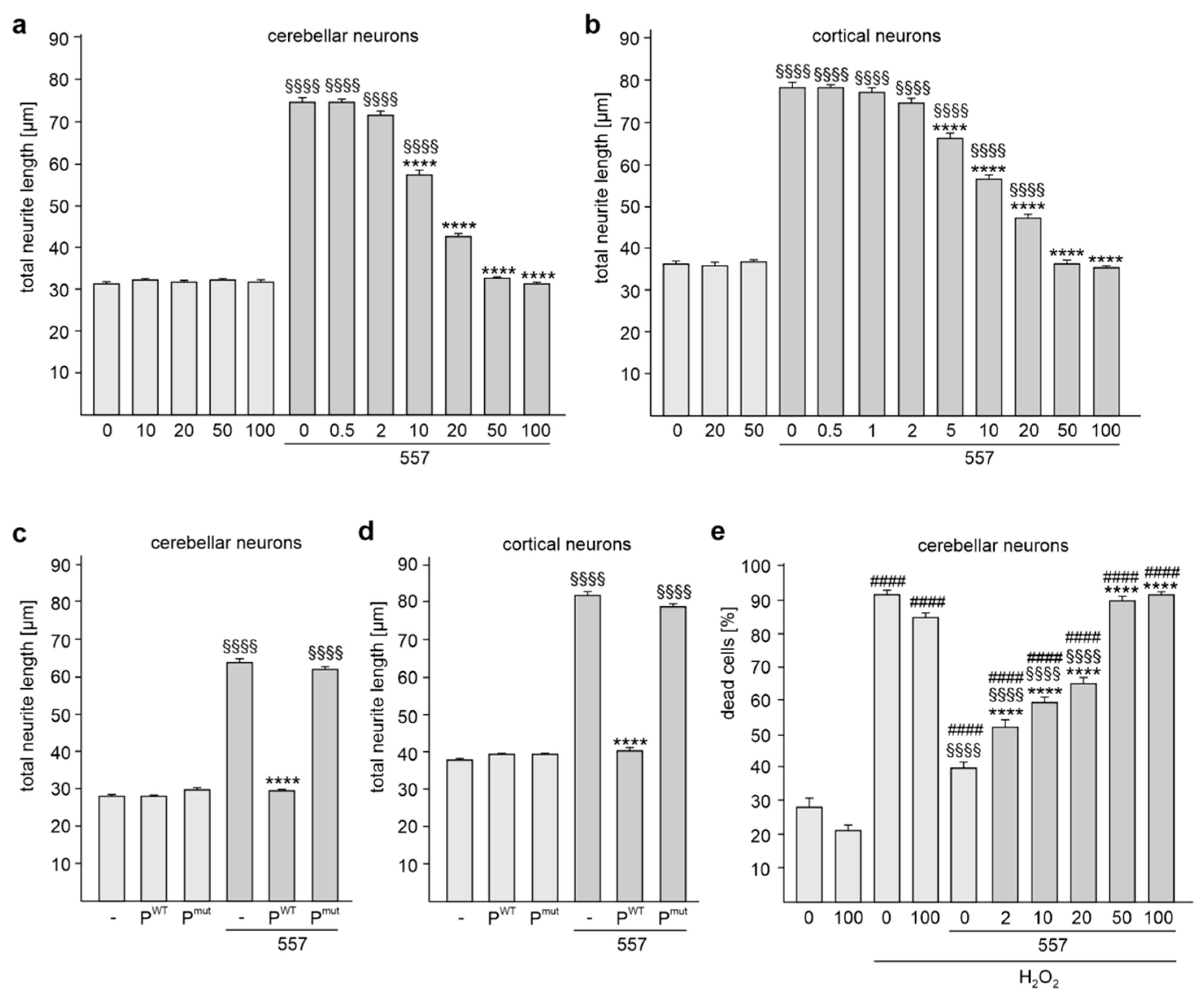
2.2. The KDET Motif Is Essential for L1-Dependent Neurite Outgrowth and Neuronal Survival
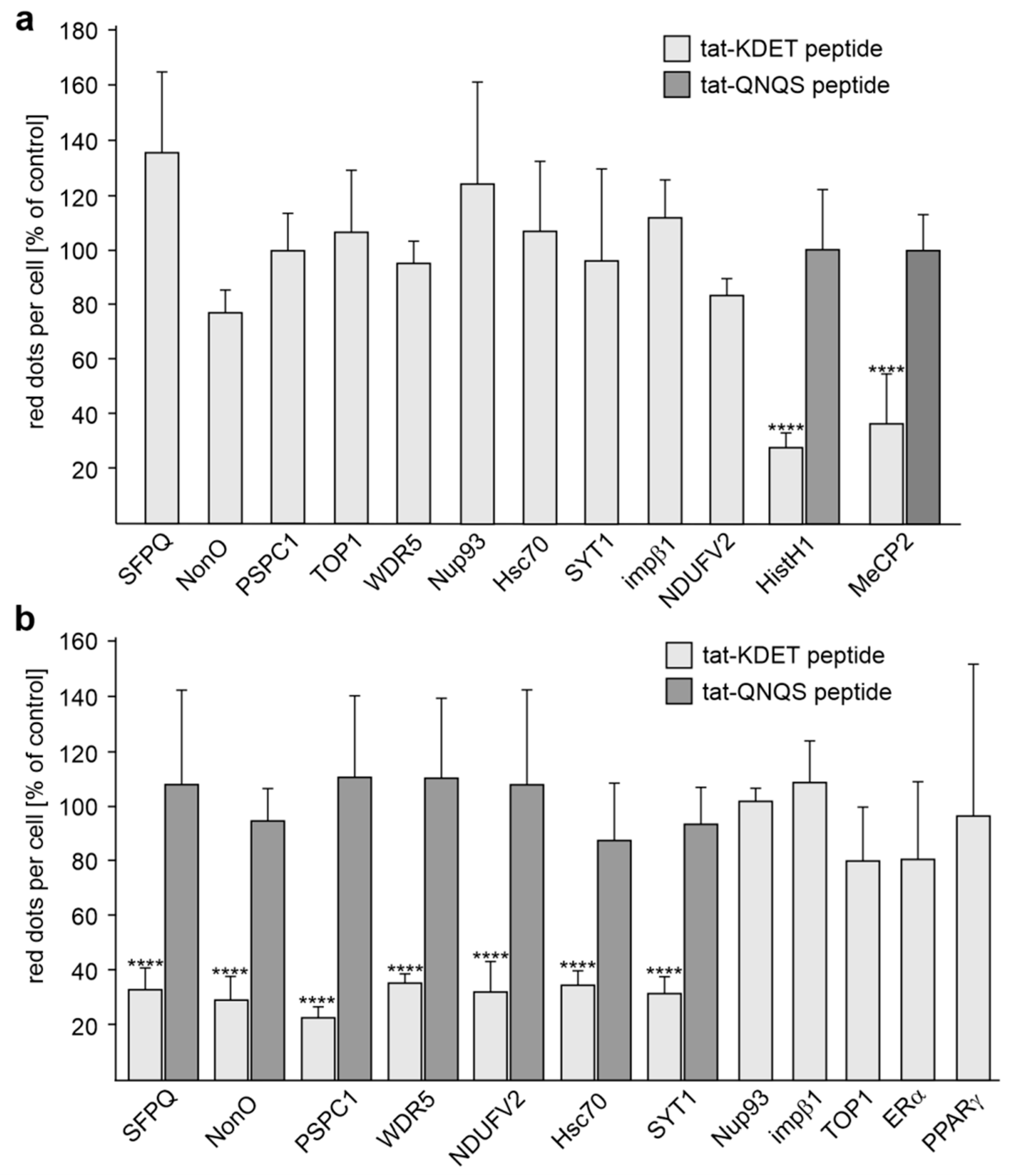
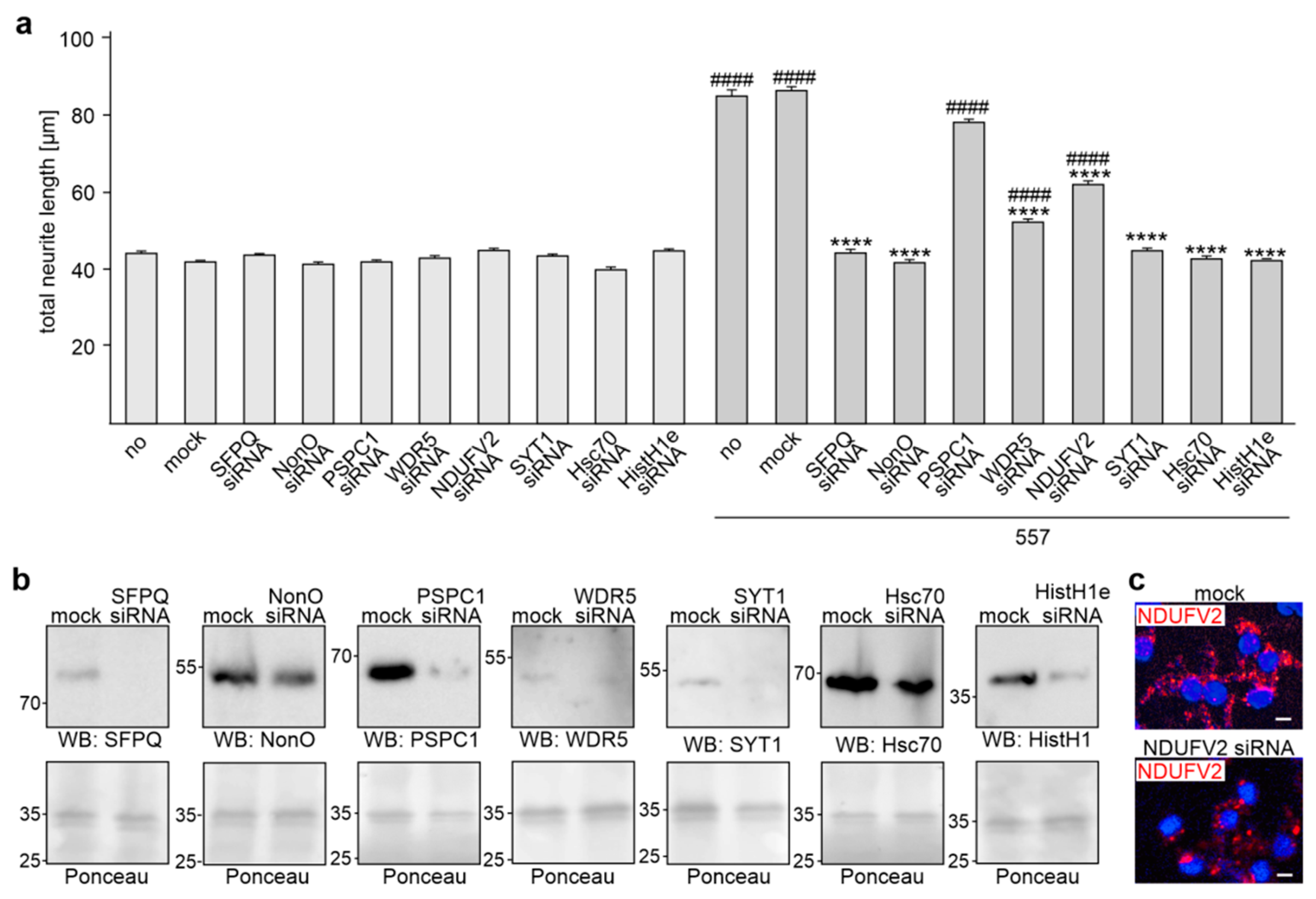
2.3. Reduction of HistH1e, NDUFV2, SFPQ, NonO, WDR5, Hsc70, or SYT1 Expression Decreases L1-Dependent Neurite Outgrowth
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Reagents and Antibodies
4.3. ELISA
4.4. Cultures of Cerebellar and Cortical Neurons
4.5. Proximity Ligation Assay and Immunostaining with Cerebellar and Cortical Neurons
4.6. Determination of Neurite Outgrowth and Neuronal Survival
4.7. Immunorecipitation and Western Blot Analysis
4.8. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Loers, G.; Schachner, M. Recognition molecules and neural repair. J. Neurochem. 2007, 101, 865–882. [Google Scholar] [CrossRef] [PubMed]
- Maness, P.F.; Schachner, M. Neural recognition molecules of the immunoglobulin superfamily: Signaling transducers of axon guidance and neuronal migration. Nat. Neurosci. 2007, 10, 19–26. [Google Scholar] [CrossRef] [PubMed]
- Schafer, M.K.; Altevogt, P. L1CAM malfunction in the nervous system and human carcinomas. Cell Mol. Life Sci. 2010, 67, 2425–2437. [Google Scholar] [CrossRef] [PubMed]
- Sytnyk, V.; Leshchyns’ka, I.; Schachner, M. Neural cell adhesion molecules of the immunoglobulin superfamily regulate synapse formation, maintenance, and function. Trends Neurosci. 2017, 40, 295–308. [Google Scholar] [CrossRef] [PubMed]
- Lutz, D.; Kataria, H.; Kleene, R.; Loers, G.; Chaudhary, H.; Guseva, D.; Wu, B.; Jakovcevski, I.; Schachner, M. Myelin basic protein cleaves cell adhesion molecule L1 and improves regeneration after injury. Mol. Neurobiol. 2016, 53, 3360–3376. [Google Scholar] [CrossRef]
- Fransen, E.; Lemmon, V.; Van Camp, G.; Vits, L.; Coucke, P.; Willems, P.J. CRASH syndrome: Clinical spectrum of corpus callosum hypoplasia, retardation, adducted thumbs, spastic paraparesis and hydrocephalus due to mutations in one single gene, L1. Eur. J. Hum. Genet. 1995, 3, 273–284. [Google Scholar] [CrossRef]
- Fransen, E.; Vits, L.; Van Camp, G.; Willems, P.J. The clinical spectrum of mutations in L1, a neuronal cell adhesion molecule. Am. J. Med. Genet. 1996, 64, 73–77. [Google Scholar] [CrossRef]
- Hortsch, M.; Nagaraj, K.; Mualla, R. The L1 family of cell adhesion molecules: A sickening number of mutations and protein functions. Adv. Neurobiol. 2014, 8, 195–229. [Google Scholar]
- Zhang, L. CRASH syndrome: Does it teach us about neurotrophic functions of cell adhesion molecules? Neuroscientist 2010, 16, 470–474. [Google Scholar] [CrossRef]
- Dahme, M.; Bartsch, U.; Martini, R.; Anliker, B.; Schachner, M.; Mantei, N. Disruption of the mouse L1 gene leads to malformations of the nervous system. Nat. Genet. 1997, 17, 346–349. [Google Scholar] [CrossRef]
- Fransen, E.; D’Hooge, R.; Van Camp, G.; Verhoye, M.; Sijbers, J.; Reyniers, E.; Soriano, P.; Kamiguchi, H.; Willemsen, R.; Koekkoek, S.K.; et al. L1 knockout mice show dilated ventricles, vermis hypoplasia and impaired exploration patterns. Hum. Mol. Genet. 1998, 7, 999–1009. [Google Scholar] [CrossRef]
- Katidou, M.; Vidaki, M.; Strigini, M.; Karagogeos, D. The immunoglobulin superfamily of neuronal cell adhesion molecules: Lessons from animal models and correlation with human disease. Biotechnol. J. 2008, 3, 1564–1580. [Google Scholar] [CrossRef]
- Wei, C.H.; Ryu, S.E. Homophilic interaction of the L1 family of cell adhesion molecules. Exp. Mol. Med. 2012, 44, 413–423. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kleene, R.; Lutz, D.; Loers, G.; Bork, U.; Borgmeyer, U.; Hermans-Borgmeyer, I.; Schachner, M. Revisiting the proteolytic processing of cell adhesion molecule L1. J. Neurochem. 2021, 157, 1102–1117. [Google Scholar] [CrossRef]
- Appel, F.; Holm, J.; Conscience, J.F.; Schachner, M. Several extracellular domains of the neural cell adhesion molecule L1 are involved in neurite outgrowth and cell body adhesion. J. Neurosci. 1993, 13, 4764–4775. [Google Scholar] [CrossRef] [Green Version]
- Holm, J.; Appel, F.; Schachner, M. Several extracellular domains of the neural cell adhesion molecule L1 are involved in homophilic interactions. J. Neurosci. Res. 1995, 42, 9–20. [Google Scholar] [CrossRef]
- Kraus, K.; Kleene, R.; Braren, I.; Loers, G.; Lutz, D.; Schachner, M. A fragment of adhesion molecule L1 is imported into mitochondria, and regulates mitochondrial metabolism and trafficking. J. Cell Sci. 2018, 131, jcs210500. [Google Scholar] [CrossRef] [Green Version]
- Kraus, K.; Kleene, R.; Henis, M.; Braren, I.; Kataria, H.; Sharaf, A.; Loers, G.; Schachner, M.; Lutz, D. A fragment of adhesion molecule L1 binds to nuclear receptors to regulate synaptic plasticity and motor coordination. Mol. Neurobiol. 2018, 55, 7164–7178. [Google Scholar] [CrossRef]
- Lutz, D.; Loers, G.; Kleene, R.; Oezen, I.; Kataria, H.; Katagihallimath, N.; Braren, I.; Harauz, G.; Schachner, M. Myelin basic protein cleaves cell adhesion molecule L1 and promotes neuritogenesis and cell survival. J. Biol. Chem. 2014, 289, 13503–13518. [Google Scholar] [CrossRef] [Green Version]
- Lutz, D.; Sharaf, A.; Drexler, D.; Kataria, H.; Wolters-Eisfeld, G.; Brunne, B.; Kleene, R.; Loers, G.; Frotscher, M.; Schachner, M. Proteolytic cleavage of transmembrane cell adhesion molecule L1 by extracellular matrix molecule Reelin is important for mouse brain development. Sci. Rep. 2017, 7, 15268. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lutz, D.; Wolters-Eisfeld, G.; Joshi, G.; Djogo, N.; Jakovcevski, I.; Schachner, M.; Kleene, R. Generation and nuclear translocation of sumoylated transmembrane fragment of cell adhesion molecule L1. J. Biol. Chem. 2012, 287, 17161–17175. [Google Scholar] [CrossRef] [PubMed]
- Lutz, D.; Wolters-Eisfeld, G.; Schachner, M.; Kleene, R. Cathepsin E generates a sumoylated intracellular fragment of the cell adhesion molecule L1 to promote neuronal and Schwann cell migration as well as myelination. J. Neurochem. 2014, 128, 713–724. [Google Scholar] [CrossRef] [PubMed]
- Kalus, I.; Schnegelsberg, B.; Seidah, N.G.; Kleene, R.; Schachner, M. The proprotein convertase PC5A and a metalloprotease are involved in the proteolytic processing of the neural adhesion molecule L1. J. Biol. Chem. 2003, 278, 10381–10388. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mechtersheimer, S.; Gutwein, P.; Agmon-Levin, N.; Stoeck, A.; Oleszewski, M.; Riedle, S.; Postina, R.; Fahrenholz, F.; Fogel, M.; Lemmon, V.; et al. Ectodomain shedding of L1 adhesion molecule promotes cell migration by autocrine binding to integrins. J. Cell Biol. 2001, 155, 661–673. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Congiu, L.; Granato, V.; Loers, G.; Kleene, R.; Schachner, M. Mitochondrial and neuronal dysfunctions in L1 mutant mice. Int. J. Mol. Sci. 2022, 23, 4337. [Google Scholar] [CrossRef]
- Kleene, R.; Loers, G.; Castillo, G.; Schachner, M. Cell adhesion molecule L1 interacts with the chromo shadow domain of heterochromatin protein 1 isoforms alpha, beta, and via its intracellular domain. FASEB J. 2022, 36, e22074. [Google Scholar] [CrossRef]
- Loers, G.; Kleene, R.; Girbes Minguez, M.; Schachner, M. The cell adhesion molecule L1 interacts with methyl CpG binding protein 2 via its intracellular domain. Int. J. Mol. Sci. 2022, 23, 3554. [Google Scholar] [CrossRef]
- Girbes Minguez, M.; Wolters-Eisfeld, G.; Lutz, D.; Buck, F.; Schachner, M.; Kleene, R. The cell adhesion molecule L1 interacts with nuclear proteins via its intracellular domain. FASEB J. 2020, 34, 9869–9883. [Google Scholar] [CrossRef]
- Hu, J.; Lin, S.L.; Schachner, M. A fragment of cell adhesion molecule L1 reduces amyloid-beta plaques in a mouse model of Alzheimer’s disease. Cell Death Dis. 2022, 13, 48. [Google Scholar] [CrossRef]
- Leshchyns’ka, I.; Sytnyk, V.; Richter, M.; Andreyeva, A.; Puchkov, D.; Schachner, M. The adhesion molecule CHL1 regulates uncoating of clathrin-coated synaptic vesicles. Neuron 2006, 52, 1011–1025. [Google Scholar] [CrossRef] [Green Version]
- Appel, F.; Holm, J.; Conscience, J.F.; von Bohlen und Halbach, F.; Faissner, A.; James, P.; Schachner, M. Identification of the border between fibronectin type III homologous repeats 2 and 3 of the neural cell adhesion molecule L1 as a neurite outgrowth promoting and signal transducing domain. J. Neurobiol. 1995, 28, 297–312. [Google Scholar] [CrossRef]
- Kataria, H.; Lutz, D.; Chaudhary, H.; Schachner, M.; Loers, G. Small molecule agonists of cell adhesion molecule L1 mimic L1 functions in vivo. Mol. Neurobiol. 2016, 53, 4461–4483. [Google Scholar]
- Daujat, S.; Zeissler, U.; Waldmann, T.; Happel, N.; Schneider, R. HP1 binds specifically to Lys26-methylated histone H1.4, whereas simultaneous Ser27 phosphorylation blocks HP1 binding. J. Biol. Chem. 2005, 280, 38090–38095. [Google Scholar] [CrossRef] [Green Version]
- Ghosh, R.P.; Horowitz-Scherer, R.A.; Nikitina, T.; Shlyakhtenko, L.S.; Woodcock, C.L. MeCP2 binds cooperatively to its substrate and competes with histone H1 for chromatin binding sites. Mol. Cell Biol. 2010, 30, 4656–4670. [Google Scholar] [CrossRef] [Green Version]
- Fukuda, M.; Mikoshiba, K. Expression of synaptotagmin I or II promotes neurite outgrowth in PC12 cells. Neurosci. Lett. 2000, 295, 33–36. [Google Scholar] [CrossRef]
- Kabayama, H.; Takei, K.; Fukuda, M.; Ibata, K.; Mikoshiba, K. Functional involvement of synaptotagmin I/II C2A domain in neurite outgrowth of chick dorsal root ganglion neuron. Neuroscience 1999, 88, 999–1003. [Google Scholar] [CrossRef]
- Mikoshiba, K.; Fukuda, M.; Ibata, K.; Kabayama, H.; Mizutani, A. Role of synaptotagmin, a Ca2+ and inositol polyphosphate binding protein, in neurotransmitter release and neurite outgrowth. Chem. Phys. Lipids 1999, 98, 59–67. [Google Scholar] [CrossRef]
- Baker, K.; Gordon, S.L.; Melland, H.; Bumbak, F.; Scott, D.J.; Jiang, T.J.; Owen, D.; Turner, B.J.; Boyd, S.G.; Rossi, M.; et al. SYT1-associated neurodevelopmental disorder: A case series. Brain 2018, 141, 2576–2591. [Google Scholar] [CrossRef] [Green Version]
- Chen, T.; Wu, Q.; Zhang, Y.; Zhang, D. NDUFV2 regulates neuronal migration in the developing cerebral cortex through modulation of the multipolar-bipolar transition. Brain Res. 2015, 1625, 102–110. [Google Scholar] [CrossRef]
- Nakagawa, T.; Xiong, Y. X-linked mental retardation gene CUL4B targets ubiquitylation of H3K4 methyltransferase component WDR5 and regulates neuronal gene expression. Mol. Cell 2011, 43, 381–391. [Google Scholar] [CrossRef] [Green Version]
- Tremblay, M.W.; Green, M.V.; Goldstein, B.M.; Aldridge, A.I.; Rosenfeld, J.A.; Streff, H.; Tan, W.D.; Craigen, W.; Bekheirnia, N.; Al Tala, S.; et al. Mutations of the histone linker H1-4 in neurodevelopmental disorders and functional characterization of neurons expressing C-terminus frameshift mutant H1.4. Hum. Mol. Genet. 2022, 31, 1430–1442. [Google Scholar] [CrossRef] [PubMed]
- Takeuchi, A.; Iida, K.; Tsubota, T.; Hosokawa, M.; Denawa, M.; Brown, J.B.; Ninomiya, K.; Ito, M.; Kimura, H.; Abe, T.; et al. Loss of Sfpq causes long-gene transcriptopathy in the brain. Cell Rep. 2018, 23, 1326–1341. [Google Scholar] [CrossRef] [PubMed]
- Cosker, K.E.; Fenstermacher, S.J.; Pazyra-Murphy, M.F.; Elliott, H.L.; Segal, R.A. The RNA-binding protein SFPQ orchestrates an RNA regulon to promote axon viability. Nat. Neurosci. 2016, 19, 690–696. [Google Scholar] [CrossRef] [PubMed]
- Mircsof, D.; Langouet, M.; Rio, M.; Moutton, S.; Siquier-Pernet, K.; Bole-Feysot, C.; Cagnard, N.; Nitschke, P.; Gaspar, L.; Znidaric, M.; et al. Mutations in NONO lead to syndromic intellectual disability and inhibitory synaptic defects. Nat. Neurosci. 2015, 18, 1731–1736. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheng, L.; Wang, Y.; Xiang, L.; Qi, J. Heat shock cognate 70 kDa protein is the target of tetradecyl 2,3-dihydroxybenzoate for neuritogenic effect in PC12 Cells. Biomedicines 2021, 9, 1483. [Google Scholar] [CrossRef]
- Kilkenny, C.; Browne, W.; Cuthill, I.C.; Emerson, M.; Altman, D.G. Animal research: Reporting in vivo experiments: The ARRIVE guidelines. J. Gene Med. 2010, 12, 561–563. [Google Scholar] [CrossRef]
- Kleene, R.; Cassens, C.; Bahring, R.; Theis, T.; Xiao, M.F.; Dityatev, A.; Schafer-Nielsen, C.; Doring, F.; Wischmeyer, E.; Schachner, M. Functional consequences of the interactions among the neural cell adhesion molecule NCAM, the receptor tyrosine kinase TrkB, and the inwardly rectifying K+ channel KIR3.3. J. Biol. Chem. 2010, 285, 28968–28979. [Google Scholar] [CrossRef] [Green Version]
- Andreyeva, A.; Leshchyns’ka, I.; Knepper, M.; Betzel, C.; Redecke, L.; Sytnyk, V.; Schachner, M. CHL1 is a selective organizer of the presynaptic machinery chaperoning the SNARE complex. PLoS ONE 2010, 5, e12018. [Google Scholar] [CrossRef] [Green Version]
- Xiao, M.F.; Xu, J.C.; Tereshchenko, Y.; Novak, D.; Schachner, M.; Kleene, R. Neural cell adhesion molecule modulates dopaminergic signaling and behavior by regulating dopamine D2 receptor internalization. J. Neurosci. 2009, 29, 14752–14763. [Google Scholar] [CrossRef] [Green Version]
- Schwarze, S.R.; Dowdy, S.F. In vivo protein transduction: Intracellular delivery of biologically active proteins, compounds and DNA. Trends Pharmacol. Sci. 2000, 21, 45–48. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| AR | androgen receptor |
| ERα | estrogen receptor α |
| ERβ | estrogen receptor β |
| HistH1e | histone H1.4 |
| hnRNP A1 | heterogeneous nuclear ribonucleoprotein A1 |
| hnRNP A2/B1 | heterogeneous nuclear ribonucleoprotein A2/B1 |
| hnRNP A3 | heterogeneous nuclear ribonucleoprotein A3 |
| impβ1 | importin β1 |
| MeCP2 | methyl CpG binding protein 2 |
| NDUFV2 | NADH dehydrogenase ubiquinone flavoprotein 2 |
| NonO | non-POU domain containing octamer-binding protein |
| Nup93 | nucleoporin 93 kDa |
| PPARγ | peroxisome proliferator-activated receptor γ |
| PSPC1 | paraspeckle component 1 |
| RXRβ | retinoid X receptor β |
| SFPQ | splicing factor proline/glutamine-rich |
| SYT1 | synaptotagmin 1 |
| TOP1 | DNA topoisomerase I |
| VDR | vitamin D receptor |
| WDR5 | WD-repeat protein 5 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kleene, R.; Loers, G.; Schachner, M. The KDET Motif in the Intracellular Domain of the Cell Adhesion Molecule L1 Interacts with Several Nuclear, Cytoplasmic, and Mitochondrial Proteins Essential for Neuronal Functions. Int. J. Mol. Sci. 2023, 24, 932. https://doi.org/10.3390/ijms24020932
Kleene R, Loers G, Schachner M. The KDET Motif in the Intracellular Domain of the Cell Adhesion Molecule L1 Interacts with Several Nuclear, Cytoplasmic, and Mitochondrial Proteins Essential for Neuronal Functions. International Journal of Molecular Sciences. 2023; 24(2):932. https://doi.org/10.3390/ijms24020932
Chicago/Turabian StyleKleene, Ralf, Gabriele Loers, and Melitta Schachner. 2023. "The KDET Motif in the Intracellular Domain of the Cell Adhesion Molecule L1 Interacts with Several Nuclear, Cytoplasmic, and Mitochondrial Proteins Essential for Neuronal Functions" International Journal of Molecular Sciences 24, no. 2: 932. https://doi.org/10.3390/ijms24020932