
Molecular Mechanism and Agricultural Application of the NifA–NifL System for Nitrogen Fixation

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Nitrogenase and Its Transcriptional Regulation
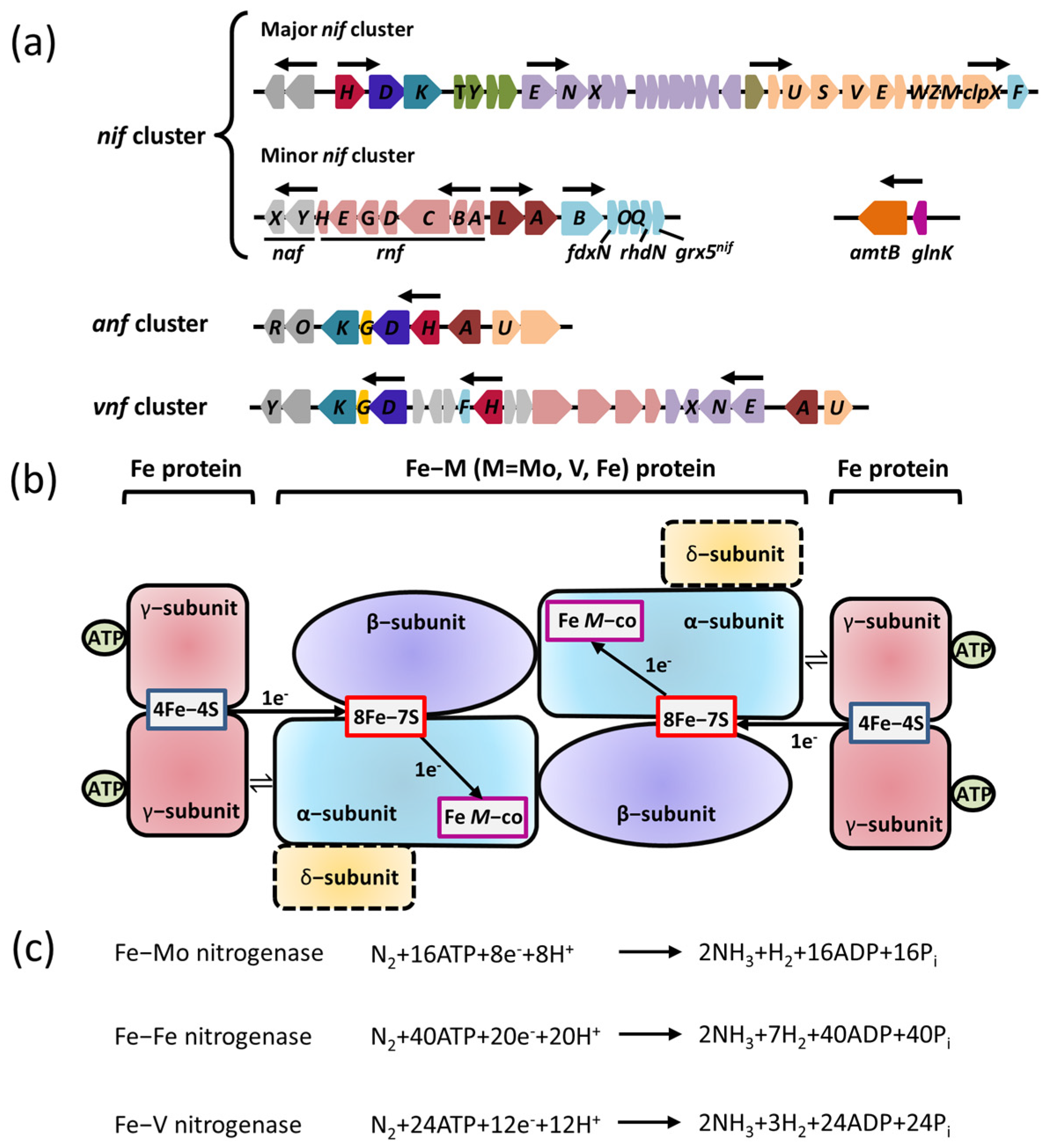
2.1. Nitrogenase
2.2. Transcriptional Regulation of Nitrogenase
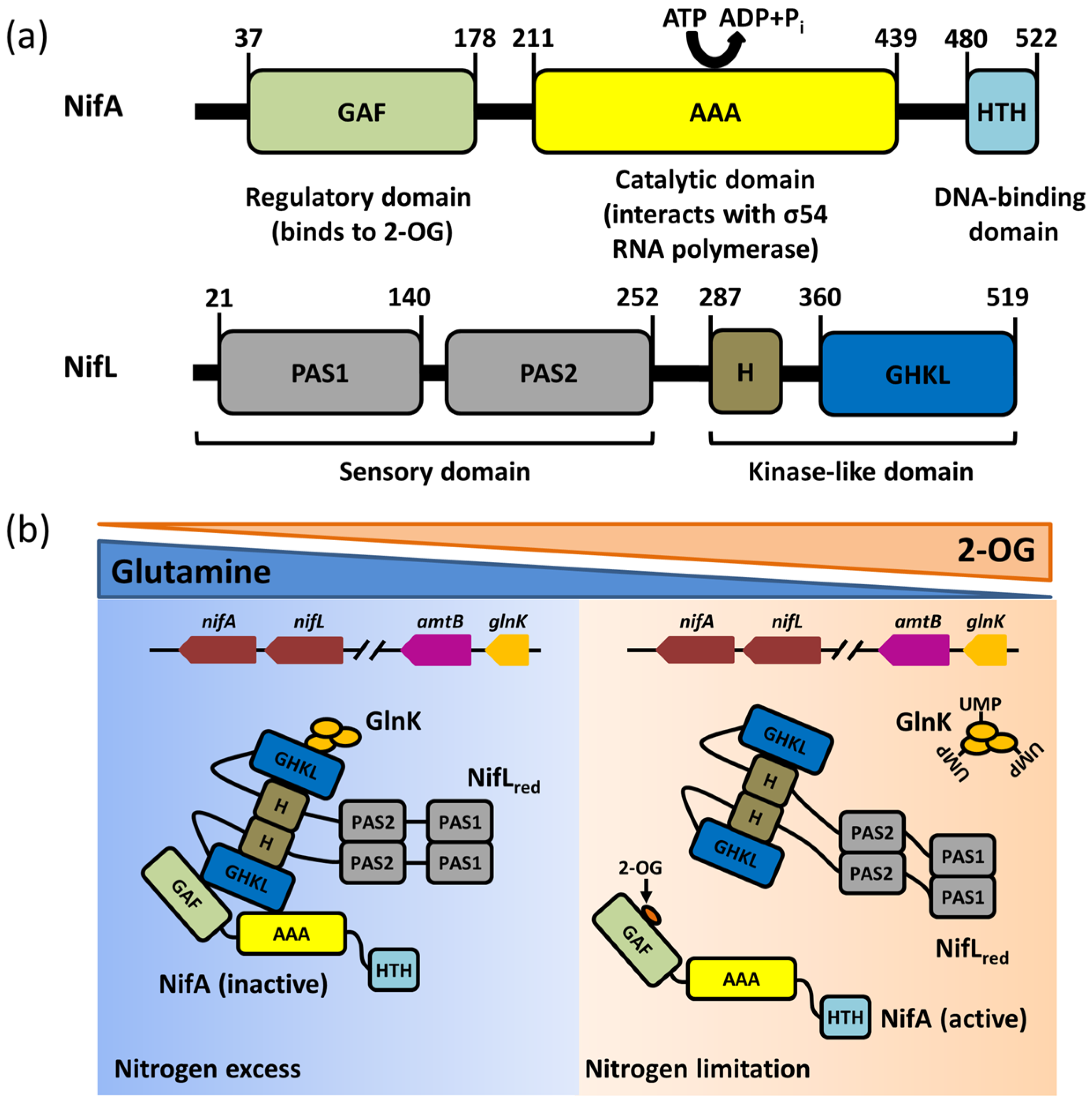
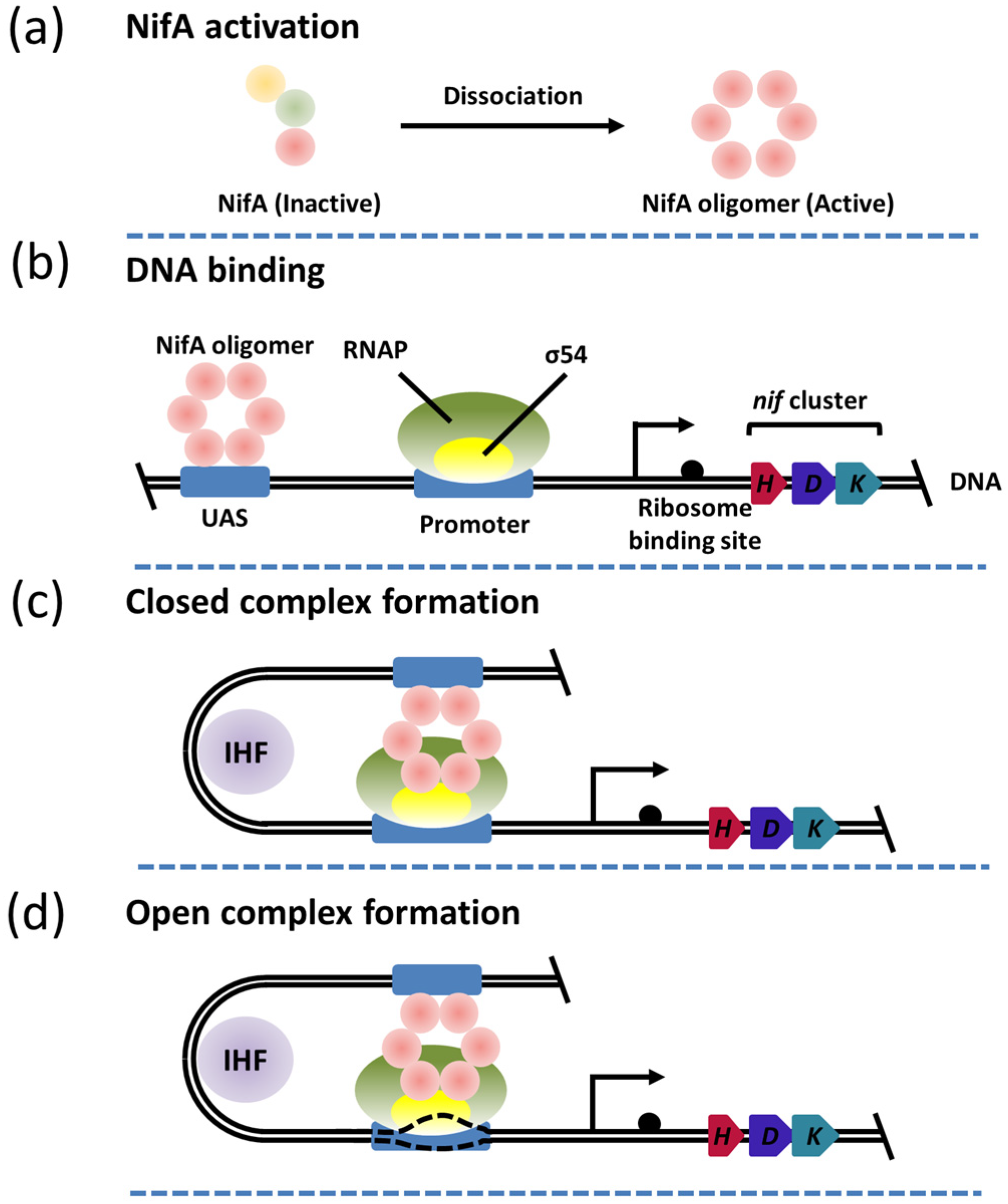
3. The Function and Structure of NifA
4. Structural Characteristics and Functions of the NifL Protein
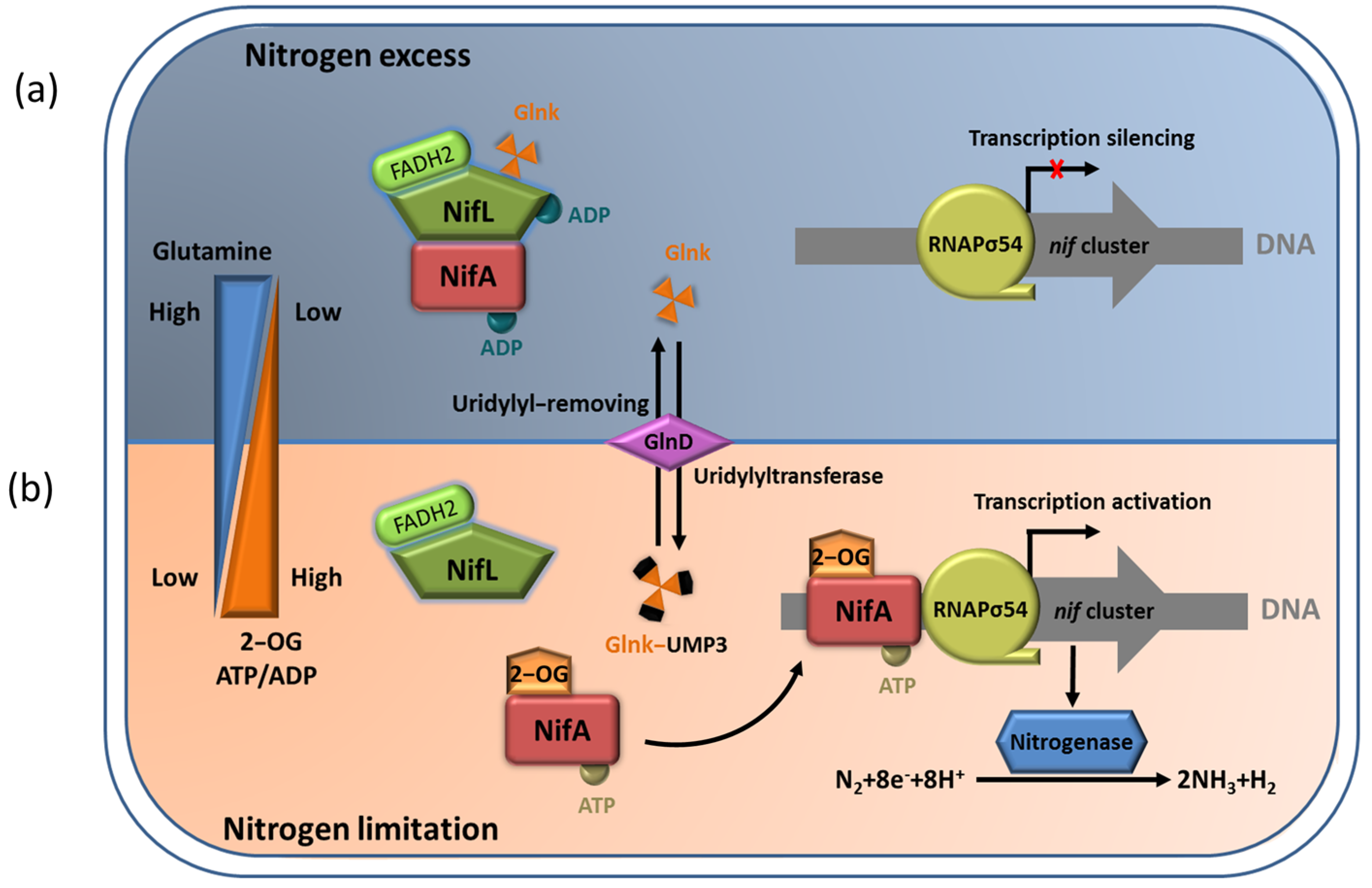
5. The NifL–NifA System Responds to the Transcriptional Regulation of Nitrogenase via Environmental Signaling Molecules
5.1. Regulation of NifA Function by 2–OG
5.2. Effects of ADP and FAD Molecules on NifL Function
5.3. The NifL–NifA System Regulated by GlnK
6. The Differences in the NifA–NifL System for the Different Nitrogen–Fixing Bacteria
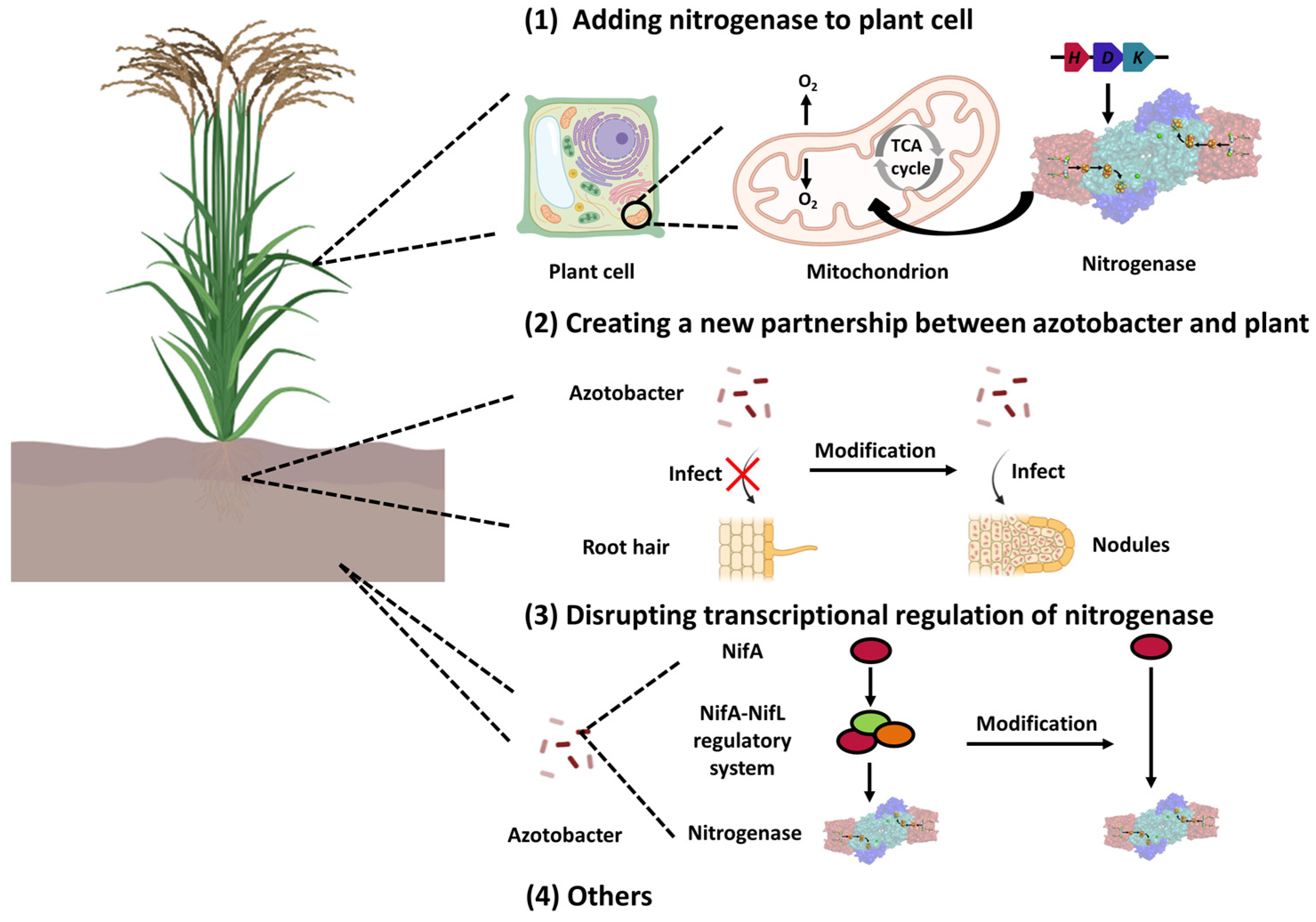
7. The Potential Applications of the NifA–NifL System in Agricultural Development
8. Summary and Prospect
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Stokstad, E. The nitrogen fix. Science 2016, 353, 1225–1227. [Google Scholar] [CrossRef]
- Fan, K.; Delgado-Baquerizo, M.; Guo, X.; Wang, D.; Wu, Y.; Zhu, M.; Yu, W.; Yao, H.; Zhu, Y.G.; Chu, H. Suppressed N fixation and diazotrophs after four decades of fertilization. Microbiome 2019, 7, 143. [Google Scholar] [CrossRef] [PubMed]
- Bonilla Cedrez, C.; Chamberlin, J.; Guo, Z.; Hijmans, R.J. Spatial variation in fertilizer prices in Sub-Saharan Africa. PLoS ONE 2020, 15, e0227764. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ye, Y.; Ngo, H.H.; Guo, W.; Chang, S.W.; Nguyen, D.D.; Varjani, S.; Ding, A.; Bui, X.T.; Nguyen, D.P. Bio-membrane based integrated systems for nitrogen recovery in wastewater treatment: Current applications and future perspectives. Chemosphere 2021, 265, 129076. [Google Scholar] [CrossRef] [PubMed]
- Kumar, M.; Tomar, R.S.; Lade, H.; Paul, D. Methylotrophic bacteria in sustainable agriculture. World J. Microbiol. Biotechnol. 2016, 32, 120. [Google Scholar] [CrossRef]
- Canfield, D.E.; Glazer, A.N.; Falkowski, P.G. The evolution and future of Earth’s nitrogen cycle. Science 2010, 330, 192–196. [Google Scholar] [CrossRef] [Green Version]
- Hu, Y.; Ribbe, M.W. Biosynthesis of the Metalloclusters of Nitrogenases. Annu. Rev. Biochem. 2016, 85, 455–483. [Google Scholar] [CrossRef]
- Das, H.K. Azotobacters as biofertilizer. Adv. Appl. Microbiol. 2019, 108, 1–43. [Google Scholar] [CrossRef]
- Romero-Perdomo, F.; Abril, J.; Camelo, M.; Moreno-Galvan, A.; Pastrana, I.; Rojas-Tapias, D.; Bonilla, R. Azotobacter chroococcum as a potentially useful bacterial biofertilizer for cotton (Gossypium hirsutum): Effect in reducing N fertilization. Rev. Argent. Microbiol. 2017, 49, 377–383. [Google Scholar] [CrossRef]
- Zambrano-Mendoza, J.L.; Sangoquiza-Caiza, C.A.; Campaa-Cruz, D.F.; Yánez-Guzmán, C.F. Use of Biofertilizers in Agricultural Production. In Technology in Agriculture; IntechOpen: London, UK, 2021; ISBN 978-1-83881-921-7. [Google Scholar]
- Hakeem, K.R.; Akhtar, J.; Sabir, M.J.S.I.P. Azotobacter chroococcum—A Potential Biofertilizer in Agriculture: An Overview. In Soil Science: Agricultural and Environmental Prospectives; Springer: Cham, Switzerland, 2016; pp. 333–348. [Google Scholar]
- Bueno Batista, M.; Dixon, R. Manipulating nitrogen regulation in diazotrophic bacteria for agronomic benefit. Biochem. Soc. Trans. 2019, 47, 603–614. [Google Scholar] [CrossRef]
- Wilson, P.W.; Burris, R.H.; Lind, C.J. The Dissociation Constant in Nitrogen Fixation by Azotobacter. Proc. Natl. Acad. Sci. USA 1942, 28, 243–250. [Google Scholar] [CrossRef] [Green Version]
- Seefeldt, L.C.; Hoffman, B.M.; Dean, D.R. Mechanism of Mo-dependent nitrogenase. Annu. Rev. Biochem. 2009, 78, 701–722. [Google Scholar] [CrossRef]
- Harwood, C.S. Iron-Only and Vanadium Nitrogenases: Fail-Safe Enzymes or Something More? Annu. Rev. Microbiol. 2020, 74, 247–266. [Google Scholar] [CrossRef]
- Rutledge, H.L.; Cook, B.D.; Nguyen, H.P.M.; Herzik, M.A., Jr.; Tezcan, F.A. Structures of the nitrogenase complex prepared under catalytic turnover conditions. Science 2022, 377, 865–869. [Google Scholar] [CrossRef]
- Seefeldt, L.C.; Yang, Z.Y.; Duval, S.; Dean, D.R. Nitrogenase reduction of carbon-containing compounds. Biochim. Biophys. Acta 2013, 1827, 1102–1111. [Google Scholar] [CrossRef] [Green Version]
- Rubio, L.M.; Ludden, P.W. Biosynthesis of the iron-molybdenum cofactor of nitrogenase. Annu. Rev. Microbiol. 2008, 62, 93–111. [Google Scholar] [CrossRef] [Green Version]
- Boyd, E.S.; Hamilton, T.L.; Peters, J.W. An alternative path for the evolution of biological nitrogen fixation. Front. Microbiol. 2011, 2, 205. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.; Zhang, L.; Liu, Z.; Zhao, D.; Liu, X.; Zhang, B.; Xie, J.; Hong, Y.; Li, P.; Chen, S.; et al. A minimal nitrogen fixation gene cluster from Paenibacillus sp. WLY78 enables expression of active nitrogenase in Escherichia coli. PLoS Genet. 2013, 9, e1003865. [Google Scholar] [CrossRef]
- Curatti, L.; Brown, C.S.; Ludden, P.W.; Rubio, L.M. Genes required for rapid expression of nitrogenase activity in Azotobacter vinelandii. Proc. Natl. Acad. Sci. USA 2005, 102, 6291–6296. [Google Scholar] [CrossRef] [Green Version]
- Arnold, W.; Rump, A.; Klipp, W.; Priefer, U.B.; Puhler, A. Nucleotide sequence of a 24,206-base-pair DNA fragment carrying the entire nitrogen fixation gene cluster of Klebsiella pneumoniae. J. Mol. Biol. 1988, 203, 715–738. [Google Scholar] [CrossRef] [PubMed]
- Glyde, R.; Ye, F.; Darbari, V.C.; Zhang, N.; Buck, M.; Zhang, X. Structures of RNA Polymerase Closed and Intermediate Complexes Reveal Mechanisms of DNA Opening and Transcription Initiation. Mol. Cell. 2017, 67, 106–116 e104. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Levdikov, V.M.; Blagova, E.; Colledge, V.L.; Lebedev, A.A.; Williamson, D.C.; Sonenshein, A.L.; Wilkinson, A.J. Structural rearrangement accompanying ligand binding in the GAF domain of CodY from Bacillus subtilis. J. Mol. Biol. 2009, 390, 1007–1018. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sysoeva, T.A.; Chowdhury, S.; Guo, L.; Nixon, B.T. Nucleotide-induced asymmetry within ATPase activator ring drives sigma54-RNAP interaction and ATP hydrolysis. Genes. Dev. 2013, 27, 2500–2511. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gao, F.; Danson, A.E.; Ye, F.; Jovanovic, M.; Buck, M.; Zhang, X. Bacterial Enhancer Binding Proteins-AAA(+) Proteins in Transcription Activation. Biomolecules 2020, 10, 351. [Google Scholar] [CrossRef] [Green Version]
- Batchelor, J.D.; Lee, P.S.; Wang, A.C.; Doucleff, M.; Wemmer, D.E. Structural mechanism of GAF-regulated sigma(54) activators from Aquifex aeolicus. J. Mol. Biol. 2013, 425, 156–170. [Google Scholar] [CrossRef] [Green Version]
- Anantharaman, V.; Balaji, S.; Aravind, L. The signaling helix: A common functional theme in diverse signaling proteins. Biol. Direct. 2006, 1, 25. [Google Scholar] [CrossRef] [Green Version]
- Little, R.; Martinez-Argudo, I.; Dixon, R. Role of the central region of NifL in conformational switches that regulate nitrogen fixation. Biochem. Soc. Trans. 2006, 34, 162–164. [Google Scholar] [CrossRef]
- Austin, S.; Buck, M.; Cannon, W.; Eydmann, T.; Dixon, R. Purification and in vitro activities of the native nitrogen fixation control proteins NifA and NifL. J. Bacteriol. 1994, 176, 3460–3465. [Google Scholar] [CrossRef] [Green Version]
- Martinez-Argudo, I.; Little, R.; Shearer, N.; Johnson, P.; Dixon, R. The NifL-NifA System: A multidomain transcriptional regulatory complex that integrates environmental signals. J. Bacteriol. 2004, 186, 601–610. [Google Scholar] [CrossRef] [Green Version]
- Rojas-Pirela, M.; Rigden, D.J.; Michels, P.A.; Caceres, A.J.; Concepcion, J.L.; Quinones, W. Structure and function of Per-ARNT-Sim domains and their possible role in the life-cycle biology of Trypanosoma cruzi. Mol. Biochem. Parasitol. 2018, 219, 52–66. [Google Scholar] [CrossRef]
- Henry, J.T.; Crosson, S. Ligand-binding PAS domains in a genomic, cellular, and structural context. Annu. Rev. Microbiol. 2011, 65, 261–286. [Google Scholar] [CrossRef] [Green Version]
- Vogt, J.H.; Schippers, J.H. Setting the PAS, the role of circadian PAS domain proteins during environmental adaptation in plants. Front. Plant Sci. 2015, 6, 513. [Google Scholar] [CrossRef] [Green Version]
- Little, R.; Slavny, P.; Dixon, R. Influence of PAS domain flanking regions on oligomerisation and redox signalling by NifL. PLoS ONE 2012, 7, e46651. [Google Scholar] [CrossRef]
- Martinez-Argudo, I.; Little, R.; Dixon, R. Role of the amino-terminal GAF domain of the NifA activator in controlling the response to the antiactivator protein NifL. Mol. Microbiol. 2004, 52, 1731–1744. [Google Scholar] [CrossRef]
- Barrett, J.; Ray, P.; Sobczyk, A.; Little, R.; Dixon, R. Concerted inhibition of the transcriptional activation functions of the enhancer-binding protein NIFA by the anti-activator NIFL. Mol. Microbiol. 2001, 39, 480–493. [Google Scholar] [CrossRef] [Green Version]
- Martinez-Argudo, I.; Little, R.; Dixon, R. A crucial arginine residue is required for a conformational switch in NifL to regulate nitrogen fixation in Azotobacter vinelandii. Proc. Natl. Acad. Sci. USA 2004, 101, 16316–16321. [Google Scholar] [CrossRef] [Green Version]
- Reyes-Ramirez, F.; Little, R.; Dixon, R. Mutant forms of the Azotobacter vinelandii transcriptional activator NifA resistant to inhibition by the NifL regulatory protein. J. Bacteriol. 2002, 184, 6777–6785. [Google Scholar] [CrossRef] [Green Version]
- Reyes-Ramirez, F.; Little, R.; Dixon, R. Role of Escherichia coli nitrogen regulatory genes in the nitrogen response of the Azotobacter vinelandii NifL-NifA complex. J. Bacteriol. 2001, 183, 3076–3082. [Google Scholar] [CrossRef] [Green Version]
- Huergo, L.F.; Dixon, R. The Emergence of 2-Oxoglutarate as a Master Regulator Metabolite. Microbiol. Mol. Biol. Rev. 2015, 79, 419–435. [Google Scholar] [CrossRef] [Green Version]
- Senior, P.J. Regulation of nitrogen metabolism in Escherichia coli and Klebsiella aerogenes: Studies with the continuous-culture technique. J. Bacteriol. 1975, 123, 407–418. [Google Scholar] [CrossRef] [PubMed]
- Little, R.; Dixon, R. The amino-terminal GAF domain of Azotobacter vinelandii NifA binds 2-oxoglutarate to resist inhibition by NifL under nitrogen-limiting conditions. J. Biol. Chem. 2003, 278, 28711–28718. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Little, R.; Colombo, V.; Leech, A.; Dixon, R. Direct interaction of the NifL regulatory protein with the GlnK signal transducer enables the Azotobacter vinelandii NifL-NifA regulatory system to respond to conditions replete for nitrogen. J. Biol. Chem. 2002, 277, 15472–15481. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hill, S.; Austin, S.; Eydmann, T.; Jones, T.; Dixon, R. Azotobacter vinelandii NIFL is a flavoprotein that modulates transcriptional activation of nitrogen-fixation genes via a redox-sensitive switch. Proc. Natl. Acad. Sci. USA 1996, 93, 2143–2148. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hill, S.; Kennedy, C.; Kavanagh, E.; Goldberg, R.B.; Hanau, R. Nitrogen fixation gene (nifL) involved in oxygen regulation of nitrogenase synthesis in K. pneumoniae. Nature 1981, 290, 424–426. [Google Scholar] [CrossRef]
- Key, J.; Hefti, M.; Purcell, E.B.; Moffat, K. Structure of the redox sensor domain of Azotobacter vinelandii NifL at atomic resolution: Signaling, dimerization, and mechanism. Biochemistry 2007, 46, 3614–3623. [Google Scholar] [CrossRef]
- Little, R.; Salinas, P.; Slavny, P.; Clarke, T.A.; Dixon, R. Substitutions in the redox-sensing PAS domain of the NifL regulatory protein define an inter-subunit pathway for redox signal transmission. Mol. Microbiol. 2011, 82, 222–235. [Google Scholar] [CrossRef] [Green Version]
- Little, R.; Reyes-Ramirez, F.; Zhang, Y.; van Heeswijk, W.C.; Dixon, R. Signal transduction to the Azotobacter vinelandii NIFL-NIFA regulatory system is influenced directly by interaction with 2-oxoglutarate and the PII regulatory protein. EMBO J. 2000, 19, 6041–6050. [Google Scholar] [CrossRef] [Green Version]
- He, S.; Chen, M.; Xie, Z.; Yan, Y.; Li, H.; Fan, Y.; Ping, S.; Lin, M.; Elmerich, C. Involvement of GlnK, a PII protein, in control of nitrogen fixation and ammonia assimilation in Pseudomonas stutzeri A1501. Arch Microbiol. 2008, 190, 1–10. [Google Scholar] [CrossRef]
- Rudnick, P.; Kunz, C.; Gunatilaka, M.K.; Hines, E.R.; Kennedy, C. Role of GlnK in NifL-mediated regulation of NifA activity in Azotobacter vinelandii. J. Bacteriol. 2002, 184, 812–820. [Google Scholar] [CrossRef] [Green Version]
- Oliveira, M.A.; Aquino, B.; Bonatto, A.C.; Huergo, L.F.; Chubatsu, L.S.; Pedrosa, F.O.; Souza, E.M.; Dixon, R.; Monteiro, R.A. Interaction of GlnK with the GAF domain of Herbaspirillum seropedicae NifA mediates NH(4)(+)-regulation. Biochimie 2012, 94, 1041–1047. [Google Scholar] [CrossRef]
- Meletzus, D.; Rudnick, P.; Doetsch, N.; Green, A.; Kennedy, C. Characterization of the glnK-amtB operon of Azotobacter vinelandii. J. Bacteriol. 1998, 180, 3260–3264. [Google Scholar] [CrossRef] [Green Version]
- Oliveira, M.A.; Gerhardt, E.C.; Huergo, L.F.; Souza, E.M.; Pedrosa, F.O.; Chubatsu, L.S. 2-Oxoglutarate levels control adenosine nucleotide binding by Herbaspirillum seropedicae PII proteins. FEBS J. 2015, 282, 4797–4809. [Google Scholar] [CrossRef] [PubMed]
- Perry, S.; Shearer, N.; Little, R.; Dixon, R. Mutational analysis of the nucleotide-binding domain of the anti-activator NifL. J. Mol. Biol. 2005, 346, 935–949. [Google Scholar] [CrossRef]
- Xu, Y.; Cheah, E.; Carr, P.D.; van Heeswijk, W.C.; Westerhoff, H.V.; Vasudevan, S.G.; Ollis, D.L. GlnK, a PII-homologue: Structure reveals ATP binding site and indicates how the T-loops may be involved in molecular recognition. J. Mol. Biol. 1998, 282, 149–165. [Google Scholar] [CrossRef]
- Hesketh, A.; Fink, D.; Gust, B.; Rexer, H.U.; Scheel, B.; Chater, K.; Wohlleben, W.; Engels, A. The GlnD and GlnK homologues of Streptomyces coelicolor A3(2) are functionally dissimilar to their nitrogen regulatory system counterparts from enteric bacteria. Mol. Microbiol. 2002, 46, 319–330. [Google Scholar] [CrossRef]
- Jiang, P.; Ninfa, A.J. Alpha-ketoglutarate controls the ability of the Escherichia coli PII signal transduction protein to regulate the activities of NRII (NrB but does not control the binding of PII to NRII. Biochemistry 2009, 48, 11514–11521. [Google Scholar] [CrossRef] [Green Version]
- Jiang, P.; Ninfa, A.J. Sensation and signaling of alpha-ketoglutarate and adenylylate energy charge by the Escherichia coli PII signal transduction protein require cooperation of the three ligand-binding sites within the PII trimer. Biochemistry 2009, 48, 11522–11531. [Google Scholar] [CrossRef] [Green Version]
- Plunkett, M.H.; Knutson, C.M.; Barney, B.M. Key factors affecting ammonium production by an Azotobacter vinelandii strain deregulated for biological nitrogen fixation. Microb. Cell Fact. 2020, 19, 107. [Google Scholar] [CrossRef]
- Portugal, M.E.; Souza, E.M.; Pedrosa, F.O.; Benelli, E.M. Streptococcus mutans GlnK protein: An unusual PII family member. Braz. J. Med. Biol. Res. 2011, 44, 394–401. [Google Scholar] [CrossRef] [Green Version]
- Arcondeguy, T.; van Heeswijk, W.C.; Merrick, M. Studies on the roles of GlnK and GlnB in regulating Klebsiella pneumoniae NifL-dependent nitrogen control. FEMS Microbiol. Lett. 1999, 180, 263–270. [Google Scholar] [CrossRef] [PubMed]
- Gloer, J.; Thummer, R.; Ullrich, H.; Schmitz, R.A. Towards understanding the nitrogen signal transduction for nif gene expression in Klebsiella pneumoniae. FEBS J. 2008, 275, 6281–6294. [Google Scholar] [CrossRef] [PubMed]
- Stefanello, A.A.; Oliveira, M.A.S.; Souza, E.M.; Pedrosa, F.O.; Chubatsu, L.S.; Huergo, L.F.; Dixon, R.; Monteiro, R.A. Regulation of Herbaspirillum seropedicae NifA by the GlnK PII signal transduction protein is mediated by effectors binding to allosteric sites. Biochim. Biophys. Acta Proteins. Proteom. 2020, 1868, 140348. [Google Scholar] [CrossRef] [PubMed]
- Egener, T.; Sarkar, A.; Martin, D.E.; Reinhold-Hurek, B. Identification of a NifL-like protein in a diazotroph of the beta-subgroup of the Proteobacteria, Azoarcus sp. strain BH72. Microbiology (Reading) 2002, 148, 3203–3212. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fischer, H.M. Genetic regulation of nitrogen fixation in rhizobia. Microbiol. Rev. 1994, 58, 352–386. [Google Scholar] [CrossRef]
- Steenhoudt, O.; Vanderleyden, J. Azospirillum, a free-living nitrogen-fixing bacterium closely associated with grasses: Genetic, biochemical and ecological aspects. FEMS Microbiol. Rev. 2000, 24, 487–506. [Google Scholar] [CrossRef]
- Inaba, J.; Thornton, J.; Huergo, L.F.; Monteiro, R.A.; Klassen, G.; Pedrosa Fde, O.; Merrick, M.; de Souza, E.M. Mutational analysis of GlnB residues critical for NifA activation in Azospirillum brasilense. Microbiol. Res. 2015, 171, 65–72. [Google Scholar] [CrossRef]
- Sotomaior, P.; Araujo, L.M.; Nishikawa, C.Y.; Huergo, L.F.; Monteiro, R.A.; Pedrosa, F.O.; Chubatsu, L.S.; Souza, E.M. Effect of ATP and 2-oxoglutarate on the in vitro interaction between the NifA GAF domain and the GlnB protein of Azospirillum brasilense. Braz. J. Med. Biol. Res. 2012, 45, 1135–1140. [Google Scholar] [CrossRef] [Green Version]
- Masepohl, B.; Drepper, T.; Paschen, A.; Gross, S.; Pawlowski, A.; Raabe, K.; Riedel, K.U.; Klipp, W. Regulation of nitrogen fixation in the phototrophic purple bacterium Rhodobacter capsulatus. J. Mol. Microbiol. Biotechnol. 2002, 4, 243–248. [Google Scholar]
- Wang, X.; Bai, J.; Xie, T.; Wang, W.; Zhang, G.; Yin, S.; Wang, D. Effects of biological nitrification inhibitors on nitrogen use efficiency and greenhouse gas emissions in agricultural soils: A review. Ecotoxicol. Environ. Saf. 2021, 220, 112338. [Google Scholar] [CrossRef]
- Dixon, R.A.; Postgate, J.R. Genetic transfer of nitrogen fixation from Klebsiella pneumoniae to Escherichia coli. Nature 1972, 237, 102–103. [Google Scholar] [CrossRef] [PubMed]
- Xiang, N.; Guo, C.; Liu, J.; Xu, H.; Dixon, R.; Yang, J.; Wang, Y.P. Using synthetic biology to overcome barriers to stable expression of nitrogenase in eukaryotic organelles. Proc. Natl. Acad. Sci. USA 2020, 117, 16537–16545. [Google Scholar] [CrossRef] [PubMed]
- Ivleva, N.B.; Groat, J.; Staub, J.M.; Stephens, M. Expression of Active Subunit of Nitrogenase via Integration into Plant Organelle Genome. PLoS ONE 2016, 11, e0160951. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lopez-Torrejon, G.; Jimenez-Vicente, E.; Buesa, J.M.; Hernandez, J.A.; Verma, H.K.; Rubio, L.M. Expression of a functional oxygen-labile nitrogenase component in the mitochondrial matrix of aerobically grown yeast. Nat. Commun. 2016, 7, 11426. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Allen, R.S.; Tilbrook, K.; Warden, A.C.; Campbell, P.C.; Rolland, V.; Singh, S.P.; Wood, C.C. Expression of 16 Nitrogenase Proteins within the Plant Mitochondrial Matrix. Front. Plant Sci. 2017, 8, 287. [Google Scholar] [CrossRef] [Green Version]
- Buren, S.; Young, E.M.; Sweeny, E.A.; Lopez-Torrejon, G.; Veldhuizen, M.; Voigt, C.A.; Rubio, L.M. Formation of Nitrogenase NifDK Tetramers in the Mitochondria of Saccharomyces cerevisiae. ACS Synth. Biol. 2017, 6, 1043–1055. [Google Scholar] [CrossRef]
- Eseverri, A.; Lopez-Torrejon, G.; Jiang, X.; Buren, S.; Rubio, L.M.; Caro, E. Use of synthetic biology tools to optimize the production of active nitrogenase Fe protein in chloroplasts of tobacco leaf cells. Plant Biotechnol. J. 2020, 18, 1882–1896. [Google Scholar] [CrossRef] [Green Version]
- Knutson, C.M.; Pieper, M.N.; Barney, B.M. Gene Fitness of Azotobacter vinelandii under Diazotrophic Growth. J. Bacteriol. 2021, 203, e0040421. [Google Scholar] [CrossRef]
- Dos Santos, P.C. Genomic Manipulations of the Diazotroph Azotobacter vinelandii. Methods Mol. Biol. 2019, 1876, 91–109. [Google Scholar] [CrossRef]
- Su, Y.; Wang, T.; Wu, N.; Li, D.; Fan, X.; Xu, Z.; Mishra, S.K.; Yang, M. Alpha-ketoglutarate extends Drosophila lifespan by inhibiting mTOR and activating AMPK. Aging (Albany N. Y.) 2019, 11, 4183–4197. [Google Scholar] [CrossRef]
- Plecita-Hlavata, L.; Engstova, H.; Holendova, B.; Tauber, J.; Spacek, T.; Petraskova, L.; Kren, V.; Spackova, J.; Gotvaldova, K.; Jezek, J.; et al. Mitochondrial Superoxide Production Decreases on Glucose-Stimulated Insulin Secretion in Pancreatic beta Cells Due to Decreasing Mitochondrial Matrix NADH/NAD(+) Ratio. Antioxid. Redox Signal 2020, 33, 789–815. [Google Scholar] [CrossRef]
- Bueno Batista, M.; Brett, P.; Appia-Ayme, C.; Wang, Y.P.; Dixon, R. Disrupting hierarchical control of nitrogen fixation enables carbon-dependent regulation of ammonia excretion in soil diazotrophs. PLoS Genet. 2021, 17, e1009617. [Google Scholar] [CrossRef]
- Reis, V.M.; Teixeira, K.R. Nitrogen fixing bacteria in the family Acetobacteraceae and their role in agriculture. J. Basic Microbiol. 2015, 55, 931–949. [Google Scholar] [CrossRef]
- Barney, B.M.; Eberhart, L.J.; Ohlert, J.M.; Knutson, C.M.; Plunkett, M.H. Gene Deletions Resulting in Increased Nitrogen Release by Azotobacter vinelandii: Application of a Novel Nitrogen Biosensor. Appl. Environ. Microbiol. 2015, 81, 4316–4328. [Google Scholar] [CrossRef] [Green Version]
- Brewin, B.; Woodley, P.; Drummond, M. The basis of ammonium release in nifL mutants of Azotobacter vinelandii. J. Bacteriol. 1999, 181, 7356–7362. [Google Scholar] [CrossRef] [Green Version]
- Michel-Reydellet, N.; Kaminski, P.A. Azorhizobium caulinodans PII and GlnK proteins control nitrogen fixation and ammonia assimilation. J. Bacteriol. 1999, 181, 2655–2658. [Google Scholar] [CrossRef] [Green Version]
- Jaggi, R.; van Heeswijk, W.C.; Westerhoff, H.V.; Ollis, D.L.; Vasudevan, S.G. The two opposing activities of adenylyl transferase reside in distinct homologous domains, with intramolecular signal transduction. EMBO J. 1997, 16, 5562–5571. [Google Scholar] [CrossRef] [Green Version]
- Schnabel, T.; Sattely, E. Engineering Posttranslational Regulation of Glutamine Synthetase for Controllable Ammonia Production in the Plant Symbiont Azospirillum brasilense. Appl. Environ. Microbiol. 2021, 87, e0058221. [Google Scholar] [CrossRef]
- Jonsson, A.; Nordlund, S.; Teixeira, P.F. Reduced activity of glutamine synthetase in Rhodospirillum rubrum mutants lacking the adenylyltransferase GlnE. Res. Microbiol. 2009, 160, 581–584. [Google Scholar] [CrossRef]
- Mus, F.; Tseng, A.; Dixon, R.; Peters, J.W. Diazotrophic Growth Allows Azotobacter vinelandii To Overcome the Deleterious Effects of a glnE Deletion. Appl. Environ. Microbiol. 2017, 83, e00808-17. [Google Scholar] [CrossRef] [Green Version]
- Carroll, P.; Pashley, C.A.; Parish, T. Functional analysis of GlnE, an essential adenylyl transferase in Mycobacterium tuberculosis. J. Bacteriol. 2008, 190, 4894–4902. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ryu, M.H.; Zhang, J.; Toth, T.; Khokhani, D.; Geddes, B.A.; Mus, F.; Garcia-Costas, A.; Peters, J.W.; Poole, P.S.; Ane, J.M.; et al. Control of nitrogen fixation in bacteria that associate with cereals. Nat. Microbiol. 2020, 5, 314–330. [Google Scholar] [CrossRef]
- Haskett, T.L.; Paramasivan, P.; Mendes, M.D.; Green, P.; Geddes, B.A.; Knights, H.E.; Jorrin, B.; Ryu, M.H.; Brett, P.; Voigt, C.A.; et al. Engineered plant control of associative nitrogen fixation. Proc. Natl. Acad. Sci. USA 2022, 119, e2117465119. [Google Scholar] [CrossRef] [PubMed]
- Geddes, B.A.; Paramasivan, P.; Joffrin, A.; Thompson, A.L.; Christensen, K.; Jorrin, B.; Brett, P.; Conway, S.J.; Oldroyd, G.E.D.; Poole, P.S. Engineering transkingdom signalling in plants to control gene expression in rhizosphere bacteria. Nat. Commun. 2019, 10, 3430. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haskett, T.L.; Karunakaran, R.; Bueno Batista, M.; Dixon, R.; Poole, P.S. Control of nitrogen fixation and ammonia excretion in Azorhizobium caulinodans. PLoS Genet. 2022, 18, e1010276. [Google Scholar] [CrossRef]
- Oldroyd, G.E.; Dixon, R. Biotechnological solutions to the nitrogen problem. Curr. Opin Biotechnol. 2014, 26, 19–24. [Google Scholar] [CrossRef]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, W.; Chen, Y.; Huang, K.; Wang, F.; Mei, Z. Molecular Mechanism and Agricultural Application of the NifA–NifL System for Nitrogen Fixation. Int. J. Mol. Sci. 2023, 24, 907. https://doi.org/10.3390/ijms24020907
Zhang W, Chen Y, Huang K, Wang F, Mei Z. Molecular Mechanism and Agricultural Application of the NifA–NifL System for Nitrogen Fixation. International Journal of Molecular Sciences. 2023; 24(2):907. https://doi.org/10.3390/ijms24020907
Chicago/Turabian StyleZhang, Wenyao, Yihang Chen, Keyang Huang, Feng Wang, and Ziqing Mei. 2023. "Molecular Mechanism and Agricultural Application of the NifA–NifL System for Nitrogen Fixation" International Journal of Molecular Sciences 24, no. 2: 907. https://doi.org/10.3390/ijms24020907