

The Involvement of Natural Polyphenols in Molecular Mechanisms Inducing Apoptosis in Tumor Cells: A Promising Adjuvant in Cancer Therapy
 and
and 
Abstract
:1. Introduction
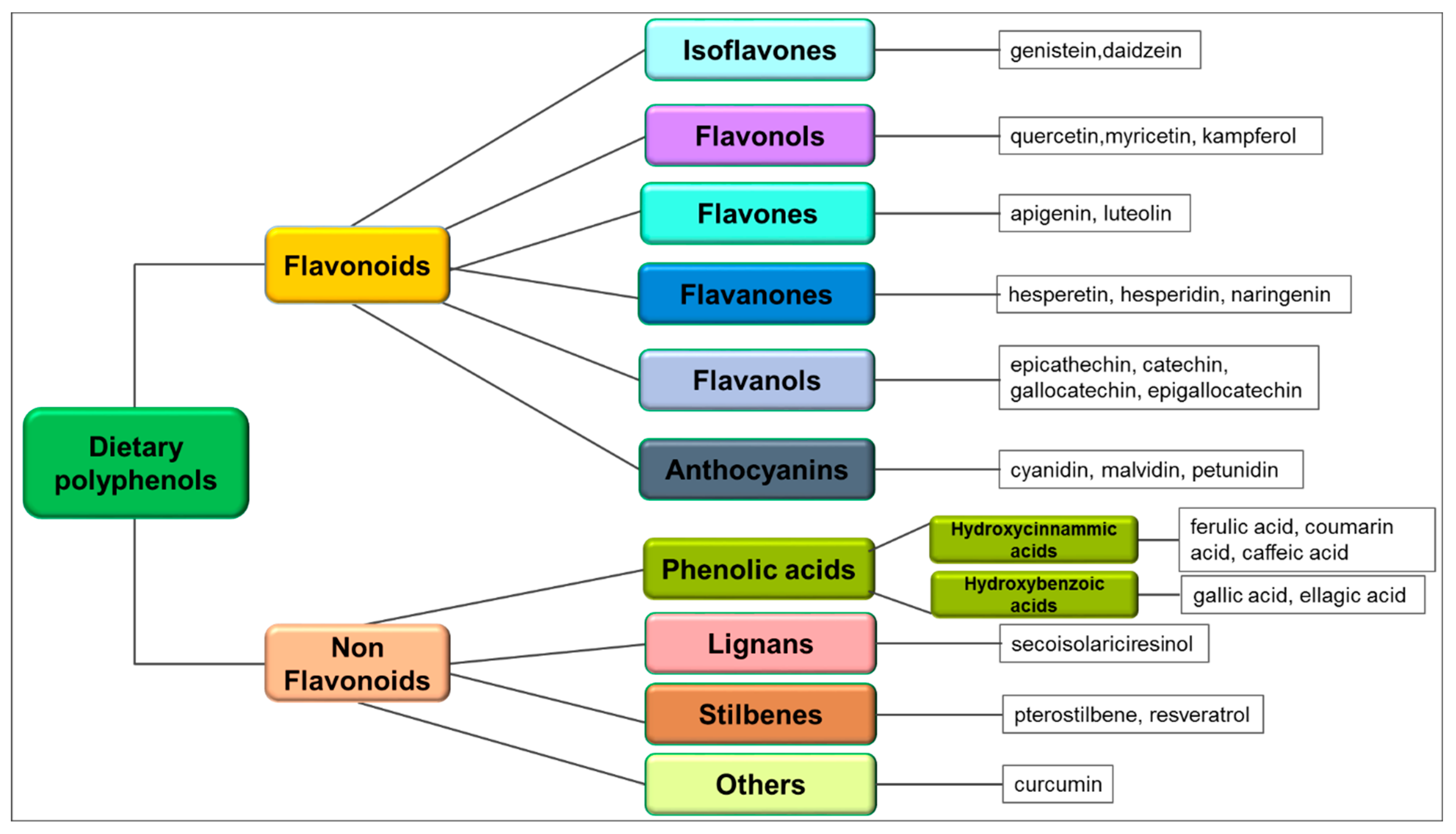
2. Chemical Structure, Sources and Pharmacokinetics of Dietary Polyphenols
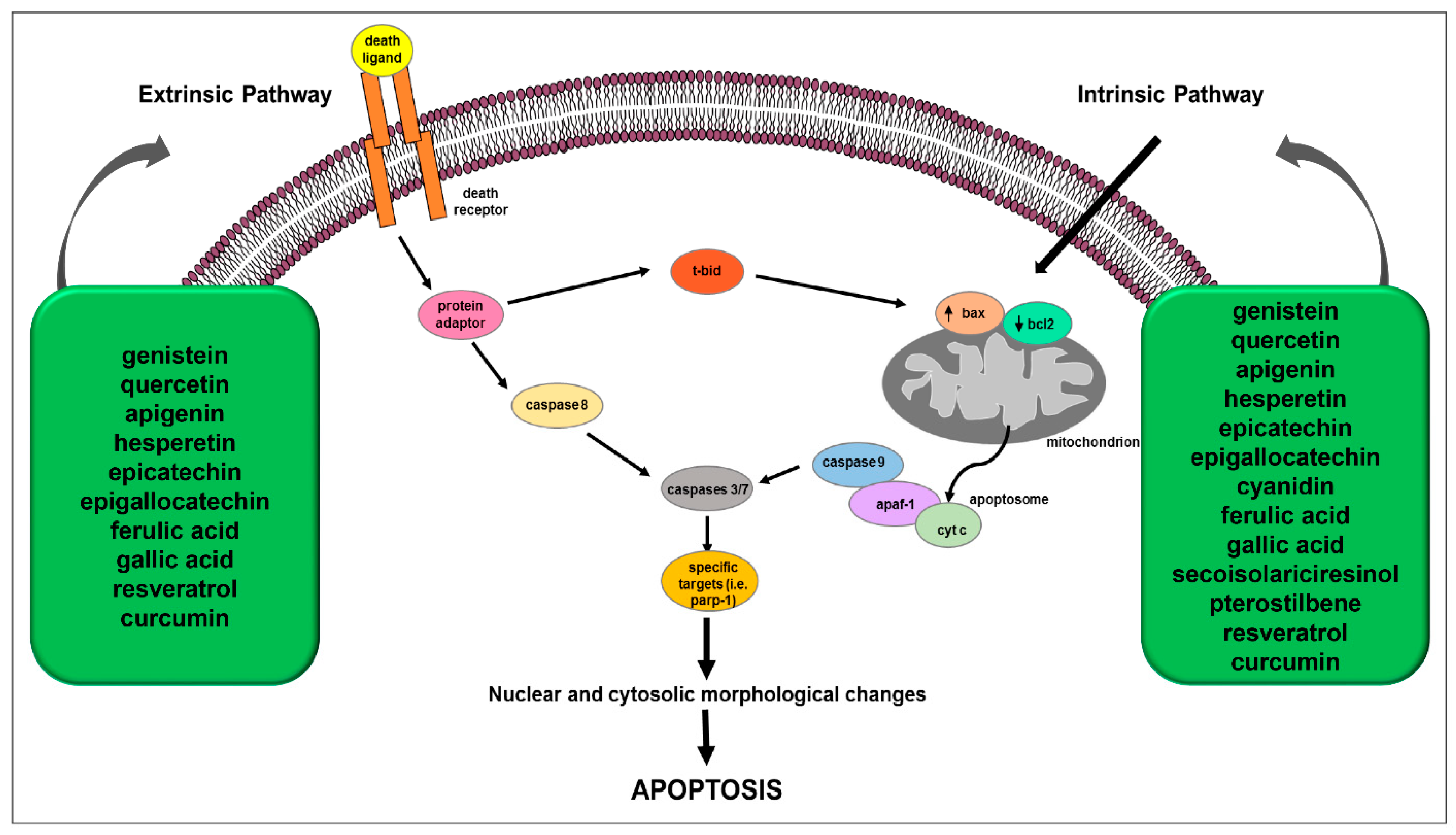
3. Polyphenol-Mediated Apoptosis as Anticancer Mechanism
3.1. Isoflavones and Apoptosis
3.2. Flavonols and Apoptosis
3.3. Flavones and Apoptosis
3.4. Flavanones and Apoptosis
3.5. Flavanols and Apoptosis
3.6. Anthocyanins and Apoptosis
3.7. Phenolic Acids and Apoptosis
3.8. Lignans and Apoptosis
3.9. Stilbenes and Apoptosis
3.10. Curcuminoids and Apoptosis

{kind=link}
{kind=link}
| Family | Phytochemical Compounds | Types or Names of Cancer | Mechanism Leading to Apoptosis | References |
|---|---|---|---|---|
| Flavonoids | ||||
| Isoflavones | genistein | breast | bax↑, bcl-2↓ | [97] |
| bcl-2↓, bcl-xL↓, ROS↑ | [98] | |||
| NSCL | ROS↑, MMP↓, cytochrome c↑, bcl-2↓, bax↑, FOXO3A↑, PUMA↑ | [99] | ||
| leukemia | ROS↑, MMP↓, bcl-2↓, bid↓, ATF-6α↑, GRP78↑, bax↑, bad↑, bak↑, cleaved parp-1↑, caspase-9↑, caspase-3↑ | [100] | ||
| bladder | bcl-2/bax ratio↓, caspase-3, -8 and -9 activation, cytochrome c↑, cleaved parp-1↑ | [101] | ||
| liver | ROS↑, cytochrome c↑, Bax↑, cleaved caspase-3↑, cleaved caspase-9↑, bcl-2↓ | [102] | ||
| colon | caspase-3 activation | [103] | ||
| caspase-3 activation, bax↑ | [104] | |||
| ovarian | bcl-2↓, bcl-xL↓, c-IAP↓, survivin↓ | [105] | ||
| prostate | TRIAL↑, MMP↓ | [106] | ||
| DNA fragmentation↑ | [107] | |||
| Flavonols | quercetin | breast | bax↑, bcl-2↓ | [110,111] |
| AIF↑, MMP↓, caspase-6, -9 activation | [111] | |||
| caspase-8, -3 activation, cleaved parp-1↑ | [116] | |||
| leukemia | bax↑, bad↑, bcl-2↓, cytochrome c↑, caspase-3 activation, cleaved parp-1↑ | [112] | ||
| lung | bax↑, bcl-2↑, caspase-3 activation | [113] | ||
| ovarian | caspase-3 activation | [114] | ||
| bax↑, p21↑, bcl-2↓, p53↑ | [115] | |||
| cervical | TRAIL↑, FasL↑, TNF↑, Fas↑, TNFSF10↑, TNFRSF10A↑, TNFRSF10B↑, TNFRSF1A↑, TNFRSF1B↑, TNFRSF21↑, TNFRSF25↑, TRADD↑, CRADD↑, DEDD↑, caspase-8↑, -10↑, -3↑, -7↑ | [117] | ||
| Flavones | apigenin | prostate | bax↑, bcl-2↓, bcl-xL↓, c-IAP1↓, c-IAP2↓, XIAP↓, survivin↓, cytochrome c↑ | [120] |
| bladder | bax↑, bad↑, bak↑, bcl-2↓, bcl-xL↓, mcl-1↓, caspase-9, -3, -7 activation, cytochrome c↑, cleaved parp-1↑ | [121] | ||
| melanoma | bcl-2↓, bax↑, cleaved caspase-9 ↑, cleaved parp-1↑, p53 ↑ | [122] | ||
| HNSCC | TNF-R↑, TRAIL-R↑, bcl-2↓, caspase activation | [123] | ||
| mesothelioma | bax/bcl-2 ratio↑, p53↑, caspase-8, -9 activation, cleaved parp-1↑ | [124] | ||
| Flavanones | hesperetin | gastric | bax↑, apaf-1↑, bcl-2↓, ROS↑, caspase-9, -3 activation, MMP↓, cytochrome c↑, DNA fragmentation | [126] |
| esophageal | ROS↑, bax↑, apaf-1↑, SuFu↑, bcl-2↓, caspase-9, -3 activation | [127] | ||
| colon | ROS↑, bax↑, bcl-2↓, cytochrome c↑, cleaved caspase-3 ↑, SOD↓, CAT↓, GPx↓ | [128] | ||
| breast | ROS↑, bax/bcl-2 ratio↑, phosphatydilserine externalization, caspase-9, -7 activation, MMP↓, cytochrome c↑,cleaved parp-1↑, DNA fragmentation | [129] | ||
| lung | Fas↑, FADD↑, caspase-8 activation | [130] | ||
| cytosolic bax↓, mitochondrial bax↑ | [131] | |||
| cervical | extrinsic and intrinsic apoptotic pathways | [132] | ||
| Flavanols | epicatechin | breast | DR4↑, DR5↑, ROS↑, MMP↓, cytochrome c↑, Smac/Diablo↑, HtrA2/Omi↑, TRAIL↑, bad↑, bax ↑ | [134] |
| bax↑, cytochrome c↑, caspase-3 activation | [136] | |||
| colon | bcl-2↓, bax↑, p53↑, DNA fragmentation | [135] | ||
| prostate | bax↑, cytochrome c↑, caspase-3 activation | [136] | ||
| Anthocyanins | cyanidin | prostate | caspase-3 activation, DNA fragmentation | [138] |
| glioblastoma | bax↑, p53↑, bcl-2↓ | [139] | ||
| osteosarcoma | bax↑, PPARγ↑, p21↑ | [140] | ||
| Non Flavonoids | ||||
| Hydroxybenzoic acids | ferulic acid | breast | caspase-8, -9 activation | [142] |
| bax/bcl-2 ratio↑ | [143] | |||
| hepatic | caspase-8, -9 activation | [142] | ||
| cervical | bax↑, bcl-2↓, mcl-1↓ | [144] | ||
| Hydroxycinnamic acids | gallic acid | gastric | Fas↑, FasL↑, DR5↑, caspase-8, -9, -3 activation, bad↑, bak↑, bcl-2↓, p53↑ | [145] |
| breast | MMP↓, caspase-8, -9 activation, cytochrome c↑ | [146] | ||
| bax↑, bcl-2↓, caspase-3 ↑, p53↑ | [147] | |||
| Lignans | secoisolariciresinol | colon | AIF↑, caspase-3↑ | [149] |
| breast | bcl-2↓ | [150] | ||
| Stilbenes | pterostilbene | osteosarcoma | bax↑, bak↑, bcl-xL↓, mcl-1↓, caspase-3 ↑, cytochrome c↑ | [152] |
| breast | superoxide anion↑, MMP↓, caspase-3, -7 activation | [153] | ||
| bax↑, p53 mutant↓ | [154] | |||
| bax↑ | [155] | |||
| endometrial | bcl-2↓, bcl-xL↓, caspase-3 activation | [156] | ||
| cervical | ROS↑, bcl-2↓, bcl-xL↓, caspase-9, -3 activation | [157] | ||
| liver | ROS↑, SOD2↓, p53↑ | [158] | ||
| Curcuminoids | curcumin | leukemia | ROS↑ | [161] |
| melanoma | ROS↑ | [162] | ||
| caspase-8 activation | [173] | |||
| breast | ROS↑, TRAIL↑ | [163] | ||
| p53↑, bax↑ | [169] | |||
| bcl-2↓ | [171] | |||
| renal | ROS↑, TRAIL↑ | [164] | ||
| TRAIL↑ | [168] | |||
| glioblastoma | TRAIL↑ | [165] | ||
| prostate | TRAIL↑ | [166] | ||
| bcl-2↓ | [172] | |||
| hepatocellular carcinoma | FasL↑ caspase-3 activation | [167] | ||
| laryngeal | bcl-2↓ | [170] | ||
| colon | p53↑, bax↑ | [36] | ||
| neuroblastoma | caspase-3 activation | [174] | ||
| linfoma | caspase-3 activation | [175] | ||
| lung | caspase-3 activation | [176] | ||
| schwannoma | bcl-2↓, bax↑, parp-1↑, caspase-3↑, caspase-9↑ | [177] | ||
| thyroid | ROS↑, caspase-3 activation, bcl-2↓, bcl-xL↓, bax↑ | [178] |
4. Dietary Polyphenols and Drug Synergism in Cancer Therapy
| Phytochemical Compounds/ Chemotherapy Drugs | Cell Lines | Outcomes | References |
|---|---|---|---|
| genistein+cisplatin | CaSki cervical cancer | ERK1/2 phosphorylation↓, bcl-2↓, p53↑, cleaved caspase-3↑ | [181] |
| genistein+centchroman | MCF-7, MDA-MB-231 breast cancer | PI3K/AKT/NF-κB phosphorylation↓, cell cycle arrest at G2/M phase, ROS↑, bax↑, bcl-2↓, caspase-3, -7, -9↑, cleaved parp-1↑ | [182] |
| quercetin+cisplatin | HeLa, SiHa cervical cancer | cell viability↓, apoptosis↑ | [183] |
| quercetin+methotrexate | Saos-2 osteosarcoma | cell viability↓, apoptosis↑, p53↑, CBX7↑, CYLD ↑, bcl-2↓, miR-223↓ | [184] |
| quercetin+tamoxifen | MCF-7 breast cancer | apoptosis↑, p53↑, bax↑, p21↑, and bcl-2↓ | [185] |
| apigenin+cetuximab | HONE1, CNE2 nasopharyngeal carcinoma | cell viability↓, apoptosis↑, bax/bcl-2 ratio↑, caspase-3↑, p-EGFR↓, p-AKT↓, p-STAT3↓, cyclin D1↓ | [186] |
| apigenin+cisplatin | A549 lung, MCF-7 breast, HCT 116 colorectal, HeLa cervical cancers | cell viability↓, apoptosis↑, p53↑, DNA damage↑ | [187] |
| apigenin+paclitaxel | HeLa cervical, A549 lung, Hep3B negroid hepatocyte, HEK293A embryonic kidney cancer | cell viability↓ | [188] |
| HeLa cervical cancer | apoptosis↑, ROS↑, caspase-2↑, MMP ↓ | [188] | |
| apigenin+etoposide apigenin+cyclophosphamide | Jurkat lymphoid and THP-1 myeloid leukaemia | apoptosis↑, cytochrome c↑, Smac/Diablo↑, HtrA2/Omi ↑, caspase-9, -3 activation | [189] |
| apigenin+quercetin+emodin+rhein+ cisplatin and cyclophosphamide apigenin+rhein+ chlorambucil | Jurkat and CCRF-CEM lymphoid leukaemia | apoptosis↑, ATP↓, glutathione↓, cell cycle arrest, DNA damage↑ | [190] |
| apigenin+quercetin,+emodin+cisplatin, cyclophosphamide and chlorambucil | THP1 and KG1a myeloid leukaemia | apoptosis↑, ATP↓, glutathione↓, cell cycle arrest, DNA damage↑ | [190] |
| hesperetin+cisplatin | HGC-27, SGC-7901, and MGC-803 gastric | PTEN↑, cytosolic cytochrome c↑, bax↑, cleaved caspase-9 and -3↑, AIF↑, p-AKT↓, bcl-2↓ | [191] |
| cyanidin-3-O-glucoside+ cisplatin | HeLa cervical cancer | Nrf2↓, HO-1↓, NQO1↓, bax/bcl-2 ratio↑ | [192] |
| gallic acid+cisplatin | A549 non-small cell lung cancer | JAK/STAT3↓, bax↑, bcl-2↓ | [193] |
| pterostilbene+gemcitabine | MIA PaCa-2, MIA PaCa-2 GEMR (gemcitabine-resistant) pancreatic ductal adenocarcinoma | S-phase cell cycle arrest, apoptosis↑, MDR1↓, RAGE/PI3K/AKT signaling↓ | [194] |
| pterostilbene+epigallocatechin gallate | MIA PaCa-2 pancreatic cancer | cell proliferation↓, mitochondrial depolarization, cytochrome c release↑ | [195] |
| PANC-1 pancreatic cancer | cell proliferation↓ | [195] | |
| curcumin+5-fluorouracil or doxorubicin | NT8e head and neck squamous cell carcinoma | cell proliferation↓, EGFR/ERK1/2 signaling↓, G1/S cell cycle arrest, apoptosis↑, bcl-2↓, bax↑, caspase-3 activation, cleaved parp-1↑ | [196] |
| curcumin+paclitaxel | MCF-7 breast cancer | cell proliferation↓, EGFR signaling↓, apoptosis↑, bcl-2↓, bax↑ | [197] |
| curcumin+paclitaxel | MCF-7, MDA-MB-231 breast cancer | apoptosis↑, caspase-3↑, cleaved parp-1↑ | [198] |
| curcumin+paclitaxel | CaSki, HeLa cervical cancer | NF-κB signaling↓, p53↑, caspase-3↑ | [199] |
| curcumin+quercetin | K562 chronic myeloid leukemia | CDKN1B↓, AKT1↓, IFN-γ↓, BTG2↑, p21↑, p27↑, CDKN1A↑, Fas↑, FasL↑ | [200] |
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| AGS | gastric adenocarcinoma |
| AKT | protein-chinasi B |
| APAF-1 | apoptotic protease activating factor-1 |
| ATF6 | activating transcription factor 6 |
| ATP | adenosine triphosphate |
| BAD | BCL-2 associated agonist of cell death |
| BAK | BCL-2 antagonist/killer |
| BAX | BCL-2-associated X protein |
| BCL-2 | B-cell lymphoma-2 |
| BCL-w | B-cell lymphoma-w |
| BCL-XL | B-cell lymphoma-extra large |
| BFL-1/A1 | Bcl-2-related protein A1 |
| BIC | bone-inducing protein complex |
| BIM | BCL-2-interacting mediator of cell death |
| BMF | BCL-2 modifying factor |
| BOC | cell adhesion associated, oncogene regulated |
| BTG2 | BTG family member 2 |
| c-XIAP1 | cellular inhibitor of apoptosis protein 1 |
| c-XIAP2 | cellular inhibitor of apoptosis protein 2 |
| CASPASE | cysteine-dependent aspartate-directed protease |
| CAT | catalase |
| CBX7 | chromobox protein homolog 7 |
| CDKN1A | cyclin dependent kinase inhibitor 1A |
| CDKN1B | cyclin-dependent kinase inhibitor 1B |
| c-IAP1 | cellular inhibitor of apoptosis protein 1 |
| COX2 | cyclooxygenase 2 |
| CRADD | CASP2 and RIPK1 domain containing adaptor with death domain |
| CYLD | ubiquitin carboxyl-terminal hydrolase |
| DEDD | death effector domain containing protein |
| DNA | deoxyribonucleic acid |
| DOX | doxorubicin |
| DR | death receptor |
| DR-5 | death receptor 5 |
| EGCG | epigallocatechin gallate |
| EGFR | epidermal growth factor receptor |
| ER | estrogen receptor |
| ERK | extracellular signal-regulated kinase |
| ERK1/2 | extracellular signal-regulated kinase 1 |
| ERα | estrogen receptor alpha |
| ERβ | estrogen receptor beta |
| FADD | fas-associated death domain |
| FAS | fas cell surface death receptor |
| FASL | fas ligand |
| FOXO3a | forkhead box O3a |
| GPx | glutathione peroxidase |
| GRP78 | glucose-regulated protein 78 |
| GSH | reduced glutathione |
| HNSCC | head and neck squamous cell carcinoma |
| HO-1 | heme oxygenase-1 |
| HONE1 | epithelial tumor cell lines 1 |
| HRK | harakiri, BCL-2 interacting protein |
| HTRA2 | high-temperature requirement protein A2 |
| IFN-γ | interferon gamma |
| IGF-IR | type 1 insulin-like growth factor receptor |
| JAK | janus kinase |
| JNK | c-Jun N-terminal kinase |
| LPH | phloridzine hydrolase |
| MAPK | mitogen-activated protein kinase |
| MCL-1 | myeloid cell leukemia-1 |
| MDR1 | multidrug resistance protein 1 |
| MMP | mitochondrial membrane potential |
| mTOR | mammalian target of rapamycin |
| NAC | N-acetylcysteine |
| ncRNAs | non-coding RNAs |
| NF-κB | nuclear factor kappa B |
| NOXA | phorbol-12-myristate-13-acetate-induced protein 1 |
| NQO1 | NAD(P)H quinone oxidoreductase |
| NRF2 | nuclear factor erythroid 2–related factor 2 |
| NSCLC | non-small cell lung cancer |
| PARP-1 | poly [ADP-ribose] polymerase 1 |
| PI3K | phoshatidylinositol-3 kinase |
| PKC | protein kinase C |
| PPARγ | peroxisome proliferator-activated receptor gamma |
| PTC | papillary thyroid cancer |
| PTEN | phosphatase and tensin homolog |
| PTK | protein tyrosine kinase |
| PUMA | p53-up-regulated modulator of apoptosis |
| RAGE | receptor for advanced glycation end products |
| ROS | reactive oxygen species |
| SMAC/DIABLO | second mitochondria-derived activator of caspase/direct inhibitor of apoptosis-binding protein with low isoelectric point |
| SOD | superoxide dismutase |
| STAT | signal transducer and activator of transcription |
| STAT3 | signal transducer and activator of transcription 3 |
| TNF | tumor necrosis factor |
| TNF-α | tumor necrosis factor alpha |
| TNF-R | tumor necrosis factor receptor |
| TNFRSF10A | tumor necrosis factor receptor superfamily 10A |
| TNFRSF10B | tumor necrosis factor receptor superfamily 10B |
| TNFRSF1B | tumor necrosis factor receptor superfamily member 1B |
| TNFRSF21 | tumor necrosis factor receptor superfamily member 21 |
| TNFRSF25 | tumor necrosis factor receptor superfamily member 25 |
| TNFSF10 | tumor necrosis factor (ligand) superfamily member 10 |
| TRADD | tumor necrosis factor receptor type 1-associated death domain protein |
| TRAIL | TNF-related apoptosis-inducing ligand |
| TRAIL-R | TNF-related apoptosis-inducing ligand receptor |
| XIAP | X-linked inhibitor of apoptosis |
| ZIP9 | Zrt- and Irt-like protein 9 |
References
- Hanahan, D. Hallmarks of Cancer: New Dimensions. Cancer Discov. 2022, 12, 31–46. [Google Scholar] [CrossRef] [PubMed]
- Global Burden of Disease 2019 Cancer Collaboration; Kocarnik, J.M.; Compton, K.; Dean, F.E.; Fu, W.; Gaw, B.L.; Harvey, J.D.; Henrikson, H.J.; Lu, D.; Pennini, A.; et al. Cancer Incidence, Mortality, Years of Life Lost, Years Lived With Disability, and Disability-Adjusted Life Years for 29 Cancer Groups From 2010 to 2019: A Systematic Analysis for the Global Burden of Disease Study 2019. JAMA Oncol. 2022, 8, 420–444. [Google Scholar] [CrossRef] [PubMed]
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global Cancer Statistics 2020: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef]
- Ansari, K.K.; Jha, A. Causes of Cancer in the World: Comparative Risk Assessment of Nine Behavioral and Environmental Risk Factors. Cureus 2022, 14, e28875. [Google Scholar] [CrossRef]
- Emanuele Liardo, R.L.; Borzi, A.M.; Spatola, C.; Martino, B.; Privitera, G.; Basile, F.; Biondi, A.; Vacante, M. Effects of infections on the pathogenesis of cancer. Indian J. Med. Res. 2021, 153, 431–445. [Google Scholar] [CrossRef] [PubMed]
- Neben, C.L.; Zimmer, A.D.; Stedden, W.; van den Akker, J.; O’Connor, R.; Chan, R.C.; Chen, E.; Tan, Z.; Leon, A.; Ji, J.; et al. Multi-Gene Panel Testing of 23,179 Individuals for Hereditary Cancer Risk Identifies Pathogenic Variant Carriers Missed by Current Genetic Testing Guidelines. J. Mol. Diagn. 2019, 21, 646–657. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- GBD 2019 Cancer Risk Factors Collaborators. The global burden of cancer attributable to risk factors, 2010–2019: A systematic analysis for the Global Burden of Disease Study 2019. Lancet 2022, 400, 563–591. [Google Scholar] [CrossRef]
- Wong, R.S. Apoptosis in cancer: From pathogenesis to treatment. J. Exp. Clin. Cancer Res. 2011, 30, 87. [Google Scholar] [CrossRef] [Green Version]
- Fernald, K.; Kurokawa, M. Evading apoptosis in cancer. Trends Cell Biol. 2013, 23, 620–633. [Google Scholar] [CrossRef] [Green Version]
- Jan, R.; Chaudhry, G.E.S. Understanding Apoptosis and Apoptotic Pathways Targeted Cancer Therapeutics. Adv. Pharm. Bull. 2019, 9, 205–218. [Google Scholar] [CrossRef]
- Vali-Pour, M.; Lehner, B.; Supek, F. The impact of rare germline variants on human somatic mutation processes. Nat. Commun. 2022, 13, 3724. [Google Scholar] [CrossRef] [PubMed]
- Levatic, J.; Salvadores, M.; Fuster-Tormo, F.; Supek, F. Mutational signatures are markers of drug sensitivity of cancer cells. Nat. Commun. 2022, 13, 2926. [Google Scholar] [CrossRef] [PubMed]
- Zhivotovsky, B.; Kroemer, G. Apoptosis and genomic instability. Nat. Rev. Mol. Cell Biol. 2004, 5, 752–762. [Google Scholar] [CrossRef] [PubMed]
- Bonora, M.; Missiroli, S.; Perrone, M.; Fiorica, F.; Pinton, P.; Giorgi, C. Mitochondrial Control of Genomic Instability in Cancer. Cancers 2021, 13, 1914. [Google Scholar] [CrossRef] [PubMed]
- DePinho, R.A. The age of cancer. Nature 2000, 408, 248–254. [Google Scholar] [CrossRef]
- Konishi, A.; Shimizu, S.; Hirota, J.; Takao, T.; Fan, Y.; Matsuoka, Y.; Zhang, L.; Yoneda, Y.; Fujii, Y.; Skoultchi, A.I.; et al. Involvement of histone H1.2 in apoptosis induced by DNA double-strand breaks. Cell 2003, 114, 673–688. [Google Scholar] [CrossRef] [Green Version]
- Storchova, Z.; Pellman, D. From polyploidy to aneuploidy, genome instability and cancer. Nat. Rev. Mol. Cell Biol. 2004, 5, 45–54. [Google Scholar] [CrossRef]
- Chen, J.G.; Yang, C.P.; Cammer, M.; Horwitz, S.B. Gene expression and mitotic exit induced by microtubule-stabilizing drugs. Cancer Res. 2003, 63, 7891–7899. [Google Scholar]
- Rallis, K.S.; Lai Yau, T.H.; Sideris, M. Chemoradiotherapy in Cancer Treatment: Rationale and Clinical Applications. Anticancer Res. 2021, 41, 1–7. [Google Scholar] [CrossRef]
- Kocakavuk, E.; Anderson, K.J.; Varn, F.S.; Johnson, K.C.; Amin, S.B.; Sulman, E.P.; Lolkema, M.P.; Barthel, F.P.; Verhaak, R.G.W. Radiotherapy is associated with a deletion signature that contributes to poor outcomes in patients with cancer. Nat. Genet. 2021, 53, 1088–1096. [Google Scholar] [CrossRef]
- Liu, Y.P.; Zheng, C.C.; Huang, Y.N.; He, M.L.; Xu, W.W.; Li, B. Molecular mechanisms of chemo- and radiotherapy resistance and the potential implications for cancer treatment. Med. Comm. 2021, 2, 315–340. [Google Scholar] [CrossRef] [PubMed]
- Maleki Dana, P.; Sadoughi, F.; Asemi, Z.; Yousefi, B. The role of polyphenols in overcoming cancer drug resistance: A comprehensive review. Cell. Mol. Biol. Lett. 2022, 27, 1. [Google Scholar] [CrossRef] [PubMed]
- Chairez-Ramirez, M.H.; de la Cruz-Lopez, K.G.; Garcia-Carranca, A. Polyphenols as Antitumor Agents Targeting Key Players in Cancer-Driving Signaling Pathways. Front. Pharmacol. 2021, 12, 710304. [Google Scholar] [CrossRef] [PubMed]
- Montane, X.; Kowalczyk, O.; Reig-Vano, B.; Bajek, A.; Roszkowski, K.; Tomczyk, R.; Pawliszak, W.; Giamberini, M.; Mocek-Plociniak, A.; Tylkowski, B. Current Perspectives of the Applications of Polyphenols and Flavonoids in Cancer Therapy. Molecules 2020, 25, 3342. [Google Scholar] [CrossRef]
- Rudrapal, M.; Khairnar, S.J.; Khan, J.; Dukhyil, A.B.; Ansari, M.A.; Alomary, M.N.; Alshabrmi, F.M.; Palai, S.; Deb, P.K.; Devi, R. Dietary Polyphenols and Their Role in Oxidative Stress-Induced Human Diseases: Insights Into Protective Effects, Antioxidant Potentials and Mechanism(s) of Action. Front. Pharmacol. 2022, 13, 806470. [Google Scholar] [CrossRef]
- Gasmi, A.; Mujawdiya, P.K.; Noor, S.; Lysiuk, R.; Darmohray, R.; Piscopo, S.; Lenchyk, L.; Antonyak, H.; Dehtiarova, K.; Shanaida, M.; et al. Polyphenols in Metabolic Diseases. Molecules 2022, 27, 6280. [Google Scholar] [CrossRef]
- Yahfoufi, N.; Alsadi, N.; Jambi, M.; Matar, C. The Immunomodulatory and Anti-Inflammatory Role of Polyphenols. Nutrients 2018, 10, 1618. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Y.; Zhang, X. Interactions of tea polyphenols with intestinal microbiota and their implication for anti-obesity. J. Sci. Food Agric. 2020, 100, 897–903. [Google Scholar] [CrossRef]
- Grosso, G.; Godos, J.; Lamuela-Raventos, R.; Ray, S.; Micek, A.; Pajak, A.; Sciacca, S.; D’Orazio, N.; Del Rio, D.; Galvano, F. A comprehensive meta-analysis on dietary flavonoid and lignan intake and cancer risk: Level of evidence and limitations. Mol. Nutr. Food Res. 2017, 61, 1600930. [Google Scholar] [CrossRef]
- Messina, M. Impact of Soy Foods on the Development of Breast Cancer and the Prognosis of Breast Cancer Patients. Forsch. Komplementmed. 2016, 23, 75–80. [Google Scholar] [CrossRef]
- Gardeazabal, I.; Romanos-Nanclares, A.; Martinez-Gonzalez, M.A.; Sanchez-Bayona, R.; Vitelli-Storelli, F.; Gaforio, J.J.; Aramendia-Beitia, J.M.; Toledo, E. Total polyphenol intake and breast cancer risk in the Seguimiento Universidad de Navarra (SUN) cohort. Br. J. Nutr. 2019, 122, 542–551. [Google Scholar] [CrossRef] [PubMed]
- Di Lorenzo, C.; Colombo, F.; Biella, S.; Stockley, C.; Restani, P. Polyphenols and Human Health: The Role of Bioavailability. Nutrients 2021, 13, 273. [Google Scholar] [CrossRef] [PubMed]
- Jang, Y.G.; Go, R.E.; Hwang, K.A.; Choi, K.C. Resveratrol inhibits DHT-induced progression of prostate cancer cell line through interfering with the AR and CXCR4 pathway. J. Steroid Biochem. 2019, 192, 105406. [Google Scholar] [CrossRef] [PubMed]
- Deb, G.; Shankar, E.; Thakur, V.S.; Ponsky, L.E.; Bodner, D.R.; Fu, P.F.; Gupta, S. Green tea-induced epigenetic reactivation of tissue inhibitor of matrix metalloproteinase-3 suppresses prostate cancer progression through histone-modifying enzymes. Mol. Carcinog. 2019, 58, 1194–1207. [Google Scholar] [CrossRef] [PubMed]
- Yuan, L.; Zhou, M.M.; Huang, D.W.; Wasan, H.S.; Zhang, K.; Sun, L.T.; Huang, H.; Ma, S.L.; Shen, M.H.; Ruan, S.M. Resveratrol inhibits the invasion and metastasis of colon cancer through reversal of epithelial-mesenchymal transition via the AKT/GSK-3 beta/Snail signaling pathway. Mol. Med. Rep. 2022, 26, 354. [Google Scholar] [CrossRef]
- Li, P.H.; Pu, S.M.; Lin, C.; He, L.; Zhao, H.X.; Yang, C.; Guo, Z.Q.; Xu, S.S.; Zhou, Z.P. Curcumin selectively induces colon cancer cell apoptosis and S cell cycle arrest by regulates Rb/E2F/p53 pathway. J. Mol. Struct. 2022, 1263, 133180. [Google Scholar] [CrossRef]
- Zan, L.L.; Chen, Q.F.; Zhang, L.; Li, X.N. Epigallocatechin gallate (EGCG) suppresses growth and tumorigenicity in breast cancer cells by downregulation of miR-25. Bioengineered 2019, 10, 374–382. [Google Scholar] [CrossRef] [Green Version]
- Bayat, S.; Derakhshan, S.M.; Derakhshan, N.M.; Khaniani, M.S.; Alivand, M.R. Downregulation of HDAC2 and HDAC3 via oleuropein as a potent prevention and therapeutic agent in MCF-7 breast cancer cells. J. Cell. Biochem. 2019, 120, 9172–9180. [Google Scholar] [CrossRef]
- Alsamri, H.; Alneyadi, A.; Muhammad, K.; Ayoub, M.A.; Eid, A.; Iratni, R. Carnosol Induces p38-Mediated ER Stress Response and Autophagy in Human Breast Cancer Cells. Front. Oncol. 2022, 12, 911615. [Google Scholar] [CrossRef]
- Wu, M.F.; Huang, Y.H.; Chiu, L.Y.; Cherng, S.H.; Sheu, G.T.; Yang, T.Y. Curcumin Induces Apoptosis of Chemoresistant Lung Cancer Cells via ROS-Regulated p38 MAPK Phosphorylation. Int. J. Mol. Sci. 2022, 23, 8248. [Google Scholar] [CrossRef]
- Sun, X.L.; Xiang, Z.M.; Xie, Y.R.; Zhang, N.; Wang, L.X.; Wu, Y.L.; Zhang, D.Y.; Wang, X.J.; Sheng, J.; Zi, C.T. Dimeric-(−)-epigallocatechin-3-gallate inhibits the proliferation of lung cancer cells by inhibiting the EGFR signaling pathway. Chem.-Biol. Interact. 2022, 365, 110084. [Google Scholar] [CrossRef] [PubMed]
- Barros, T.M.B.; Lima, A.P.B.; Almeida, T.C.; da Silva, G.N. Inhibition of urinary bladder cancer cell proliferation by silibinin. Environ. Mol. Mutagen. 2020, 61, 445–455. [Google Scholar] [CrossRef] [PubMed]
- Adami, B.S.; Diz, F.M.; Goncalves, G.P.O.; Reghelin, C.K.; Scherer, M.; Dutra, A.P.; Papaleo, R.M.; de Oliveira, J.R.; Morrone, F.B.; Wieck, A.; et al. Morphological and mechanical changes induced by quercetin in human T24 bladder cancer cells. Micron 2021, 151, 103152. [Google Scholar] [CrossRef] [PubMed]
- Wei, R.; Penso, N.E.C.; Hackman, R.M.; Wang, Y.F.; Mackenzie, G.G. Epigallocatechin-3-Gallate (EGCG) Suppresses Pancreatic Cancer Cell Growth, Invasion, and Migration partly through the Inhibition of Akt Pathway and Epithelial-Mesenchymal Transition: Enhanced Efficacy When Combined with Gemcitabine. Nutrients 2019, 11, 1856. [Google Scholar] [CrossRef] [Green Version]
- Ratajczak, K.; Glatzel-Plucinska, N.; Ratajczak-Wielgomas, K.; Nowinska, K.; Borska, S. Effect of Resveratrol Treatment on Human Pancreatic Cancer Cells through Alterations of Bcl-2 Family Members. Molecules 2021, 26, 6560. [Google Scholar] [CrossRef]
- Jokar, M.H.; Sedighi, S.; Moradzadeh, M. A comparative study of anti-leukemic effects of kaempferol and epigallocatechin-3-gallate (EGCG) on human leukemia HL-60 cells. Avicenna J. Phytomed. 2021, 11, 314–323. [Google Scholar] [CrossRef]
- Kuttikrishnan, S.; Siveen, K.S.; Prabhu, K.S.; Khan, A.Q.; Ahmed, E.I.; Akhtar, S.; Ali, T.A.; Merhi, M.; Dermime, S.; Steinhoff, M.; et al. Curcumin Induces Apoptotic Cell Death via Inhibition of PI3-Kinase/AKT Pathway in B-Precursor Acute Lymphoblastic Leukemia. Front. Oncol. 2019, 9, 484. [Google Scholar] [CrossRef] [Green Version]
- Dong, C.Q.; Wang, Z.G.; Shen, P.; Chen, Y.G.; Wang, J.S.; Wang, H.B. Epigallocatechin-3-gallate suppresses the growth of human osteosarcoma by inhibiting the Wnt/beta-catenin signalling pathway. Bioengineered 2022, 13, 8490–8502. [Google Scholar] [CrossRef]
- De Luca, A.; Bellavia, D.; Raimondi, L.; Carina, V.; Costa, V.; Fini, M.; Giavaresi, G. Multiple Effects of Resveratrol on Osteosarcoma Cell Lines. Pharmaceuticals 2022, 15, 342. [Google Scholar] [CrossRef]
- Qin, J.L.; Fu, M.L.; Wang, J.; Huang, F.X.; Liu, H.P.; Huangfu, M.J.; Yu, D.; Liu, H.W.; Li, X.M.; Guan, X.; et al. PTEN/AKT/mTOR signaling mediates anticancer effects of epigallocatechin-3-gallate in ovarian cancer. Oncol. Rep. 2020, 43, 1885–1896. [Google Scholar] [CrossRef]
- Sun, S.F.; Fang, H.L. Curcumin inhibits ovarian cancer progression by regulating circ-PLEKHM3/miR-320a/SMG1 axis. J. Ovarian Res. 2021, 14, 158. [Google Scholar] [CrossRef] [PubMed]
- Dei Cas, M.; Ghidoni, R. Cancer Prevention and Therapy with Polyphenols: Sphingolipid-Mediated Mechanisms. Nutrients 2018, 10, 940. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bhosale, P.B.; Ha, S.E.; Vetrivel, P.; Kim, H.H.; Kim, S.M.; Kim, G.S. Functions of polyphenols and its anticancer properties in biomedical research: A narrative review. Transl. Cancer Res. 2020, 9, 7619–7631. [Google Scholar] [CrossRef] [PubMed]
- Selvakumar, P.; Badgeley, A.; Murphy, P.; Anwar, H.; Sharma, U.; Lawrence, K.; Lakshmikuttyamma, A. Flavonoids and Other Polyphenols Act as Epigenetic Modifiers in Breast Cancer. Nutrients 2020, 12, 761. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Curti, V.; Di Lorenzo, A.; Dacrema, M.; Xiao, J.B.; Nabavi, S.M.; Daglia, M. In vitro polyphenol effects on apoptosis: An update of literature data. Semin. Cancer Biol. 2017, 46, 119–131. [Google Scholar] [CrossRef]
- Antonsson, B.; Martinou, J.C. The Bcl-2 protein family. Exp. Cell. Res. 2000, 256, 50–57. [Google Scholar] [CrossRef]
- Li, J.; Yuan, J. Caspases in apoptosis and beyond. Oncogene 2008, 27, 6194–6206. [Google Scholar] [CrossRef] [Green Version]
- Aubrey, B.J.; Kelly, G.L.; Janic, A.; Herold, M.J.; Strasser, A. How does p53 induce apoptosis and how does this relate to p53-mediated tumour suppression? Cell Death Differ. 2018, 25, 104–113. [Google Scholar] [CrossRef] [Green Version]
- Singla, R.K.; Dubey, A.K.; Garg, A.; Sharma, R.K.; Fiorino, M.; Ameen, S.M.; Haddad, M.A.; Al-Hiary, M. Natural Polyphenols: Chemical Classification, Definition of Classes, Subcategories, and Structures. J. Aoac. Int. 2019, 102, 1397–1400. [Google Scholar] [CrossRef]
- Lopez-Lazaro, M. Flavonoids as anticancer agents: Structure-activity relationship study. Curr. Med. Chem. Anticancer Agents 2002, 2, 691–714. [Google Scholar] [CrossRef]
- Khare, S.; Dewangan, R.P.; Kumar, A. Structure-Activity Relationship of Flavonoids: Recent Updates. In The Chemistry Inside Spices & Herbs: Research and Development; Bentham Science Publishers: Sharjah, United Arab Emirates, 2022; pp. 235–256. [Google Scholar] [CrossRef]
- Kim, K.; Vance, T.M.; Chun, O.K. Estimated intake and major food sources of flavonoids among US adults: Changes between 1999–2002 and 2007–2010 in NHANES. Eur. J. Nutr. 2016, 55, 833–843. [Google Scholar] [CrossRef] [PubMed]
- Janabi, A.H.W.; Kamboh, A.A.; Saeed, M.; Lu, X.Y.; BiBi, J.; Majeed, F.; Naveed, M.; Mughal, M.J.; Korejo, N.A.; Kamboh, R.; et al. Flavonoid-rich foods (FRF): A promising nutraceutical approach against lifespan-shortening diseases. Iran. J. Basic Med. Sci 2020, 23, 140–153. [Google Scholar] [CrossRef]
- Zaheer, K.; Humayoun Akhtar, M. An updated review of dietary isoflavones: Nutrition, processing, bioavailability and impacts on human health. Crit. Rev. Food Sci. Nutr. 2017, 57, 1280–1293. [Google Scholar] [CrossRef] [PubMed]
- Barreca, D.; Trombetta, D.; Smeriglio, A.; Mandalari, G.; Romeo, O.; Felice, M.R.; Gattuso, G.; Nabavi, S.M. Food flavonols: Nutraceuticals with complex health benefits and functionalities. Trends Food Sci. Tech. 2021, 117, 194–204. [Google Scholar] [CrossRef]
- Hostetler, G.L.; Ralston, R.A.; Schwartz, S.J. Flavones: Food Sources, Bioavailability, Metabolism, and Bioactivity. Adv. Nutr. 2017, 8, 423–435. [Google Scholar] [CrossRef] [Green Version]
- Khan, M.K.; Dangles, O. A comprehensive review on flavanones, the major citrus polyphenols. J. Food Compos. Anal. 2014, 33, 85–104. [Google Scholar] [CrossRef]
- Luo, Y.; Jian, Y.Q.; Liu, Y.K.; Jiang, S.; Muhammad, D.; Wang, W. Flavanols from Nature: A Phytochemistry and Biological Activity Review. Molecules 2022, 27, 719. [Google Scholar] [CrossRef]
- Abudureheman, B.; Yu, X.; Fang, D.; Zhang, H. Enzymatic Oxidation of Tea Catechins and Its Mechanism. Molecules 2022, 27, 942. [Google Scholar] [CrossRef]
- Khoo, H.E.; Azlan, A.; Tang, S.T.; Lim, S.M. Anthocyanidins and anthocyanins: Colored pigments as food, pharmaceutical ingredients, and the potential health benefits. Food Nutr. Res. 2017, 61, 1361779. [Google Scholar] [CrossRef] [Green Version]
- Kumar, N.; Goel, N. Phenolic acids: Natural versatile molecules with promising therapeutic applications. Biotechnol. Rep. 2019, 24, e00370. [Google Scholar] [CrossRef]
- Rodriguez-Garcia, C.; Sanchez-Quesada, C.; Toledo, E.; Delgado-Rodriguez, M.; Gaforio, J.J. Naturally Lignan-Rich Foods: A Dietary Tool for Health Promotion? Molecules 2019, 24, 917. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Teka, T.; Zhang, L.; Ge, X.; Li, Y.; Han, L.; Yan, X. Stilbenes: Source plants, chemistry, biosynthesis, pharmacology, application and problems related to their clinical Application-A comprehensive review. Phytochemistry 2022, 197, 113128. [Google Scholar] [CrossRef]
- Ramesh, T.N.; Paul, M.; Manikanta, K.; Girish, K.S. Structure and morphological studies of curcuminoids and curcuminoid mixture. J. Cryst. Growth 2020, 547, 125812. [Google Scholar] [CrossRef]
- D’Archivio, M.; Filesi, C.; Vari, R.; Scazzocchio, B.; Masella, R. Bioavailability of the Polyphenols: Status and Controversies. Int. J. Mol. Sci. 2010, 11, 1321–1342. [Google Scholar] [CrossRef] [PubMed]
- Day, A.J.; Canada, F.J.; Diaz, J.C.; Kroon, P.A.; Mclauchlan, R.; Faulds, C.B.; Plumb, G.W.; Morgan, M.R.A.; Williamson, G. Dietary flavonoid and isoflavone glycosides are hydrolysed by the lactase site of lactase phlorizin hydrolase. Febs Lett. 2000, 468, 166–170. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ray, S.K.; Mukherjee, S. Evolving Interplay Between Dietary Polyphenols and Gut Microbiota-An Emerging Importance in Healthcare. Front. Nutr. 2021, 8, 634944. [Google Scholar] [CrossRef] [PubMed]
- Nocito, M.C.; De Luca, A.; Prestia, F.; Avena, P.; La Padula, D.; Zavaglia, L.; Sirianni, R.; Casaburi, I.; Puoci, F.; Chimento, A.; et al. Antitumoral Activities of Curcumin and Recent Advances to ImProve Its Oral Bioavailability. Biomedicines 2021, 9, 1476. [Google Scholar] [CrossRef]
- Chimento, A.; De Amicis, F.; Sirianni, R.; Sinicropi, M.S.; Puoci, F.; Casaburi, I.; Saturnino, C.; Pezzi, V. Progress to Improve Oral Bioavailability and Beneficial Effects of Resveratrol. Int. J. Mol. Sci. 2019, 20, 1381. [Google Scholar] [CrossRef] [Green Version]
- Enaru, B.; Socaci, S.; Farcas, A.; Socaciu, C.; Danciu, C.; Stanila, A.; Diaconeasa, Z. Novel Delivery Systems of Polyphenols and Their Potential Health Benefits. Pharmaceuticals 2021, 14, 946. [Google Scholar] [CrossRef]
- Sharma, E.; Attri, D.C.; Sati, P.; Dhyani, P.; Szopa, A.; Sharifi-Rad, J.; Hano, C.; Calina, D.; Cho, W.C. Recent updates on anticancer mechanisms of polyphenols. Front. Cell Dev. Biol. 2022, 10, 1005910. [Google Scholar] [CrossRef]
- Suraweera, T.L.; Rupasinghe, H.P.V.; Dellaire, G.; Xu, Z.L. Regulation of Nrf2/ARE Pathway by Dietary Flavonoids: A Friend or Foe for Cancer Management? Antioxidants 2020, 9, 973. [Google Scholar] [CrossRef] [PubMed]
- Abotaleb, M.; Samuel, S.M.; Varghese, E.; Varghese, S.; Kubatka, P.; Liskova, A.; Busselberg, D. Flavonoids in Cancer and Apoptosis. Cancers 2019, 11, 28. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hazafa, A.; Iqbal, M.O.; Javaid, U.; Tareen, M.B.K.; Amna, D.; Ramzan, A.; Piracha, S.; Naeem, M. Inhibitory effect of polyphenols (phenolic acids, lignans, and stilbenes) on cancer by regulating signal transduction pathways: A review. Clin. Transl. Oncol. 2022, 24, 432–445. [Google Scholar] [CrossRef] [PubMed]
- Hazafa, A.; Rehman, K.U.; Jahan, N.; Jabeen, Z. The Role of Polyphenol (Flavonoids) Compounds in the Treatment of Cancer Cells. Nutr. Cancer 2020, 72, 386–397. [Google Scholar] [CrossRef] [PubMed]
- Elmore, S. Apoptosis: A review of programmed cell death. Toxicol. Pathol. 2007, 35, 495–516. [Google Scholar] [CrossRef] [PubMed]
- Hengartner, M.O. The biochemistry of apoptosis. Nature 2000, 407, 770–776. [Google Scholar] [CrossRef]
- Kale, J.; Osterlund, E.J.; Andrews, D.W. BCL-2 family proteins: Changing partners in the dance towards death. Cell Death Differ. 2018, 25, 65–80. [Google Scholar] [CrossRef] [Green Version]
- Chimento, A.; De Luca, A.; Avena, P.; De Amicis, F.; Casaburi, I.; Sirianni, R.; Pezzi, V. Estrogen Receptors-Mediated Apoptosis in Hormone-Dependent Cancers. Int. J. Mol. Sci. 2022, 23, 1242. [Google Scholar] [CrossRef]
- Fridman, J.S.; Lowe, S.W. Control of apoptosis by p53. Oncogene 2003, 22, 9030–9040. [Google Scholar] [CrossRef] [Green Version]
- Singh, K.K. Mitochondria damage checkpoint in apoptosis and genome stability. Fems Yeast Res. 2004, 5, 127–132. [Google Scholar] [CrossRef]
- Rapoport, B.L.; Anderson, R. Realizing the Clinical Potential of Immunogenic Cell Death in Cancer Chemotherapy and Radiotherapy. Int. J. Mol. Sci. 2019, 20, 959. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fan, Y.; Wang, M.; Li, Z.; Jiang, H.; Shi, J.; Shi, X.; Liu, S.; Zhao, J.; Kong, L.; Zhang, W.; et al. Intake of Soy, Soy Isoflavones and Soy Protein and Risk of Cancer Incidence and Mortality. Front. Nutr. 2022, 9, 847421. [Google Scholar] [CrossRef] [PubMed]
- Tuli, H.S.; Tuorkey, M.J.; Thakral, F.; Sak, K.; Kumar, M.; Sharma, A.K.; Sharma, U.; Jain, A.; Aggarwal, V.; Bishayee, A. Molecular Mechanisms of Action of Genistein in Cancer: Recent Advances. Front. Pharmacol. 2019, 10, 1336. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, L.; Rios, E.; Castro, L.; Liu, J.; Yan, Y.; Dixon, D. Genistein: Dual Role in Women’s Health. Nutrients 2021, 13, 3048. [Google Scholar] [CrossRef]
- Kuiper, G.G.; Lemmen, J.G.; Carlsson, B.; Corton, J.C.; Safe, S.H.; van der Saag, P.T.; van der Burg, B.; Gustafsson, J.A. Interaction of estrogenic chemicals and phytoestrogens with estrogen receptor beta. Endocrinology 1998, 139, 4252–4263. [Google Scholar] [CrossRef] [PubMed]
- Choi, E.J.; Jung, J.Y.; Kim, G.H. Genistein inhibits the proliferation and differentiation of MCF-7 and 3T3-L1 cells via the regulation of ER alpha expression and induction of apoptosis. Exp. Ther. Med. 2014, 8, 454–458. [Google Scholar] [CrossRef] [Green Version]
- Ullah, M.F.; Ahmad, A.; Zubair, H.; Khan, H.Y.; Wang, Z.W.; Sarkar, F.H.; Hadi, S.M. Soy isoflavone genistein induces cell death in breast cancer cells through mobilization of endogenous copper ions and generation of reactive oxygen species. Mol. Nutr. Food Res. 2011, 55, 553–559. [Google Scholar] [CrossRef]
- Chan, L.J.; Pang, Y.H.; Wang, Y.J.; Zhu, D.; Taledaohan, A.; Jia, Y.J.; Zhao, L.C.; Wang, W.J. Genistein-induced mitochondrial dysfunction and FOXO3a/PUMA expression in non-small lung cancer cells. Pharm. Biol. 2022, 60, 1876–1883. [Google Scholar] [CrossRef]
- Hsiao, Y.C.; Peng, S.F.; Lai, K.C.; Liao, C.L.; Huang, Y.P.; Lin, C.C.; Lin, M.L.; Liu, K.C.; Tsai, C.C.; Ma, Y.S.; et al. Genistein induces apoptosis in vitro and has antitumor activity against human leukemia HL-60 cancer cell xenograft growth in vivo. Environ. Toxicol. 2019, 34, 443–456. [Google Scholar] [CrossRef]
- Park, C.; Cha, H.J.; Lee, H.; Hwang-Bo, H.; Ji, S.Y.; Kim, M.Y.; Hong, S.H.; Jeong, J.W.; Han, M.H.; Choi, S.H.; et al. Induction of G2/M Cell Cycle Arrest and Apoptosis by Genistein in Human Bladder Cancer T24 Cells through Inhibition of the ROS-Dependent PI3k/Akt Signal Transduction Pathway. Antioxidants 2019, 8, 327. [Google Scholar] [CrossRef]
- Zhang, Q.; Bao, J.; Yang, J. Genistein-triggered anticancer activity against liver cancer cell line HepG2 involves ROS generation, mitochondrial apoptosis, G2/M cell cycle arrest and inhibition of cell migration. Arch. Med. Sci. 2019, 15, 1001–1009. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shafiee, G.; Saidijam, M.; Tavilani, H.; Ghasemkhani, N.; Khodadadi, I. Genistein Induces Apoptosis and Inhibits Proliferation of HT29 Colon Cancer Cells. Int. J. Mol. Cell. Med. 2016, 5, 178–191. [Google Scholar] [PubMed]
- Qin, J.; Teng, J.A.; Zhu, Z.; Chen, J.X.; Huang, W.J. Genistein induces activation of the mitochondrial apoptosis pathway by inhibiting phosphorylation of Akt in colorectal cancer cells. Pharm. Biol. 2016, 54, 74–79. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Solomon, L.A.; Ali, S.; Banerjee, S.; Munkarah, A.R.; Morris, R.T.; Sarkar, F.H. Sensitization of ovarian cancer cells to cisplatin by genistein: The role of NF-kappaB. J. Ovarian Res. 2008, 1, 9. [Google Scholar] [CrossRef] [Green Version]
- Szliszka, E.; Krol, W. Soy isoflavones augment the effect of TRAIL-mediated apoptotic death in prostate cancer cells. Oncol. Rep. 2011, 26, 533–541. [Google Scholar] [CrossRef] [PubMed]
- Dong, X.; Xu, W.Q.; Sikes, R.A.; Wu, C.Q. Combination of low dose of genistein and daidzein has synergistic preventive effects on isogenic human prostate cancer cells when compared with individual soy isoflavone. Food Chem. 2013, 141, 1923–1933. [Google Scholar] [CrossRef] [PubMed]
- Asgharian, P.; Tazekand, A.P.; Hosseini, K.; Forouhandeh, H.; Ghasemnejad, T.; Ranjbar, M.; Hasan, M.; Kumar, M.; Beirami, S.M.; Tarhriz, V.; et al. Potential mechanisms of quercetin in cancer prevention: Focus on cellular and molecular targets. Cancer Cell Int. 2022, 22, 257. [Google Scholar] [CrossRef]
- Akbari Kordkheyli, V.; Khonakdar Tarsi, A.; Mishan, M.A.; Tafazoli, A.; Bardania, H.; Zarpou, S.; Bagheri, A. Effects of quercetin on microRNAs: A mechanistic review. J. Cell. Biochem. 2019, 120, 12141–12155. [Google Scholar] [CrossRef] [PubMed]
- Duo, J.; Ying, G.G.; Wang, G.W.; Zhang, L. Quercetin inhibits human breast cancer cell proliferation and induces apoptosis via Bcl-2 and Bax regulation. Mol. Med. Rep. 2012, 5, 1453–1456. [Google Scholar] [CrossRef]
- Chou, C.C.; Yang, J.S.; Lu, H.F.; Ip, S.W.; Lo, C.; Wu, C.C.; Lin, J.P.; Tang, N.Y.; Chung, J.G.; Chou, M.J.; et al. Quercetin-mediated Cell Cycle Arrest and Apoptosis Involving Activation of a Caspase Cascade through the Mitochondria! Pathway in Human Breast Cancer MCF-7 Cells. Arch. Pharm. Res. 2010, 33, 1181–1191. [Google Scholar] [CrossRef]
- Niu, G.M.; Yin, S.M.; Xie, S.F.; Li, Y.Q.; Nie, D.N.; Ma, L.P.; Wang, X.J.; Wu, Y.D. Quercetin induces apoptosis by activating caspase-3 and regulating Bcl-2 and cyclooxygenase-2 pathways in human HL-60 cells. Acta Biochim. Biophys. Sin. 2011, 43, 30–37. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Farrag, I.M.; Belal, A.; Badawi, M.H.A.; Abdelhady, A.A.; Galala, F.; El-Sharkawy, A.; El-Dahshan, A.A.; Mehany, A.B.M. Antiproliferative, Apoptotic Effects and Suppression of Oxidative Stress of Quercetin against Induced Toxicity in Lung Cancer Cells of Rats: In vitro and In vivo Study. J. Cancer 2021, 12, 5249–5259. [Google Scholar] [CrossRef]
- Zhou, J.; Gong, J.; Ding, C.; Chen, G. Quercetin induces the apoptosis of human ovarian carcinoma cells by upregulating the expression of microRNA-145. Mol. Med. Rep. 2015, 12, 3127–3131. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gong, C.; Yang, Z.Y.; Zhang, L.Y.; Wang, Y.H.; Gong, W.; Liu, Y. Quercetin suppresses DNA double-strand break repair and enhances the radiosensitivity of human ovarian cancer cells via p53-dependent endoplasmic reticulum stress pathway. OncoTargets Ther. 2018, 11, 17–27. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seo, H.S.; Ku, J.M.; Choi, H.S.; Choi, Y.K.; Woo, J.K.; Kim, M.; Kim, I.; Na, C.H.; Hur, H.; Jang, B.H.; et al. Quercetin induces caspase-dependent extrinsic apoptosis through inhibition of signal transducer and activator of transcription 3 signaling in HER2-overexpressing BT-474 breast cancer cells. Oncol. Rep. 2016, 36, 31–42. [Google Scholar] [CrossRef] [Green Version]
- Kedhari Sundaram, M.; Raina, R.; Afroze, N.; Bajbouj, K.; Hamad, M.; Haque, S.; Hussain, A. Quercetin modulates signaling pathways and induces apoptosis in cervical cancer cells. Biosci. Rep. 2019, 39, BSR20190720. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Imran, M.; Aslam Gondal, T.; Atif, M.; Shahbaz, M.; Batool Qaisarani, T.; Hanif Mughal, M.; Salehi, B.; Martorell, M.; Sharifi-Rad, J. Apigenin as an anticancer agent. Phytother. Res. 2020, 34, 1812–1828. [Google Scholar] [CrossRef]
- Rahmani, A.H.; Alsahli, M.A.; Almatroudi, A.; Almogbel, M.A.; Khan, A.A.; Anwar, S.; Almatroodi, S.A. The Potential Role of Apigenin in Cancer Prevention and Treatment. Molecules 2022, 27, 6051. [Google Scholar] [CrossRef] [PubMed]
- Shukla, S.; Fu, P.F.; Gupta, S. Apigenin induces apoptosis by targeting inhibitor of apoptosis proteins and Ku70-Bax interaction in prostate cancer. Apoptosis 2014, 19, 883–894. [Google Scholar] [CrossRef]
- Shi, M.D.; Shiao, C.K.; Lee, Y.C.; Shih, Y.W. Apigenin, a dietary flavonoid, inhibits proliferation of human bladder cancer T-24 cells via blocking cell cycle progression and inducing apoptosis. Cancer Cell Int. 2015, 15, 33. [Google Scholar] [CrossRef]
- Woo, J.S.; Choo, G.S.; Yoo, E.S.; Kim, S.H.; Lee, J.H.; Han, S.H.; Kim, H.J.; Jung, S.H.; Park, Y.S.; Kim, B.S.; et al. Apigenin induces apoptosis by regulating Akt and MAPK pathways in human melanoma cell A375SM. Mol. Med. Rep. 2020, 22, 4877–4889. [Google Scholar] [CrossRef]
- Chan, L.P.; Chou, T.H.; Ding, H.Y.; Chen, P.R.; Chiang, F.Y.; Kuo, P.L.; Liang, C.H. Apigenin induces apoptosis via tumor necrosis factor receptor- and Bcl-2-mediated pathway and enhances susceptibility of head and neck squamous cell carcinoma to 5-fluorouracil and cisplatin. Biochim. Biophys. Acta-Gen. Subj. 2012, 1820, 1081–1091. [Google Scholar] [CrossRef] [PubMed]
- Masuelli, L.; Benvenuto, M.; Mattera, R.; Di Stefano, E.; Zago, E.; Taffera, G.; Tresoldi, I.; Giganti, M.G.; Frajese, G.V.; Berardi, G.; et al. In Vitro and In Vivo Anti-tumoral Effects of the Flavonoid Apigenin in Malignant Mesothelioma. Front. Pharmacol. 2017, 8, 373. [Google Scholar] [CrossRef] [Green Version]
- Sohel, M.; Sultana, H.; Sultana, T.; Al Amin, M.; Aktar, S.; Ali, M.C.; Rahim, Z.B.; Hossain, M.A.; Al Mamun, A.; Amin, M.N.; et al. Chemotherapeutic potential of hesperetin for cancer treatment, with mechanistic insights: A comprehensive review. Heliyon 2022, 8, e08815. [Google Scholar] [CrossRef]
- Zhang, J.X.; Wu, D.D.; Song, J.; Wang, J.; Yi, J.S.; Dong, W.G. Hesperetin Induces the Apoptosis of Gastric Cancer Cells via Activating Mitochondrial Pathway by Increasing Reactive Oxygen Species. Dig. Dis. Sci. 2015, 60, 2985–2995. [Google Scholar] [CrossRef] [PubMed]
- Wu, D.D.; Zhang, J.X.; Wang, J.; Li, J.; Liao, F.; Dong, W.G. Hesperetin induces apoptosis of esophageal cancer cells via mitochondrial pathway mediated by the increased intracellular reactive oxygen species. Tumor Biol. 2016, 37, 3451–3459. [Google Scholar] [CrossRef] [PubMed]
- Sivagami, G.; Vinothkumar, R.; Preethy, C.P.; Riyasdeen, A.; Akbarsha, M.A.; Menon, V.P.; Nalini, N. Role of hesperetin (a natural flavonoid) and its analogue on apoptosis in HT-29 human colon adenocarcinoma cell line—A comparative study. Food Chem. Toxicol. 2012, 50, 660–671. [Google Scholar] [CrossRef] [PubMed]
- Palit, S.; Kar, S.; Sharma, G.; Das, P.K. Hesperetin Induces Apoptosis in Breast Carcinoma by Triggering Accumulation of ROS and Activation of ASK1/JNK Pathway. J. Cell. Physiol. 2015, 230, 1729–1739. [Google Scholar] [CrossRef]
- Elango, R.; Athinarayanan, J.; Subbarayan, V.P.; Lei, D.K.Y.; Alshatwi, A.A. Hesperetin induces an apoptosis-triggered extrinsic pathway and a p53-independent pathway in human lung cancer H522 cells. J. Asian Nat. Prod. Res. 2018, 20, 559–569. [Google Scholar] [CrossRef]
- Tanaka, M.; Endo, H.; Sakusa, K.; Yano, M. Hesperetin induces apoptosis in A549 cells via the Hsp70-mediated activation of Bax. Int. J. Oncol. 2022, 61, 143. [Google Scholar] [CrossRef]
- Alshatwi, A.A.; Ramesh, E.; Periasamy, V.S.; Subash-Babu, P. The apoptotic effect of hesperetin on human cervical cancer cells is mediated through cell cycle arrest, death receptor, and mitochondrial pathways. Fundam. Clin. Pharmacol. 2013, 27, 581–592. [Google Scholar] [CrossRef] [PubMed]
- Prakash, M.; Basavaraj, B.V.; Murthy, K.N.C. Biological functions of epicatechin: Plant cell to human cell health. J. Funct. Foods 2019, 52, 14–24. [Google Scholar] [CrossRef]
- Pereyra-Vergara, F.; Olivares-Corichi, I.M.; Perez-Ruiz, A.G.; Luna-Arias, J.P.; Garcia-Sanchez, J.R. Apoptosis Induced by (-)-Epicatechin in Human Breast Cancer Cells is Mediated by Reactive Oxygen Species. Molecules 2020, 25, 1020. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, D.; Mollah, M.L.; Kim, K. Induction of Apoptosis of SW480 Human Colon Cancer Cells by (-)-Epicatechin Isolated from Bulnesia sarmienti. Anticancer Res. 2012, 32, 5353–5361. [Google Scholar]
- Thomas, P.; Dong, J. (-)-Epicatechin acts as a potent agonist of the membrane androgen receptor, ZIP9 (SLC39A9), to promote apoptosis of breast and prostate cancer cells. J. Steroid Biochem. 2021, 211, 105906. [Google Scholar] [CrossRef]
- Chen, J.L.; Xu, B.J.; Sun, J.X.; Jiang, X.W.; Bai, W.B. Anthocyanin supplement as a dietary strategy in cancer prevention and management: A comprehensive review. Crit. Rev. Food Sci. Nutr. 2022, 62, 7242–7254. [Google Scholar] [CrossRef]
- Sorrenti, V.; Vanella, L.; Acquaviva, R.; Cardile, V.; Giofre, S.; Di Giacomo, C. Cyanidin induces apoptosis and differentiation in prostate cancer cells. Int. J. Oncol. 2015, 47, 1303–1310. [Google Scholar] [CrossRef] [Green Version]
- Hosseini, M.M.; Karimi, A.; Behroozaghdam, M.; Javidi, M.A.; Ghiasvand, S.; Bereimipour, A.; Aryan, H.; Nassiri, F.; Jangholi, E. Cytotoxic and Apoptogenic Effects of Cyanidin-3-Glucoside on the Glioblastoma Cell Line. World Neurosurg. 2017, 108, 94–100. [Google Scholar] [CrossRef]
- Atashi, H.A.; Arani, H.Z.; Shekarriz, A.; Nazari, H.; Zabolian, A.; Rakhshan, R.; Olya, M. Cyanidin 3-O-Glucoside Induces the Apoptosis in the Osteosarcoma Cells through Upregulation of the PPAR gamma and P21: An In Vitro Study. Anticancer Agents Med. Chem. 2020, 20, 1087–1093. [Google Scholar] [CrossRef]
- Abotaleb, M.; Liskova, A.; Kubatka, P.; Busselberg, D. Therapeutic Potential of Plant Phenolic Acids in the Treatment of Cancer. Biomolecules 2020, 10, 221. [Google Scholar] [CrossRef] [Green Version]
- ElKhazendar, M.; Chalak, J.; El-Huneidi, W.; Vinod, A.; Abdel-Rahman, W.M.; Abu-Gharbieh, E. Antiproliferative and proapoptotic activities of ferulic acid in breast and liver cancer cell lines. Trop. J. Pharm. Res. 2019, 18, 2571–2576. [Google Scholar] [CrossRef]
- Cheng, W.J.; Zhang, P.P.; Luo, Q.Q.; Deng, S.M.; Jia, A.Q. The chemosensitizer ferulic acid enhances epirubicin-induced apoptosis in MDA-MB-231 cells. J. Funct. Foods 2020, 73, 104130. [Google Scholar] [CrossRef]
- Luo, L.P.; Zhu, S.H.; Tong, Y.; Peng, S.W. Ferulic Acid Induces Apoptosis of HeLa and Caski Cervical Carcinoma Cells by Down-Regulating the Phosphatidylinositol 3-Kinase (PI3K)/Akt Signaling Pathway. Med. Sci. Monit. 2020, 26, e920095. [Google Scholar] [CrossRef] [PubMed]
- Tsai, C.L.; Chiu, Y.M.; Ho, T.Y.; Hsieh, C.T.; Shieh, D.C.; Lee, Y.J.; Tsay, G.J.; Wu, Y.Y. Gallic Acid Induces Apoptosis in Human Gastric Adenocarcinoma Cells. Anticancer Res. 2018, 38, 2057–2067. [Google Scholar] [CrossRef] [PubMed]
- Yahya, S.A.; Hassan, F.A. Cytotoxic Effects Of Gallic Acid In Human Breast Cancer Cell Mda-Mb-231. J. Pharm. Negat. Result 2022, 13, 1337–1344. [Google Scholar] [CrossRef]
- Aborehab, N.M.; Elnagar, M.R.; Waly, N.E. Gallic acid potentiates the apoptotic effect of paclitaxel and carboplatin via overexpression of Bax and P53 on the MCF-7 human breast cancer cell line. J. Biochem. Mol. Toxicol. 2021, 35, e22638. [Google Scholar] [CrossRef] [PubMed]
- Mottaghi, S.; Abbaszadeh, H. A comprehensive mechanistic insight into the dietary and estrogenic lignans, arctigenin and sesamin as potential anticarcinogenic and anticancer agents. Current status, challenges, and future perspectives. Crit. Rev. Food Sci. Nutr. 2022, 62, 7301–7318. [Google Scholar] [CrossRef]
- Ozgocmen, M.; Bayram, D.; Turel, G.Y.; Togay, V.A.; Calapoglu, N.S. Secoisolariciresinol diglucoside induces caspase-3-mediated apoptosis in monolayer and spheroid cultures of human colon carcinoma cells. J. Food Biochem. 2021, 45, e13719. [Google Scholar] [CrossRef]
- Chen, J.; Saggar, J.K.; Corey, P.; Thompson, L.U. Flaxseed and Pure Secoisolariciresinol Diglucoside, but Not Flaxseed Hull, Reduce Human Breast Tumor Growth (MCF-7) in Athymic Mice. J. Nutr. 2009, 139, 2061–2066. [Google Scholar] [CrossRef] [Green Version]
- Sirerol, J.A.; Rodriguez, M.L.; Mena, S.; Asensi, M.A.; Estrela, J.M.; Ortega, A.L. Role of Natural Stilbenes in the Prevention of Cancer. Oxid. Med. Cell. Longev. 2016, 2016, 3128951. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.W.; Wang, L.J.; Wu, Y.P.; Lv, C.W.; Li, X.K.; Cao, X.R.; Yang, M.; Feng, D.P.; Luo, Z.J. Pterostilbene exerts antitumor activity against human osteosarcoma cells by inhibiting the JAK2/STAT3 signaling pathway. Toxicology 2013, 304, 120–131. [Google Scholar] [CrossRef]
- Alosi, J.A.; McDonald, D.E.; Schneider, J.S.; Privette, A.R.; McFadden, D.W. Pterostilbene Inhibits Breast Cancer In Vitro Through Mitochondrial Depolarization and Induction of Caspase-Dependent Apoptosis. J. Surg. Res. 2010, 161, 195–201. [Google Scholar] [CrossRef] [PubMed]
- Elsherbini, A.M.; Sheweita, S.A.; Sultan, A.S. Pterostilbene as a Phytochemical Compound Induces Signaling Pathways Involved in the Apoptosis and Death of Mutant P53-Breast Cancer Cell Lines. Nutr. Cancer 2021, 73, 1976–1984. [Google Scholar] [CrossRef] [PubMed]
- Wakimoto, R.; Ono, M.; Takeshima, M.; Higuchi, T.; Nakano, S. Differential Anticancer Activity of Pterostilbene Against Three Subtypes of Human Breast Cancer Cells. Anticancer Res. 2017, 37, 6153–6159. [Google Scholar] [CrossRef] [PubMed]
- Wen, W.; Lowe, G.; Roberts, C.M.; Finlay, J.; Han, E.S.; Glackin, C.A.; Dellinger, T.H. Pterostilbene, a natural phenolic compound, synergizes the antineoplastic effects of megestrol acetate in endometrial cancer. Sci. Rep. 2017, 7, 12754. [Google Scholar] [CrossRef] [Green Version]
- Shin, H.J.; Han, J.M.; Choi, Y.S.; Jung, H.J. Pterostilbene Suppresses both Cancer Cells and Cancer Stem-Like Cells in Cervical Cancer with Superior Bioavailability to Resveratrol. Molecules 2020, 25, 228. [Google Scholar] [CrossRef] [Green Version]
- Guo, L.Y.; Tan, K.; Wang, H.; Zhang, X. Pterostilbene inhibits hepatocellular carcinoma through p53/SOD2/ROS-mediated mitochondrial apoptosis. Oncol. Rep. 2016, 36, 3233–3240. [Google Scholar] [CrossRef] [Green Version]
- Degot, P.; Huber, V.; Touraud, D.; Kunz, W. Curcumin extracts from Curcuma Longa—Improvement of concentration, purity, and stability in food-approved and water-soluble surfactant-free microemulsions. Food Chem. 2021, 339, 128140. [Google Scholar] [CrossRef]
- Yang, Z.J.; Huang, S.Y.; Zhou, D.D.; Xiong, R.G.; Zhao, C.N.; Fang, A.P.; Zhang, Y.J.; Li, H.B.; Zhu, H.L. Effects and Mechanisms of Curcumin for the Prevention and Management of Cancers: An Updated Review. Antioxidants 2022, 11, 1481. [Google Scholar] [CrossRef]
- Shakor, A.B.A.; Atia, M.; Ismail, I.A.; Alshehri, A.; El-Refaey, H.; Kwiatkowska, K.; Sobota, A. Curcumin induces apoptosis of multidrug-resistant human leukemia HL60 cells by complex pathways leading to ceramide accumulation. Bba-Mol. Cell Biol. L. 2014, 1841, 1672–1682. [Google Scholar] [CrossRef]
- Yu, T.; Ji, J.; Guo, Y.L. MST1 activation by curcumin mediates JNK activation, Foxo3a nuclear translocation and apoptosis in melanoma cells. Biochem. Bioph. Res. Commun. 2013, 441, 53–58. [Google Scholar] [CrossRef] [PubMed]
- Park, S.; Cho, D.H.; Andera, L.; Suh, N.; Kim, I. Curcumin enhances TRAIL-induced apoptosis of breast cancer cells by regulating apoptosis-related proteins. Mol. Cell. Biochem. 2013, 383, 39–48. [Google Scholar] [CrossRef] [PubMed]
- Jung, E.M.; Park, J.W.; Choi, K.S.; Park, J.W.; Lee, H.I.L.; Lee, K.S.; Kwon, T.K. Curcumin sensitizes tumor necrosis factor-related apoptosis-inducing ligand (TRAIL)-mediated apoptosis through CHOP-independent DR5 upregulation. Carcinogenesis 2006, 27, 2008–2017. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kwon, D.; Oh, S.; Park, J.H.; Lee, S.; Lee, S. The TRAIL Sensitization Effect of Substituted Triazolyl Curcumin Mimics Against Brain Cancer Cells. Bull. Korean Chem. Soc. 2014, 35, 3339–3342. [Google Scholar] [CrossRef] [Green Version]
- Shankar, S.; Ganapathy, S.; Chen, Q.H.; Srivastava, R.K. Curcumin sensitizes TRAIL-resistant xenografts: Molecular mechanisms of apoptosis, metastasis and angiogenesis. Mol. Cancer 2008, 7, 16. [Google Scholar] [CrossRef] [Green Version]
- Wang, W.Z.; Li, L.; Liu, M.Y.; Jin, X.B.; Mao, J.W.; Pu, Q.H.; Meng, M.J.; Chen, X.G.; Zhu, J.Y. Curcumin induces FasL-related apoptosis through p38 activation in human hepatocellular carcinoma Huh7 cells. Life Sci. 2013, 92, 352–358. [Google Scholar] [CrossRef]
- Obaidi, I.; Blanco Fernandez, A.; McMorrow, T. Curcumin Sensitises Cancerous Kidney Cells to TRAIL Induced Apoptosis via Let-7C Mediated Deregulation of Cell Cycle Proteins and Cellular Metabolism. Int. J. Mol. Sci. 2022, 23, 9569. [Google Scholar] [CrossRef] [PubMed]
- Choudhuri, T.; Pal, S.; Agwarwal, M.L.; Das, T.; Sa, G. Curcumin induces apoptosis in human breast cancer cells through p53-dependent Bax induction. Febs Lett. 2002, 512, 334–340. [Google Scholar] [CrossRef] [Green Version]
- Mou, S.; Zhou, Z.; He, Y.; Liu, F.; Gong, L. Curcumin inhibits cell proliferation and promotes apoptosis of laryngeal cancer cells through Bcl-2 and PI3K/Akt, and by upregulating miR-15a. Oncol. Lett. 2017, 14, 4937–4942. [Google Scholar] [CrossRef] [Green Version]
- Zhou, Q.M.; Sun, Y.; Lu, Y.Y.; Zhang, H.; Chen, Q.L.; Su, S.B. Curcumin reduces mitomycin C resistance in breast cancer stem cells by regulating Bcl-2 family-mediated apoptosis. Cancer Cell Int. 2017, 17, 84. [Google Scholar] [CrossRef] [Green Version]
- Sha, J.; Li, J.; Wang, W.; Pan, L.; Cheng, J.; Li, L.; Zhao, H.; Lin, W. Curcumin induces G0/G1 arrest and apoptosis in hormone independent prostate cancer DU-145 cells by down regulating Notch signaling. Biomed. Pharmacother. 2016, 84, 177–184. [Google Scholar] [CrossRef] [PubMed]
- Bush, J.A.; Cheung, K.J., Jr.; Li, G. Curcumin induces apoptosis in human melanoma cells through a Fas receptor/caspase-8 pathway independent of p53. Exp. Cell. Res. 2001, 271, 305–314. [Google Scholar] [CrossRef] [PubMed]
- Aravindan, N.; Madhusoodhanan, R.; Ahmad, S.; Johnson, D.; Herman, T.S. Curcumin inhibits NFkappaB mediated radioprotection and modulate apoptosis related genes in human neuroblastoma cells. Cancer Biol. Ther. 2008, 7, 569–576. [Google Scholar] [CrossRef] [Green Version]
- Qiao, Q.; Jiang, Y.; Li, G. Inhibition of the PI3K/AKT-NF-kappaB pathway with curcumin enhanced radiation-induced apoptosis in human Burkitt’s lymphoma. J. Pharmacol. Sci. 2013, 121, 247–256. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, S.H.; Hang, L.W.; Yang, J.S.; Chen, H.Y.; Lin, H.Y.; Chiang, J.H.; Lu, C.C.; Yang, J.L.; Lai, T.Y.; Ko, Y.C.; et al. Curcumin induces apoptosis in human non-small cell lung cancer NCI-H460 cells through ER stress and caspase cascade- and mitochondria-dependent pathways. Anticancer Res. 2010, 30, 2125–2133. [Google Scholar]
- Sohn, E.J.; Bak, K.M.; Nam, Y.K.; Park, H.T. Upregulation of microRNA 344a-3p is involved in curcumin induced apoptosis in RT4 schwannoma cells. Cancer Cell Int. 2018, 18, 199. [Google Scholar] [CrossRef] [Green Version]
- Khan, A.Q.; Ahmed, E.I.; Elareer, N.; Fathima, H.; Prabhu, K.S.; Siveen, K.S.; Kulinski, M.; Azizi, F.; Dermime, S.; Ahmad, A.; et al. Curcumin-Mediated Apoptotic Cell Death in Papillary Thyroid Cancer and Cancer Stem-Like Cells through Targeting of the JAK/STAT3 Signaling Pathway. Int. J. Mol. Sci. 2020, 21, 438. [Google Scholar] [CrossRef] [Green Version]
- Vladu, A.F.; Ficai, D.; Ene, A.G.; Ficai, A. Combination Therapy Using Polyphenols: An Efficient Way to Improve Antitumoral Activity and Reduce Resistance. Int. J. Mol. Sci. 2022, 23, 10244. [Google Scholar] [CrossRef]
- Patra, S.; Pradhan, B.; Nayak, R.; Behera, C.; Das, S.; Patra, S.K.; Efferth, T.; Jena, M.; Bhutia, S.K. Dietary polyphenols in chemoprevention and synergistic effect in cancer: Clinical evidences and molecular mechanisms of action. Phytomedicine 2021, 90, 153554. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Lee, G.; Lee, J.I.; Ahn, T.G.; Kim, S.A. Effects of genistein on anti-tumor activity of cisplatin in human cervical cancer cell lines. Obstet. Gynecol. Sci. 2019, 62, 322–328. [Google Scholar] [CrossRef]
- Kaushik, S.; Shyam, H.; Agarwal, S.; Sharma, R.; Nag, T.C.; Dwivedi, A.K.; Balapure, A.K. Genistein potentiates Centchroman induced antineoplasticity in breast cancer via PI3K/Akt deactivation and ROS dependent induction of apoptosis. Life Sci. 2019, 239, 117073. [Google Scholar] [CrossRef] [PubMed]
- Xu, W.; Xie, S.; Chen, X.; Pan, S.; Qian, H.; Zhu, X. Effects of Quercetin on the Efficacy of Various Chemotherapeutic Drugs in Cervical Cancer Cells. Drug Des. Devel. Ther. 2021, 15, 577–588. [Google Scholar] [CrossRef] [PubMed]
- Mohammadi, E.; Alemi, F.; Maleki, M.; Malakoti, F.; Farsad-Akhtar, N.; Yousefi, B. Quercetin and Methotrexate in Combination have Anticancer Activity in Osteosarcoma Cells and Repress Oncogenic MicroRNA-223. Drug Res. 2022, 72, 226–233. [Google Scholar] [CrossRef] [PubMed]
- Xu, Z.X.; Zhao, D.M.; Zheng, X.Y.; Huang, B.; Xia, X.S.; Pan, X.J. Quercetin exerts bidirectional regulation effects on the efficacy of tamoxifen in estrogen receptor-positive breast cancer therapy: An in vitro study. Environ. Toxicol. 2020, 35, 1179–1193. [Google Scholar] [CrossRef] [PubMed]
- Hu, W.J.; Liu, J.; Zhong, L.K.; Wang, J. Apigenin enhances the antitumor effects of cetuximab in nasopharyngeal carcinoma by inhibiting EGFR signaling. Biomed. Pharmacother. 2018, 102, 681–688. [Google Scholar] [CrossRef]
- Liu, R.; Ji, P.; Liu, B.; Qiao, H.S.; Wang, X.; Zhou, L.K.; Deng, T.; Ba, Y. Apigenin enhances the cisplatin cytotoxic effect through p53-modulated apoptosis. Oncol. Lett. 2017, 13, 1024–1030. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, Y.M.; Xin, Y.Q.; Diao, Y.; Lu, C.Y.; Fu, J.; Luo, L.; Yin, Z.M. Synergistic Effects of Apigenin and Paclitaxel on Apoptosis of Cancer Cells. PLoS ONE 2011, 6, e29169. [Google Scholar] [CrossRef] [Green Version]
- Mahbub, A.A.; Le Maitre, C.L.; Cross, N.A.; Jordan-Mahy, N. The effect of apigenin and chemotherapy combination treatments on apoptosis-related genes and proteins in acute leukaemia cell lines. Sci. Rep. 2022, 12, 8858. [Google Scholar] [CrossRef]
- Mahbub, A.A.; Maitre, C.L.L.; Haywood-Small, S.; Cross, N.A.; Jordan-Mahy, N. Polyphenols enhance the activity of alkylating agents in leukaemia cell lines. Oncotarget 2019, 10, 4570–4586. [Google Scholar] [CrossRef] [Green Version]
- He, P.; Ma, J.; Liu, Y.; Deng, H.; Dong, W. Hesperetin Promotes Cisplatin-Induced Apoptosis of Gastric Cancer In Vitro and In Vivo by Upregulating PTEN Expression. Front. Pharmacol. 2020, 11, 1326. [Google Scholar] [CrossRef]
- Li, X.; Mu, J.; Lin, Y.; Zhao, J.; Meng, X. Combination of cyanidin-3-O-glucoside and cisplatin induces oxidative stress and apoptosis in HeLa cells by reducing activity of endogenous antioxidants, increasing bax/bcl-2 mRNA expression ratio, and downregulating Nrf2 expression. J. Food Biochem. 2021, 45, e13806. [Google Scholar] [CrossRef] [PubMed]
- Zhang, T.; Ma, L.; Wu, P.; Li, W.; Li, T.; Gu, R.; Dan, X.; Li, Z.; Fan, X.; Xiao, Z. Gallic acid has anticancer activity and enhances the anticancer effects of cisplatin in non--small cell lung cancer A549 cells via the JAK/STAT3 signaling pathway. Oncol. Rep. 2019, 41, 1779–1788. [Google Scholar] [CrossRef] [PubMed]
- Hsu, Y.H.; Chen, S.Y.; Wang, S.Y.; Lin, J.A.; Yen, G.C. Pterostilbene Enhances Cytotoxicity and Chemosensitivity in Human Pancreatic Cancer Cells. Biomolecules 2020, 10, 709. [Google Scholar] [CrossRef]
- Kostin, S.F.; McDonald, D.E.; McFadden, D.W. Inhibitory effects of (-)-epigallocatechin-3-gallate and pterostilbene on pancreatic cancer growth in vitro. J. Surg. Res. 2012, 177, 255–262. [Google Scholar] [CrossRef]
- Sivanantham, B.; Sethuraman, S.; Krishnan, U.M. Combinatorial Effects of Curcumin with an Anti-Neoplastic Agent on Head and Neck Squamous Cell Carcinoma Through the Regulation of EGFR-ERK1/2 and Apoptotic Signaling Pathways. Acs Comb. Sci. 2016, 18, 22–35. [Google Scholar] [CrossRef] [PubMed]
- Zhan, Y.Z.; Chen, Y.N.; Liu, R.; Zhang, H.; Zhang, Y.M. Potentiation of paclitaxel activity by curcumin in human breast cancer cell by modulating apoptosis and inhibiting EGFR signaling. Arch. Pharm. Res. 2014, 37, 1086–1095. [Google Scholar] [CrossRef]
- Calaf, G.M.; Ponce-Cusi, R.; Carrion, F. Curcumin and paclitaxel induce cell death in breast cancer cell lines. Oncol. Rep. 2018, 40, 2381–2388. [Google Scholar] [CrossRef]
- Dang, Y.P.; Yuan, X.Y.; Tian, R.; Li, D.G.; Liu, W. Curcumin improves the paclitaxel-induced apoptosis of HPV-positive human cervical cancer cells via the NF-kappa B-p53-caspase-3 pathway. Exp. Ther. Med. 2015, 9, 1470–1476. [Google Scholar] [CrossRef] [Green Version]
- Altundag, E.M.; Yilmaz, A.M.; Serdar, B.S.; Jannuzzi, A.T.; Kocturk, S.; Yalcin, A.S. Synergistic Induction of Apoptosis by Quercetin and Curcumin in Chronic Myeloid Leukemia (K562) Cells: II. Signal Transduction Pathways Involved. Nutr. Cancer 2021, 73, 703–712. [Google Scholar] [CrossRef]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chimento, A.; De Luca, A.; D’Amico, M.; De Amicis, F.; Pezzi, V. The Involvement of Natural Polyphenols in Molecular Mechanisms Inducing Apoptosis in Tumor Cells: A Promising Adjuvant in Cancer Therapy. Int. J. Mol. Sci. 2023, 24, 1680. https://doi.org/10.3390/ijms24021680
Chimento A, De Luca A, D’Amico M, De Amicis F, Pezzi V. The Involvement of Natural Polyphenols in Molecular Mechanisms Inducing Apoptosis in Tumor Cells: A Promising Adjuvant in Cancer Therapy. International Journal of Molecular Sciences. 2023; 24(2):1680. https://doi.org/10.3390/ijms24021680
Chicago/Turabian StyleChimento, Adele, Arianna De Luca, Maria D’Amico, Francesca De Amicis, and Vincenzo Pezzi. 2023. "The Involvement of Natural Polyphenols in Molecular Mechanisms Inducing Apoptosis in Tumor Cells: A Promising Adjuvant in Cancer Therapy" International Journal of Molecular Sciences 24, no. 2: 1680. https://doi.org/10.3390/ijms24021680