
Episodic Binge-like Ethanol Reduces Skeletal Muscle Strength Associated with Atrophy, Fibrosis, and Inflammation in Young Rats

 and
and 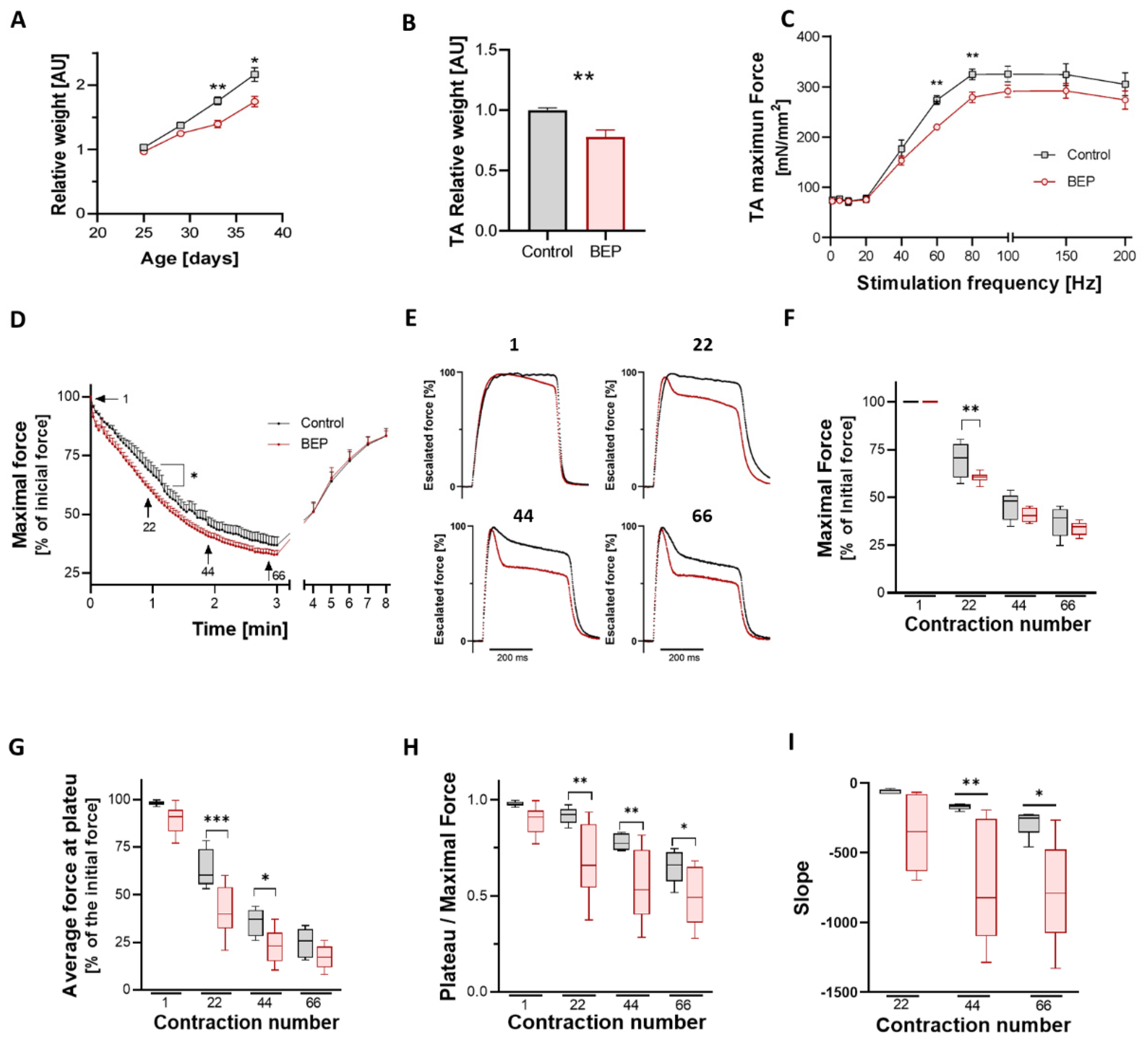{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Repetitive Binge-like Ethanol Administration Decreases TA Muscle Strength and Increases Muscle Fatigability
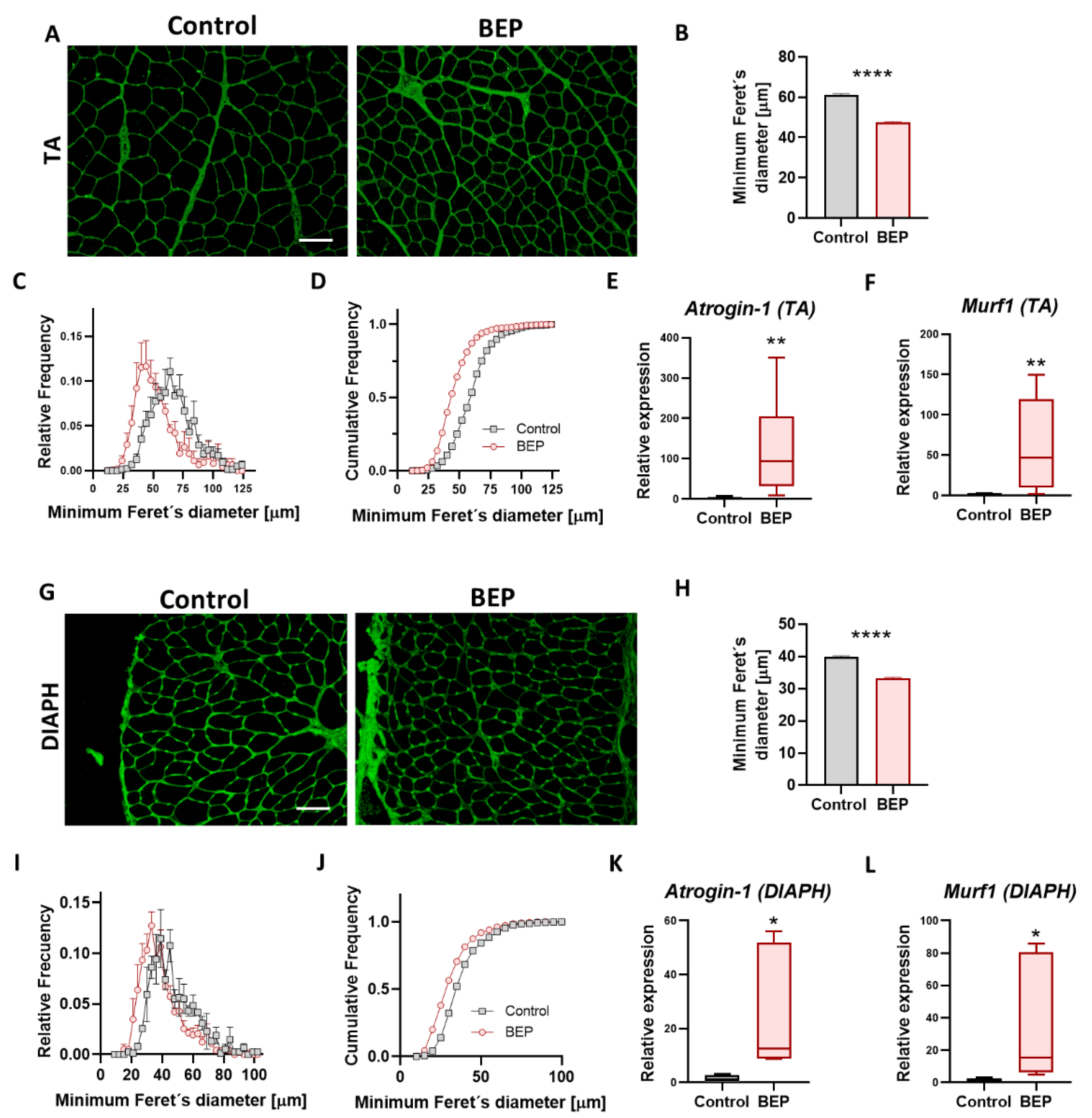
2.2. Repetitive Binge-like Ethanol Administration Causes a Reduction in Muscle Fiber Size
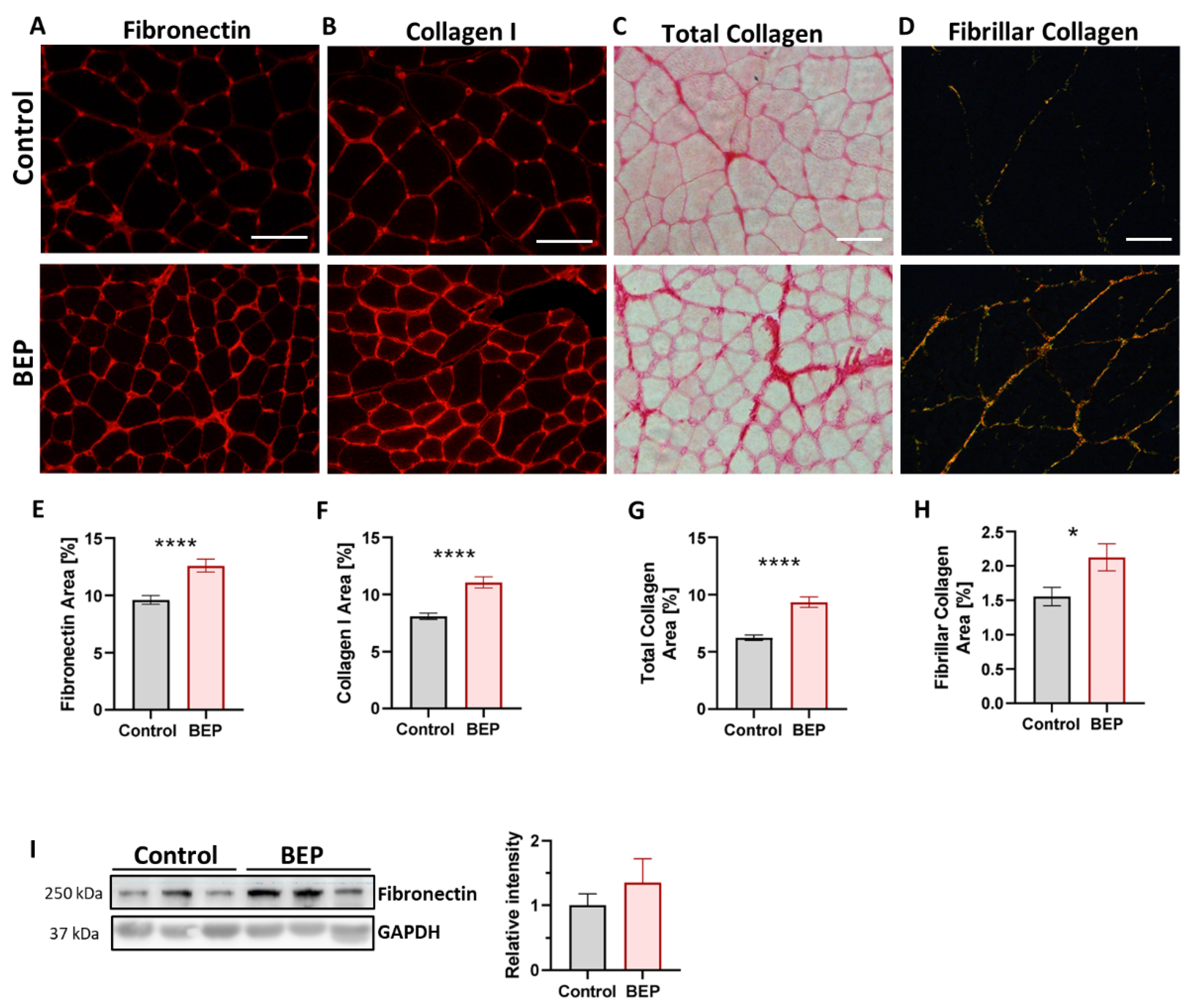
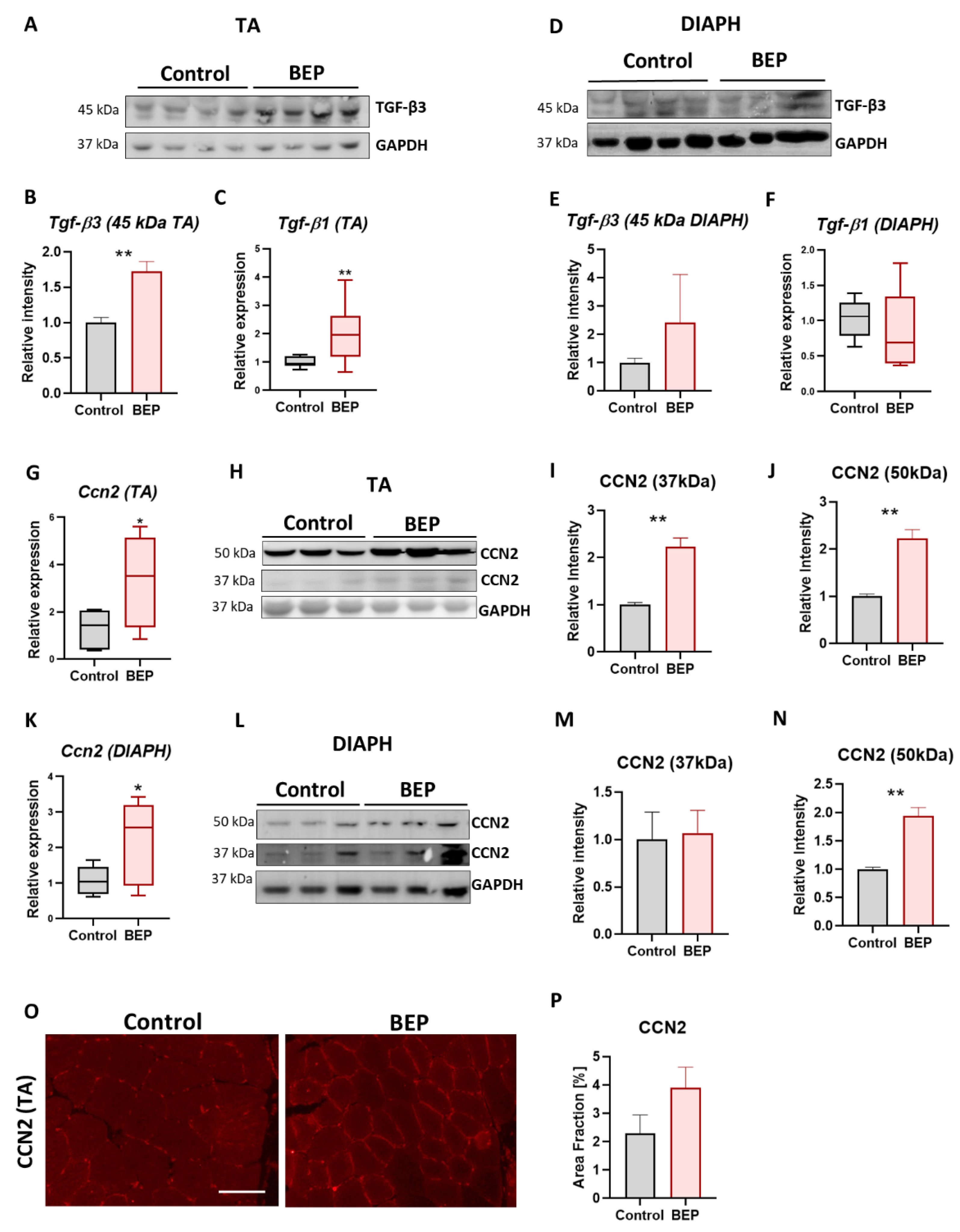
2.3. Exposure to Repetitive Binge-like Ethanol Leads to a Fibrotic Phenotype
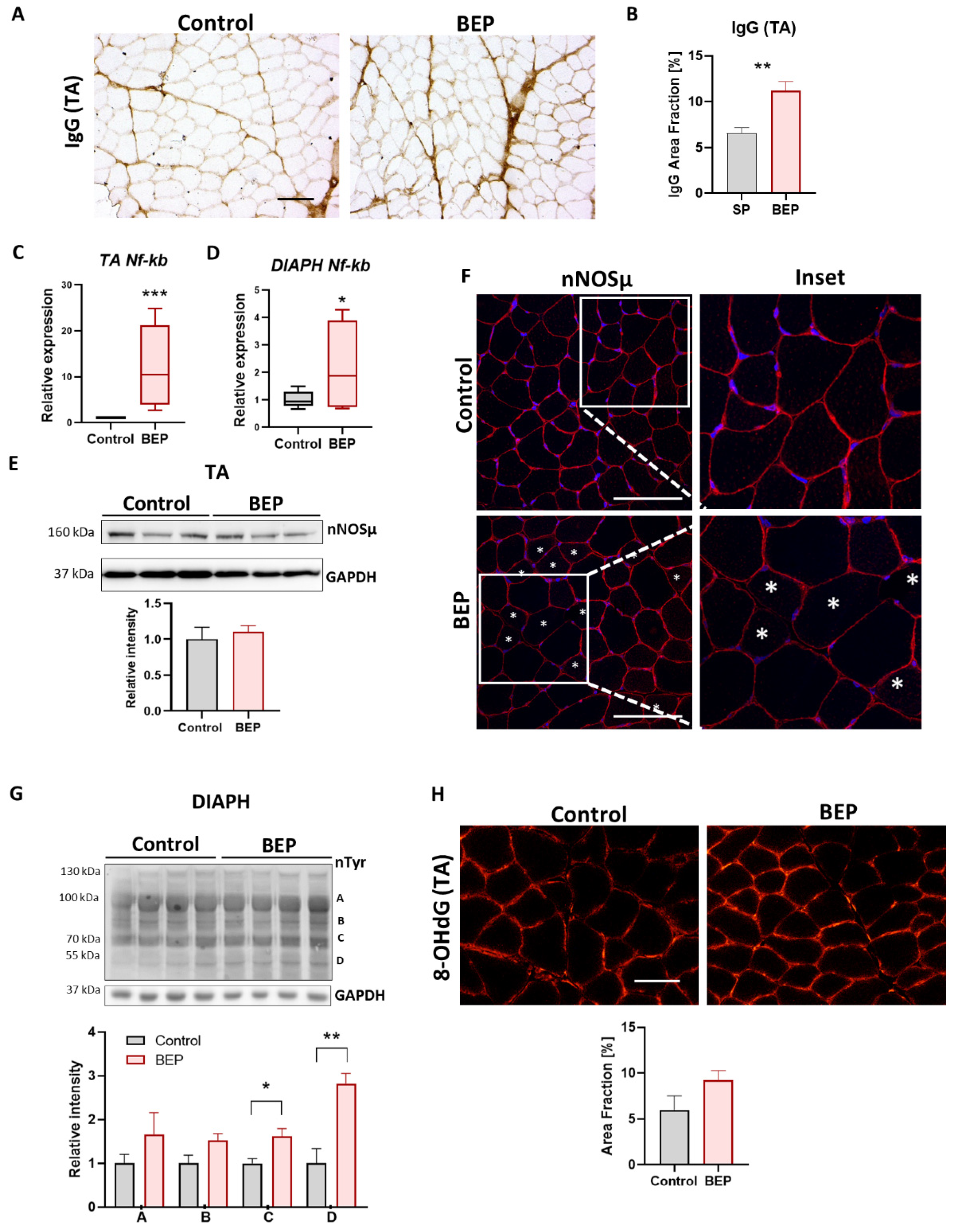
2.4. BEP-Treated Rats Exhibit Skeletal Muscle Pathological Markers
3. Discussion
Summary and Perspectives
4. Materials and Methods
4.1. Binge-like Ethanol Protocol in Rats
4.2. In Situ Analysis of Muscle Contractile Properties
4.3. Immunohistochemistry
4.4. Indirect Immunofluorescence
4.5. Sirius Red Staining
4.6. Determination of Occupied Area and Fiber Diameter
4.7. Determination of Fiber Type Percentage
4.8. Western Blot
4.9. RNA Isolation, Reverse Transcription, and Quantitative Real-Time PCR
4.10. Hematoxilin & Eosin Staining
4.11. Statistical Analyses
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hendriks, H.F.J. Alcohol and Human Health: What Is the Evidence? Annu. Rev. Food Sci. Technol. 2020, 11, 1–21. [Google Scholar] [CrossRef] [Green Version]
- WHO. Global Status Report on Alcohol and Health; World Health Organization: Geneva, Switzerland, 2018. [Google Scholar]
- Courtney, K.E.; Polich, J. Binge drinking in young adults: Data, definitions, and determinants. Psychol. Bull. 2009, 135, 142–156. [Google Scholar] [CrossRef] [Green Version]
- Kuntsche, E.; Kuntsche, S.; Thrul, J.; Gmel, G. Binge drinking: Health impact, prevalence, correlates and interventions. Psychol. Health 2017, 32, 976–1017. [Google Scholar] [CrossRef] [Green Version]
- Chung, T.; Creswell, K.G.; Bachrach, R.; Clark, D.B.; Martin, C.S. Adolescent Binge Drinking. Alcohol Res 2018, 39, 5–15. [Google Scholar] [PubMed]
- Chen, Y.; Houston, S. Risk Perception, Social Suppor, and Alcohol Use among U.S. Adolescents. Int. J. Commun. Health 2018, 13, 11–22. [Google Scholar]
- González Angulo, P.; Hernández Martínez, E.K.; Rodríguez Puente, L.A.; Castillo Vargas, R.; Salazar Mendoza, J.; Camacho Martínez, J.U. Percepción de riesgo ante el consumo de alcohol y tabaco en estudiantes de ciencias de la salud de Saltillo. Enfermería Glob. 2019, 18, 398–422. [Google Scholar] [CrossRef]
- Mendez-Ruiz, M.D.; Alonso-Castillo, M.T.D.J.; Alonso-Castillo, M.M.; Uribe-Alvarado, J.I.; Armendáriz-García, N.A. Relação de percepção de risco e consumo de álcool por adolescentes. SMAD. Rev. Eletrônica Saúde Ment. Álcool Drog. (Edição Em Port.) 2015, 11, 161. [Google Scholar] [CrossRef] [Green Version]
- Valencia Martin, J.L.; Gonzalez, M.J.; Galan, I. Methodological issues in the measurement of alcohol consumption: The importance of drinking patterns. Rev Esp Salud Publica 2014, 88, 433–446. [Google Scholar] [CrossRef] [Green Version]
- Chaiyasong, S.; Huckle, T.; Mackintosh, A.-M.; Meier, P.; Parry, C.D.H.; Callinan, S.; Viet Cuong, P.; Kazantseva, E.; Gray-Phillip, G.; Parker, K.; et al. Drinking patterns vary by gender, age and country-level income: Cross-country analysis of the International Alcohol Control Study. Drug Alcohol. Rev. 2018, 37, S53–S62. [Google Scholar] [CrossRef]
- Holahan, C.J.; Schutte, K.K.; Brennan, P.L.; Holahan, C.K.; Moos, R.H. Episodic heavy drinking and 20-year total mortality among late-life moderate drinkers. Alcohol. Clin. Exp. Res. 2014, 38, 1432–1438. [Google Scholar] [CrossRef]
- Vera, B.D.V.; Pilatti, A.; Pautassi, R.M. ELSA 2014 Cohort: Risk Factors Associated with Heavy Episodic Drinking Trajectories in Argentinean College Students. Front. Behav. Neurosci. 2020, 14, 105. [Google Scholar] [CrossRef] [PubMed]
- Green, K.; Nelson, T.F.; Hartmann, D. Binge drinking and sports participation in college: Patterns among athletes and former athletes. Int. Rev. Sociol. Sport 2014, 49, 417–434. [Google Scholar] [CrossRef] [Green Version]
- Caceres-Ayala, C.; Pautassi, R.M.; Acuña, M.J.; Cerpa, W.; Rebolledo, D.L. The functional and molecular effects of problematic alcohol consumption on skeletal muscle: A focus on athletic performance. Am. J. Drug Alcohol Abus. 2022, 48, 133–147. [Google Scholar] [CrossRef]
- O’Brien, K.S.; Kypri, K. Alcohol industry sponsorship and hazardous drinking among sportspeople. Addiction 2008, 103, 1961–1966. [Google Scholar] [CrossRef] [PubMed]
- O’Brien, K.S.; Forrest, W.; Greenlees, I.; Rhind, D.; Jowett, S.; Pinsky, I.; Espelt, A.; Bosque-Prous, M.; Sonderlund, A.L.; Vergani, M.; et al. Alcohol consumption, masculinity, and alcohol-related violence and anti-social behaviour in sportspeople. J. Sci. Med. Sport 2018, 21, 335–341. [Google Scholar] [CrossRef]
- O’Brien, K.S.; Ali, A.; Cotter, J.D.; O’Shea, R.P.; Stannard, S. Hazardous drinking in New Zealand sportspeople: Level of sporting participation and drinking motives. Alcohol Alcohol. 2007, 42, 376–382. [Google Scholar] [CrossRef] [Green Version]
- Preedy, V.R.; Ohlendieck, K.; Adachi, J.; Koll, M.; Sneddon, A.; Hunter, R.; Rajendram, R.; Mantle, D.; Peters, T.J. The importance of alcohol-induced muscle disease. J. Muscle Res. Cell Motil. 2003, 24, 55–63. [Google Scholar] [CrossRef]
- Simon, L.; Jolley, S.E.; Molina, P.E. Alcoholic Myopathy: Pathophysiologic Mechanisms and Clinical Implications. Alcohol. Res. 2017, 38, 207–217. [Google Scholar]
- Hanid, A.; Slavin, G.; Mair, W.; Sowter, C.; Ward, P.; Webb, J.; Levi, J. Fibre type changes in striated muscle of alcoholics. J. Clin. Pathol. 1981, 34, 991–995. [Google Scholar] [CrossRef] [Green Version]
- Zinovyeva, O.E.; Emelyanova, A.Y.; Samhaeva, N.D.; Sheglova, N.S.; Shenkman, B.S.; Nemirovskaya, T.L. Physiological basis of alcohol-induced skeletal muscle injury. Fiziol. Cheloveka 2016, 42, 130–136. [Google Scholar] [CrossRef]
- Fernandez-Sola, J.; Preedy, V.R.; Lang, C.H.; Gonzalez-Reimers, E.; Arno, M.; Lin, J.C.; Wiseman, H.; Zhou, S.; Emery, P.W.; Nakahara, T.; et al. Molecular and cellular events in alcohol-induced muscle disease. Alcohol. Clin. Exp. Res. 2007, 31, 1953–1962. [Google Scholar] [CrossRef]
- Molina, P.E.; Nelson, S. Binge Drinking’s Effects on the Body. Alcohol Res 2018, 39, 99–109. [Google Scholar]
- Crowell, K.T.; Laufenberg, L.J.; Lang, C.H. Chronic Alcohol Consumption, but not Acute Intoxication, Decreases In Vitro Skeletal Muscle Contractile Function. Alcohol. Clin. Exp. Res. 2019, 43, 2090–2099. [Google Scholar] [CrossRef]
- Laudato, J.A.; Tice, A.L.; Call, J.A.; Gordon, B.S.; Steiner, J.L. Effects of alcohol on skeletal muscle contractile performance in male and female mice. PLoS ONE 2021, 16, e0255946. [Google Scholar] [CrossRef] [PubMed]
- Pascual, M.; Blanco, A.M.; Cauli, O.; Miñarro, J.; Guerri, C. Intermittent ethanol exposure induces inflammatory brain damage and causes long-term behavioural alterations in adolescent rats. Eur. J. Neurosci. 2007, 25, 541–550. [Google Scholar] [CrossRef] [PubMed]
- Torres, A.K.; Tapia-Rojas, C.; Cerpa, W.; Quintanilla, R.A. Stimulation of Melanocortin Receptor-4 (MC4R) Prevents Mitochondrial Damage Induced by Binge Ethanol Protocol in Adolescent Rat Hippocampus. Neuroscience 2020, 438, 70–85. [Google Scholar] [CrossRef] [PubMed]
- Gómez, G.I.; Falcon, R.V.; Maturana, C.J.; Labra, V.C.; Salgado, N.; Rojas, C.A.; Oyarzun, J.E.; Cerpa, W.; Quintanilla, R.A.; Orellana, J.A. Heavy Alcohol Exposure Activates Astroglial Hemichannels and Pannexons in the Hippocampus of Adolescent Rats: Effects on Neuroinflammation and Astrocyte Arborization. Front. Cell. Neurosci. 2018, 12, 472. [Google Scholar] [CrossRef]
- Tapia-Rojas, C.; Carvajal, F.J.; Mira, R.G.; Arce, C.; Lerma-Cabrera, J.M.; Orellana, J.A.; Cerpa, W.; Quintanilla, R.A. Adolescent Binge Alcohol Exposure Affects the Brain Function Through Mitochondrial Impairment. Mol. Neurobiol. 2018, 55, 4473–4491. [Google Scholar] [CrossRef] [Green Version]
- Rebolledo, D.L.; Kim, M.J.; Whitehead, N.P.; Adams, M.E.; Froehner, S.C. Sarcolemmal targeting of nNOSμ improves contractile function of mdx muscle. Hum. Mol. Genet. 2016, 25, 158–166. [Google Scholar] [CrossRef] [Green Version]
- Adachi, J.; Asano, M.; Ueno, Y.; Niemelä, O.; Ohlendieck, K.; Peters, T.J.; Preedy, V.R. Alcoholic muscle disease and biomembrane perturbations (review). J. Nutr. Biochem. 2003, 14, 616–625. [Google Scholar] [CrossRef]
- Fanzani, A.; Conraads, V.M.; Penna, F.; Martinet, W. Molecular and cellular mechanisms of skeletal muscle atrophy: An update. J Cachexia Sarcopenia Muscle 2012, 3, 163–179. [Google Scholar] [CrossRef] [PubMed]
- Meng, S.J.; Yu, L.J. Oxidative stress, molecular inflammation and sarcopenia. Int. J. Mol. Sci. 2010, 11, 1509–1526. [Google Scholar] [CrossRef] [Green Version]
- von Haehling, S. Wasting away: How to treat cachexia and muscle wasting in chronic disease? Br. J. Clin. Pharmacol. 2017, 83, 2599–2601. [Google Scholar] [CrossRef] [PubMed]
- Schiaffino, S.; Dyar, K.A.; Ciciliot, S.; Blaauw, B.; Sandri, M. Mechanisms regulating skeletal muscle growth and atrophy. FEBS J. 2013, 280, 4294–4314. [Google Scholar] [CrossRef]
- Rebolledo, D.L.; Gonzalez, D.; Faundez-Contreras, J.; Contreras, O.; Vio, C.P.; Murphy-Ullrich, J.E.; Lipson, K.E.; Brandan, E. Denervation-induced skeletal muscle fibrosis is mediated by CTGF/CCN2 independently of TGF-beta. Matrix Biol. J. Int. Soc. Matrix Biol. 2019, 82, 20–37. [Google Scholar] [CrossRef]
- Gonzalez, D.; Contreras, O.; Rebolledo, D.L.; Espinoza, J.P.; van Zundert, B.; Brandan, E. ALS skeletal muscle shows enhanced TGF-beta signaling, fibrosis and induction of fibro/adipogenic progenitor markers. PLoS ONE 2017, 12, e0177649. [Google Scholar] [CrossRef] [Green Version]
- Bodine, S.C.; Baehr, L.M. Skeletal muscle atrophy and the E3 ubiquitin ligases MuRF1 and MAFbx/atrogin-1. Am. J. Physiol. Endocrinol. Metab. 2014, 307, E469–E484. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bonaldo, P.; Sandri, M. Cellular and molecular mechanisms of muscle atrophy. Dis. Model. Mech. 2013, 6, 25–39. [Google Scholar] [CrossRef] [Green Version]
- Foletta, V.C.; White, L.J.; Larsen, A.E.; Leger, B.; Russell, A.P. The role and regulation of MAFbx/atrogin-1 and MuRF1 in skeletal muscle atrophy. Pflug. Arch. 2011, 461, 325–335. [Google Scholar] [CrossRef]
- Rom, O.; Reznick, A.Z. The role of E3 ubiquitin-ligases MuRF-1 and MAFbx in loss of skeletal muscle mass. Free Radic. Biol. Med. 2016, 98, 218–230. [Google Scholar] [CrossRef]
- Sandri, M.; Sandri, C.; Gilbert, A.; Skurk, C.; Calabria, E.; Picard, A.; Walsh, K.; Schiaffino, S.; Lecker, S.H.; Goldberg, A.L. Foxo transcription factors induce the atrophy-related ubiquitin ligase atrogin-1 and cause skeletal muscle atrophy. Cell 2004, 117, 399–412. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yin, L.; Li, N.; Jia, W.; Wang, N.; Liang, M.; Yang, X.; Du, G. Skeletal muscle atrophy: From mechanisms to treatments. Pharm. Res 2021, 172, 105807. [Google Scholar] [CrossRef]
- Pette, D.; Staron, R.S. Myosin isoforms, muscle fiber types, and transitions. Microsc. Res. Tech. 2000, 50, 500–509. [Google Scholar] [CrossRef] [PubMed]
- Ciciliot, S.; Rossi, A.C.; Dyar, K.A.; Blaauw, B.; Schiaffino, S. Muscle type and fiber type specificity in muscle wasting. Int. J. Biochem. Cell Biol. 2013, 45, 2191–2199. [Google Scholar] [CrossRef]
- Evans, W.J.; Lexell, J. Human Aging, Muscle Mass, and Fiber Type Composition. J. Gerontol. Ser. A 1995, 50, 11–16. [Google Scholar] [CrossRef] [PubMed]
- Zierath, J.R.; Hawley, J.A. Skeletal Muscle Fiber Type: Influence on Contractile and Metabolic Properties. PLoS Biol. 2004, 2, e348. [Google Scholar] [CrossRef]
- Jiménez-Fuentes, M.A.; Gea, J.; Mariñán, M.; Gáldiz, J.B.; Gallego, F.; Broquetas, J.M. [Cellular homogeneity in diverse portions of the diaphragm]. Arch. Bronconeumol. 1998, 34, 82–86. [Google Scholar] [CrossRef] [PubMed]
- Rebolledo, D.L.; Lipson, K.E.; Brandan, E. Driving fibrosis in neuromuscular diseases: Role and regulation of Connective tissue growth factor (CCN2/CTGF). Matrix Biol. Plus 2021, 11, 100059. [Google Scholar] [CrossRef] [PubMed]
- Wynn, T.A.; Ramalingam, T.R. Mechanisms of fibrosis: Therapeutic translation for fibrotic disease. Nat. Med. 2012, 18, 1028–1040. [Google Scholar] [CrossRef] [Green Version]
- Serrano, A.L.; Munoz-Canoves, P. Regulation and dysregulation of fibrosis in skeletal muscle. Exp. Cell Res. 2010, 316, 3050–3058. [Google Scholar] [CrossRef]
- Mahdy, M.A.A. Skeletal muscle fibrosis: An overview. Cell Tissue Res. 2019, 375, 575–588. [Google Scholar] [CrossRef]
- Serrano, A.L.; Muñoz-Cánoves, P. Fibrosis development in early-onset muscular dystrophies: Mechanisms and translational implications. Semin. Cell Dev. Biol. 2017, 64, 181–190. [Google Scholar] [CrossRef]
- Song, Y.; Yao, S.; Liu, Y.; Long, L.; Yang, H.; Li, Q.; Liang, J.; Li, X.; Lu, Y.; Zhu, H.; et al. Expression levels of TGF-β1 and CTGF are associated with the severity of Duchenne muscular dystrophy. Exp. Ther. Med. 2017, 13, 1209–1214. [Google Scholar] [CrossRef]
- Gonzalez, D.; Rebolledo, D.L.; Correa, L.M.; Court, F.A.; Cerpa, W.; Lipson, K.E.; van Zundert, B.; Brandan, E. The inhibition of CTGF/CCN2 activity improves muscle and locomotor function in a murine ALS model. Hum. Mol. Genet. 2018, 27, 2913–2926. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rittié, L. Method for Picrosirius Red-Polarization Detection of Collagen Fibers in Tissue Sections. Methods Mol. Biol. 2017, 1627, 395–407. [Google Scholar] [CrossRef] [PubMed]
- Taglietti, V.; Kefi, K.; Bronisz-Budzyńska, I.; Mirciloglu, B.; Rodrigues, M.; Cardone, N.; Coulpier, F.; Periou, B.; Gentil, C.; Goddard, M.; et al. Duchenne muscular dystrophy trajectory in R-DMDdel52 preclinical rat model identifies COMP as biomarker of fibrosis. Acta Neuropathol. Commun. 2022, 10, 60. [Google Scholar] [CrossRef] [PubMed]
- Ishizaki, M.; Suga, T.; Kimura, E.; Shiota, T.; Kawano, R.; Uchida, Y.; Uchino, K.; Yamashita, S.; Maeda, Y.; Uchino, M. Mdx respiratory impairment following fibrosis of the diaphragm. Neuromuscul. Disord. 2008, 18, 342–348. [Google Scholar] [CrossRef]
- Stedman, H.H.; Sweeney, H.L.; Shrager, J.B.; Maguire, H.C.; Panettieri, R.A.; Petrof, B.; Narusawa, M.; Leferovich, J.M.; Sladky, J.T.; Kelly, A.M. The mdx mouse diaphragm reproduces the degenerative changes of Duchenne muscular dystrophy. Nature 1991, 352, 536–539. [Google Scholar] [CrossRef]
- Rebolledo, D.L.; Acuna, M.J.; Brandan, E. Role of Matricellular CCN Proteins in Skeletal Muscle: Focus on CCN2/CTGF and Its Regulation by Vasoactive Peptides. Int. J. Mol. Sci. 2021, 22, 5234. [Google Scholar] [CrossRef]
- Leask, A.; Parapuram, S.K.; Shi-Wen, X.; Abraham, D.J. Connective tissue growth factor (CTGF, CCN2) gene regulation: A potent clinical bio-marker of fibroproliferative disease? J. Cell Commun. Signal. 2009, 3, 89–94. [Google Scholar] [CrossRef] [Green Version]
- Ramazani, Y.; Knops, N.; Elmonem, M.A.; Nguyen, T.Q.; Arcolino, F.O.; Van Den Heuvel, L.; Levtchenko, E.; Kuypers, D.; Goldschmeding, R. Connective tissue growth factor (CTGF) from basics to clinics. Matrix Biol. 2018, 68–69, 44–66. [Google Scholar] [CrossRef]
- Steiner, J.L.; Pruznak, A.M.; Navaratnarajah, M.; Lang, C.H. Alcohol Differentially Alters Extracellular Matrix and Adhesion Molecule Expression in Skeletal Muscle and Heart. Alcohol. Clin. Exp. Res. 2015, 39, 1330–1340. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dodd, T.; Simon, L.; LeCapitaine, N.J.; Zabaleta, J.; Mussell, J.; Berner, P.; Ford, S.; Dufour, J.; Bagby, G.J.; Nelson, S.; et al. Chronic binge alcohol administration accentuates expression of pro-fibrotic and inflammatory genes in the skeletal muscle of simian immunodeficiency virus-infected macaques. Alcohol. Clin. Exp. Res. 2014, 38, 2697–2706. [Google Scholar] [CrossRef]
- Levitt, D.E.; Yeh, A.Y.; Prendergast, M.J.; Budnar, J.R.G.; Adler, K.A.; Cook, G.; Molina, P.E.; Simon, L. Chronic Alcohol Dysregulates Skeletal Muscle Myogenic Gene Expression after Hind Limb Immobilization in Female Rats. Biomolecules 2020, 10, 441. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dekeyser, G.J.; Clary, C.R.; Otis, J.S. Chronic alcohol ingestion delays skeletal muscle regeneration following injury. Regen. Med. Res. 2013, 1, 2. [Google Scholar] [CrossRef] [Green Version]
- Messina, S.; Vita, G.L.; Aguennouz, M.; Sframeli, M.; Romeo, S.; Rodolico, C.; Vita, G. Activation of NF-kappaB pathway in Duchenne muscular dystrophy: Relation to age. Acta Myol. 2011, 30, 16–23. [Google Scholar]
- Yang, Q.; Tang, Y.; Imbrogno, K.; Lu, A.; Proto, J.D.; Chen, A.; Guo, F.; Fu, F.H.; Huard, J.; Wang, B. AAV-based shRNA silencing of NF-κB ameliorates muscle pathologies in mdx mice. Gene Ther. 2012, 19, 1196–1204. [Google Scholar] [CrossRef] [Green Version]
- Cabrera, D.; Gutiérrez, J.; Cabello-Verrugio, C.; Morales, M.G.; Mezzano, S.; Fadic, R.; Casar, J.C.; Hancke, J.L.; Brandan, E. Andrographolide attenuates skeletal muscle dystrophy in mdx mice and increases efficiency of cell therapy by reducing fibrosis. Skelet. Muscle 2014, 4, 6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thoma, A.; Lightfoot, A.P. NF-kB and Inflammatory Cytokine Signalling: Role in Skeletal Muscle Atrophy. Adv. Exp. Med. Biol. 2018, 1088, 267–279. [Google Scholar] [CrossRef]
- Fang, W.Y.; Tseng, Y.T.; Lee, T.Y.; Fu, Y.C.; Chang, W.H.; Lo, W.W.; Lin, C.L.; Lo, Y.C. Triptolide prevents LPS-induced skeletal muscle atrophy via inhibiting NF-κB/TNF-α and regulating protein synthesis/degradation pathway. Br. J. Pharmacol. 2021, 178, 2998–3016. [Google Scholar] [CrossRef]
- Peterson, J.M.; Bakkar, N.; Guttridge, D.C. Chapter four—NF-κB Signaling in Skeletal Muscle Health and Disease. In Current Topics in Developmental Biology; Pavlath, G.K., Ed.; Academic Press: Cambridge, MA, USA, 2011; Volume 96, pp. 85–119. [Google Scholar]
- Li, H.; Malhotra, S.; Kumar, A. Nuclear factor-kappa B signaling in skeletal muscle atrophy. J. Mol. Med. 2008, 86, 1113–1126. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thomas, G.D.; Shaul, P.W.; Yuhanna, I.S.; Froehner, S.C.; Adams, M.E. Vasomodulation by skeletal muscle-derived nitric oxide requires alpha-syntrophin-mediated sarcolemmal localization of neuronal Nitric oxide synthase. Circ. Res. 2003, 92, 554–560. [Google Scholar] [CrossRef] [Green Version]
- Brenman, J.E.; Chao, D.S.; Xia, H.; Aldape, K.; Bredt, D.S. Nitric oxide synthase complexed with dystrophin and absent from skeletal muscle sarcolemma in Duchenne muscular dystrophy. Cell 1995, 82, 743–752. [Google Scholar] [CrossRef] [Green Version]
- Suzuki, N.; Mizuno, H.; Warita, H.; Takeda, S.; Itoyama, Y.; Aoki, M. Neuronal NOS is dislocated during muscle atrophy in amyotrophic lateral sclerosis. J. Neurol. Sci. 2010, 294, 95–101. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, N.; Motohashi, N.; Uezumi, A.; Fukada, S.; Yoshimura, T.; Itoyama, Y.; Aoki, M.; Miyagoe-Suzuki, Y.; Takeda, S. NO production results in suspension-induced muscle atrophy through dislocation of neuronal NOS. J. Clin. Investig. 2007, 117, 2468–2476. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kobayashi, Y.M.; Rader, E.P.; Crawford, R.W.; Iyengar, N.K.; Thedens, D.R.; Faulkner, J.A.; Parikh, S.V.; Weiss, R.M.; Chamberlain, J.S.; Moore, S.A.; et al. Sarcolemma-localized nNOS is required to maintain activity after mild exercise. Nature 2008, 456, 511–515. [Google Scholar] [CrossRef] [Green Version]
- Percival, J.M. nNOS regulation of skeletal muscle fatigue and exercise performance. Biophys. Rev. 2011, 3, 9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, J.; Chu, H.; Zhao, H.; Cheng, X.; Liu, Y.; Jin, W.; Zhao, J.; Liu, B.; Ding, Y.; Ma, H. Nitricoxide synthase-induced oxidative stress in prolonged alcoholic myopathies of rats. Mol. Cell. Biochem. 2007, 304, 135–142. [Google Scholar] [CrossRef] [PubMed]
- Beckendorf, L.; Linke, W.A. Emerging importance of oxidative stress in regulating striated muscle elasticity. J. Muscle Res. Cell Motil. 2015, 36, 25–36. [Google Scholar] [CrossRef] [Green Version]
- Jang, Y.C.; Rodriguez, K.; Lustgarten, M.S.; Muller, F.L.; Bhattacharya, A.; Pierce, A.; Choi, J.J.; Lee, N.H.; Chaudhuri, A.; Richardson, A.G.; et al. Superoxide-mediated oxidative stress accelerates skeletal muscle atrophy by synchronous activation of proteolytic systems. Geroscience 2020, 42, 1579–1591. [Google Scholar] [CrossRef]
- Powers, S.K.; Morton, A.B.; Ahn, B.; Smuder, A.J. Redox control of skeletal muscle atrophy. Free Radic. Biol. Med. 2016, 98, 208–217. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salanova, M.; Schiffl, G.; Gutsmann, M.; Felsenberg, D.; Furlan, S.; Volpe, P.; Clarke, A.; Blottner, D. Nitrosative stress in human skeletal muscle attenuated by exercise countermeasure after chronic disuse. Redox Biol 2013, 1, 514–526. [Google Scholar] [CrossRef] [PubMed]
- Avin, K.G.; Chen, N.X.; Organ, J.M.; Zarse, C.; O’Neill, K.; Conway, R.G.; Konrad, R.J.; Bacallao, R.L.; Allen, M.R.; Moe, S.M. Skeletal Muscle Regeneration and Oxidative Stress Are Altered in Chronic Kidney Disease. PLoS ONE 2016, 11, e0159411. [Google Scholar] [CrossRef] [Green Version]
- Souza, L.B.; Maziero, C.; Lazzarin, M.C.; Quintana, H.T.; Tome, T.C.; Baptista, V.I.A.; de Oliveira, F. Presence of metalloproteinases 2 and 9 and 8-OHdG in the fibrotic process in skeletal muscle of Mdx mice. Acta Histochem. 2020, 122, 151458. [Google Scholar] [CrossRef] [PubMed]
- Yu, J.; Lü, G.; Xu, J. [The effects of ethanol on neuromuscular junctions of adult toad]. Hua Xi Yi Ke Da Xue Xue Bao = J. West China Univ. Med. Sci. = Huaxi Yike Daxue Xuebao 2001, 32, 274–276. [Google Scholar]
- Prentice, C.; Stannard, S.R.; Barnes, M.J. Effects of heavy episodic drinking on physical performance in club level rugby union players. J. Sci. Med. Sport 2015, 18, 268–271. [Google Scholar] [CrossRef] [PubMed]
- Forcina, L.; Cosentino, M.; Musarò, A. Mechanisms Regulating Muscle Regeneration: Insights into the Interrelated and Time-Dependent Phases of Tissue Healing. Cells 2020, 9, 1297. [Google Scholar] [CrossRef]
- Vary, T.C.; Frost, R.A.; Lang, C.H. Acute alcohol intoxication increases atrogin-1 and MuRF1 mRNA without increasing proteolysis in skeletal muscle. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2008, 294, R1777–R1789. [Google Scholar] [CrossRef] [Green Version]
- Reed, C.H.; Buhr, T.J.; Tystahl, A.C.; Bauer, E.E.; Clark, P.J.; Valentine, R.J. The effects of voluntary binge-patterned ethanol ingestion and daily wheel running on signaling of muscle protein synthesis and degradation in female mice. Alcohol 2022, 104, 45–52. [Google Scholar] [CrossRef]
- Tice, A.L.; Laudato, J.A.; Fadool, D.A.; Gordon, B.S.; Steiner, J.L. Acute binge alcohol alters whole body metabolism and the time-dependent expression of skeletal muscle-specific metabolic markers for multiple days in mice. Am. J. Physiol. Endocrinol. Metab. 2022, 323, E215–E230. [Google Scholar] [CrossRef]
- Otis, J.S.; Brown, L.A.; Guidot, D.M. Oxidant-induced atrogin-1 and transforming growth factor-beta1 precede alcohol-related myopathy in rats. Muscle Nerve 2007, 36, 842–848. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Otis, J.S.; Guidot, D.M. Procysteine increases alcohol-depleted glutathione stores in rat plantaris following a period of abstinence. Alcohol Alcohol. 2010, 45, 495–500. [Google Scholar] [CrossRef] [Green Version]
- Gumucio, J.P.; Mendias, C.L. Atrogin-1, MuRF-1, and sarcopenia. Endocrine 2013, 43, 12–21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barbe, M.F.; Hilliard, B.A.; Amin, M.; Harris, M.Y.; Hobson, L.J.; Cruz, G.E.; Popoff, S.N. Blocking CTGF/CCN2 reduces established skeletal muscle fibrosis in a rat model of overuse injury. FASEB J. 2020, 34, 6554–6569. [Google Scholar] [CrossRef] [Green Version]
- Lipson, K.E.; Wong, C.; Teng, Y.; Spong, S. CTGF is a central mediator of tissue remodeling and fibrosis and its inhibition can reverse the process of fibrosis. Fibrogenesis Tissue Repair. 2012, 5, S24. [Google Scholar] [CrossRef]
- Zhou, J.; Sun, C.; Yang, L.; Wang, J.; Jn-Simon, N.; Zhou, C.; Bryant, A.; Cao, Q.; Li, C.; Petersen, B.; et al. Liver regeneration and ethanol detoxification: A new link in YAP regulation of ALDH1A1 during alcohol-related hepatocyte damage. FASEB J. 2022, 36, e22224. [Google Scholar] [CrossRef]
- Robinson, P.M.; Smith, T.S.; Patel, D.; Dave, M.; Lewin, A.S.; Pi, L.; Scott, E.W.; Tuli, S.S.; Schultz, G.S. Proteolytic Processing of Connective Tissue Growth Factor in Normal Ocular Tissues and during Corneal Wound Healing. Investig. Opthalmol. Vis. Sci. 2012, 53, 8093. [Google Scholar] [CrossRef] [Green Version]
- Kaasbøll, O.J.; Gadicherla, A.K.; Wang, J.H.; Monsen, V.T.; Hagelin, E.M.V.; Dong, M.Q.; Attramadal, H. Connective tissue growth factor (CCN2) is a matricellular preproprotein controlled by proteolytic activation. J. Biol. Chem. 2018, 293, 17953–17970. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Córdova, G.; Rochard, A.; Riquelme-Guzmán, C.; Cofré, C.; Scherman, D.; Bigey, P.; Brandan, E. SMAD3 and SP1/SP3 Transcription Factors Collaborate to Regulate Connective Tissue Growth Factor Gene Expression in Myoblasts in Response to Transforming Growth Factor β. J. Cell. Biochem. 2015, 116, 1880–1887. [Google Scholar] [CrossRef] [PubMed]
- Györfi, A.H.; Matei, A.E.; Distler, J.H.W. Targeting TGF-β signaling for the treatment of fibrosis. Matrix Biol. J. Int. Soc. Matrix Biol. 2018, 68–69, 8–27. [Google Scholar] [CrossRef]
- Leask, A.; Abraham, D.J. TGF-beta signaling and the fibrotic response. FASEB J. 2004, 18, 816–827. [Google Scholar] [CrossRef] [PubMed]
- Valle-Tenney, R.; Rebolledo, D.L.; Lipson, K.E.; Brandan, E. Role of hypoxia in skeletal muscle fibrosis: Synergism between hypoxia and TGF-β signaling upregulates CCN2/CTGF expression specifically in muscle fibers. Matrix Biol. J. Int. Soc. Matrix Biol. 2020, 87, 48–65. [Google Scholar] [CrossRef] [PubMed]
- Valle-Tenney, R.; Rebolledo, D.; Acuña, M.J.; Brandan, E. HIF-hypoxia signaling in skeletal muscle physiology and fibrosis. J. Cell Commun. Signal. 2020, 14, 147–158. [Google Scholar] [CrossRef]
- Gallardo, F.S.; Córdova-Casanova, A.; Brandan, E. The linkage between inflammation and fibrosis in muscular dystrophies: The axis autotaxin-lysophosphatidic acid as a new therapeutic target? J. Cell Commun. Signal. 2021, 15, 317–334. [Google Scholar] [CrossRef] [PubMed]
- Córdova-Casanova, A.; Cruz-Soca, M.; Chun, J.; Casar, J.C.; Brandan, E. Activation of the ATX/LPA/LPARs axis induces a fibrotic response in skeletal muscle. Matrix Biol. J. Int. Soc. Matrix Biol. 2022, 109, 121–139. [Google Scholar] [CrossRef] [PubMed]
- Cabello-Verrugio, C.; Córdova, G.; Vial, C.; Zúñiga, L.M.; Brandan, E. Connective tissue growth factor induction by lysophosphatidic acid requires transactivation of transforming growth factor type β receptors and the JNK pathway. Cell. Signal. 2011, 23, 449–457. [Google Scholar] [CrossRef]
- Huang, S.; Liu, Z.; Qian, X.; Li, L.; Zhang, H.; Li, S.; Liu, Z. YAP/TAZ Promote Fibrotic Activity in Human Trabecular Meshwork Cells by Sensing Cytoskeleton Structure Alternation. Chemosensors 2022, 10, 235. [Google Scholar] [CrossRef]
- Di Benedetto, A.; Mottolese, M.; Sperati, F.; Ercolani, C.; Di Lauro, L.; Pizzuti, L.; Vici, P.; Terrenato, I.; Sperduti, I.; Shaaban, A.M.; et al. The Hippo transducers TAZ/YAP and their target CTGF in male breast cancer. Oncotarget 2016, 7, 43188–43198. [Google Scholar] [CrossRef] [Green Version]
- Ni, H.M.; Bhakta, A.; Wang, S.; Li, Z.; Manley, S.; Huang, H.; Copple, B.; Ding, W.X. Role of hypoxia inducing factor-1β in alcohol-induced autophagy, steatosis and liver injury in mice. PLoS ONE 2014, 9, e115849. [Google Scholar] [CrossRef]
- Zelickson, B.R.; Benavides, G.A.; Johnson, M.S.; Chacko, B.K.; Venkatraman, A.; Landar, A.; Betancourt, A.M.; Bailey, S.M.; Darley-Usmar, V.M. Nitric oxide and hypoxia exacerbate alcohol-induced mitochondrial dysfunction in hepatocytes. Biochim. Biophys. Acta 2011, 1807, 1573–1582. [Google Scholar] [CrossRef] [Green Version]
- Arteel, G.E.; Iimuro, Y.; Yin, M.; Raleigh, J.A.; Thurman, R.G. Chronic enteral ethanol treatment causes hypoxia in rat liver tissue in vivo. Hepatology 1997, 25, 920–926. [Google Scholar] [CrossRef]
- Nath, B.; Levin, I.; Csak, T.; Petrasek, J.; Mueller, C.; Kodys, K.; Catalano, D.; Mandrekar, P.; Szabo, G. Hepatocyte-specific hypoxia-inducible factor-1α is a determinant of lipid accumulation and liver injury in alcohol-induced steatosis in mice. Hepatology 2011, 53, 1526–1537. [Google Scholar] [CrossRef] [Green Version]
- Foitzik, T.; Fernández-del Castillo, C.; Rattner, D.W.; Klar, E.; Warshaw, A.L. Alcohol selectively impairs oxygenation of the pancreas. Arch. Surg. 1995, 130, 357–360; discussion 361. [Google Scholar] [CrossRef] [PubMed]
- McKim, S.E.; Uesugi, T.; Raleigh, J.A.; McClain, C.J.; Arteel, G.E. Chronic intragastric alcohol exposure causes hypoxia and oxidative stress in the rat pancreas. Arch. Biochem. Biophys. 2003, 417, 34–43. [Google Scholar] [CrossRef] [PubMed]
- Morris, N.; Yeligar, S. Role of HIF-1α in Alcohol-Mediated Multiple Organ Dysfunction. Biomolecules 2018, 8, 170. [Google Scholar] [CrossRef] [PubMed]
- Percival, J.M.; Anderson, K.N.; Gregorevic, P.; Chamberlain, J.S.; Froehner, S.C. Functional deficits in nNOSmu-deficient skeletal muscle: Myopathy in nNOS knockout mice. PLoS ONE 2008, 3, e3387. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.; Yang, H.T.; Wasala, L.; Zhang, K.; Yue, Y.; Duan, D.; Lai, Y. Dystrophin R16/17 protein therapy restores sarcolemmal nNOS in trans and improves muscle perfusion and function. Mol. Med. 2019, 25, 31. [Google Scholar] [CrossRef]
- Tapia-Rojas, C.; Mira, R.G.; Torres, A.K.; Jara, C.; Pérez, M.J.; Vergara, E.H.; Cerpa, W.; Quintanilla, R.A. Alcohol consumption during adolescence: A link between mitochondrial damage and ethanol brain intoxication. Birth Defects Res 2017, 109, 1623–1639. [Google Scholar] [CrossRef]
- Tapia-Rojas, C.; Torres, A.K.; Quintanilla, R.A. Adolescence binge alcohol consumption induces hippocampal mitochondrial impairment that persists during the adulthood. Neuroscience 2019, 406, 356–368. [Google Scholar] [CrossRef]
- Acuña, M.J.; Salas, D.; Córdova-Casanova, A.; Cruz-Soca, M.; Céspedes, C.; Vio, C.P.; Brandan, E. Blockade of Bradykinin receptors worsens the dystrophic phenotype of mdx mice: Differential effects for B1 and B2 receptors. J. Cell Commun. Signal. 2018, 12, 589–601. [Google Scholar] [CrossRef]
- Morales, M.G.; Cabello-Verrugio, C.; Santander, C.; Cabrera, D.; Goldschmeding, R.; Brandan, E. CTGF/CCN-2 over-expression can directly induce features of skeletal muscle dystrophy. J. Pathol. 2011, 225, 490–501. [Google Scholar] [CrossRef] [PubMed]
- Morales, M.G.; Gutierrez, J.; Cabello-Verrugio, C.; Cabrera, D.; Lipson, K.E.; Goldschmeding, R.; Brandan, E. Reducing CTGF/CCN2 slows down mdx muscle dystrophy and improves cell therapy. Hum. Mol. Genet. 2013, 22, 4938–4951. [Google Scholar] [CrossRef] [PubMed]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cáceres-Ayala, C.; Mira, R.G.; Acuña, M.J.; Brandan, E.; Cerpa, W.; Rebolledo, D.L. Episodic Binge-like Ethanol Reduces Skeletal Muscle Strength Associated with Atrophy, Fibrosis, and Inflammation in Young Rats. Int. J. Mol. Sci. 2023, 24, 1655. https://doi.org/10.3390/ijms24021655
Cáceres-Ayala C, Mira RG, Acuña MJ, Brandan E, Cerpa W, Rebolledo DL. Episodic Binge-like Ethanol Reduces Skeletal Muscle Strength Associated with Atrophy, Fibrosis, and Inflammation in Young Rats. International Journal of Molecular Sciences. 2023; 24(2):1655. https://doi.org/10.3390/ijms24021655
Chicago/Turabian StyleCáceres-Ayala, Constanza, Rodrigo G. Mira, María José Acuña, Enrique Brandan, Waldo Cerpa, and Daniela L. Rebolledo. 2023. "Episodic Binge-like Ethanol Reduces Skeletal Muscle Strength Associated with Atrophy, Fibrosis, and Inflammation in Young Rats" International Journal of Molecular Sciences 24, no. 2: 1655. https://doi.org/10.3390/ijms24021655