
Differential Regulation of Innate Lymphoid Cells in Human and Murine Oral Squamous Cell Carcinoma
 ,
,
Abstract
:1. Introduction
2. Results
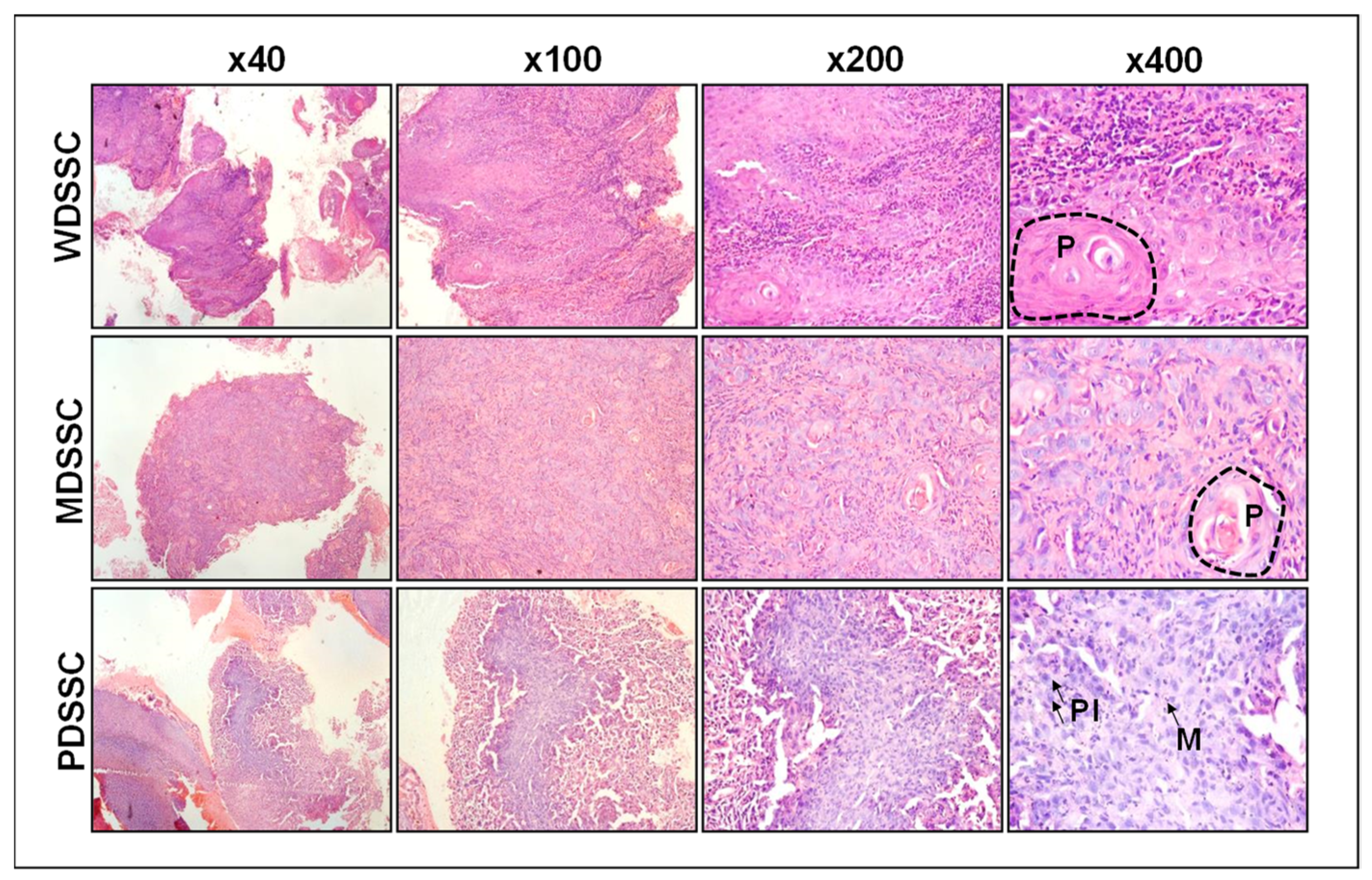
2.1. Clinicopathologic Characteristics of Human Oral Squamous Cell Carcinoma
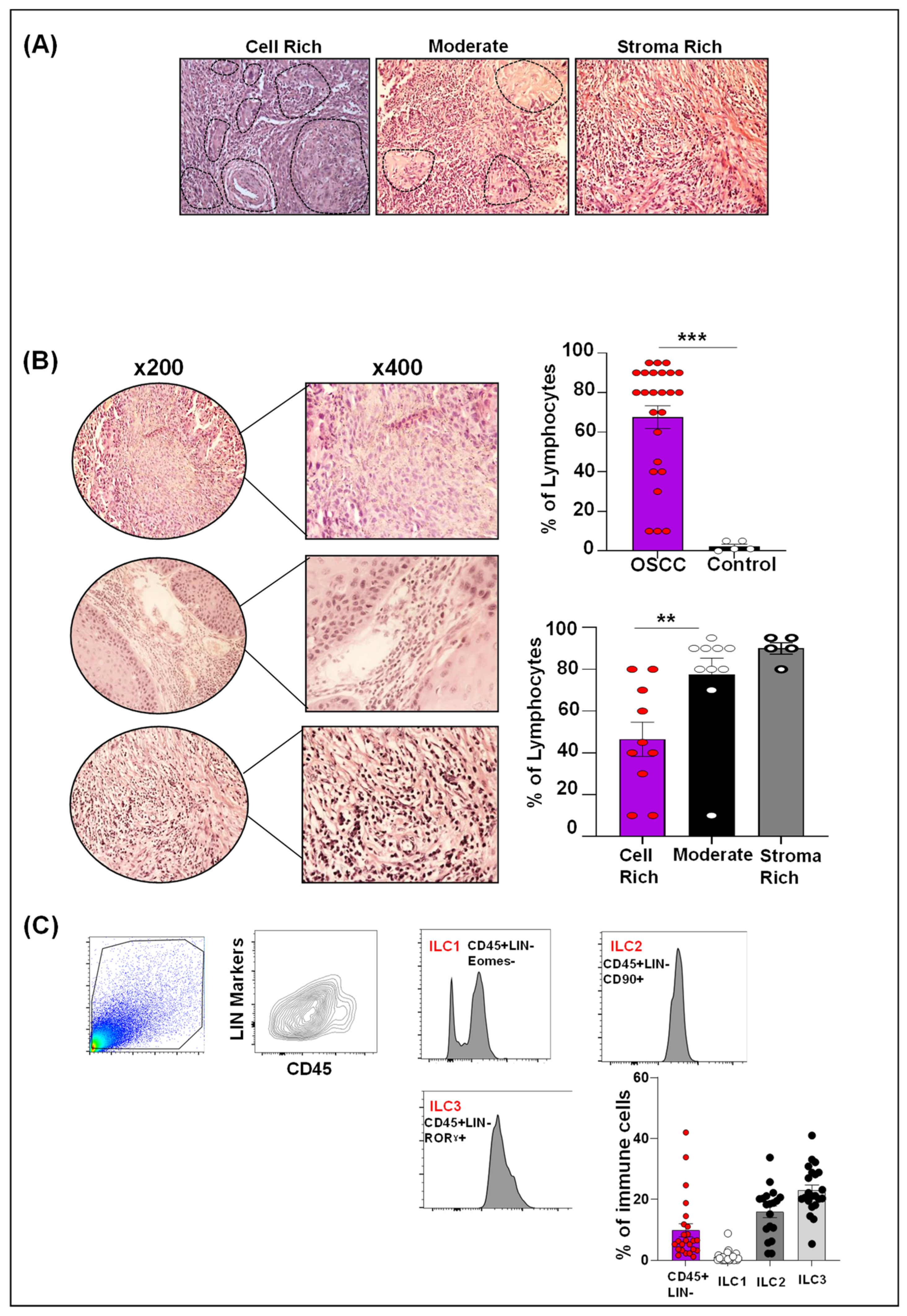
2.2. Human OSCC Microenvironment Exhibit Increased Lymphocytic Infiltrate
2.3. Differential ILCs Expansion in Human OSCC
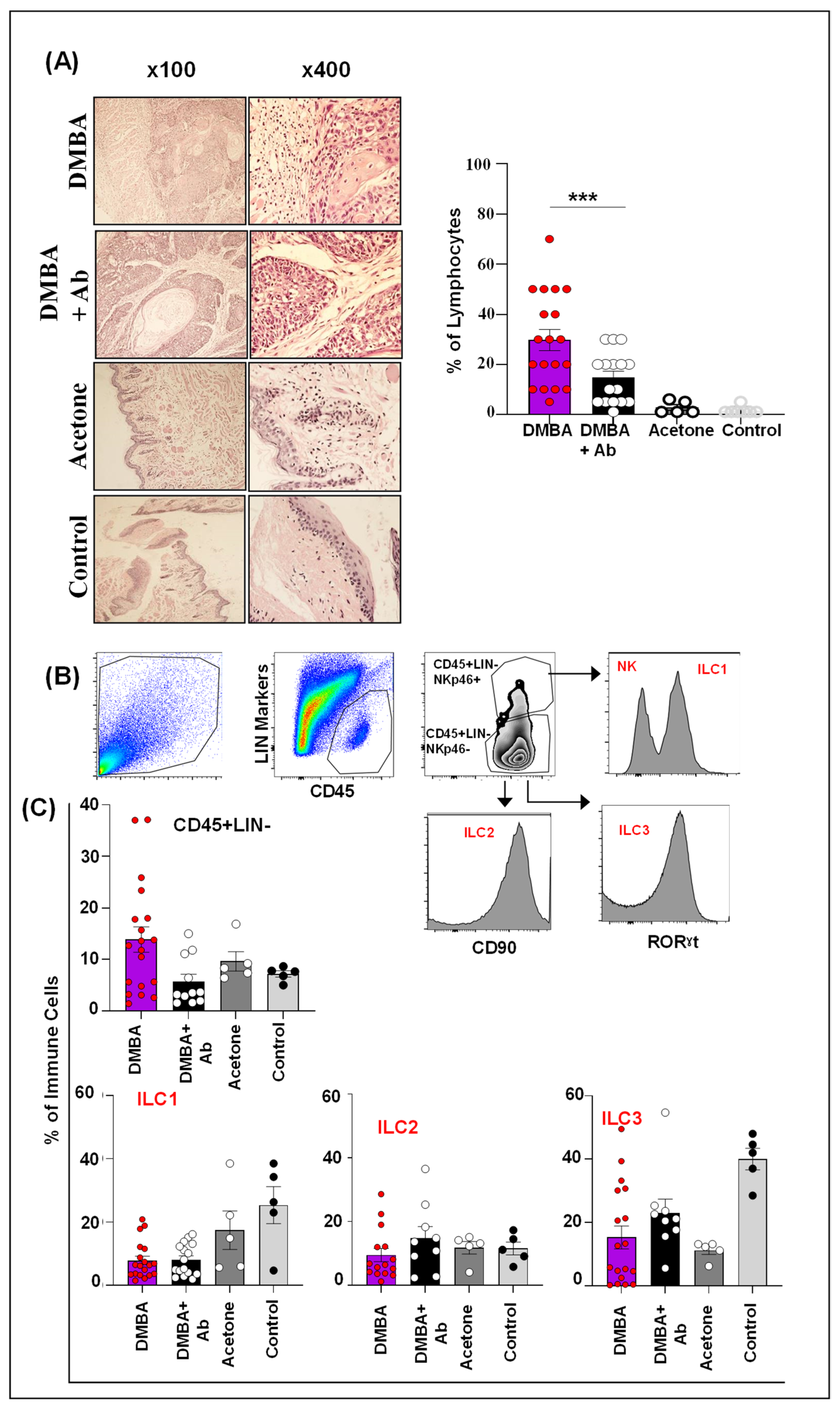
2.4. ILC1 and ILC3 Are Deregulated in Murine Model of OSCC
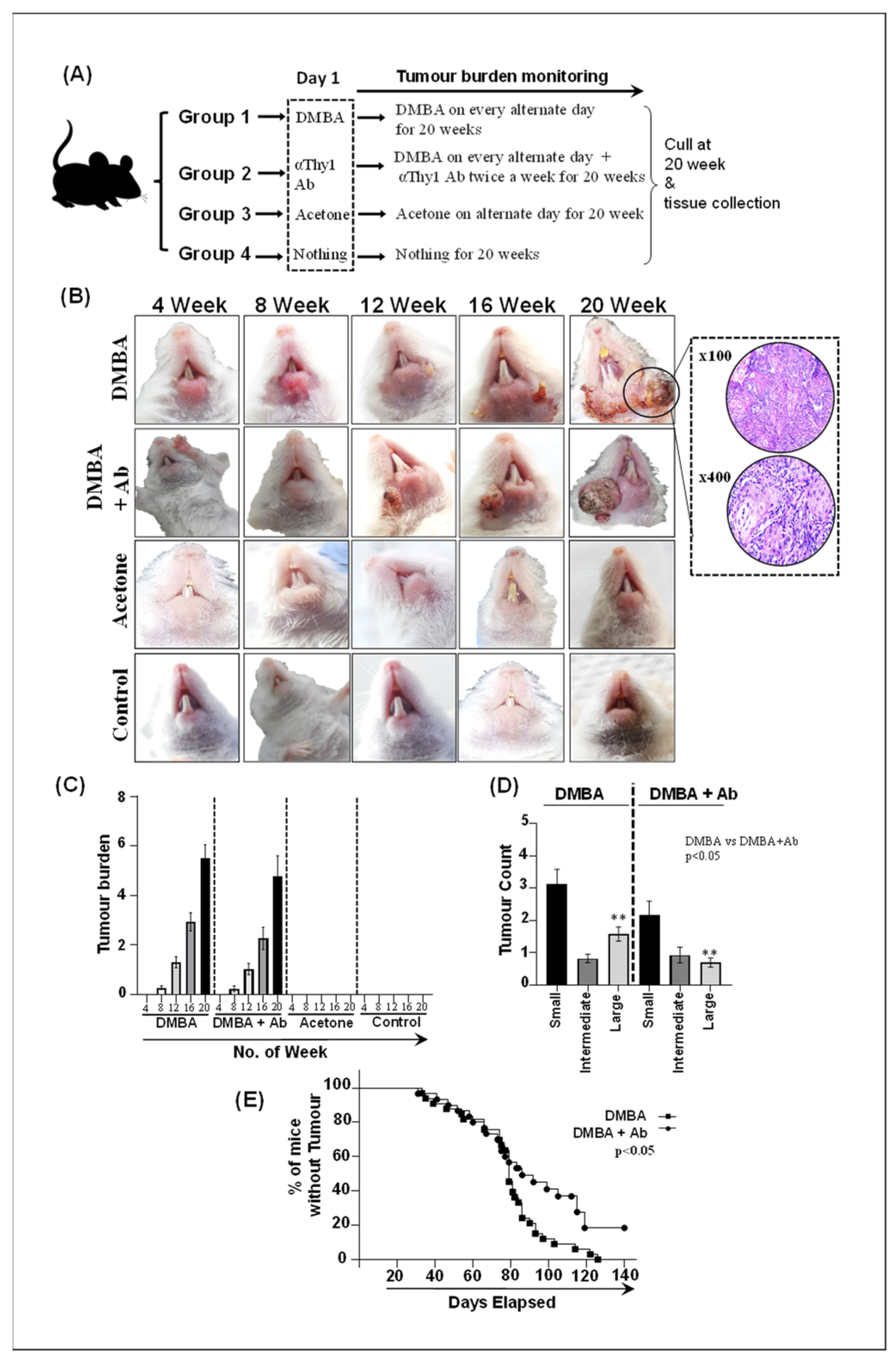
2.5. ILC Inhibition Decelerates Tumorigenesis and Limits Tumor Expansion In Vivo
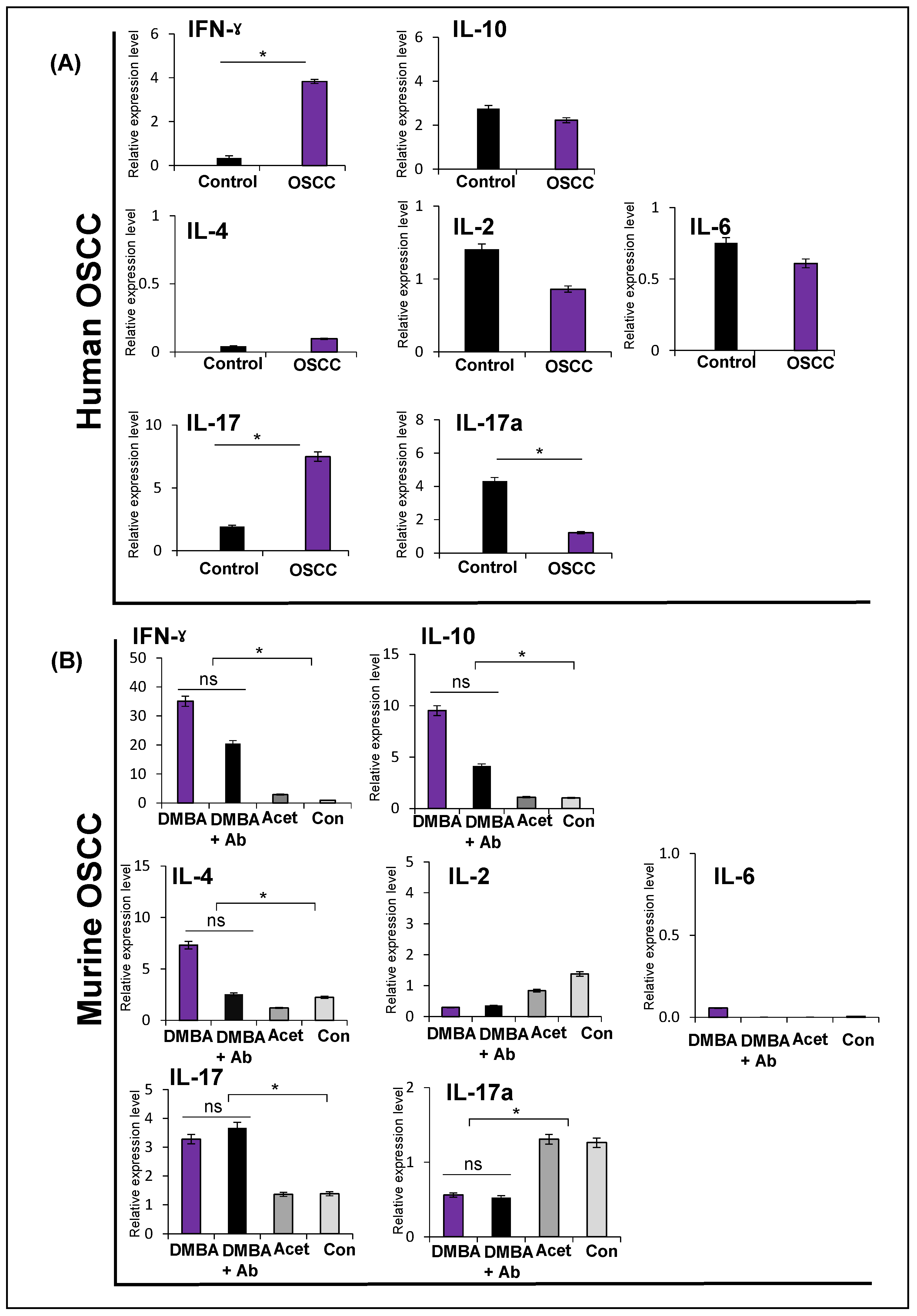
2.6. Expression of Cytokines in Human and Murine Oral Squamous Cell Carcinoma
3. Discussion
4. Materials and Methods
4.1. Patients
4.2. Murine Experiments
4.3. Quantification of Lymphocyte Infiltration in Human and Murine OSCC
4.4. Isolation of Mononuclear Cells
4.5. ILCs Identification via Flowcytometry
4.6. RNA Extraction from Murine and Human OSCC Tissues
4.7. Complementary DNA (cDNA) Synthesis
4.8. Quantitative Real Time Polymerase Chain Reaction (RT-qPCR)
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Anwar, N.; Pervez, S.; Chundriger, Q.; Awan, S.; Moatter, T.; Ali, T.S. Oral cancer: Clinicopathological features and associated risk factors in a high-risk population presenting to a major tertiary care center in Pakistan. PLoS ONE 2020, 15, e0236359. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.Z.; Xie, L.; Shang, Z.J. Burden of Oral Cancer on the 10 Most Populous Countries from 1990 to 2019: Estimates from the Global Burden of Disease Study 2019. Int. J. Environ. Res. Public Health 2022, 19, 875. [Google Scholar] [CrossRef] [PubMed]
- Peltanova, B.; Raudenska, M.; Masarik, M. Effect of tumor microenvironment on pathogenesis of the head and neck squamous cell carcinoma: A systematic review. Mol. Cancer 2019, 18, 63. [Google Scholar] [CrossRef] [Green Version]
- Niklander, S.E. Inflammatory Mediators in Oral Cancer: Pathogenic Mechanisms and Diagnostic Potential. Front. Oral Health 2021, 2, 642238. [Google Scholar] [CrossRef]
- Ralli, M.; Grasso, M.; Gilardi, A.; Ceccanti, M.; Messina, M.P.; Tirassa, P.; Fiore, M.; Altissimi, G.; Salzano, A.F.; De Vincentiis, M.; et al. The role of cytokines in head and neck squamous cell carcinoma: A review. Clin. Ter. 2020, 171, e268–e274. [Google Scholar] [CrossRef]
- Yuan, X.; Rasul, F.; Nashan, B.; Sun, C. Innate lymphoid cells and cancer: Role in tumor progression and inhibition. Eur. J. Immunol. 2021, 51, 2188–2205. [Google Scholar] [CrossRef]
- Moreno-Nieves, U.Y.; Tay, J.K.; Saumyaa, S.; Horowitz, N.B.; Shin, J.H.; Mohammad, I.A.; Luca, B.; Mundy, D.C.; Gulati, G.S.; Bedi, N.; et al. Landscape of innate lymphoid cells in human head and neck cancer reveals divergent NK cell states in the tumor microenvironment. Proc. Natl. Acad. Sci. USA 2021, 118, e2101169118. [Google Scholar] [CrossRef] [PubMed]
- Vivier, E.; Artis, D.; Colonna, M.; Diefenbach, A.; Di Santo, J.P.; Eberl, G.; Koyasu, S.; Locksley, R.M.; McKenzie, A.N.J.; Mebius, R.E.; et al. Innate lymphoid cells: 10 years on. Cell 2018, 174, 1054–1066. [Google Scholar] [CrossRef] [Green Version]
- Sun, J.; Tang, Q.; Zhang, J.; Chen, G.; Peng, J.; Chen, L. Possible Immunotherapeutic Strategies Based on Carcinogen-Dependent Subgroup Classification for Oral Cancer. Front. Mol. Biosci. 2021, 8, 717038. [Google Scholar] [CrossRef]
- Kamran, N.; Alam, S.; Nasir, S.; Ali, K.; Bakhtiar, S.; Khurshid, A.; Ullah, I. Expression of CD3 Levels of Tumor Infiltrating Lymphocytes (TILs) in Various Grades of Oral Squamous Cell Carcinoma of Tertiary Care Hospitals of Peshawar. Pak. J. Med. Health Sci. 2022, 16, 630. [Google Scholar] [CrossRef]
- Shaban, M.; Khurram, S.A.; Fraz, M.M.; Alsubaie, N.; Masood, I.; Mushtaq, S.; Hassan, M.; Loya, A.; Rajpoot, N.M. A Novel Digital Score for Abundance of Tumour Infiltrating Lymphocytes Predicts Disease Free Survival in Oral Squamous Cell Carcinoma. Sci. Rep. 2019, 9, 13341. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elmusrati, A.; Wang, J.; Wang, C.Y. Tumor microenvironment and immune evasion in head and neck squamous cell carcinoma. Int. J. Oral Sci. 2021, 13, 24. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.K.; Busuttil, R.A.; Boussioutas, A. The Role of Innate Immune Cells in Tumor Invasion and Metastasis. Cancers 2021, 13, 5885. [Google Scholar] [CrossRef] [PubMed]
- Xu, Q.; Wang, C.; Yuan, X.; Feng, Z.; Han, Z. Prognostic Value of Tumor-Infiltrating Lymphocytes for Patients with Head and Neck Squamous Cell Carcinoma. Transl. Oncol. 2017, 10, 10–16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salgado, R.; Denkert, C.; Demaria, S.; Sirtaine, N.; Klauschen, F.; Pruneri, G.; Wienert, S.; Vanden, E.G.; Baehner, F.L.; Penault-Llorca, F.; et al. International TILs Working Group 2014. The evaluation of tumor-infiltrating lymphocytes (TILs) in breast cancer: Recommendations by an International TILs Working Group 2014. Ann. Oncol. 2015, 26, 259–271. [Google Scholar] [CrossRef]
- Heikkinen, I.; Bello, I.O.; Wahab, A.; Hagström, J.; Haglund, C.; Coletta, R.D.; Nieminen, P.; Mäkitie, A.A.; Salo, T.; Leivo, I.; et al. Assessment of Tumor-infiltrating Lymphocytes Predicts the Behavior of Early-stage Oral Tongue Cancer. Am. J. Surg. Pathol. 2019, 43, 1392–1396. [Google Scholar] [CrossRef]
- Almangush, A.; De Keukeleire, S.; Rottey, S.; Ferdinande, L.; Vermassen, T.; Leivo, I.; Mäkitie, A.A. Tumor-Infiltrating Lymphocytes in Head and Neck Cancer: Ready for Prime Time? Cancers 2022, 14, 1558. [Google Scholar] [CrossRef]
- De Keukeleire, S.J.; Vermassen, T.; De Meulenaere, A.; Deron, P.; Huvenne, W.; Duprez, F.; Creytens, D.; Van Dorpe, J.; Rottey, S.; Ferdinande, L. Tumour infiltrating lymphocytes in oropharyngeal carcinoma: Prognostic value and evaluation of a standardised method. Pathology 2021, 53, 836–843. [Google Scholar] [CrossRef]
- Koh, J.; Kim, H.Y.; Lee, Y.; Park, I.K.; Kang, C.H.; Kim, Y.T.; Kim, J.E.; Choi, M.; Lee, W.W.; Jeon, Y.K.; et al. IL23-Producing Human Lung Cancer Cells Promote Tumor Growth via Conversion of Innate Lymphoid Cell 1 (ILC1) into ILC3. Clin. Cancer Res. 2019, 25, 4026–4037. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Song, Y.; Lin, D.; Lei, L.; Mei, Y.; Jin, Z.; Gong, H.; Zhu, Y.; Hu, B.; Zhang, Y.; et al. NCR− group 3 innate lymphoid cells orchestrate IL-23/IL-17 axis to promote hepatocellular carcinoma development. EBioMedicine 2019, 41, 333–344. [Google Scholar] [CrossRef]
- Ghaedi, M.; Ohashi, P.S. ILC transdifferentiation: Roles in cancer progression. Cell Res. 2020, 30, 562–563. [Google Scholar] [CrossRef] [PubMed]
- Goertzen, C.; Mahdi, H.; Laliberte, C.; Meirson, T.; Eymael, D.; Gil-Henn, H.; Magalhaes, M. Oral inflammation promotes oral squamous cell carcinoma invasion. Oncotarget 2018, 9, 29047–29063. [Google Scholar] [CrossRef] [Green Version]
- Warnakulasuriya, S.; Straif, K. Carcinogenicity of smokeless tobacco: Evidence from studies in humans & experimental animals. Indian J. Med. Res. 2018, 148, 681–686. [Google Scholar] [CrossRef]
- Edirisinghe, S.T.; Weerasekera, M.; De Silva, D.K.; Liyanage, I.; Niluka, M.; Madushika, K.; Deegodagamage, S.; Wijesundara, C.; Rich, A.M.; De Silva, H.; et al. The Risk of Oral Cancer among Different Categories of Exposure to Tobacco Smoking in Sri Lanka. Asian Pac. J. Cancer Prev. 2022, 23, 2929–2935. [Google Scholar] [CrossRef]
- Ghias, K.; Rehmani, S.S.; Razzak, S.A.; Madhani, S.; Azim, M.K.; Ahmed, R.; Khan, M.J. Mutational landscape of head and neck squamous cell carcinomas in a South Asian population. Genet. Mol. Biol. 2019, 4242, 526–542. [Google Scholar] [CrossRef] [PubMed]
- Entwistle, L.J.; Gregory, L.G.; Oliver, R.A.; Branchett, W.J.; Puttur, F.; Lloyd, C.M. Pulmonary Group 2 Innate Lymphoid Cell Phenotype Is Context Specific: Determining the Effect of Strain, Location, and Stimuli. Front. Immunol. 2020, 10, 3114. [Google Scholar] [CrossRef]
- Bie, Q.; Zhang, P.; Su, Z.; Zheng, D.; Ying, X.; Wu, Y.; Yang, H.; Chen, D.; Wang, S.; Xu, H. Polarization of ILC2s in peripheral blood might contribute to immunosuppressive microenvironment in patients with gastric cancer. J. Immunol. Res. 2014, 2014, 923135. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, Y.; Yan, Y.; Su, Z.; Bie, Q.; Chen, X.; Barnie, P.A.; Guo, Q.; Wang, S.; Xu, H. Enhanced circulating ILC2s and MDSCs may contribute to ensure maintenance of Th2 predominant in patients with lung cancer. Mol. Med. Rep. 2017, 15, 4374–4381. [Google Scholar] [CrossRef] [Green Version]
- Trabanelli, S.; Curti, A.; Lecciso, M.; Salomé, B.; Riether, C.; Ochsenbein, A.; Romero, P.; Jandus, C. CD127+ innate lymphoid cells are dysregulated in treatment naïve acute myeloid leukemia patients at diagnosis. Haematologica 2015, 100, e257–e260. [Google Scholar] [CrossRef] [Green Version]
- Salimi, M.; Wang, R.; Yao, X.; Li, X.; Wang, X.; Hu, Y.; Chang, X.; Fan, P.; Dong, T.; Ogg, G. Activated innate lymphoid cell populations accumulate in human tumour tissues. BMC Cancer 2018, 18, 341. [Google Scholar] [CrossRef]
- Irshad, S.; Flores-Borja, F.; Lawler, K.; Monypenny, J.; Evans, R.; Male, V.; Gordon, P.; Cheung, A.; Gazinska, P.; Noor, F.; et al. RORγt+ Innate Lymphoid Cells Promote Lymph Node Metastasis of Breast Cancers. Cancer Res. 2017, 77, 1083–1096. [Google Scholar] [CrossRef] [Green Version]
- Brown, J.L.; Campbell, L.; Malcolm, J.; Adrados, P.A.; Butcher, J.P.; Culshaw, S. Enrichment of Innate Lymphoid Cell Populations in Gingival Tissue. J. Dent. Res. 2018, 97, 1399–1405. [Google Scholar] [CrossRef] [Green Version]
- Pandiyan, P.; Bhaskaran, N.; Zhang, Y.; Weinberg, A. Isolation of T cells from mouse oral tissues. Biol. Proced. Online 2014, 16, 4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shannon, J.P.; Vrba, S.M.; Reynoso, G.V.; Wynne-Jones, E.; Kamenyeva, O.; Malo, C.S.; Cherry, C.R.; McManus, D.T.; Hickman, H.D. Group 1 innate lymphoid-cell-derived interferon-γ maintains anti-viral vigilance in the mucosal epithelium. Immunity 2021, 54, 276–290.e5. [Google Scholar] [CrossRef] [PubMed]
- Warner, K.; Ghaedi, M.; Chung, D.C.; Jacquelot, N.; Ohashi, P.S. Innate lymphoid cells in early tumor development. Front. Immunol. 2022, 13, 948358. [Google Scholar] [CrossRef]
- Wan, J.; Wu, Y.; Huang, L.; Tian, Y.; Ji, X.; Abdelaziz, M.H.; Cai, W.; Dineshkumar, K.; Lei, Y.; Yao, S. ILC2-derived IL-9 inhibits colorectal cancer progression by activating CD8+ T cells. Cancer Lett. 2021, 502, 34–43. [Google Scholar] [CrossRef]
- Wang, S.; Qu, Y.; Xia, P.; Chen, Y.; Zhu, X.; Zhang, J.; Wang, G.; Tian, Y.; Ying, J.; Fan, Z. Transdifferentiation of tumor infiltrating innate lymphoid cells during progression of colorectal cancer. Cell Res. 2020, 30, 610–622. [Google Scholar] [CrossRef]
- Kondoh, N.; Mizuno-Kamiya, M.; Umemura, N.; Takayama, E.; Kawaki, H.; Mitsudo, K.; Muramatsu, Y.; Sumitomo, S. Immunomodulatory aspects in the progression and treatment of oral malignancy. Jpn. Dent. Sci. Rev. 2019, 55, 113–120. [Google Scholar] [CrossRef]
- Yu, M.; Qian, X.X.; Li, G.; Cheng, Z.; Lin, Z. Prognostic biomarker IL17A correlated with immune infiltrates in head and neck cancer. World J. Surg. Oncol. 2022, 20, 243. [Google Scholar] [CrossRef]
- Wei, T.; Cong, X.; Wang, X.T.; Xu, X.J.; Min, S.N.; Ye, P.; Peng, X.; Wu, L.L.; Yu, G.Y. Interleukin-17A promotes tongue squamous cell carcinoma metastasis through activating miR-23b/versican pathway. Oncotarget 2017, 8, 6663–6680. [Google Scholar] [CrossRef] [PubMed]
- Woodford, D.; Johnson, S.D.; De Costa, A.M.; Young, M.R. An Inflammatory Cytokine Milieu is Prominent in Premalignant Oral Lesions, but Subsides when Lesions Progress to Squamous Cell Carcinoma. J. Clin. Cell Immunol. 2014, 5, 230. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tait, W.E.D.; Beamer, C.A. Isolation and Identification of Innate Lymphoid Cells (ILCs) for Immunotoxicity Testing. Methods Mol. Biol. 2018, 1803, 353–370. [Google Scholar] [CrossRef]
- Bando, J.K.; Colonna, M. Innate lymphoid cell function in the context of adaptive immunity. Nat. Immunol. 2016, 17, 783–789. [Google Scholar] [CrossRef]
- Syed, S.A.; Qureshi, M.A.; Khan, S.; Kumar, R.; Shafique, Y.; Khan, B.A.; Safdar, J. Development of a murine model of oral carcinogenesis: An accelerated tool for biomarker and anti-tumour drug discovery. Ecancermedicalscience 2022, 16, 1413. [Google Scholar] [CrossRef] [PubMed]
- Overbergh, L.; Giulietti, A.; Valckx, D.; Decallonne, B.; Bouillon, R.; Mathieu, C. The use of real-time reverse transcriptase PCR for the quantification of cytokine gene expression. J. Biomol. Tech. 2003, 14, 33–43. [Google Scholar]
- Capomaccio, S.; Cappelli, K.; Spinsanti, G.; Mencarelli, M.; Muscettola, M.; Felicetti, M.; Supplizi, A.V.; Bonifazi, M. Athletic humans and horses: Comparative analysis of interleukin-6 (IL-6) and IL-6 receptor (IL-6R) expression in peripheral blood mononuclear cells in trained and untrained subjects at rest. BMC Physiol. 2011, 11, 3. [Google Scholar] [CrossRef] [Green Version]
- Wang, Q.; Zheng, Y.; Huang, Z.; Tian, Y.; Zhou, J.; Mao, Q.; Wu, Y.; Ni, B. Activated IL-23/IL-17 pathway closely correlates with increased Foxp3 expression in livers of chronic hepatitis B patients. BMC Immunol. 2011, 12, 25. [Google Scholar] [CrossRef] [Green Version]
- Vazquez-Tello, A.; Halwani, R.; Li, R.; Nadigel, J.; Bar, A.; Mazer, B.D.; Eidelman, D.H.; Al-Muhsen, S.; Hamid, Q. IL-17A and IL-17F Expression in B Lymphocytes. Int. Arch. Allergy Immunol. 2012, 157, 406–416. [Google Scholar] [CrossRef]
- Smith, E.; Stark, M.A.; Zarbock, A.; Burcin, T.L.; Bruce, A.C.; Vaswani, D.; Foley, P.; Ley, K. IL-17A inhibits the expansion of IL-17A-producing T cells in mice through "short-loop" inhibition via IL-17 receptor. J. Immunol. 2008, 181, 1357–1364. [Google Scholar] [CrossRef]
- Torabi, A.; Tahmoorespur, M.; Vahedi, F.; Mosavari, N.; Nassiri, M. Quantiation of IL-4, IL-10 and IFN-γ genes expression after immunization of mice with CFP-10 and ESAT-6 containing vectors. Iran. J. Immunol. 2013, 10, 205–215. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Characteristics (N = 52) | Frequency (%) |
|---|---|
| Mean Age in years | |
| Male | 46 |
| Female | 57.4 |
| Gender | |
| Male | 42 (80.8) |
| Female | 10 (19.2) |
| Site | |
| Buccal mucosa | 28 (53.8) |
| Tongue | 10 (19.2) |
| Lip | 05 (9.7) |
| Alveolus | 05 (9.7) |
| Hard palate | 04 (7.6) |
| Tobacco Habits | |
| Yes | 42 (80.8) |
| No | 10 (19.2) |
| Type of biopsy | |
| Surgical excision | 29 (55.8) |
| Punch/incisional | 23 (44.2) |
| Grade | |
| Moderately differentiated | 38 (73.1) |
| Well differentiated | 09 (17.3) |
| Poorly differentiated | 05 (9.6) |
| Size * (n = 29) | |
| pT1 | 02 (7.0) |
| pT2 | 13 (44.8) |
| pT3 | 03 (10.3) |
| pT4 | 11 (37.9) |
| Nodal metastasis * (n = 29) | |
| pN0 | 15 (51.7) |
| pN1 | 03 (10.3) |
| pN2 | 06 (20.8) |
| pN3 | 05 (17.2) |
| Lymphovascular invasion (n = 29) | |
| No | 27 (93.1) |
| Yes | 02 (6.9) |
| Perineural invasion (n = 29) | |
| No | 18 (62.1) |
| Yes | 11 (37.9) |
| Human Cytokines | Primer Sequence (5′–3′) | Length |
|---|---|---|
| IL-2 [45] | F: AACTCACCAGGATGCTCACATTTA R: TCCCTGGGTCTTAAGTGAAAGTTT | 24 24 |
| IL-4 [45] | F: CCACGGACACAAGTGCGATA R: CCCTGCAGAAGGTTTCCTTCT | 20 21 |
| IL-6 [46] | F: CCACTCACCTCTTCAGAACGAAT R: TTGGAAGCATCCATCTTTTTCA | 23 22 |
| IL-10 [45] | F: GTGATGCCCCAAGCTGAGA R: CACGGCCTTGCTCTTGTTTT | 19 20 |
| IL-17 [47] | F: TCAACCCGATTGTCCACCAT R: GAGTTTAGTCCGAAATGAGGCTG | 20 23 |
| IL-17a [48] | F: CAT CCA TAA CCG GAA TAC CAA TA R: TAG TCC ACG TTC CCA TCA GC | 23 20 |
| IFN-γ [45] | F: TCAGCTCTGCATCGTTTTGG R: GTTCCATTATCCGCTACATCTGAA | 20 24 |
| GAPDH | F: GCATCCTGGGCTACACTGA R: CCACCACCCTGTTGCTGTA | 19 19 |
| Murine Cytokines | Primer Sequence (5′–3′) | Length |
|---|---|---|
| IL-2 [45] | F: CCTGAGCAGGATGGAGAATTACA R: TCCAGAACATGCCGCAGAG | 23 19 |
| IL-4 [45] | F: ACAGGAGAAGGGACGCCAT R: GAAGCCCTACAGACGAGCTCA | 19 21 |
| IL-6 [45] | F: GAGGATACCACTCCCAACAGACC R: AAGTGCATCATCGTTGTTCATACA | 23 24 |
| IL-10 [45] | F: GGTTGCCAAGCCTTATCGGA R: ACCTGCTCCACTGCCTTGCT | 20 20 |
| IL-17 [45] | F: GCTCCAGAAGGCCCTCAGA R: AGCTTTCCCTCCGCATTGA | 19 19 |
| IL-17a [49] | F: ATCCCTCAAAGCTCAGCGTGTC R: GGGTCTTCATTGCGGTGGAGAG | 22 22 |
| IFN-γ [45] | F: TCAAGTGGCATAGATGTGGAAGAA R: TGGCTCTGCAGGATTTTCATG | 24 21 |
| GAPDH [50] | F: TTCACCACCATGGAGAAGGC R: GGCATGGACTGTGGTCATGA | 20 20 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Syed, S.A.; Qureshi, M.A.; Khan, S.; Kumar, R.; Khyani, I.A.M.; Khan, B.A.; Safdar, J. Differential Regulation of Innate Lymphoid Cells in Human and Murine Oral Squamous Cell Carcinoma. Int. J. Mol. Sci. 2023, 24, 1627. https://doi.org/10.3390/ijms24021627
Syed SA, Qureshi MA, Khan S, Kumar R, Khyani IAM, Khan BA, Safdar J. Differential Regulation of Innate Lymphoid Cells in Human and Murine Oral Squamous Cell Carcinoma. International Journal of Molecular Sciences. 2023; 24(2):1627. https://doi.org/10.3390/ijms24021627
Chicago/Turabian StyleSyed, Sofia Ali, Muhammad Asif Qureshi, Saeed Khan, Rajesh Kumar, Iqbal A. Muhammad Khyani, Bilal Ahmed Khan, and Jawad Safdar. 2023. "Differential Regulation of Innate Lymphoid Cells in Human and Murine Oral Squamous Cell Carcinoma" International Journal of Molecular Sciences 24, no. 2: 1627. https://doi.org/10.3390/ijms24021627