
Analysis of Serial Neuroblastoma PDX Passages in Mice Allows the Identification of New Mediators of Neuroblastoma Aggressiveness

 , ,
, ,  , , , and
, , , and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Successive PDX Passages in Mice Show a Pattern of Increased Aggressiveness
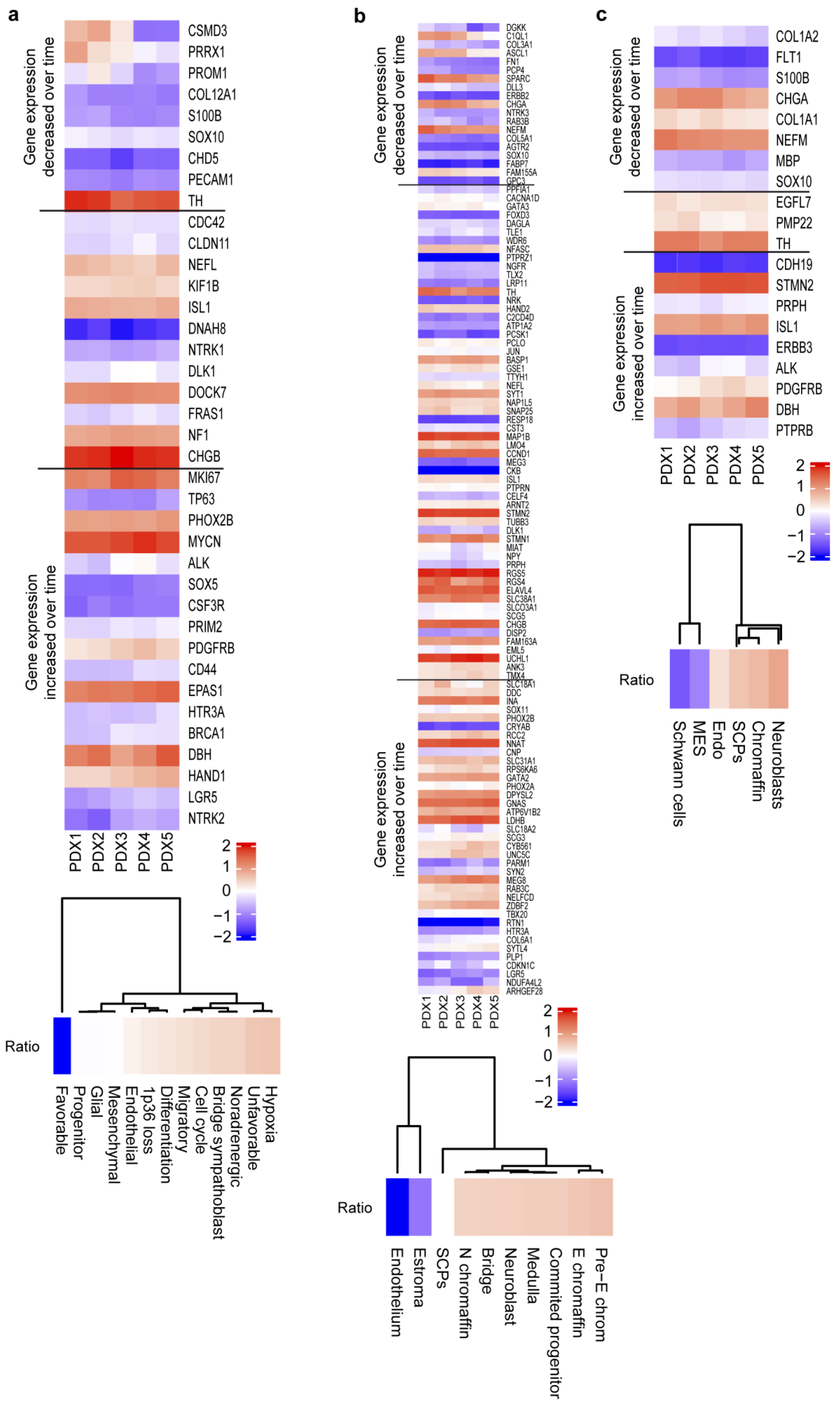
2.2. PDX Gene Expression Patterns over Time Associate with Changes in Undifferentiation and Proliferation Genes
2.3. miRNA for Relevant Neuroblastoma Targets Are Associated with PDX Passage
2.4. PDX Passages Permit the Identification of Genes with Relevance in Neuroblastoma Progression
3. Discussion
4. Materials and Methods
4.1. Patient-Derived Xenografts (PDXs)
4.2. RNA Extraction and mRNA and miRNA Array
4.3. Microarray Analysis
4.4. Patient Data Correlation
4.5. Immunohistochemistry
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Matthay, K.K.; Maris, J.M.; Schleiermacher, G.; Nakagawara, A.; Mackall, C.L.; Diller, L.; Weiss, W.A. Neuroblastoma. Nat. Rev. Dis. Prim. 2016, 2, 16078. [Google Scholar] [CrossRef] [PubMed]
- Johnsen, J.I.; Dyberg, C.; Wickström, M. Neuroblastoma-A Neural Crest Derived Embryonal Malignancy. Front. Mol. Neurosci. 2019, 12, 9. [Google Scholar] [CrossRef] [PubMed]
- Gomez, R.L.; Ibragimova, S.; Ramachandran, R.; Philpott, A.; Ali, F.R. Tumoral Heterogeneity in Neuroblastoma. Biochim. Biophys. Acta—Rev. Cancer 2022, 1877, 188805. [Google Scholar] [CrossRef] [PubMed]
- Zeineldin, M.; Patel, A.G.; Dyer, M.A. Neuroblastoma: When Differentiation Goes Awry. Neuron 2022, 110, 2916–2928. [Google Scholar] [CrossRef] [PubMed]
- van Groningen, T.; Koster, J.; Valentijn, L.J.; Zwijnenburg, D.A.; Akogul, N.; Hasselt, N.E.; Broekmans, M.; Haneveld, F.; Nowakowska, N.E.; Bras, J.; et al. Neuroblastoma Is Composed of Two Super-Enhancer-Associated Differentiation States. Nat. Genet. 2017, 49, 1261–1266. [Google Scholar] [CrossRef]
- Bedoya-Reina, O.C.; Li, W.; Arceo, M.; Plescher, M.; Bullova, P.; Pui, H.; Kaucka, M.; Kharchenko, P.; Martinsson, T.; Holmberg, J.; et al. Single-Nuclei Transcriptomes from Human Adrenal Gland Reveal Distinct Cellular Identities of Low and High-Risk Neuroblastoma Tumors. Nat. Commun. 2021, 12, 5309. [Google Scholar] [CrossRef]
- Jansky, S.; Sharma, A.K.; Körber, V.; Quintero, A.; Toprak, U.H.; Wecht, E.M.; Gartlgruber, M.; Greco, A.; Chomsky, E.; Grünewald, T.G.P.; et al. Single-Cell Transcriptomic Analyses Provide Insights into the Developmental Origins of Neuroblastoma. Nat. Genet. 2021, 53, 683–693. [Google Scholar] [CrossRef]
- Westermark, U.K.; Wilhelm, M.; Frenzel, A.; Henriksson, M.A. The MYCN Oncogene and Differentiation in Neuroblastoma. Semin. Cancer Biol. 2011, 21, 256–266. [Google Scholar] [CrossRef]
- Nolan, J.C.; Frawley, T.; Tighe, J.; Soh, H.; Curtin, C.; Piskareva, O. Preclinical Models for Neuroblastoma: Advances and Challenges. Cancer Lett. 2020, 474, 53–62. [Google Scholar] [CrossRef]
- Kamili, A.; Atkinson, C.; Trahair, T.N.; Fletcher, J.I. Mouse Models of High-Risk Neuroblastoma. Cancer Metastasis Rev. 2020, 39, 261–274. [Google Scholar] [CrossRef]
- Kamili, A.; Gifford, A.J.; Li, N.; Mayoh, C.; Chow, S.-O.; Failes, T.W.; Eden, G.L.; Cadiz, R.; Xie, J.; Lukeis, R.E.; et al. Accelerating Development of High-Risk Neuroblastoma Patient-Derived Xenograft Models for Preclinical Testing and Personalised Therapy. Br. J. Cancer 2020, 122, 680–691. [Google Scholar] [CrossRef] [Green Version]
- Braekeveldt, N.; Bexell, D. Patient-Derived Xenografts as Preclinical Neuroblastoma Models. Cell Tissue Res. 2018, 372, 233–243. [Google Scholar] [CrossRef]
- Mañas, A.; Aaltonen, K.; Andersson, N.; Hansson, K.; Adamska, A.; Seger, A.; Yasui, H.; van den Bos, H.; Radke, K.; Esfandyari, J.; et al. Clinically Relevant Treatment of PDX Models Reveals Patterns of Neuroblastoma Chemoresistance. Sci. Adv. 2022, 8, 4617. [Google Scholar] [CrossRef]
- Braekeveldt, N.; von Stedingk, K.; Fransson, S.; Martinez-Monleon, A.; Lindgren, D.; Axelson, H.; Levander, F.; Willforss, J.; Hansson, K.; Øra, I.; et al. Patient-Derived Xenograft Models Reveal Intratumor Heterogeneity and Temporal Stability in Neuroblastoma. Cancer Res. 2018, 78, 5958–5969. [Google Scholar] [CrossRef] [Green Version]
- Braekeveldt, N.; Wigerup, C.; Gisselsson, D.; Mohlin, S.; Merselius, M.; Beckman, S.; Jonson, T.; Börjesson, A.; Backman, T.; Tadeo, I.; et al. Neuroblastoma Patient-Derived Orthotopic Xenografts Retain Metastatic Patterns and Geno- and Phenotypes of Patient Tumours. Int. J. Cancer 2015, 136, E252–E261. [Google Scholar] [CrossRef] [Green Version]
- Stewart, E.; Federico, S.M.; Chen, X.; Shelat, A.A.; Bradley, C.; Gordon, B.; Karlstrom, A.; Twarog, N.R.; Clay, M.R.; Bahrami, A.; et al. Orthotopic Patient-Derived Xenografts of Paediatric Solid Tumours. Nature 2017, 549, 96–100. [Google Scholar] [CrossRef] [Green Version]
- Woo, X.Y.; Giordano, J.; Srivastava, A.; Zhao, Z.-M.; Lloyd, M.W.; de Bruijn, R.; Suh, Y.-S.; Patidar, R.; Chen, L.; Scherer, S.; et al. Conservation of Copy Number Profiles during Engraftment and Passaging of Patient-Derived Cancer Xenografts. Nat. Genet. 2021, 53, 86–99. [Google Scholar] [CrossRef]
- Ernst, J.; Bar-Joseph, Z. STEM: A Tool for the Analysis of Short Time Series Gene Expression Data. BMC Bioinform. 2006, 7, 1–11. [Google Scholar] [CrossRef] [Green Version]
- De Preter, K.; Mestdagh, P.; Vermeulen, J.; Zeka, F.; Naranjo, A.; Bray, I.; Castel, V.; Chen, C.; Drozynska, E.; Eggert, A.; et al. MiRNA Expression Profiling Enables Risk Stratification in Archived and Fresh Neuroblastoma Tumor Samples. Clin. Cancer Res. 2011, 17, 7684–7692. [Google Scholar] [CrossRef] [Green Version]
- Hanemaaijer, E.S.; Margaritis, T.; Sanders, K.; Bos, F.L.; Candelli, T.; Al-Saati, H.; van Noesel, M.M.; Meyer-Wentrup, F.A.G.; van de Wetering, M.; Holstege, F.C.P.; et al. Single-Cell Atlas of Developing Murine Adrenal Gland Reveals Relation of Schwann Cell Precursor Signature to Neuroblastoma Phenotype. Proc. Natl. Acad. Sci. USA 2021, 118, e2022350118. [Google Scholar] [CrossRef]
- Abdolahi, S.; Ghazvinian, Z.; Muhammadnejad, S.; Saleh, M.; Asadzadeh Aghdaei, H.; Baghaei, K. Patient-Derived Xenograft (PDX) Models, Applications and Challenges in Cancer Research. J. Transl. Med. 2022, 20, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Blomme, A.; Van Simaeys, G.; Doumont, G.; Costanza, B.; Bellier, J.; Otaka, Y.; Sherer, F.; Lovinfosse, P.; Boutry, S.; Palacios, A.P.; et al. Murine Stroma Adopts a Human-like Metabolic Phenotype in the PDX Model of Colorectal Cancer and Liver Metastases. Oncogene 2018, 37, 1237–1250. [Google Scholar] [CrossRef] [PubMed]
- Huang, W.-T.; Lin, T.-S.; Wu, J.-Y.; Hong, J.-M.; Chen, Y.-L.; Qiu, F.-N. Evaluation of MiR-429 as a Novel Serum Biomarker for Pancreatic Ductal Adenocarcinoma and Analysis Its Tumor Suppressor Function and Target Genes. Eur. Rev. Med. Pharmacol. Sci. 2022, 26, 4638–4653. [Google Scholar] [CrossRef] [PubMed]
- Gu, Y.; Zhou, Z. Berberine Inhibits the Proliferation, Invasion and Migration of Endometrial Stromal Cells by Downregulating MiR-429. Mol. Med. Rep. 2021, 23, 416. [Google Scholar] [CrossRef] [PubMed]
- Zhou, G.-H.; Lu, Y.-Y.; Xie, J.-L.; Gao, Z.-K.; Wu, X.-B.; Yao, W.-S.; Gu, W.-G. Overexpression of MiR-758 Inhibited Proliferation, Migration, Invasion, and Promoted Apoptosis of Non-Small Cell Lung Cancer Cells by Negatively Regulating HMGB. Biosci. Rep. 2019, 39, BSR20180855. [Google Scholar] [CrossRef] [Green Version]
- Mlakar, V.; Jurkovic Mlakar, S.; Lopez, G.; Maris, J.M.; Ansari, M.; Gumy-Pause, F. 11q Deletion in Neuroblastoma: A Review of Biological and Clinical Implications. Mol. Cancer 2017, 16, 114. [Google Scholar] [CrossRef] [Green Version]
- Pallarès-Albanell, J.; Zomeño-Abellán, M.T.; Escaramís, G.; Pantano, L.; Soriano, A.; Segura, M.F.; Martí, E. A High-Throughput Screening Identifies MicroRNA Inhibitors That Influence Neuronal Maintenance and/or Response to Oxidative Stress. Mol. Ther.—Nucleic Acids 2019, 17, 374–387. [Google Scholar] [CrossRef] [Green Version]
- Cattaneo, M.G.; Taylor, J.E.; Culler, M.D.; Nisoli, E.; Vicentini, L.M. Selective Stimulation of Somatostatin Receptor Subtypes: Differential Effects on Ras/MAP Kinase Pathway and Cell Proliferation in Human Neuroblastoma Cells. FEBS Lett. 2000, 481, 271–276. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Cao, L.; Song, Y.; Kang, Z.; Liu, T.; Ding, J.; Hu, G.; Lu, M. Mitochondrial Glutamine Transporter SLC1A5_var, a Potential Target to Suppress Astrocyte Reactivity in Parkinson’s Disease. Cell Death Dis. 2022, 13, 946. [Google Scholar] [CrossRef]
- Pan, B.; Li, Y.; Xu, Z.; Miao, Y.; Yin, H.; Kong, Y.; Zhang, X.; Liang, J.; Xia, Y.; Wang, L.; et al. Identifying a Novel Ferroptosis-Related Prognostic Score for Predicting Prognosis in Chronic Lymphocytic Leukemia. Front. Immunol. 2022, 13, 962000. [Google Scholar] [CrossRef]
- Hassanein, M.; Qian, J.; Hoeksema, M.D.; Wang, J.; Jacobovitz, M.; Ji, X.; Harris, F.T.; Harris, B.K.; Boyd, K.L.; Chen, H.; et al. Targeting SLC1a5-Mediated Glutamine Dependence in Non-Small Cell Lung Cancer. Int. J. Cancer 2015, 137, 1587–1597. [Google Scholar] [CrossRef] [Green Version]
- Crone, M.; Mah, J.K. Current and Emerging Therapies for Duchenne Muscular Dystrophy. Curr. Treat. Options Neurol. 2018, 20, 31. [Google Scholar] [CrossRef]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gómez-Muñoz, M.A.; Aguilar-Morante, D.; Colmenero-Repiso, A.; Amador-Álvarez, A.; Ojeda-Puertas, M.; Cordero Varela, J.A.; Rodríguez-Prieto, I.; Pardal, R.; Vega, F.M. Analysis of Serial Neuroblastoma PDX Passages in Mice Allows the Identification of New Mediators of Neuroblastoma Aggressiveness. Int. J. Mol. Sci. 2023, 24, 1590. https://doi.org/10.3390/ijms24021590
Gómez-Muñoz MA, Aguilar-Morante D, Colmenero-Repiso A, Amador-Álvarez A, Ojeda-Puertas M, Cordero Varela JA, Rodríguez-Prieto I, Pardal R, Vega FM. Analysis of Serial Neuroblastoma PDX Passages in Mice Allows the Identification of New Mediators of Neuroblastoma Aggressiveness. International Journal of Molecular Sciences. 2023; 24(2):1590. https://doi.org/10.3390/ijms24021590
Chicago/Turabian StyleGómez-Muñoz, María A., Diana Aguilar-Morante, Ana Colmenero-Repiso, Aida Amador-Álvarez, Mónica Ojeda-Puertas, Juan Antonio Cordero Varela, Ismael Rodríguez-Prieto, Ricardo Pardal, and Francisco M. Vega. 2023. "Analysis of Serial Neuroblastoma PDX Passages in Mice Allows the Identification of New Mediators of Neuroblastoma Aggressiveness" International Journal of Molecular Sciences 24, no. 2: 1590. https://doi.org/10.3390/ijms24021590