
Human Regulatory T Cells: Understanding the Role of Tregs in Select Autoimmune Skin Diseases and Post-Transplant Nonmelanoma Skin Cancers

Abstract
:1. Introduction
2. Classification of Tregs
3. Tissue-Specific Tregs
4. Mechanism of Action
4.1. Starving T Cells of IL-2
4.2. Induction of T Cell Apoptosis
4.3. Production of Inhibitory Cytokines
4.4. Inhibiting Immunostimulatory Signals and Metabolic Activity of APCs
5. Role of Tregs in Select Autoimmune Skin Diseases
5.1. Psoriasis
5.2. Vitiligo
5.3. Systemic Sclerosis
6. The Role of Tregs in Transplantation and Skin Cancer
6.1. Squamous Cell Carcinoma
6.2. Basal Cell Carcinoma
7. Regulatory T Cell Therapies
7.1. IL-2 Based Approaches: Low Dose IL-2 and IL-2 Complexes
7.2. Adoptive Treg Therapy: Polyclonal and Antigen-Specific Tregs
8. Future Directions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Hori, S.; Nomura, T.; Sakaguchi, S. Control of regulatory T cell development by the transcription factor Foxp3. Science 2003, 299, 1057–1061. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martin, B.; Banz, A.; Bienvenu, B.; Cordier, C.; Dautigny, N.; Becourt, C.; Lucas, B. Suppression of CD4+ T lymphocyte effector functions by CD4+CD25+ cells in vivo. J. Immunol. 2004, 172, 3391–3398. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stassen, M.; Jonuleit, H.; Muller, C.; Klein, M.; Richter, C.; Bopp, T.; Schmitt, S.; Schmitt, E. Differential regulatory capacity of CD25+ T regulatory cells and preactivated CD25+ T regulatory cells on development, functional activation, and proliferation of Th2 cells. J. Immunol. 2004, 173, 267–274. [Google Scholar] [CrossRef] [Green Version]
- Oldenhove, G.; de Heusch, M.; Urbain-Vansanten, G.; Urbain, J.; Maliszewski, C.; Leo, O.; Moser, M. CD4+ CD25+ regulatory T cells control T helper cell type 1 responses to foreign antigens induced by mature dendritic cells in vivo. J. Exp. Med. 2003, 198, 259–266. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fontenot, J.D.; Gavin, M.A.; Rudensky, A.Y. Foxp3 programs the development and function of CD4+CD25+ regulatory T cells. Nat. Immunol. 2003, 4, 330–336. [Google Scholar] [CrossRef]
- Rubtsov, Y.P.; Niec, R.E.; Josefowicz, S.; Li, L.; Darce, J.; Mathis, D.; Benoist, C.; Rudensky, A.Y. Stability of the regulatory T cell lineage in vivo. Science 2010, 329, 1667–1671. [Google Scholar] [CrossRef] [Green Version]
- Bailey-Bucktrout, S.L.; Martinez-Llordella, M.; Zhou, X.; Anthony, B.; Rosenthal, W.; Luche, H.; Fehling, H.J.; Bluestone, J.A. Self-antigen-driven activation induces instability of regulatory T cells during an inflammatory autoimmune response. Immunity 2013, 39, 949–962. [Google Scholar] [CrossRef] [Green Version]
- Komatsu, N.; Okamoto, K.; Sawa, S.; Nakashima, T.; Oh-hora, M.; Kodama, T.; Tanaka, S.; Bluestone, J.A.; Takayanagi, H. Pathogenic conversion of Foxp3+ T cells into TH17 cells in autoimmune arthritis. Nat. Med. 2014, 20, 62–68. [Google Scholar] [CrossRef]
- Zhou, X.; Bailey-Bucktrout, S.; Jeker, L.T.; Bluestone, J.A. Plasticity of CD4(+) FoxP3(+) T cells. Curr. Opin. Immunol. 2009, 21, 281–285. [Google Scholar] [CrossRef] [Green Version]
- Hori, S. Lineage stability and phenotypic plasticity of Foxp3(+) regulatory T cells. Immunol. Rev. 2014, 259, 159–172. [Google Scholar] [CrossRef]
- Ohkura, N.; Hamaguchi, M.; Morikawa, H.; Sugimura, K.; Tanaka, A.; Ito, Y.; Osaki, M.; Tanaka, Y.; Yamashita, R.; Nakano, N.; et al. T cell receptor stimulation-induced epigenetic changes and Foxp3 expression are independent and complementary events required for Treg cell development. Immunity 2012, 37, 785–799. [Google Scholar] [CrossRef] [Green Version]
- Sakaguchi, S.; Sakaguchi, N.; Asano, M.; Itoh, M.; Toda, M. Immunologic self-tolerance maintained by activated T cells expressing IL-2 receptor alpha-chains (CD25). Breakdown of a single mechanism of self-tolerance causes various autoimmune diseases. J. Immunol. 1995, 155, 1151–1164. [Google Scholar] [CrossRef]
- Miyara, M.; Yoshioka, Y.; Kitoh, A.; Shima, T.; Wing, K.; Niwa, A.; Parizot, C.; Taflin, C.; Heike, T.; Valeyre, D.; et al. Functional delineation and differentiation dynamics of human CD4+ T cells expressing the FoxP3 transcription factor. Immunity 2009, 30, 899–911. [Google Scholar] [CrossRef] [Green Version]
- Seddiki, N.; Santner-Nanan, B.; Tangye, S.G.; Alexander, S.I.; Solomon, M.; Lee, S.; Nanan, R.; Fazekas de Saint Groth, B. Persistence of naive CD45RA+ regulatory T cells in adult life. Blood 2006, 107, 2830–2838. [Google Scholar] [CrossRef] [Green Version]
- Valmori, D.; Merlo, A.; Souleimanian, N.E.; Hesdorffer, C.S.; Ayyoub, M. A peripheral circulating compartment of natural naive CD4 Tregs. J. Clin. Investig. 2005, 115, 1953–1962. [Google Scholar] [CrossRef]
- Wing, J.B.; Tanaka, A.; Sakaguchi, S. Human FOXP3(+) Regulatory T Cell Heterogeneity and Function in Autoimmunity and Cancer. Immunity 2019, 50, 302–316. [Google Scholar] [CrossRef] [Green Version]
- Tan, J.; Taitz, J.; Sun, S.M.; Langford, L.; Ni, D.; Macia, L. Your Regulatory T Cells Are What You Eat: How Diet and Gut Microbiota Affect Regulatory T Cell Development. Front. Nutr. 2022, 9, 878382. [Google Scholar] [CrossRef]
- Kanamori, M.; Nakatsukasa, H.; Okada, M.; Lu, Q.; Yoshimura, A. Induced Regulatory T Cells: Their Development, Stability, and Applications. Trends Immunol. 2016, 37, 803–811. [Google Scholar] [CrossRef]
- Takimoto, T.; Wakabayashi, Y.; Sekiya, T.; Inoue, N.; Morita, R.; Ichiyama, K.; Takahashi, R.; Asakawa, M.; Muto, G.; Mori, T.; et al. Smad2 and Smad3 are redundantly essential for the TGF-beta-mediated regulation of regulatory T plasticity and Th1 development. J. Immunol. 2010, 185, 842–855. [Google Scholar] [CrossRef] [Green Version]
- Sakai, R.; Komai, K.; Iizuka-Koga, M.; Yoshimura, A.; Ito, M. Regulatory T Cells: Pathophysiological Roles and Clinical Applications. Keio J. Med. 2020, 69, 1–15. [Google Scholar] [CrossRef]
- Yadav, M.; Louvet, C.; Davini, D.; Gardner, J.M.; Martinez-Llordella, M.; Bailey-Bucktrout, S.; Anthony, B.A.; Sverdrup, F.M.; Head, R.; Kuster, D.J.; et al. Neuropilin-1 distinguishes natural and inducible regulatory T cells among regulatory T cell subsets in vivo. J. Exp. Med. 2012, 209, 1713–1722. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, H.; Ma, Y.; Dawicki, W.; Zhang, X.; Gordon, J.R. Comparison of induced versus natural regulatory T cells of the same TCR specificity for induction of tolerance to an environmental antigen. J. Immunol. 2013, 191, 1136–1143. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yoshimura, A.; Wakabayashi, Y.; Mori, T. Cellular and molecular basis for the regulation of inflammation by TGF-beta. J. Biochem. 2010, 147, 781–792. [Google Scholar] [CrossRef]
- Kashiwagi, I.; Morita, R.; Schichita, T.; Komai, K.; Saeki, K.; Matsumoto, M.; Takeda, K.; Nomura, M.; Hayashi, A.; Kanai, T.; et al. Smad2 and Smad3 Inversely Regulate TGF-beta Autoinduction in Clostridium butyricum-Activated Dendritic Cells. Immunity 2015, 43, 65–79. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kurebayashi, Y.; Baba, Y.; Minowa, A.; Nadya, N.A.; Azuma, M.; Yoshimura, A.; Koyasu, S.; Nagai, S. TGF-beta-induced phosphorylation of Akt and Foxo transcription factors negatively regulates induced regulatory T cell differentiation. Biochem. Biophys. Res. Commun. 2016, 480, 114–119. [Google Scholar] [CrossRef]
- Kretschmer, K.; Apostolou, I.; Hawiger, D.; Khazaie, K.; Nussenzweig, M.C.; von Boehmer, H. Inducing and expanding regulatory T cell populations by foreign antigen. Nat. Immunol 2005, 6, 1219–1227. [Google Scholar] [CrossRef]
- Atarashi, K.; Tanoue, T.; Shima, T.; Imaoka, A.; Kuwahara, T.; Momose, Y.; Cheng, G.; Yamasaki, S.; Saito, T.; Ohba, Y.; et al. Induction of colonic regulatory T cells by indigenous Clostridium species. Science 2011, 331, 337–341. [Google Scholar] [CrossRef] [Green Version]
- Cipolletta, D.; Feuerer, M.; Li, A.; Kamei, N.; Lee, J.; Shoelson, S.E.; Benoist, C.; Mathis, D. PPAR-gamma is a major driver of the accumulation and phenotype of adipose tissue Treg cells. Nature 2012, 486, 549–553. [Google Scholar] [CrossRef] [Green Version]
- Schiering, C.; Krausgruber, T.; Chomka, A.; Frohlich, A.; Adelmann, K.; Wohlfert, E.A.; Pott, J.; Griseri, T.; Bollrath, J.; Hegazy, A.N.; et al. The alarmin IL-33 promotes regulatory T-cell function in the intestine. Nature 2014, 513, 564–568. [Google Scholar] [CrossRef] [Green Version]
- Bovenschen, H.J.; van de Kerkhof, P.C.; van Erp, P.E.; Woestenenk, R.; Joosten, I.; Koenen, H.J. Foxp3+ regulatory T cells of psoriasis patients easily differentiate into IL-17A-producing cells and are found in lesional skin. J. Investig. Dermatol. 2011, 131, 1853–1860. [Google Scholar] [CrossRef]
- Ohnmacht, C.; Park, J.H.; Cording, S.; Wing, J.B.; Atarashi, K.; Obata, Y.; Gaboriau-Routhiau, V.; Marques, R.; Dulauroy, S.; Fedoseeva, M.; et al. Mucosal immunology. The microbiota regulates type 2 immunity through RORgammat(+) T cells. Science 2015, 349, 989–993. [Google Scholar] [CrossRef]
- Sefik, E.; Geva-Zatorsky, N.; Oh, S.; Konnikova, L.; Zemmour, D.; McGuire, A.M.; Burzyn, D.; Ortiz-Lopez, A.; Lobera, M.; Yang, J.; et al. MUCOSAL IMMUNOLOGY. Individual intestinal symbionts induce a distinct population of RORgamma(+) regulatory T cells. Science 2015, 349, 993–997. [Google Scholar] [CrossRef] [Green Version]
- Kim, K.S.; Hong, S.W.; Han, D.; Yi, J.; Jung, J.; Yang, B.G.; Lee, J.Y.; Lee, M.; Surh, C.D. Dietary antigens limit mucosal immunity by inducing regulatory T cells in the small intestine. Science 2016, 351, 858–863. [Google Scholar] [CrossRef]
- Burzyn, D.; Kuswanto, W.; Kolodin, D.; Shadrach, J.L.; Cerletti, M.; Jang, Y.; Sefik, E.; Tan, T.G.; Wagers, A.J.; Benoist, C.; et al. A special population of regulatory T cells potentiates muscle repair. Cell 2013, 155, 1282–1295. [Google Scholar] [CrossRef] [Green Version]
- Rosenblum, M.D.; Gratz, I.K.; Paw, J.S.; Lee, K.; Marshak-Rothstein, A.; Abbas, A.K. Response to self antigen imprints regulatory memory in tissues. Nature 2011, 480, 538–542. [Google Scholar] [CrossRef] [Green Version]
- Sanchez Rodriguez, R.; Pauli, M.L.; Neuhaus, I.M.; Yu, S.S.; Arron, S.T.; Harris, H.W.; Yang, S.H.; Anthony, B.A.; Sverdrup, F.M.; Krow-Lucal, E.; et al. Memory regulatory T cells reside in human skin. J. Clin. Investig. 2014, 124, 1027–1036. [Google Scholar] [CrossRef] [Green Version]
- Ali, N.; Zirak, B.; Rodriguez, R.S.; Pauli, M.L.; Truong, H.A.; Lai, K.; Ahn, R.; Corbin, K.; Lowe, M.M.; Scharschmidt, T.C.; et al. Regulatory T Cells in Skin Facilitate Epithelial Stem Cell Differentiation. Cell 2017, 169, 1119–1129.e1111. [Google Scholar] [CrossRef] [Green Version]
- Nosbaum, A.; Prevel, N.; Truong, H.A.; Mehta, P.; Ettinger, M.; Scharschmidt, T.C.; Ali, N.H.; Pauli, M.L.; Abbas, A.K.; Rosenblum, M.D. Cutting Edge: Regulatory T Cells Facilitate Cutaneous Wound Healing. J. Immunol. 2016, 196, 2010–2014. [Google Scholar] [CrossRef] [Green Version]
- Semjen, A.; Garcia-Colera, A.; Requin, J. On controlling force and time in rhythmic movement sequences: The effect of stress location. Ann. N. Y. Acad. Sci. 1984, 423, 168–182. [Google Scholar] [CrossRef]
- Shevyrev, D.; Tereshchenko, V. Treg Heterogeneity, Function, and Homeostasis. Front. Immunol. 2019, 10, 3100. [Google Scholar] [CrossRef]
- Shi, H.; Chi, H. Metabolic Control of Treg Cell Stability, Plasticity, and Tissue-Specific Heterogeneity. Front. Immunol. 2019, 10, 2716. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rueda, C.M.; Jackson, C.M.; Chougnet, C.A. Regulatory T-Cell-Mediated Suppression of Conventional T-Cells and Dendritic Cells by Different cAMP Intracellular Pathways. Front. Immunol. 2016, 7, 216. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hope, C.M.; Coates, P.T.; Carroll, R.P. Immune profiling and cancer post transplantation. World J. Nephrol. 2015, 4, 41–56. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Welzl, K.; Weinberger, B.; Kronbichler, A.; Sturm, G.; Kern, G.; Mayer, G.; Grubeck-Loebenstein, B.; Koppelstaetter, C. How immunosuppressive therapy affects T cells from kidney transplanted patients of different age: The role of latent cytomegalovirus infection. Clin. Exp. Immunol. 2014, 176, 112–119. [Google Scholar] [CrossRef] [PubMed]
- Liberman, A.C.; Budzinski, M.L.; Sokn, C.; Gobbini, R.P.; Steininger, A.; Arzt, E. Regulatory and Mechanistic Actions of Glucocorticoids on T and Inflammatory Cells. Front. Endocrinol. (Lausanne) 2018, 9, 235. [Google Scholar] [CrossRef] [PubMed]
- Campbell, D.J.; Koch, M.A. Phenotypical and functional specialization of FOXP3+ regulatory T cells. Nat. Rev. Immunol. 2011, 11, 119–130. [Google Scholar] [CrossRef] [Green Version]
- Barron, L.; Dooms, H.; Hoyer, K.K.; Kuswanto, W.; Hofmann, J.; O’Gorman, W.E.; Abbas, A.K. Cutting edge: Mechanisms of IL-2-dependent maintenance of functional regulatory T cells. J. Immunol. 2010, 185, 6426–6430. [Google Scholar] [CrossRef] [Green Version]
- Boyman, O.; Sprent, J. The role of interleukin-2 during homeostasis and activation of the immune system. Nat. Rev. Immunol. 2012, 12, 180–190. [Google Scholar] [CrossRef]
- Yu, A.; Malek, T.R. Selective availability of IL-2 is a major determinant controlling the production of CD4+CD25+Foxp3+ T regulatory cells. J. Immunol. 2006, 177, 5115–5121. [Google Scholar] [CrossRef] [Green Version]
- Sadlack, B.; Merz, H.; Schorle, H.; Schimpl, A.; Feller, A.C.; Horak, I. Ulcerative colitis-like disease in mice with a disrupted interleukin-2 gene. Cell 1993, 75, 253–261. [Google Scholar] [CrossRef]
- Willerford, D.M.; Chen, J.; Ferry, J.A.; Davidson, L.; Ma, A.; Alt, F.W. Interleukin-2 receptor alpha chain regulates the size and content of the peripheral lymphoid compartment. Immunity 1995, 3, 521–530. [Google Scholar] [CrossRef] [Green Version]
- Chinen, T.; Kannan, A.K.; Levine, A.G.; Fan, X.; Klein, U.; Zheng, Y.; Gasteiger, G.; Feng, Y.; Fontenot, J.D.; Rudensky, A.Y. An essential role for the IL-2 receptor in Treg cell function. Nat. Immunol. 2016, 17, 1322–1333. [Google Scholar] [CrossRef]
- Grossman, W.J.; Verbsky, J.W.; Barchet, W.; Colonna, M.; Atkinson, J.P.; Ley, T.J. Human T regulatory cells can use the perforin pathway to cause autologous target cell death. Immunity 2004, 21, 589–601. [Google Scholar] [CrossRef] [Green Version]
- Gondek, D.C.; Lu, L.F.; Quezada, S.A.; Sakaguchi, S.; Noelle, R.J. Cutting edge: Contact-mediated suppression by CD4+CD25+ regulatory cells involves a granzyme B-dependent, perforin-independent mechanism. J. Immunol. 2005, 174, 1783–1786. [Google Scholar] [CrossRef] [Green Version]
- Ren, X.; Ye, F.; Jiang, Z.; Chu, Y.; Xiong, S.; Wang, Y. Involvement of cellular death in TRAIL/DR5-dependent suppression induced by CD4(+)CD25(+) regulatory T cells. Cell Death Differ. 2007, 14, 2076–2084. [Google Scholar] [CrossRef] [Green Version]
- Bodmer, J.L.; Holler, N.; Reynard, S.; Vinciguerra, P.; Schneider, P.; Juo, P.; Blenis, J.; Tschopp, J. TRAIL receptor-2 signals apoptosis through FADD and caspase-8. Nat. Cell Biol. 2000, 2, 241–243. [Google Scholar] [CrossRef]
- Schmidt, A.; Oberle, N.; Krammer, P.H. Molecular mechanisms of treg-mediated T cell suppression. Front. Immunol. 2012, 3, 51. [Google Scholar] [CrossRef] [Green Version]
- Boks, M.A.; Kager-Groenland, J.R.; Haasjes, M.S.; Zwaginga, J.J.; van Ham, S.M.; ten Brinke, A. IL-10-generated tolerogenic dendritic cells are optimal for functional regulatory T cell induction—A comparative study of human clinical-applicable DC. Clin. Immunol. 2012, 142, 332–342. [Google Scholar] [CrossRef]
- Strobl, H.; Knapp, W. TGF-beta1 regulation of dendritic cells. Microbes Infect. 1999, 1, 1283–1290. [Google Scholar] [CrossRef]
- Wallet, M.A.; Sen, P.; Tisch, R. Immunoregulation of dendritic cells. Clin. Med. Res. 2005, 3, 166–175. [Google Scholar] [CrossRef]
- Safinia, N.; Scotta, C.; Vaikunthanathan, T.; Lechler, R.I.; Lombardi, G. Regulatory T Cells: Serious Contenders in the Promise for Immunological Tolerance in Transplantation. Front. Immunol. 2015, 6, 438. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thepmalee, C.; Panya, A.; Junking, M.; Chieochansin, T.; Yenchitsomanus, P.T. Inhibition of IL-10 and TGF-beta receptors on dendritic cells enhances activation of effector T-cells to kill cholangiocarcinoma cells. Hum. Vaccin. Immunother. 2018, 14, 1423–1431. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kearley, J.; Barker, J.E.; Robinson, D.S.; Lloyd, C.M. Resolution of airway inflammation and hyperreactivity after in vivo transfer of CD4+CD25+ regulatory T cells is interleukin 10 dependent. J. Exp. Med. 2005, 202, 1539–1547. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Onishi, Y.; Fehervari, Z.; Yamaguchi, T.; Sakaguchi, S. Foxp3+ natural regulatory T cells preferentially form aggregates on dendritic cells in vitro and actively inhibit their maturation. Proc. Natl. Acad. Sci. USA 2008, 105, 10113–10118. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Akkaya, B.; Oya, Y.; Akkaya, M.; Al Souz, J.; Holstein, A.H.; Kamenyeva, O.; Kabat, J.; Matsumura, R.; Dorward, D.W.; Glass, D.D.; et al. Regulatory T cells mediate specific suppression by depleting peptide-MHC class II from dendritic cells. Nat. Immunol. 2019, 20, 218–231. [Google Scholar] [CrossRef]
- Fallarino, F.; Grohmann, U.; Hwang, K.W.; Orabona, C.; Vacca, C.; Bianchi, R.; Belladonna, M.L.; Fioretti, M.C.; Alegre, M.L.; Puccetti, P. Modulation of tryptophan catabolism by regulatory T cells. Nat. Immunol. 2003, 4, 1206–1212. [Google Scholar] [CrossRef]
- Fallarino, F.; Grohmann, U.; You, S.; McGrath, B.C.; Cavener, D.R.; Vacca, C.; Orabona, C.; Bianchi, R.; Belladonna, M.L.; Volpi, C.; et al. The combined effects of tryptophan starvation and tryptophan catabolites down-regulate T cell receptor zeta-chain and induce a regulatory phenotype in naive T cells. J. Immunol. 2006, 176, 6752–6761. [Google Scholar] [CrossRef] [Green Version]
- Sharma, M.D.; Baban, B.; Chandler, P.; Hou, D.Y.; Singh, N.; Yagita, H.; Azuma, M.; Blazar, B.R.; Mellor, A.L.; Munn, D.H. Plasmacytoid dendritic cells from mouse tumor-draining lymph nodes directly activate mature Tregs via indoleamine 2,3-dioxygenase. J. Clin. Investig. 2007, 117, 2570–2582. [Google Scholar] [CrossRef] [Green Version]
- Sharma, M.D.; Hou, D.Y.; Liu, Y.; Koni, P.A.; Metz, R.; Chandler, P.; Mellor, A.L.; He, Y.; Munn, D.H. Indoleamine 2,3-dioxygenase controls conversion of Foxp3+ Tregs to TH17-like cells in tumor-draining lymph nodes. Blood 2009, 113, 6102–6111. [Google Scholar] [CrossRef] [Green Version]
- Puccetti, P.; Grohmann, U. IDO and regulatory T cells: A role for reverse signalling and non-canonical NF-kappaB activation. Nat. Rev. Immunol. 2007, 7, 817–823. [Google Scholar] [CrossRef]
- Tagami, H. Triggering factors. Clin. Dermatol. 1997, 15, 677–685. [Google Scholar] [CrossRef]
- Ho, P.; Bruce, I.N.; Silman, A.; Symmons, D.; Newman, B.; Young, H.; Griffiths, C.E.; John, S.; Worthington, J.; Barton, A. Evidence for common genetic control in pathways of inflammation for Crohn’s disease and psoriatic arthritis. Arthritis Rheum. 2005, 52, 3596–3602. [Google Scholar] [CrossRef]
- Conrad, C.; Boyman, O.; Tonel, G.; Tun-Kyi, A.; Laggner, U.; de Fougerolles, A.; Kotelianski, V.; Gardner, H.; Nestle, F.O. Alpha1beta1 integrin is crucial for accumulation of epidermal T cells and the development of psoriasis. Nat. Med. 2007, 13, 836–842. [Google Scholar] [CrossRef]
- Love, T.J.; Zhu, Y.; Zhang, Y.; Wall-Burns, L.; Ogdie, A.; Gelfand, J.M.; Choi, H.K. Obesity and the risk of psoriatic arthritis: A population-based study. Ann. Rheum. Dis. 2012, 71, 1273–1277. [Google Scholar] [CrossRef] [Green Version]
- El-Boghdady, N.A.; Ismail, M.F.; Abd-Alhameed, M.F.; Ahmed, A.S.; Ahmed, H.H. Bidirectional Association Between Psoriasis and Obesity: Benefits and Risks. J. Interferon Cytokine Res. 2018, 38, 12–19. [Google Scholar] [CrossRef]
- Ogawa, E.; Sato, Y.; Minagawa, A.; Okuyama, R. Pathogenesis of psoriasis and development of treatment. J. Dermatol. 2018, 45, 264–272. [Google Scholar] [CrossRef] [Green Version]
- Nguyen, U.D.T.; Zhang, Y.; Lu, N.; Louie-Gao, Q.; Niu, J.; Ogdie, A.; Gelfand, J.M.; LaValley, M.P.; Dubreuil, M.; Sparks, J.A.; et al. Smoking paradox in the development of psoriatic arthritis among patients with psoriasis: A population-based study. Ann. Rheum. Dis. 2018, 77, 119–123. [Google Scholar] [CrossRef]
- Gouirand, V.; Habrylo, I.; Rosenblum, M.D. Regulatory T Cells and Inflammatory Mediators in Autoimmune Disease. J. Investig. Dermatol. 2022, 142, 774–780. [Google Scholar] [CrossRef]
- Marie, J.; Kovacs, D.; Pain, C.; Jouary, T.; Cota, C.; Vergier, B.; Picardo, M.; Taieb, A.; Ezzedine, K.; Cario-Andre, M. Inflammasome activation and vitiligo/nonsegmental vitiligo progression. Br. J. Dermatol. 2014, 170, 816–823. [Google Scholar] [CrossRef]
- Sushama, S.; Dixit, N.; Gautam, R.K.; Arora, P.; Khurana, A.; Anubhuti, A. Cytokine profile (IL-2, IL-6, IL-17, IL-22, and TNF-alpha) in vitiligo-New insight into pathogenesis of disease. J. Cosmet. Dermatol. 2019, 18, 337–341. [Google Scholar] [CrossRef]
- Dong, J.; An, X.; Zhong, H.; Wang, Y.; Shang, J.; Zhou, J. Interleukin-22 participates in the inflammatory process of vitiligo. Oncotarget 2017, 8, 109161–109174. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, X.; Guo, W.; Chang, Y.; Chen, J.; Kang, P.; Yi, X.; Cui, T.; Guo, S.; Xiao, Q.; Jian, Z.; et al. Oxidative stress-induced IL-15 trans-presentation in keratinocytes contributes to CD8(+) T cells activation via JAK-STAT pathway in vitiligo. Free Radic. Biol. Med. 2019, 139, 80–91. [Google Scholar] [CrossRef] [PubMed]
- Tomaszewska, K.; Kozlowska, M.; Kaszuba, A.; Lesiak, A.; Narbutt, J.; Zalewska-Janowska, A. Increased Serum Levels of IFN-gamma, IL-1beta, and IL-6 in Patients with Alopecia Areata and Nonsegmental Vitiligo. Oxid. Med. Cell. Longev. 2020, 2020, 5693572. [Google Scholar] [CrossRef] [PubMed]
- Atwa, M.A.; Ali, S.M.M.; Youssef, N.; Mahmoud Marie, R.E. Elevated serum level of interleukin-15 in vitiligo patients and its correlation with disease severity but not activity. J. Cosmet Dermatol. 2021, 20, 2640–2644. [Google Scholar] [CrossRef]
- Custurone, P.; Di Bartolomeo, L.; Irrera, N.; Borgia, F.; Altavilla, D.; Bitto, A.; Pallio, G.; Squadrito, F.; Vaccaro, M. Role of Cytokines in Vitiligo: Pathogenesis and Possible Targets for Old and New Treatments. Int. J. Mol. Sci. 2021, 22, 11429. [Google Scholar] [CrossRef]
- Bhatia, S.; Khaitan, B.K.; Gupta, V.; Khandpur, S.; Sahni, K.; Sreenivas, V. Efficacy of NB-UVB in Progressive Versus Non-Progressive Non-Segmental Vitiligo: A Prospective Comparative Study. Indian Dermatol. Online J. 2021, 12, 701–705. [Google Scholar] [CrossRef]
- Kim, S.R.; Heaton, H.; Liu, L.Y.; King, B.A. Rapid Repigmentation of Vitiligo Using Tofacitinib Plus Low-Dose, Narrowband UV-B Phototherapy. JAMA Dermatol. 2018, 154, 370–371. [Google Scholar] [CrossRef]
- Krasimirova, E.; Velikova, T.; Ivanova-Todorova, E.; Tumangelova-Yuzeir, K.; Kalinova, D.; Boyadzhieva, V.; Stoilov, N.; Yoneva, T.; Rashkov, R.; Kyurkchiev, D. Treg/Th17 cell balance and phytohaemagglutinin activation of T lymphocytes in peripheral blood of systemic sclerosis patients. World J. Exp. Med. 2017, 7, 84–96. [Google Scholar] [CrossRef]
- Yang, X.; Yang, J.; Xing, X.; Wan, L.; Li, M. Increased frequency of Th17 cells in systemic sclerosis is related to disease activity and collagen overproduction. Arthritis Res. Ther. 2014, 16, R4. [Google Scholar] [CrossRef] [Green Version]
- Xing, X.; Li, A.; Tan, H.; Zhou, Y. IFN-gamma(+) IL-17(+) Th17 cells regulate fibrosis through secreting IL-21 in systemic scleroderma. J. Cell. Mol. Med. 2020, 24, 13600–13608. [Google Scholar] [CrossRef]
- MacDonald, K.G.; Dawson, N.A.J.; Huang, Q.; Dunne, J.V.; Levings, M.K.; Broady, R. Regulatory T cells produce profibrotic cytokines in the skin of patients with systemic sclerosis. J. Allergy Clin. Immunol. 2015, 135, 946–955.e949. [Google Scholar] [CrossRef]
- Deegan, A.E. Bacterial endocarditis. J. Am. Dent. Assoc. 1997, 128, 1628. [Google Scholar] [CrossRef]
- Frantz, C.; Cauvet, A.; Durand, A.; Gonzalez, V.; Pierre, R.; Do Cruzeiro, M.; Bailly, K.; Andrieu, M.; Orvain, C.; Avouac, J.; et al. Driving Role of Interleukin-2-Related Regulatory CD4+ T Cell Deficiency in the Development of Lung Fibrosis and Vascular Remodeling in a Mouse Model of Systemic Sclerosis. Arthritis Rheumatol. 2022, 74, 1387–1398. [Google Scholar] [CrossRef]
- Ingegnoli, F.; Ughi, N.; Mihai, C. Update on the epidemiology, risk factors, and disease outcomes of systemic sclerosis. Best Pract. Res. Clin. Rheumatol. 2018, 32, 223–240. [Google Scholar] [CrossRef]
- Bukiri, H.; Volkmann, E.R. Current advances in the treatment of systemic sclerosis. Curr. Opin. Pharmacol. 2022, 64, 102211. [Google Scholar] [CrossRef]
- Higuchi, T.; Kawaguchi, Y.; Takagi, K.; Tochimoto, A.; Ota, Y.; Katsumata, Y.; Ichida, H.; Hanaoka, M.; Kawasumi, H.; Tochihara, M.; et al. Sildenafil attenuates the fibrotic phenotype of skin fibroblasts in patients with systemic sclerosis. Clin. Immunol. 2015, 161, 333–338. [Google Scholar] [CrossRef]
- Parisi, S.; Bruzzone, M.; Centanaro Di Vittorio, C.; Lagana, A.; Peroni, C.L.; Fusaro, E. Efficacy of bosentan in the treatment of Raynaud’s phenomenon in patients with systemic sclerosis never treated with prostanoids. Reumatismo 2014, 65, 286–291. [Google Scholar] [CrossRef] [Green Version]
- Christophers, E. Psoriasis--epidemiology and clinical spectrum. Clin. Exp. Dermatol. 2001, 26, 314–320. [Google Scholar] [CrossRef]
- Ni, X.; Lai, Y. Keratinocyte: A trigger or an executor of psoriasis? J. Leukoc. Biol. 2020, 108, 485–491. [Google Scholar] [CrossRef]
- Farber, E.M.; Nall, M.L. The natural history of psoriasis in 5600 patients. Dermatologica 1974, 148, 1–18. [Google Scholar] [CrossRef]
- Chandran, V.; Raychaudhuri, S.P. Geoepidemiology and environmental factors of psoriasis and psoriatic arthritis. J. Autoimmun. 2010, 34, J314–J321. [Google Scholar] [CrossRef]
- Ghei, O.K.; Kay, W.W. Regulation of C4-dicarboxylic acid transport in Bacillus subtilis. Can. J. Microbiol. 1975, 21, 527–536. [Google Scholar] [CrossRef] [PubMed]
- Brandrup, F.; Holm, N.; Grunnet, N.; Henningsen, K.; Hansen, H.E. Psoriasis in monozygotic twins: Variations in expression in individuals with identical genetic constitution. Acta Derm. Venereol. 1982, 62, 229–236. [Google Scholar] [PubMed]
- Swanbeck, G.; Inerot, A.; Martinsson, T.; Wahlstrom, J. A population genetic study of psoriasis. Br. J. Dermatol. 1994, 131, 32–39. [Google Scholar] [CrossRef] [PubMed]
- Kimberling, W.; Dobson, R.L. The inheritance of psoriasis. J. Investig. Dermatol. 1973, 60, 538–540. [Google Scholar] [CrossRef] [Green Version]
- Harden, J.L.; Krueger, J.G.; Bowcock, A.M. The immunogenetics of Psoriasis: A comprehensive review. J. Autoimmun. 2015, 64, 66–73. [Google Scholar] [CrossRef] [Green Version]
- Sugiyama, H.; Gyulai, R.; Toichi, E.; Garaczi, E.; Shimada, S.; Stevens, S.R.; McCormick, T.S.; Cooper, K.D. Dysfunctional blood and target tissue CD4+CD25high regulatory T cells in psoriasis: Mechanism underlying unrestrained pathogenic effector T cell proliferation. J. Immunol. 2005, 174, 164–173. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Zhang, C.; Li, B.; Yu, C.; Bai, X.; Xiao, C.; Wang, L.; Dang, E.; Yang, L.; Wang, G. A novel role of IL-17A in contributing to the impaired suppressive function of Tregs in psoriasis. J. Dermatol. Sci. 2021, 101, 84–92. [Google Scholar] [CrossRef]
- Kim, C.H. FOXP3 and its role in the immune system. Adv. Exp. Med. Biol. 2009, 665, 17–29. [Google Scholar] [CrossRef]
- Georgiev, P.; Charbonnier, L.M.; Chatila, T.A. Regulatory T Cells: The Many Faces of Foxp3. J. Clin. Immunol. 2019, 39, 623–640. [Google Scholar] [CrossRef]
- von Knethen, A.; Heinicke, U.; Weigert, A.; Zacharowski, K.; Brune, B. Histone Deacetylation Inhibitors as Modulators of Regulatory T Cells. Int. J. Mol. Sci. 2020, 21, 2356. [Google Scholar] [CrossRef] [Green Version]
- Yang, L.; Li, B.; Dang, E.; Jin, L.; Fan, X.; Wang, G. Impaired function of regulatory T cells in patients with psoriasis is mediated by phosphorylation of STAT3. J. Dermatol. Sci. 2016, 81, 85–92. [Google Scholar] [CrossRef]
- Goodman, W.A.; Levine, A.D.; Massari, J.V.; Sugiyama, H.; McCormick, T.S.; Cooper, K.D. IL-6 signaling in psoriasis prevents immune suppression by regulatory T cells. J. Immunol. 2009, 183, 3170–3176. [Google Scholar] [CrossRef] [Green Version]
- Peluso, I.; Fantini, M.C.; Fina, D.; Caruso, R.; Boirivant, M.; MacDonald, T.T.; Pallone, F.; Monteleone, G. IL-21 counteracts the regulatory T cell-mediated suppression of human CD4+ T lymphocytes. J. Immunol. 2007, 178, 732–739. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Li, S.; Li, C. Clinical Features, Immunopathogenesis, and Therapeutic Strategies in Vitiligo. Clin. Rev. Allergy Immunol. 2021, 61, 299–323. [Google Scholar] [CrossRef]
- Gellatly, K.J.; Strassner, J.P.; Essien, K.; Refat, M.A.; Murphy, R.L.; Coffin-Schmitt, A.; Pandya, A.G.; Tovar-Garza, A.; Frisoli, M.L.; Fan, X.; et al. scRNA-seq of human vitiligo reveals complex networks of subclinical immune activation and a role for CCR5 in Treg function. Sci. Transl. Med. 2021, 13, eabd8995. [Google Scholar] [CrossRef]
- Mosenson, J.A.; Zloza, A.; Klarquist, J.; Barfuss, A.J.; Guevara-Patino, J.A.; Poole, I.C. HSP70i is a critical component of the immune response leading to vitiligo. Pigment Cell Melanoma Res. 2012, 25, 88–98. [Google Scholar] [CrossRef] [Green Version]
- Ahn, Y.; Seo, J.; Lee, E.J.; Kim, J.Y.; Park, M.Y.; Hwang, S.; Almurayshid, A.; Lim, B.J.; Yu, J.W.; Oh, S.H. ATP-P2X7-Induced Inflammasome Activation Contributes to Melanocyte Death and CD8(+) T-Cell Trafficking to the Skin in Vitiligo. J. Investig. Dermatol. 2020, 140, 1794–1804.e4. [Google Scholar] [CrossRef]
- Richmond, J.M.; Bangari, D.S.; Essien, K.I.; Currimbhoy, S.D.; Groom, J.R.; Pandya, A.G.; Youd, M.E.; Luster, A.D.; Harris, J.E. Keratinocyte-Derived Chemokines Orchestrate T-Cell Positioning in the Epidermis during Vitiligo and May Serve as Biomarkers of Disease. J. Investig. Dermatol. 2017, 137, 350–358. [Google Scholar] [CrossRef] [Green Version]
- Tulic, M.K.; Cavazza, E.; Cheli, Y.; Jacquel, A.; Luci, C.; Cardot-Leccia, N.; Hadhiri-Bzioueche, H.; Abbe, P.; Gesson, M.; Sormani, L.; et al. Innate lymphocyte-induced CXCR3B-mediated melanocyte apoptosis is a potential initiator of T-cell autoreactivity in vitiligo. Nat. Commun. 2019, 10, 2178. [Google Scholar] [CrossRef]
- van den Boorn, J.G.; Konijnenberg, D.; Dellemijn, T.A.; van der Veen, J.P.; Bos, J.D.; Melief, C.J.; Vyth-Dreese, F.A.; Luiten, R.M. Autoimmune destruction of skin melanocytes by perilesional T cells from vitiligo patients. J. Investig. Dermatol. 2009, 129, 2220–2232. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, S.; Zhu, G.; Yang, Y.; Jian, Z.; Guo, S.; Dai, W.; Shi, Q.; Ge, R.; Ma, J.; Liu, L.; et al. Oxidative stress drives CD8(+) T-cell skin trafficking in patients with vitiligo through CXCL16 upregulation by activating the unfolded protein response in keratinocytes. J. Allergy Clin. Immunol. 2017, 140, 177–189.e9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- van den Wijngaard, R.; Wankowicz-Kalinska, A.; Le Poole, C.; Tigges, B.; Westerhof, W.; Das, P. Local immune response in skin of generalized vitiligo patients. Destruction of melanocytes is associated with the prominent presence of CLA+ T cells at the perilesional site. Lab. Investig. 2000, 80, 1299–1309. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mandelcorn-Monson, R.L.; Shear, N.H.; Yau, E.; Sambhara, S.; Barber, B.H.; Spaner, D.; DeBenedette, M.A. Cytotoxic T lymphocyte reactivity to gp100, MelanA/MART-1, and tyrosinase, in HLA-A2-positive vitiligo patients. J. Investig. Dermatol. 2003, 121, 550–556. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hegab, D.S.; Attia, M.A. Decreased Circulating T Regulatory Cells in Egyptian Patients with Nonsegmental Vitiligo: Correlation with Disease Activity. Dermatol. Res. Pract. 2015, 2015, 145409. [Google Scholar] [CrossRef]
- Eby, J.M.; Kang, H.K.; Tully, S.T.; Bindeman, W.E.; Peiffer, D.S.; Chatterjee, S.; Mehrotra, S.; Le Poole, I.C. CCL22 to Activate Treg Migration and Suppress Depigmentation in Vitiligo. J. Investig. Dermatol. 2015, 135, 1574–1580. [Google Scholar] [CrossRef] [Green Version]
- Lili, Y.; Yi, W.; Ji, Y.; Yue, S.; Weimin, S.; Ming, L. Global activation of CD8+ cytotoxic T lymphocytes correlates with an impairment in regulatory T cells in patients with generalized vitiligo. PLoS ONE 2012, 7, e37513. [Google Scholar] [CrossRef] [Green Version]
- Zhou, L.; Li, K.; Shi, Y.L.; Hamzavi, I.; Gao, T.W.; Henderson, M.; Huggins, R.H.; Agbai, O.; Mahmoud, B.; Mi, X.; et al. Systemic analyses of immunophenotypes of peripheral T cells in non-segmental vitiligo: Implication of defective natural killer T cells. Pigment Cell Melanoma Res. 2012, 25, 602–611. [Google Scholar] [CrossRef] [Green Version]
- Klarquist, J.; Denman, C.J.; Hernandez, C.; Wainwright, D.A.; Strickland, F.M.; Overbeck, A.; Mehrotra, S.; Nishimura, M.I.; Le Poole, I.C. Reduced skin homing by functional Treg in vitiligo. Pigment Cell Melanoma Res. 2010, 23, 276–286. [Google Scholar] [CrossRef]
- Ben Ahmed, M.; Zaraa, I.; Rekik, R.; Elbeldi-Ferchiou, A.; Kourda, N.; Belhadj Hmida, N.; Abdeladhim, M.; Karoui, O.; Ben Osman, A.; Mokni, M.; et al. Functional defects of peripheral regulatory T lymphocytes in patients with progressive vitiligo. Pigment Cell Melanoma Res. 2012, 25, 99–109. [Google Scholar] [CrossRef]
- Moftah, N.H.; El-Barbary, R.A.; Ismail, M.A.; Ali, N.A. Effect of narrow band-ultraviolet B on CD4(+) CD25(high) FoxP3(+) T-lymphocytes in the peripheral blood of vitiligo patients. Photodermatol. Photoimmunol. Photomed. 2014, 30, 254–261. [Google Scholar] [CrossRef]
- Ujiie, H. Regulatory T cells in autoimmune skin diseases. Exp. Dermatol. 2019, 28, 642–646. [Google Scholar] [CrossRef] [Green Version]
- Miller, M.C.; Mayo, K.H. Chemokines from a Structural Perspective. Int. J. Mol. Sci. 2017, 18, 88. [Google Scholar] [CrossRef] [Green Version]
- Le Poole, I.C.; Mehrotra, S. Replenishing Regulatory T Cells to Halt Depigmentation in Vitiligo. J. Investig. Dermatol. Symp. Proc. 2017, 18, S38–S45. [Google Scholar] [CrossRef] [Green Version]
- Klarquist, J.; Tobin, K.; Farhangi Oskuei, P.; Henning, S.W.; Fernandez, M.F.; Dellacecca, E.R.; Navarro, F.C.; Eby, J.M.; Chatterjee, S.; Mehrotra, S.; et al. Ccl22 Diverts T Regulatory Cells and Controls the Growth of Melanoma. Cancer Res. 2016, 76, 6230–6240. [Google Scholar] [CrossRef] [Green Version]
- Tembhre, M.K.; Parihar, A.S.; Sharma, V.K.; Sharma, A.; Chattopadhyay, P.; Gupta, S. Alteration in regulatory T cells and programmed cell death 1-expressing regulatory T cells in active generalized vitiligo and their clinical correlation. Br. J. Dermatol. 2015, 172, 940–950. [Google Scholar] [CrossRef]
- Essien, K.I.; Katz, E.L.; Strassner, J.P.; Harris, J.E. Regulatory T Cells Require CCR6 for Skin Migration and Local Suppression of Vitiligo. J. Investig. Dermatol. 2022, 142, 3158–3166. [Google Scholar] [CrossRef]
- Chen, J.; Wang, X.; Cui, T.; Ni, Q.; Zhang, Q.; Zou, D.; He, K.; Wu, W.; Ma, J.; Wang, Y.; et al. Th1-like Treg in vitiligo: An incompetent regulator in immune tolerance. J. Autoimmun. 2022, 131, 102859. [Google Scholar] [CrossRef]
- Wankowicz-Kalinska, A.; van den Wijngaard, R.M.; Tigges, B.J.; Westerhof, W.; Ogg, G.S.; Cerundolo, V.; Storkus, W.J.; Das, P.K. Immunopolarization of CD4+ and CD8+ T cells to Type-1-like is associated with melanocyte loss in human vitiligo. Lab. Investig. 2003, 83, 683–695. [Google Scholar] [CrossRef] [Green Version]
- Ferreira, C.; Barros, L.; Baptista, M.; Blankenhaus, B.; Barros, A.; Figueiredo-Campos, P.; Konjar, S.; Laine, A.; Kamenjarin, N.; Stojanovic, A.; et al. Type 1 Treg cells promote the generation of CD8(+) tissue-resident memory T cells. Nat. Immunol. 2020, 21, 766–776. [Google Scholar] [CrossRef]
- Richmond, J.M.; Strassner, J.P.; Rashighi, M.; Agarwal, P.; Garg, M.; Essien, K.I.; Pell, L.S.; Harris, J.E. Resident Memory and Recirculating Memory T Cells Cooperate to Maintain Disease in a Mouse Model of Vitiligo. J. Investig. Dermatol. 2019, 139, 769–778. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Riding, R.L.; Harris, J.E. The Role of Memory CD8(+) T Cells in Vitiligo. J. Immunol. 2019, 203, 11–19. [Google Scholar] [CrossRef] [PubMed]
- Boniface, K.; Jacquemin, C.; Darrigade, A.S.; Dessarthe, B.; Martins, C.; Boukhedouni, N.; Vernisse, C.; Grasseau, A.; Thiolat, D.; Rambert, J.; et al. Vitiligo Skin Is Imprinted with Resident Memory CD8 T Cells Expressing CXCR3. J. Investig. Dermatol. 2018, 138, 355–364. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fett, N. Scleroderma: Nomenclature, etiology, pathogenesis, prognosis, and treatments: Facts and controversies. Clin. Dermatol. 2013, 31, 432–437. [Google Scholar] [CrossRef] [PubMed]
- Cutolo, M.; Soldano, S.; Smith, V. Pathophysiology of systemic sclerosis: Current understanding and new insights. Expert Rev. Clin. Immunol. 2019, 15, 753–764. [Google Scholar] [CrossRef] [Green Version]
- Rongioletti, F.; Kaiser, F.; Cinotti, E.; Metze, D.; Battistella, M.; Calzavara-Pinton, P.G.; Damevska, K.; Girolomoni, G.; Andre, J.; Perrot, J.L.; et al. Scleredema. A multicentre study of characteristics, comorbidities, course and therapy in 44 patients. J. Eur. Acad. Dermatol. Venereol. 2015, 29, 2399–2404. [Google Scholar] [CrossRef]
- Nihtyanova, S.I.; Tang, E.C.; Coghlan, J.G.; Wells, A.U.; Black, C.M.; Denton, C.P. Improved survival in systemic sclerosis is associated with better ascertainment of internal organ disease: A retrospective cohort study. QJM 2010, 103, 109–115. [Google Scholar] [CrossRef]
- Tyndall, A.J.; Bannert, B.; Vonk, M.; Airo, P.; Cozzi, F.; Carreira, P.E.; Bancel, D.F.; Allanore, Y.; Muller-Ladner, U.; Distler, O.; et al. Causes and risk factors for death in systemic sclerosis: A study from the EULAR Scleroderma Trials and Research (EUSTAR) database. Ann. Rheum. Dis. 2010, 69, 1809–1815. [Google Scholar] [CrossRef] [Green Version]
- Romano, E.; Manetti, M.; Guiducci, S.; Ceccarelli, C.; Allanore, Y.; Matucci-Cerinic, M. The genetics of systemic sclerosis: An update. Clin. Exp. Rheumatol. 2011, 29, S75–S86. [Google Scholar]
- Katsumoto, T.R.; Whitfield, M.L.; Connolly, M.K. The pathogenesis of systemic sclerosis. Annu. Rev. Pathol. 2011, 6, 509–537. [Google Scholar] [CrossRef]
- Radstake, T.R.; van Bon, L.; Broen, J.; Wenink, M.; Santegoets, K.; Deng, Y.; Hussaini, A.; Simms, R.; Cruikshank, W.W.; Lafyatis, R. Increased frequency and compromised function of T regulatory cells in systemic sclerosis (SSc) is related to a diminished CD69 and TGFbeta expression. PLoS ONE 2009, 4, e5981. [Google Scholar] [CrossRef]
- Fenoglio, D.; Battaglia, F.; Parodi, A.; Stringara, S.; Negrini, S.; Panico, N.; Rizzi, M.; Kalli, F.; Conteduca, G.; Ghio, M.; et al. Alteration of Th17 and Treg cell subpopulations co-exist in patients affected with systemic sclerosis. Clin. Immunol. 2011, 139, 249–257. [Google Scholar] [CrossRef]
- Antiga, E.; Quaglino, P.; Bellandi, S.; Volpi, W.; Del Bianco, E.; Comessatti, A.; Osella-Abate, S.; De Simone, C.; Marzano, A.; Bernengo, M.G.; et al. Regulatory T cells in the skin lesions and blood of patients with systemic sclerosis and morphoea. Br. J. Dermatol. 2010, 162, 1056–1063. [Google Scholar] [CrossRef]
- Zhou, Y.; Hou, W.; Xu, K.; Han, D.; Jiang, C.; Mou, K.; Li, Y.; Meng, L.; Lu, S. The elevated expression of Th17-related cytokines and receptors is associated with skin lesion severity in early systemic sclerosis. Hum. Immunol. 2015, 76, 22–29. [Google Scholar] [CrossRef]
- Nakashima, T.; Jinnin, M.; Yamane, K.; Honda, N.; Kajihara, I.; Makino, T.; Masuguchi, S.; Fukushima, S.; Okamoto, Y.; Hasegawa, M.; et al. Impaired IL-17 signaling pathway contributes to the increased collagen expression in scleroderma fibroblasts. J. Immunol. 2012, 188, 3573–3583. [Google Scholar] [CrossRef] [Green Version]
- Brembilla, N.C.; Montanari, E.; Truchetet, M.E.; Raschi, E.; Meroni, P.; Chizzolini, C. Th17 cells favor inflammatory responses while inhibiting type I collagen deposition by dermal fibroblasts: Differential effects in healthy and systemic sclerosis fibroblasts. Arthritis Res. Ther. 2013, 15, R151. [Google Scholar] [CrossRef] [Green Version]
- Lei, L.; Zhao, C.; Qin, F.; He, Z.Y.; Wang, X.; Zhong, X.N. Th17 cells and IL-17 promote the skin and lung inflammation and fibrosis process in a bleomycin-induced murine model of systemic sclerosis. Clin. Exp. Rheumatol. 2016, 34 (Suppl. S100), 14–22. [Google Scholar]
- Carvalheiro, T.; Affandi, A.J.; Malvar-Fernandez, B.; Dullemond, I.; Cossu, M.; Ottria, A.; Mertens, J.S.; Giovannone, B.; Bonte-Mineur, F.; Kok, M.R.; et al. Induction of Inflammation and Fibrosis by Semaphorin 4A in Systemic Sclerosis. Arthritis Rheumatol. 2019, 71, 1711–1722. [Google Scholar] [CrossRef] [Green Version]
- Almanzar, G.; Klein, M.; Schmalzing, M.; Hilligardt, D.; El Hajj, N.; Kneitz, H.; Wild, V.; Rosenwald, A.; Benoit, S.; Hamm, H.; et al. Disease Manifestation and Inflammatory Activity as Modulators of Th17/Treg Balance and RORC/FoxP3 Methylation in Systemic Sclerosis. Int. Arch. Allergy Immunol. 2016, 171, 141–154. [Google Scholar] [CrossRef]
- Ugor, E.; Simon, D.; Almanzar, G.; Pap, R.; Najbauer, J.; Nemeth, P.; Balogh, P.; Prelog, M.; Czirjak, L.; Berki, T. Increased proportions of functionally impaired regulatory T cell subsets in systemic sclerosis. Clin. Immunol. 2017, 184, 54–62. [Google Scholar] [CrossRef]
- Hamilton, R.F., Jr.; Parsley, E.; Holian, A. Alveolar macrophages from systemic sclerosis patients: Evidence for IL-4-mediated phenotype changes. Am. J. Physiol. Lung Cell Mol. Physiol. 2004, 286, L1202–L1209. [Google Scholar] [CrossRef] [PubMed]
- Fuschiotti, P.; Medsger, T.A., Jr.; Morel, P.A. Effector CD8+ T cells in systemic sclerosis patients produce abnormally high levels of interleukin-13 associated with increased skin fibrosis. Arthritis Rheum. 2009, 60, 1119–1128. [Google Scholar] [CrossRef] [PubMed]
- Gasparini, G.; Cozzani, E.; Parodi, A. Interleukin-4 and interleukin-13 as possible therapeutic targets in systemic sclerosis. Cytokine 2020, 125, 154799. [Google Scholar] [CrossRef] [PubMed]
- Saigusa, R.; Asano, Y.; Taniguchi, T.; Hirabayashi, M.; Nakamura, K.; Miura, S.; Yamashita, T.; Takahashi, T.; Ichimura, Y.; Toyama, T.; et al. Fli1-haploinsufficient dermal fibroblasts promote skin-localized transdifferentiation of Th2-like regulatory T cells. Arthritis Res. Ther. 2018, 20, 23. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Slobodin, G.; Ahmad, M.S.; Rosner, I.; Peri, R.; Rozenbaum, M.; Kessel, A.; Toubi, E.; Odeh, M. Regulatory T cells (CD4(+)CD25(bright)FoxP3(+)) expansion in systemic sclerosis correlates with disease activity and severity. Cell Immunol. 2010, 261, 77–80. [Google Scholar] [CrossRef]
- Rice, L.M.; Padilla, C.M.; McLaughlin, S.R.; Mathes, A.; Ziemek, J.; Goummih, S.; Nakerakanti, S.; York, M.; Farina, G.; Whitfield, M.L.; et al. Fresolimumab treatment decreases biomarkers and improves clinical symptoms in systemic sclerosis patients. J. Clin. Investig. 2015, 125, 2795–2807. [Google Scholar] [CrossRef]
- Marino, J.; Paster, J.; Benichou, G. Allorecognition by T Lymphocytes and Allograft Rejection. Front. Immunol. 2016, 7, 582. [Google Scholar] [CrossRef] [Green Version]
- Ingulli, E. Mechanism of cellular rejection in transplantation. Pediatr. Nephrol. 2010, 25, 61–74. [Google Scholar] [CrossRef] [Green Version]
- Starzl, T.E. History of clinical transplantation. World J. Surg. 2000, 24, 759–782. [Google Scholar] [CrossRef] [Green Version]
- Pilch, N.A.; Bowman, L.J.; Taber, D.J. Immunosuppression trends in solid organ transplantation: The future of individualization, monitoring, and management. Pharmacotherapy 2021, 41, 119–131. [Google Scholar] [CrossRef]
- Zdanowicz, M.M. The pharmacology of immunosuppression. Am. J. Pharm. Educ. 2009, 73, 144. [Google Scholar] [CrossRef] [Green Version]
- Dantal, J.; Soulillou, J.P. Immunosuppressive drugs and the risk of cancer after organ transplantation. N. Engl. J. Med. 2005, 352, 1371–1373. [Google Scholar] [CrossRef]
- Busnach, G.; Piselli, P.; Arbustini, E.; Baccarani, U.; Burra, P.; Carrieri, M.P.; Citterio, F.; De Juli, E.; Bellelli, S.; Pradier, C.; et al. Immunosuppression and cancer: A comparison of risks in recipients of organ transplants and in HIV-positive individuals. Transplant. Proc. 2006, 38, 3533–3535. [Google Scholar] [CrossRef]
- Furukawa, A.; Wisel, S.A.; Tang, Q. Impact of Immune-Modulatory Drugs on Regulatory T Cell. Transplantation 2016, 100, 2288–2300. [Google Scholar] [CrossRef] [Green Version]
- Bernaldo-de-Quiros, E.; Lopez-Abente, J.; Camino, M.; Gil, N.; Panadero, E.; Lopez-Esteban, R.; Martinez-Bonet, M.; Pion, M.; Correa-Rocha, R. The Presence of a Marked Imbalance Between Regulatory T Cells and Effector T Cells Reveals That Tolerance Mechanisms Could Be Compromised in Heart Transplant Children. Transplant. Direct 2021, 7, e693. [Google Scholar] [CrossRef]
- Okeke, E.B.; Uzonna, J.E. The Pivotal Role of Regulatory T Cells in the Regulation of Innate Immune Cells. Front. Immunol. 2019, 10, 680. [Google Scholar] [CrossRef] [Green Version]
- Shaban, E.; Bayliss, G.; Malhotra, D.K.; Shemin, D.; Wang, L.J.; Gohh, R.; Dworkin, L.D.; Gong, R. Targeting Regulatory T Cells for Transplant Tolerance: New Insights and Future Perspectives. Kidney Dis. (Basel) 2018, 4, 205–213. [Google Scholar] [CrossRef]
- Carroll, R.P.; Segundo, D.S.; Hollowood, K.; Marafioti, T.; Clark, T.G.; Harden, P.N.; Wood, K.J. Immune phenotype predicts risk for posttransplantation squamous cell carcinoma. J. Am. Soc. Nephrol. 2010, 21, 713–722. [Google Scholar] [CrossRef] [Green Version]
- Shimizu, J.; Yamazaki, S.; Sakaguchi, S. Induction of tumor immunity by removing CD25+CD4+ T cells: A common basis between tumor immunity and autoimmunity. J. Immunol. 1999, 163, 5211–5218. [Google Scholar] [CrossRef]
- Gliwinski, M.; Piotrowska, M.; Iwaszkiewicz-Grzes, D.; Urban-Wojciuk, Z.; Trzonkowski, P. Therapy with CD4(+)CD25(+) T regulatory cells—Should we be afraid of cancer? Contemp. Oncol. (Pozn) 2019, 23, 1–6. [Google Scholar] [CrossRef]
- Biller, B.J.; Guth, A.; Burton, J.H.; Dow, S.W. Decreased ratio of CD8+ T cells to regulatory T cells associated with decreased survival in dogs with osteosarcoma. J. Vet. Intern. Med. 2010, 24, 1118–1123. [Google Scholar] [CrossRef] [PubMed]
- Facciabene, A.; Motz, G.T.; Coukos, G. T-regulatory cells: Key players in tumor immune escape and angiogenesis. Cancer Res. 2012, 72, 2162–2171. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.L.; Fang, J.H.; Lai, M.D.; Shan, Y.S. Depletion of CD4(+)CD25(+) regulatory T cells can promote local immunity to suppress tumor growth in benzo[a]pyrene-induced forestomach carcinoma. World J. Gastroenterol. 2008, 14, 5797–5809. [Google Scholar] [CrossRef] [PubMed]
- Tang, Q.; Bluestone, J.A. Regulatory T-cell therapy in transplantation: Moving to the clinic. Cold Spring Harb. Perspect. Med. 2013, 3, a015552. [Google Scholar] [CrossRef] [PubMed]
- Shang, B.; Liu, Y.; Jiang, S.J.; Liu, Y. Prognostic value of tumor-infiltrating FoxP3+ regulatory T cells in cancers: A systematic review and meta-analysis. Sci. Rep. 2015, 5, 15179. [Google Scholar] [CrossRef] [Green Version]
- Garrett, G.L.; Blanc, P.D.; Boscardin, J.; Lloyd, A.A.; Ahmed, R.L.; Anthony, T.; Bibee, K.; Breithaupt, A.; Cannon, J.; Chen, A.; et al. Incidence of and Risk Factors for Skin Cancer in Organ Transplant Recipients in the United States. JAMA Dermatol. 2017, 153, 296–303. [Google Scholar] [CrossRef] [Green Version]
- Tam, S.; Yao, C.; Amit, M.; Gajera, M.; Luo, X.; Treistman, R.; Khanna, A.; Aashiq, M.; Nagarajan, P.; Bell, D.; et al. Association of Immunosuppression With Outcomes of Patients With Cutaneous Squamous Cell Carcinoma of the Head and Neck. JAMA Otolaryngol. Head Neck Surg. 2020, 146, 128–135. [Google Scholar] [CrossRef]
- Blue, E.D.; Freeman, S.C.; Lobl, M.B.; Clarey, D.D.; Fredrick, R.L.; Wysong, A.; Whitley, M.J. Cutaneous Squamous Cell Carcinoma Arising in Immunosuppressed Patients: A Systematic Review of Tumor Profiling Studies. JID Innov. 2022, 2, 100126. [Google Scholar] [CrossRef]
- Euvrard, S.; Kanitakis, J.; Claudy, A. Skin cancers after organ transplantation. N. Engl. J. Med 2003, 348, 1681–1691. [Google Scholar] [CrossRef] [Green Version]
- Bibee, K.; Swartz, A.; Sridharan, S.; Kurten, C.H.L.; Wessel, C.B.; Skinner, H.; Zandberg, D.P. Cutaneous squamous cell carcinoma in the organ transplant recipient. Oral Oncol. 2020, 103, 104562. [Google Scholar] [CrossRef]
- Lindelof, B.; Sigurgeirsson, B.; Gabel, H.; Stern, R.S. Incidence of skin cancer in 5356 patients following organ transplantation. Br. J. Dermatol. 2000, 143, 513–519. [Google Scholar]
- Tessari, G.; Naldi, L.; Boschiero, L.; Nacchia, F.; Fior, F.; Forni, A.; Rugiu, C.; Faggian, G.; Sassi, F.; Gotti, E.; et al. Incidence and clinical predictors of a subsequent nonmelanoma skin cancer in solid organ transplant recipients with a first nonmelanoma skin cancer: A multicenter cohort study. Arch. Dermatol. 2010, 146, 294–299. [Google Scholar] [CrossRef] [Green Version]
- Moloney, F.J.; Comber, H.; O’Lorcain, P.; O’Kelly, P.; Conlon, P.J.; Murphy, G.M. A population-based study of skin cancer incidence and prevalence in renal transplant recipients. Br. J. Dermatol. 2006, 154, 498–504. [Google Scholar] [CrossRef]
- Sherston, S.N.; Vogt, K.; Schlickeiser, S.; Sawitzki, B.; Harden, P.N.; Wood, K.J. Demethylation of the TSDR is a marker of squamous cell carcinoma in transplant recipients. Am. J. Transplant. 2014, 14, 2617–2622. [Google Scholar] [CrossRef] [Green Version]
- Frazzette, N.; Khodadadi-Jamayran, A.; Doudican, N.; Santana, A.; Felsen, D.; Pavlick, A.C.; Tsirigos, A.; Carucci, J.A. Decreased cytotoxic T cells and TCR clonality in organ transplant recipients with squamous cell carcinoma. NPJ Precis. Oncol. 2020, 4, 13. [Google Scholar] [CrossRef]
- Kim, H.J.; Verbinnen, B.; Tang, X.; Lu, L.; Cantor, H. Inhibition of follicular T-helper cells by CD8(+) regulatory T cells is essential for self tolerance. Nature 2010, 467, 328–332. [Google Scholar] [CrossRef] [Green Version]
- Saligrama, N.; Zhao, F.; Sikora, M.J.; Serratelli, W.S.; Fernandes, R.A.; Louis, D.M.; Yao, W.; Ji, X.; Idoyaga, J.; Mahajan, V.B.; et al. Opposing T cell responses in experimental autoimmune encephalomyelitis. Nature 2019, 572, 481–487. [Google Scholar] [CrossRef]
- Kanitakis, J.; Alhaj-Ibrahim, L.; Euvrard, S.; Claudy, A. Basal cell carcinomas developing in solid organ transplant recipients: Clinicopathologic study of 176 cases. Arch. Dermatol. 2003, 139, 1133–1137. [Google Scholar] [CrossRef] [Green Version]
- Omland, S.H.; Nielsen, P.S.; Gjerdrum, L.M.; Gniadecki, R. Immunosuppressive Environment in Basal Cell Carcinoma: The Role of Regulatory T Cells. Acta Derm. Venereol. 2016, 96, 917–921. [Google Scholar] [CrossRef] [Green Version]
- Adolphe, C.; Xue, A.; Fard, A.T.; Genovesi, L.A.; Yang, J.; Wainwright, B.J. Genetic and functional interaction network analysis reveals global enrichment of regulatory T cell genes influencing basal cell carcinoma susceptibility. Genome Med. 2021, 13, 19. [Google Scholar] [CrossRef]
- Anaya, J.M.; Shoenfeld, Y.; Rojas-Villarraga, A.; Levy, R.A.; Cervera, R. (Eds.) Autoimmunity: From Bench to Bedside; El Rosario University Press: Bogota, Colombia, 2013. [Google Scholar]
- Vesely, M.D. Getting under the Skin: Targeting Cutaneous Autoimmune Disease. Yale J. Biol. Med. 2020, 93, 197–206. [Google Scholar] [PubMed]
- Fishman, J.A.; Rubin, R.H. Infection in organ-transplant recipients. N. Engl. J. Med. 1998, 338, 1741–1751. [Google Scholar] [CrossRef] [PubMed]
- Gutierrez-Dalmau, A.; Campistol, J.M. Immunosuppressive therapy and malignancy in organ transplant recipients: A systematic review. Drugs 2007, 67, 1167–1198. [Google Scholar] [CrossRef] [PubMed]
- Penn, I.; Starzl, T.E. Proceedings: The effect of immunosuppression on cancer. Proc. Natl. Cancer Conf. 1972, 7, 425–436. [Google Scholar] [PubMed]
- Siegel, J.P.; Sharon, M.; Smith, P.L.; Leonard, W.J. The Il-2 Receptor Beta-Chain (P70)—Role in Mediating Signals for Lak, Nk, and Proliferative Activities. Science 1987, 238, 75–78. [Google Scholar] [CrossRef]
- Shevach, E.M. Mechanisms of Foxp3(+) T Regulatory Cell-Mediated Suppression. Immunity 2009, 30, 636–645. [Google Scholar] [CrossRef] [Green Version]
- Saadoun, D.; Rosenzwajg, M.; Joly, F.; Six, A.; Carrat, F.; Thibault, V.; Sene, D.; Cacoub, P.; Klatzmann, D. Regulatory T-cell responses to low-dose interleukin-2 in HCV-induced vasculitis. N. Engl. J. Med. 2011, 365, 2067–2077. [Google Scholar] [CrossRef]
- Klatzmann, D.; Abbas, A.K. The promise of low-dose interleukin-2 therapy for autoimmune and inflammatory diseases. Nat. Rev. Immunol. 2015, 15, 283–294. [Google Scholar] [CrossRef]
- Yu, A.; Zhu, L.; Altman, N.H.; Malek, T.R. A low interleukin-2 receptor signaling threshold supports the development and homeostasis of T regulatory cells. Immunity 2009, 30, 204–217. [Google Scholar] [CrossRef] [Green Version]
- Boyman, O.; Kovar, M.; Rubinstein, M.P.; Surh, C.D.; Sprent, J. Selective stimulation of T cell subsets with antibody-cytokine immune complexes. Science 2006, 311, 1924–1927. [Google Scholar] [CrossRef] [Green Version]
- Pilat, N.; Sprent, J. Treg Therapies Revisited: Tolerance Beyond Deletion. Front. Immunol. 2020, 11, 622810. [Google Scholar] [CrossRef]
- Castela, E.; Le Duff, F.; Butori, C.; Ticchioni, M.; Hofman, P.; Bahadoran, P.; Lacour, J.P.; Passeron, T. Effects of low-dose recombinant interleukin 2 to promote T-regulatory cells in alopecia areata. JAMA Dermatol. 2014, 150, 748–751. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rosenzwajg, M.; Lorenzon, R.; Cacoub, P.; Pham, H.P.; Pitoiset, F.; El Soufi, K.; Ribet, C.; Bernard, C.; Aractingi, S.; Banneville, B.; et al. Immunological and clinical effects of low-dose interleukin-2 across 11 autoimmune diseases in a single, open clinical trial. Ann. Rheum. Dis. 2019, 78, 209–217. [Google Scholar] [CrossRef]
- Goschl, L.; Scheinecker, C.; Bonelli, M. Treg cells in autoimmunity: From identification to Treg-based therapies. Semin. Immunopathol. 2019, 41, 301–314. [Google Scholar] [CrossRef]
- Owen, C.M.; Harrison, P.V. Successful treatment of severe psoriasis with basiliximab, an interleukin-2 receptor monoclonal antibody. Clin. Exp. Dermatol. 2000, 25, 195–197. [Google Scholar] [CrossRef]
- Salim, A.; Emerson, R.M.; Dalziel, K.L. Successful treatment of severe generalized pustular psoriasis with basiliximab (interleukin-2 receptor blocker). Br. J. Dermatol. 2000, 143, 1121–1122. [Google Scholar] [CrossRef]
- Krueger, J.G.; Walters, I.B.; Miyazawa, M.; Gilleaudeau, P.; Hakimi, J.; Light, S.; Sherr, A.; Gottlieb, A.B. Successful in vivo blockade of CD25 (high-affinity interleukin 2 receptor) on T cells by administration of humanized anti-Tac antibody to patients with psoriasis. J. Am. Acad. Dermatol. 2000, 43, 448–458. [Google Scholar] [CrossRef]
- Tahvildari, M.; Dana, R. Low-Dose IL-2 Therapy in Transplantation, Autoimmunity, and Inflammatory Diseases. J. Immunol. 2019, 203, 2749–2755. [Google Scholar] [CrossRef]
- Yu, Z.; Cheng, H.; Ding, T.; Liang, Y.; Yan, C.; Gao, C.; Wen, H. Absolute decrease in regulatory T cells and low-dose interleukin-2 therapy: Restoring and expanding regulatory T cells to treat systemic sclerosis: A 24-week study. Clin. Exp. Dermatol. 2022, 47, 2188–2195. [Google Scholar] [CrossRef] [PubMed]
- Ali, H.; Mohiuddin, A.; Sharma, A.; Shaheen, I.; Kim, J.J.; El Kosi, M.; Halawa, A. Implication of interleukin-2 receptor antibody induction therapy in standard risk renal transplant in the tacrolimus era: A meta-analysis. Clin. Kidney J. 2019, 12, 592–599. [Google Scholar] [CrossRef]
- Evans, R.D.R.; Lan, J.H.; Kadatz, M.; Brar, S.; Chang, D.T.; McMichael, L.; Gill, J.; Gill, J.S. Use and Outcomes of Induction Therapy in Well-Matched Kidney Transplant Recipients. Clin. J. Am. Soc. Nephrol. 2022, 17, 271–279. [Google Scholar] [CrossRef] [PubMed]
- Trzonkowski, P.; Bieniaszewska, M.; Juscinska, J.; Dobyszuk, A.; Krzystyniak, A.; Marek, N.; Mysliwska, J.; Hellmann, A. First-in-man clinical results of the treatment of patients with graft versus host disease with human ex vivo expanded CD4+CD25+CD127- T regulatory cells. Clin. Immunol. 2009, 133, 22–26. [Google Scholar] [CrossRef]
- Di Ianni, M.; Falzetti, F.; Carotti, A.; Terenzi, A.; Castellino, F.; Bonifacio, E.; Del Papa, B.; Zei, T.; Ostini, R.I.; Cecchini, D.; et al. Tregs prevent GVHD and promote immune reconstitution in HLA-haploidentical transplantation. Blood 2011, 117, 3921–3928. [Google Scholar] [CrossRef]
- Brunstein, C.G.; Miller, J.S.; Cao, Q.; McKenna, D.H.; Hippen, K.L.; Curtsinger, J.; Defor, T.; Levine, B.L.; June, C.H.; Rubinstein, P.; et al. Infusion of ex vivo expanded T regulatory cells in adults transplanted with umbilical cord blood: Safety profile and detection kinetics. Blood 2011, 117, 1061–1070. [Google Scholar] [CrossRef] [PubMed]
- Theil, A.; Tuve, S.; Oelschlagel, U.; Maiwald, A.; Dohler, D.; Ossmann, D.; Zenkel, A.; Wilhelm, C.; Middeke, J.M.; Shayegi, N.; et al. Adoptive transfer of allogeneic regulatory T cells into patients with chronic graft-versus-host disease. Cytotherapy 2015, 17, 473–486. [Google Scholar] [CrossRef] [PubMed]
- Bluestone, J.A.; Buckner, J.H.; Fitch, M.; Gitelman, S.E.; Gupta, S.; Hellerstein, M.K.; Herold, K.C.; Lares, A.; Lee, M.R.; Li, K.; et al. Type 1 diabetes immunotherapy using polyclonal regulatory T cells. Sci. Transl. Med. 2015, 7, 315ra189. [Google Scholar] [CrossRef] [Green Version]
- Chatterjee, S.; Eby, J.M.; Al-Khami, A.A.; Soloshchenko, M.; Kang, H.K.; Kaur, N.; Naga, O.S.; Murali, A.; Nishimura, M.I.; Caroline Le Poole, I.; et al. A quantitative increase in regulatory T cells controls development of vitiligo. J. Investig. Dermatol. 2014, 134, 1285–1294. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Q.; Lu, W.; Liang, C.L.; Chen, Y.; Liu, H.; Qiu, F.; Dai, Z. Chimeric Antigen Receptor (CAR) Treg: A Promising Approach to Inducing Immunological Tolerance. Front. Immunol. 2018, 9, 2359. [Google Scholar] [CrossRef] [Green Version]
- Boardman, D.; Maher, J.; Lechler, R.; Smyth, L.; Lombardi, G. Antigen-specificity using chimeric antigen receptors: The future of regulatory T-cell therapy? Biochem. Soc. Trans. 2016, 44, 342–348. [Google Scholar] [CrossRef]
- Gross, G.; Waks, T.; Eshhar, Z. Expression of immunoglobulin-T-cell receptor chimeric molecules as functional receptors with antibody-type specificity. Proc. Natl. Acad. Sci. USA 1989, 86, 10024–10028. [Google Scholar] [CrossRef] [Green Version]
- Horton, H.; McMorrow, I.; Burke, B. Independent expression and assembly properties of heterologous lamins A and C in murine embryonal carcinomas. Eur. J. Cell Biol. 1992, 57, 172–183. [Google Scholar]
- Xue, E.; Minniti, A.; Alexander, T.; Del Papa, N.; Greco, R.; Autoimmune Diseases Working Party (ADWP) of the European Society for Blood; Marrow Transplantation (EBMT). Cellular-Based Therapies in Systemic Sclerosis: From Hematopoietic Stem Cell Transplant to Innovative Approaches. Cells 2022, 11, 3346. [Google Scholar] [CrossRef]
- Hu, G.; Li, G.; Wen, W.; Ding, W.; Zhou, Z.; Zheng, Y.; Huang, T.; Ren, J.; Chen, R.; Zhu, D.; et al. Case report: B7-H3 CAR-T therapy partially controls tumor growth in a basal cell carcinoma patient. Front. Oncol. 2022, 12, 956593. [Google Scholar] [CrossRef]
- de Nattes, T.; Camus, V.; Francois, A.; Dallet, G.; Ferrand, C.; Guerrot, D.; Lemoine, M.; Morin, F.; Thieblemont, C.; Veresezan, E.L.; et al. Kidney Transplant T Cell-Mediated Rejection Occurring After Anti-CD19 CAR T-Cell Therapy for Refractory Aggressive Burkitt-like Lymphoma With 11q Aberration: A Case Report. Am. J. Kidney Dis. 2022, 79, 760–764. [Google Scholar] [CrossRef]
- Feng, G.; Li, Q.; Zhu, H.; Jiang, Y.; Yuan, J.; Fu, Y.; Deng, Q. Safety and Efficacy of Anti-CD19-Chimeric Antigen Receptor T Cell Combined With Programmed Cell Death 1 Inhibitor Therapy in a Patient With Refractory Post-Transplant Lymphoproliferative Disease: Case Report and Literature Review. Front. Oncol. 2021, 11, 726134. [Google Scholar] [CrossRef] [PubMed]
- Hernani, R.; Sancho, A.; Amat, P.; Hernandez-Boluda, J.C.; Perez, A.; Pinana, J.L.; Carretero, C.; Goterris, R.; Gomez, M.; Saus, A.; et al. CAR-T therapy in solid transplant recipients with post-transplant lymphoproliferative disease: Case report and literature review. Curr. Res. Transl. Med. 2021, 69, 103304. [Google Scholar] [CrossRef] [PubMed]
- Krishnamoorthy, S.; Ghobadi, A.; Santos, R.D.; Schilling, J.D.; Malone, A.F.; Murad, H.; Bartlett, N.L.; Alhamad, T. CAR-T therapy in solid organ transplant recipients with treatment refractory posttransplant lymphoproliferative disorder. Am. J. Transplant. 2021, 21, 809–814. [Google Scholar] [CrossRef] [PubMed]
- Luttwak, E.; Hagin, D.; Perry, C.; Wolach, O.; Itchaki, G.; Amit, O.; Bar-On, Y.; Freund, T.; Kay, S.; Eshel, R.; et al. Anti-CD19 CAR-T therapy for EBV-negative posttransplantation lymphoproliferative disease-a single center case series. Bone Marrow Transplant. 2021, 56, 1031–1037. [Google Scholar] [CrossRef]
- Mamlouk, O.; Nair, R.; Iyer, S.P.; Edwards, A.; Neelapu, S.S.; Steiner, R.E.; Adkins, S.A.; Hawkins, M.; Saini, N.; Devashish, K.; et al. Safety of CAR T-cell therapy in kidney transplant recipients. Blood 2021, 137, 2558–2562. [Google Scholar] [CrossRef]
- Melilli, E.; Mussetti, A.; Linares, G.S.; Ruella, M.; La Salette, C.; Savchenko, A.; Taco, M.D.R.; Montero, N.; Grinyo, J.; Fava, A.; et al. Acute Kidney Injury Following Chimeric Antigen Receptor T-Cell Therapy for B-Cell Lymphoma in a Kidney Transplant Recipient. Kidney Med. 2021, 3, 665–668. [Google Scholar] [CrossRef]
- Rosler, W.; Bink, A.; Bissig, M.; Imbach, L.; Marques Maggio, E.; Manz, M.G.; Muller, T.; Roth, P.; Rushing, E.; Widmer, C.; et al. CAR T-cell Infusion Following Checkpoint Inhibition Can Induce Remission in Chemorefractory Post-transplant Lymphoproliferative Disorder of the CNS. Hemasphere 2022, 6, e733. [Google Scholar] [CrossRef]
- Portuguese, A.J.; Gauthier, J.; Tykodi, S.S.; Hall, E.T.; Hirayama, A.V.; Yeung, C.C.S.; Blosser, C.D. CD19 CAR-T therapy in solid organ transplant recipients: Case report and systematic review. Bone Marrow Transplant. 2022, 1–7. [Google Scholar] [CrossRef]


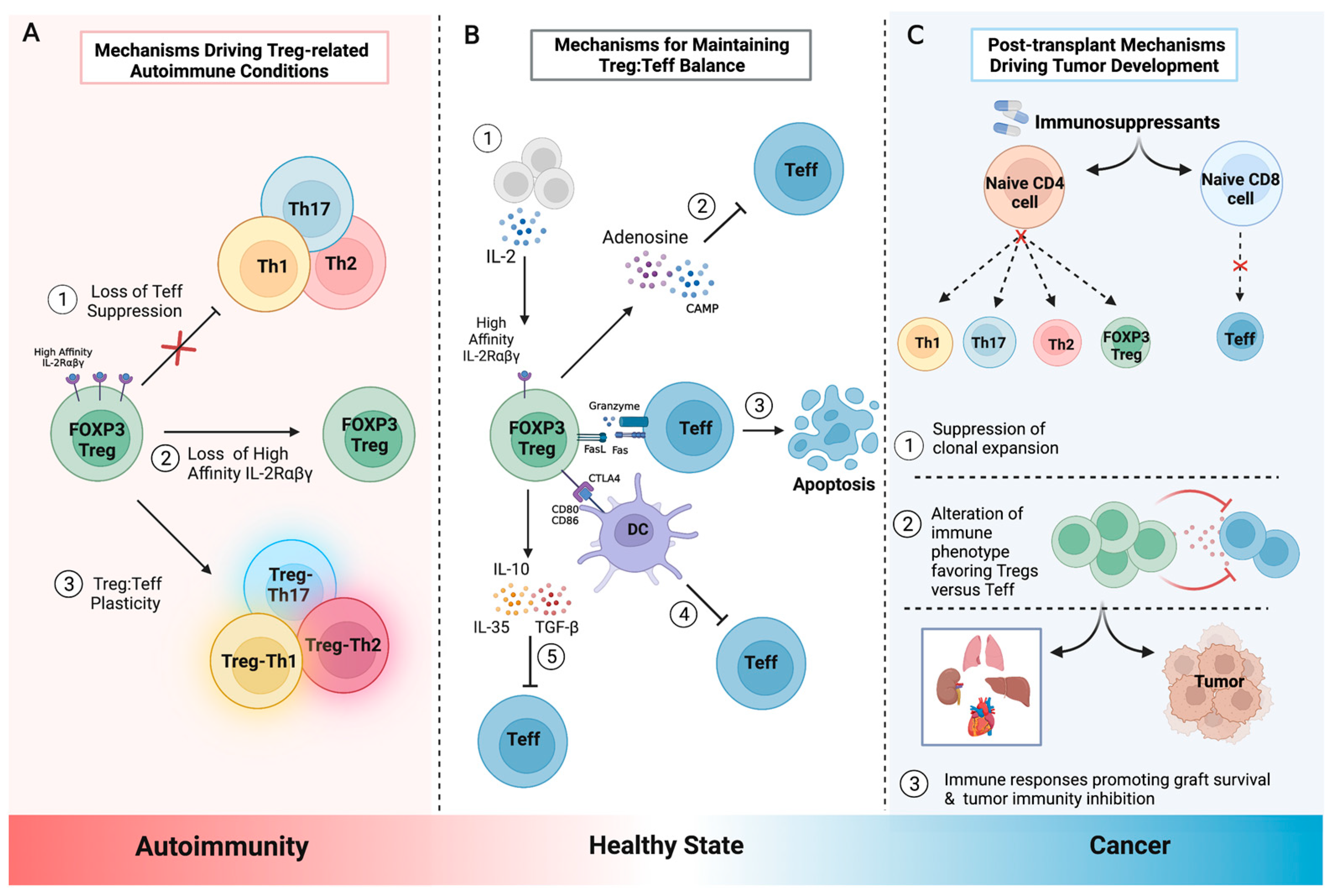{kind=link}
| Disease | Risk Factors | Teff Cells | Cytokines | Treatments | References |
|---|---|---|---|---|---|
| Psoriasis | Obesity, infection, trauma | Th1, Treg-Th17 | IL-2, IL-17, IL-22, IL-23, IL-26, TNF-α, IFN-γ | Anti-TNF-α inhibitors, T-cell-targeted therapies | [71,72,73,74,75,76,77,78] |
| Vitiligo | Genetics, trauma | Treg-Th1 | IL-1β, IL-6, IL-15, IL-22, IL-33, TNF-α, IFN-γ | JAK inhibitor, NB-UVB therapy, anti-TNF-α inhibitors | [79,80,81,82,83,84,85,86,87] |
| Systemic sclerosis | Genetics, silica, solvents, heavy metal | Treg-Th2, Treg-Th17 | IL-4, IL-13, IL-17, IL-21, IL-22, IL-33 | Low-dose IL-2 therapy, PDE-5 inhibitors, calcium channel blocker, bosentan | [88,89,90,91,92,93,94,95,96,97] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Oparaugo, N.C.; Ouyang, K.; Nguyen, N.P.N.; Nelson, A.M.; Agak, G.W. Human Regulatory T Cells: Understanding the Role of Tregs in Select Autoimmune Skin Diseases and Post-Transplant Nonmelanoma Skin Cancers. Int. J. Mol. Sci. 2023, 24, 1527. https://doi.org/10.3390/ijms24021527
Oparaugo NC, Ouyang K, Nguyen NPN, Nelson AM, Agak GW. Human Regulatory T Cells: Understanding the Role of Tregs in Select Autoimmune Skin Diseases and Post-Transplant Nonmelanoma Skin Cancers. International Journal of Molecular Sciences. 2023; 24(2):1527. https://doi.org/10.3390/ijms24021527
Chicago/Turabian StyleOparaugo, Nicole Chizara, Kelsey Ouyang, Nam Phuong N. Nguyen, Amanda M. Nelson, and George W. Agak. 2023. "Human Regulatory T Cells: Understanding the Role of Tregs in Select Autoimmune Skin Diseases and Post-Transplant Nonmelanoma Skin Cancers" International Journal of Molecular Sciences 24, no. 2: 1527. https://doi.org/10.3390/ijms24021527