
Optimized Antimicrobial Peptide Jelleine-I Derivative Br-J-I Inhibits Fusobacterium Nucleatum to Suppress Colorectal Cancer Progression
 , , ,
, , , 
Abstract
:1. Introduction
2. Results
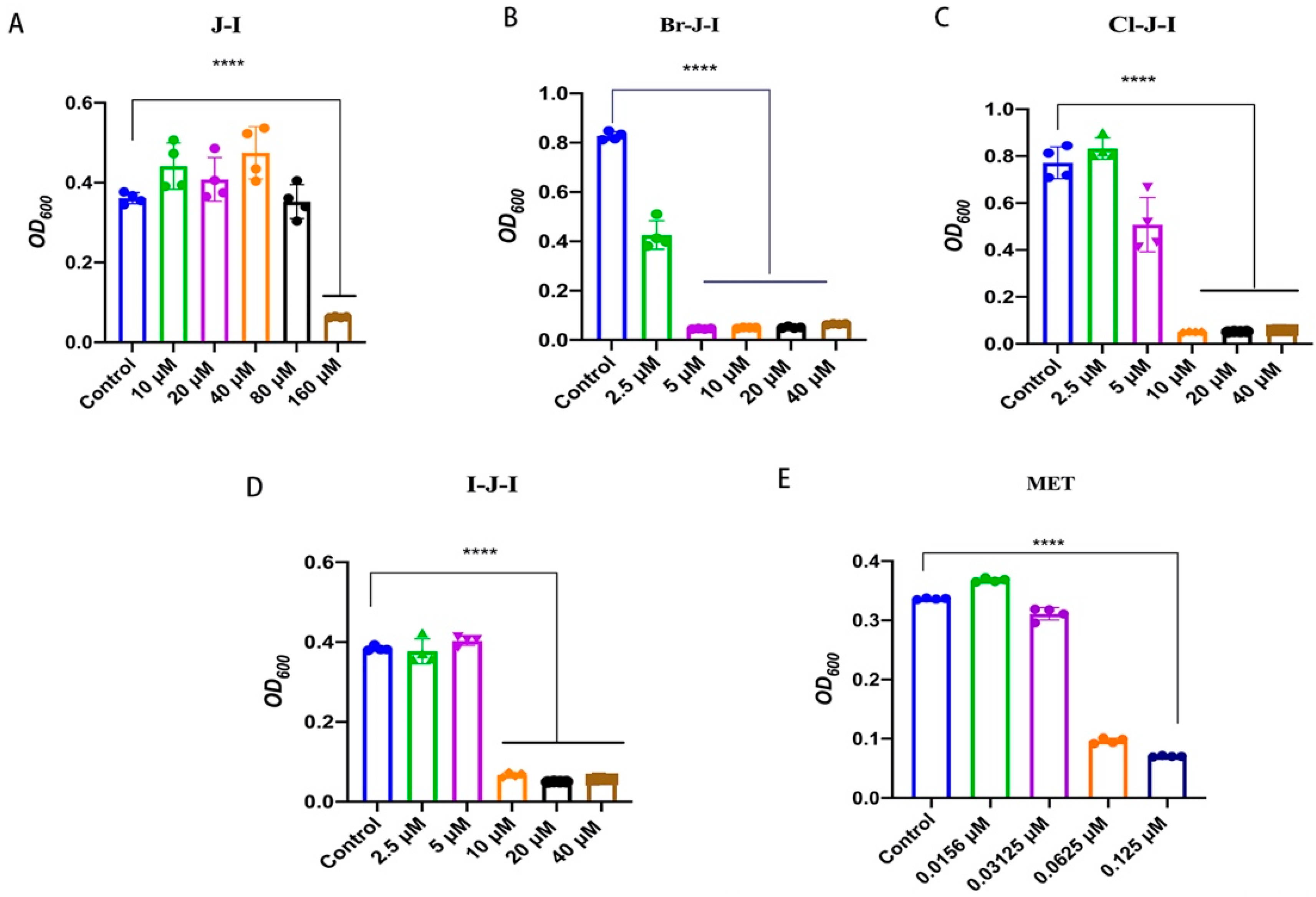
2.1. Br-J-I Shows Potent Antimicrobial Activity against Fn
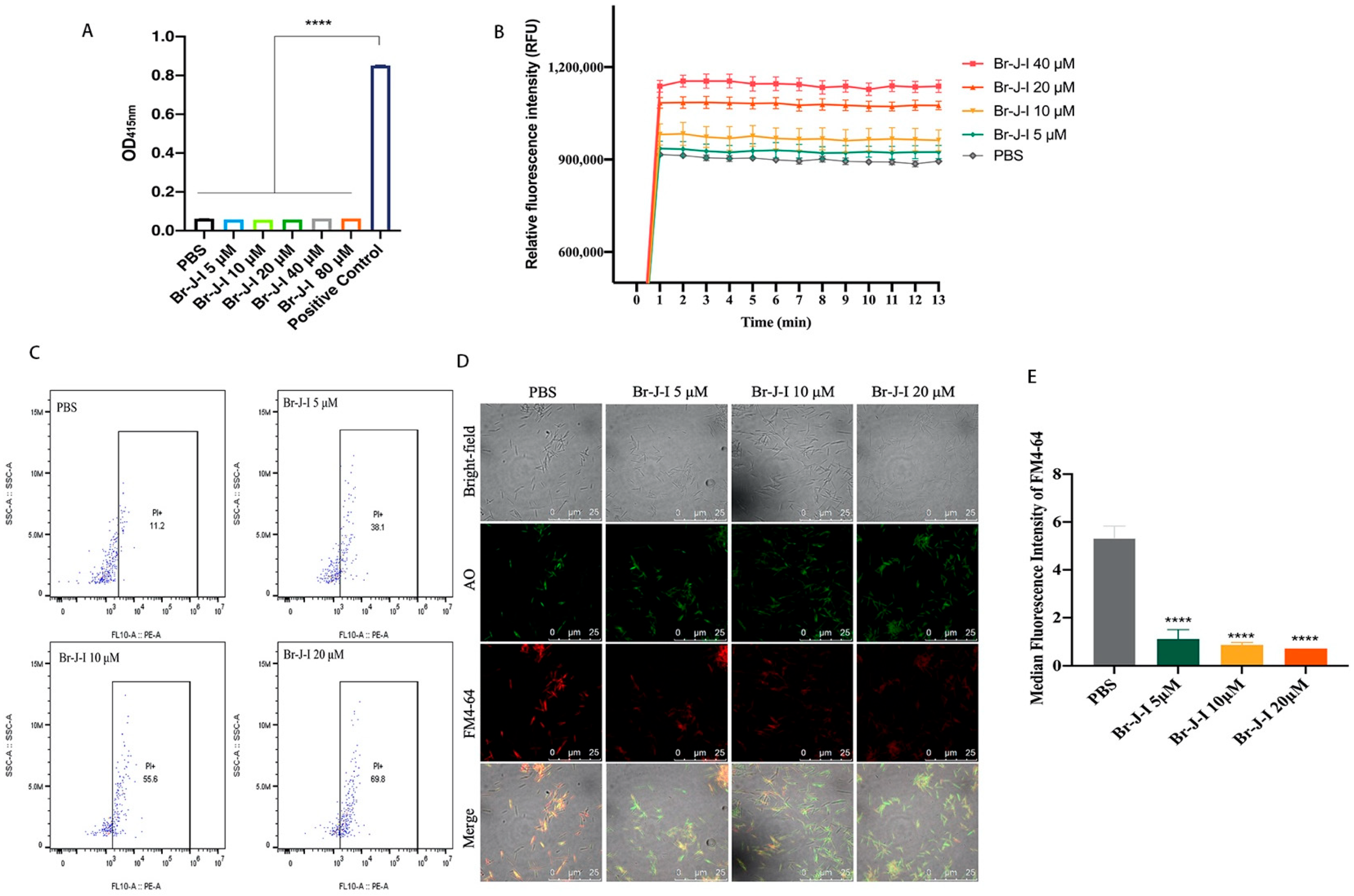
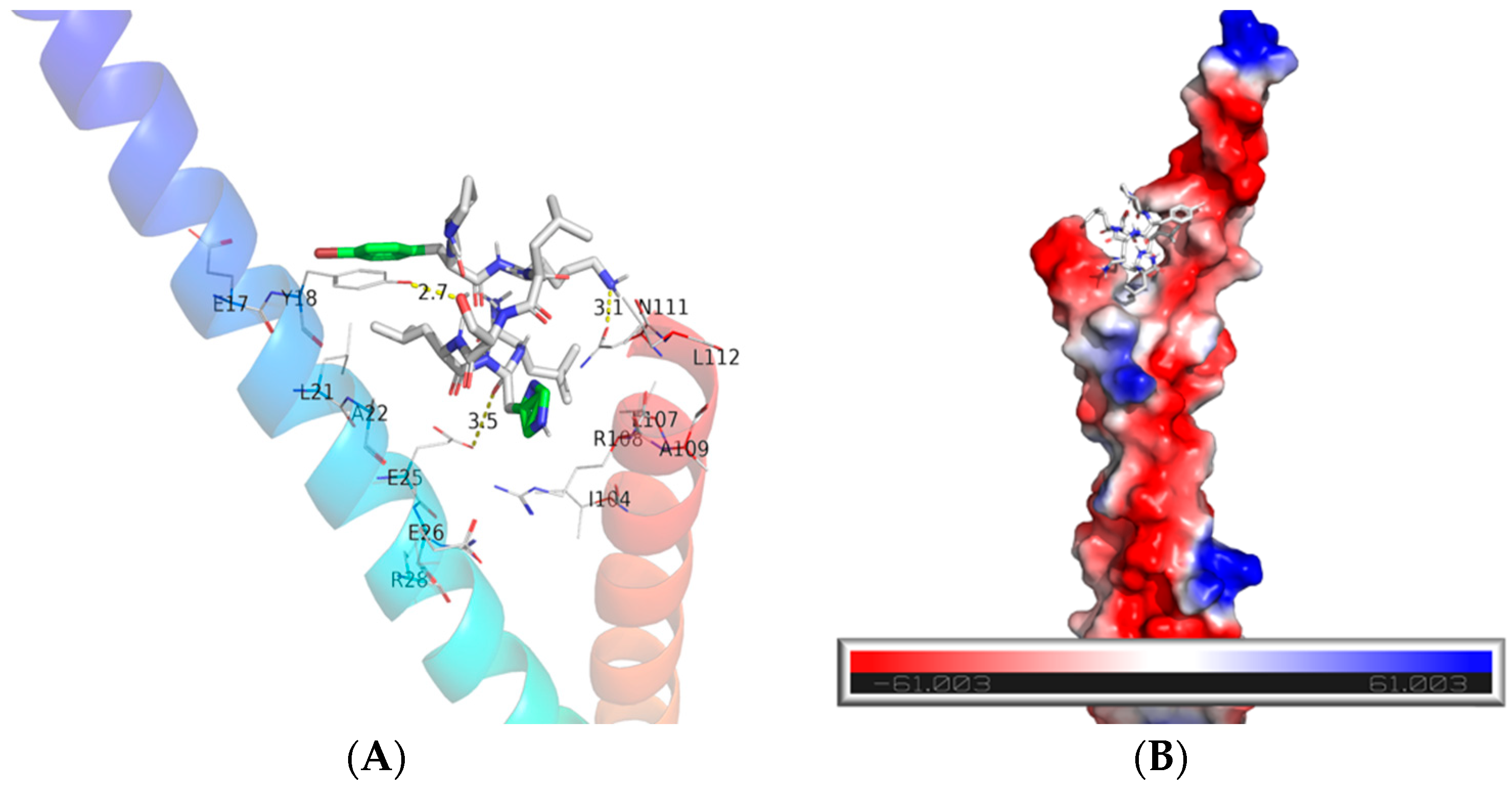
2.2. Br-J-I Induces Membrane Disruption of Fn to Inhibit Fn
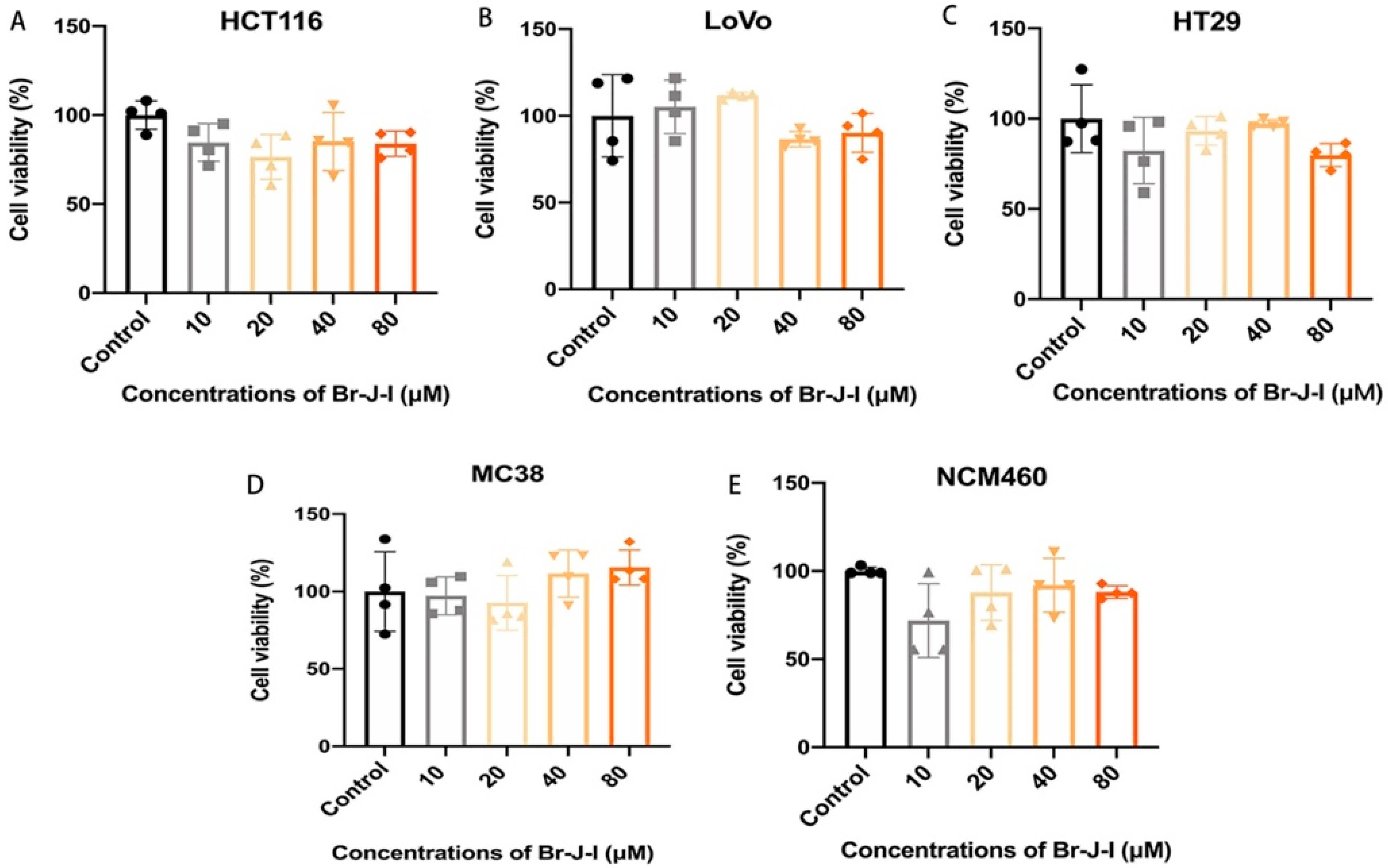
2.3. Br-J-I Exhibits Little Cytotoxicity to Colon Epithelial Cells and CRC Cells
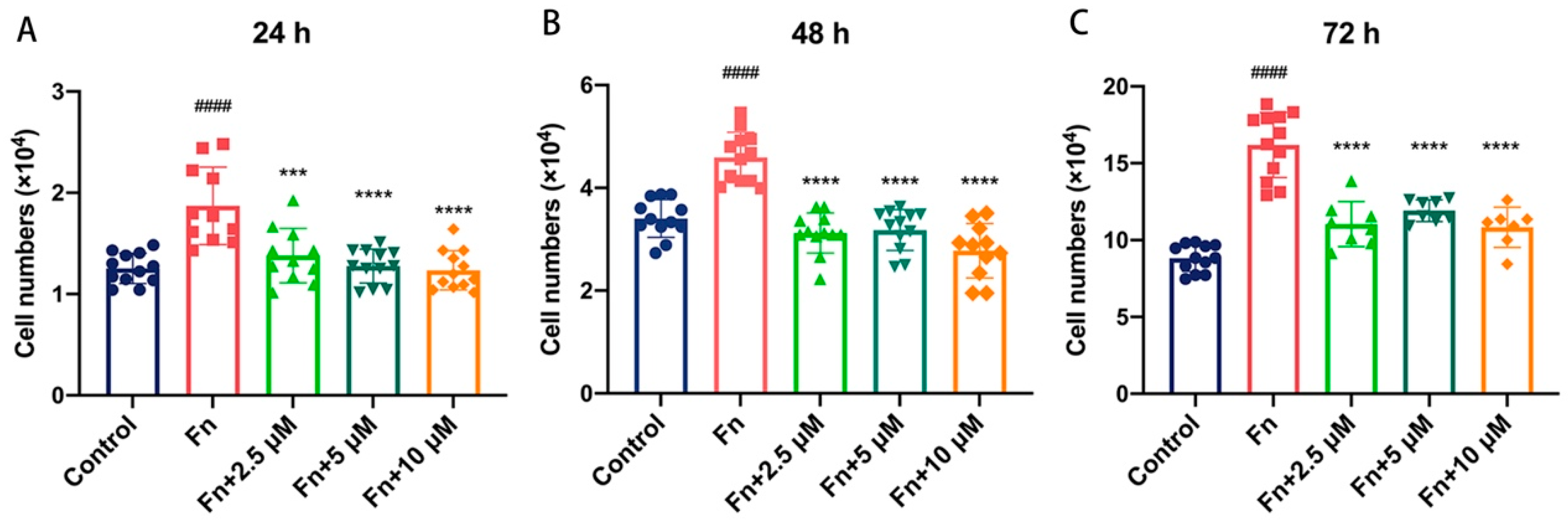
2.4. Br-J-I Suppresses the Tumor-Promoting Effect of Fn
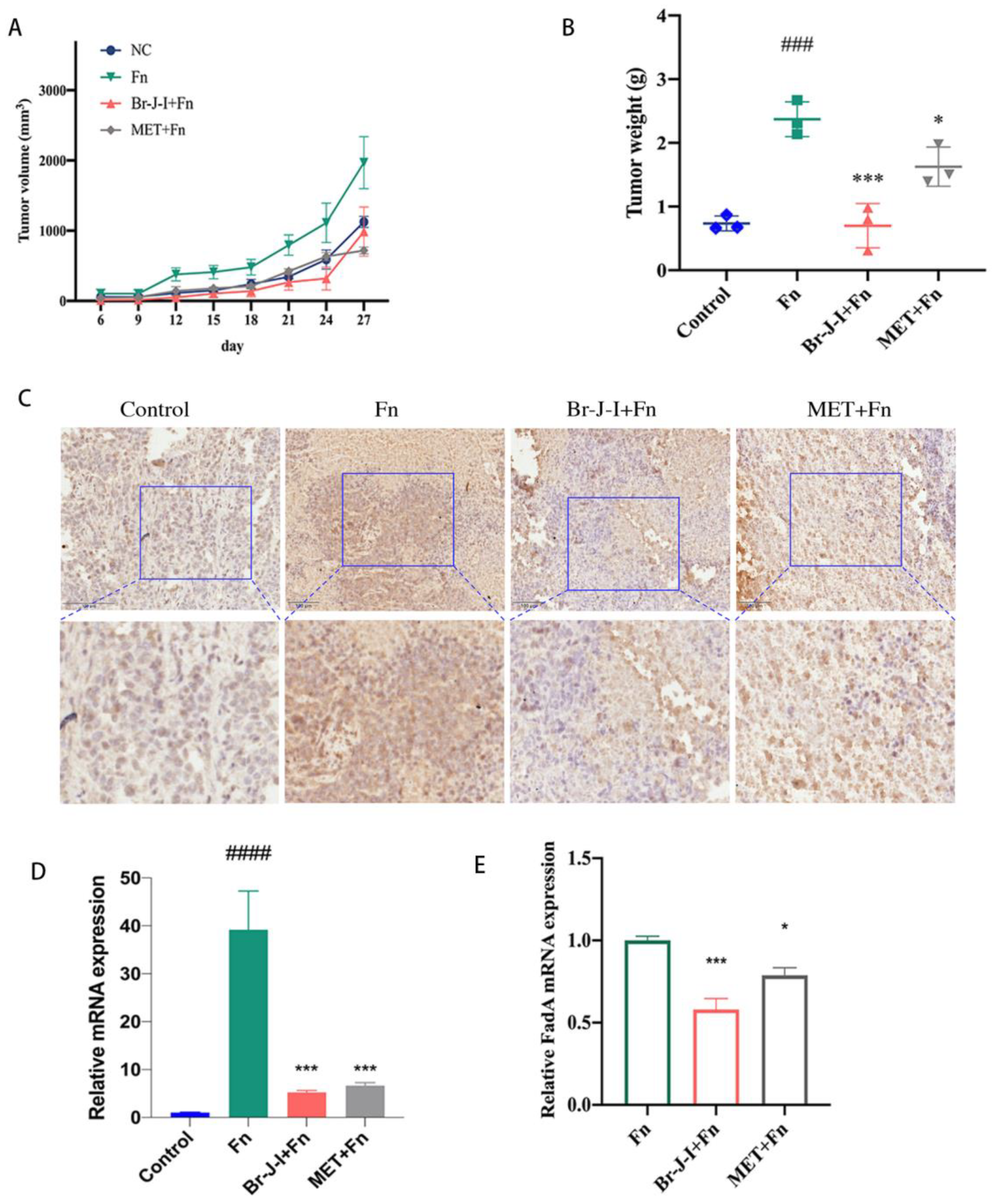
2.5. Br-J-I Inhibits the Growth of CRC Induced by Intratumoral Fn in Mice Engrafted with HCT116
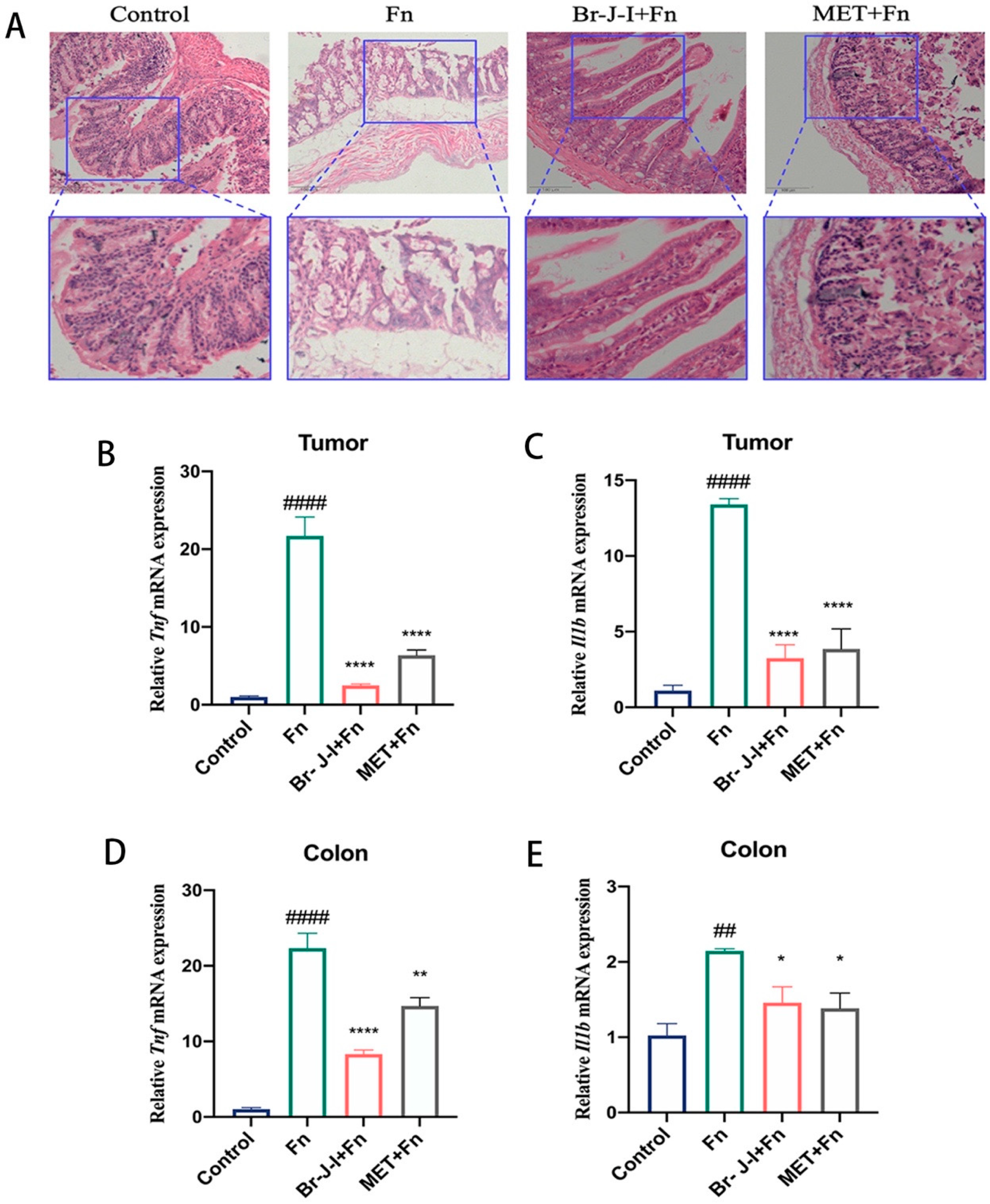
2.6. Br-J-I Inhibits Fn-Induced Inflammation
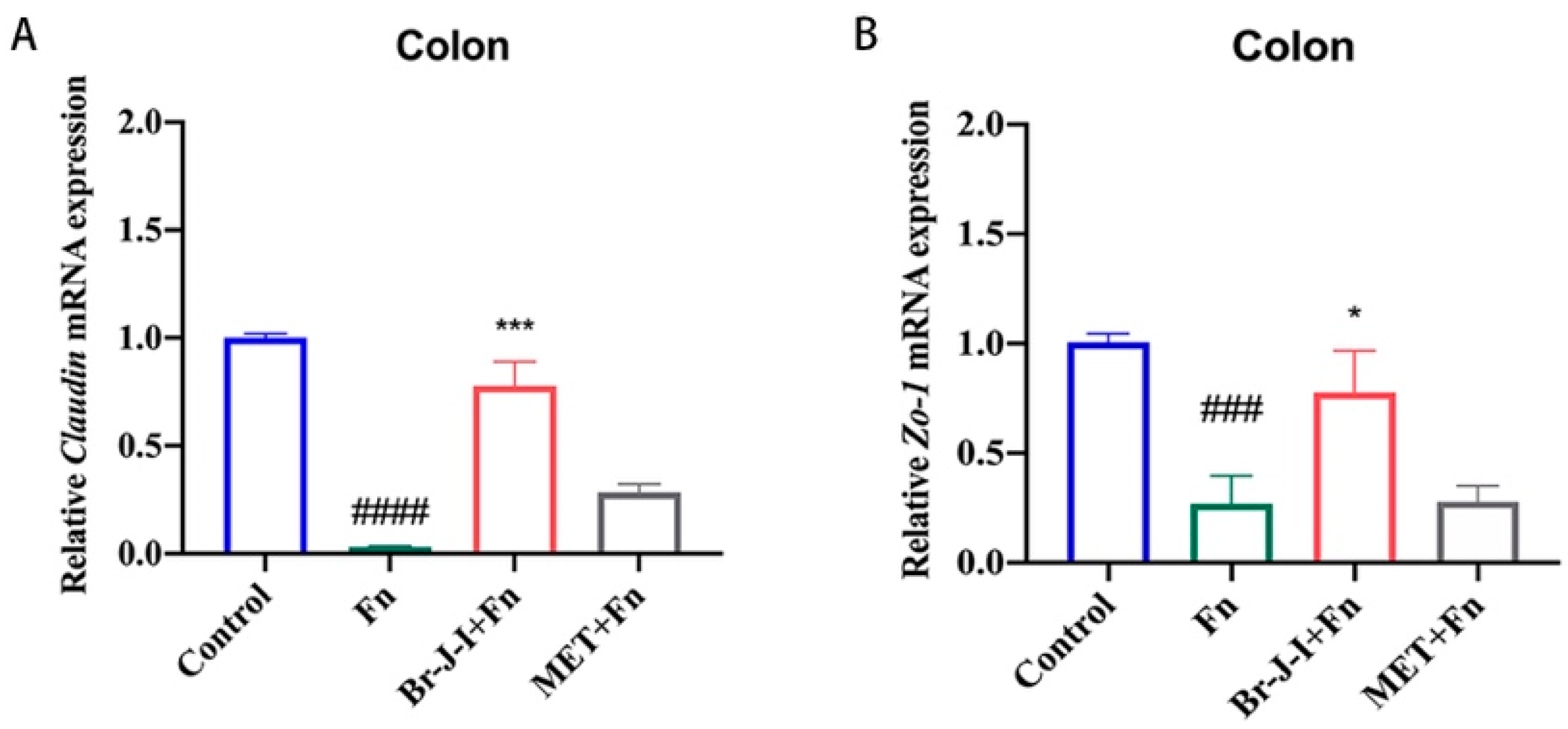
2.7. Br-J-I Improves Intestinal Mucosa Tight Junction (TJ) in HCT116 Cell-Engrafted Mice with Intratumoral Fn Colonization
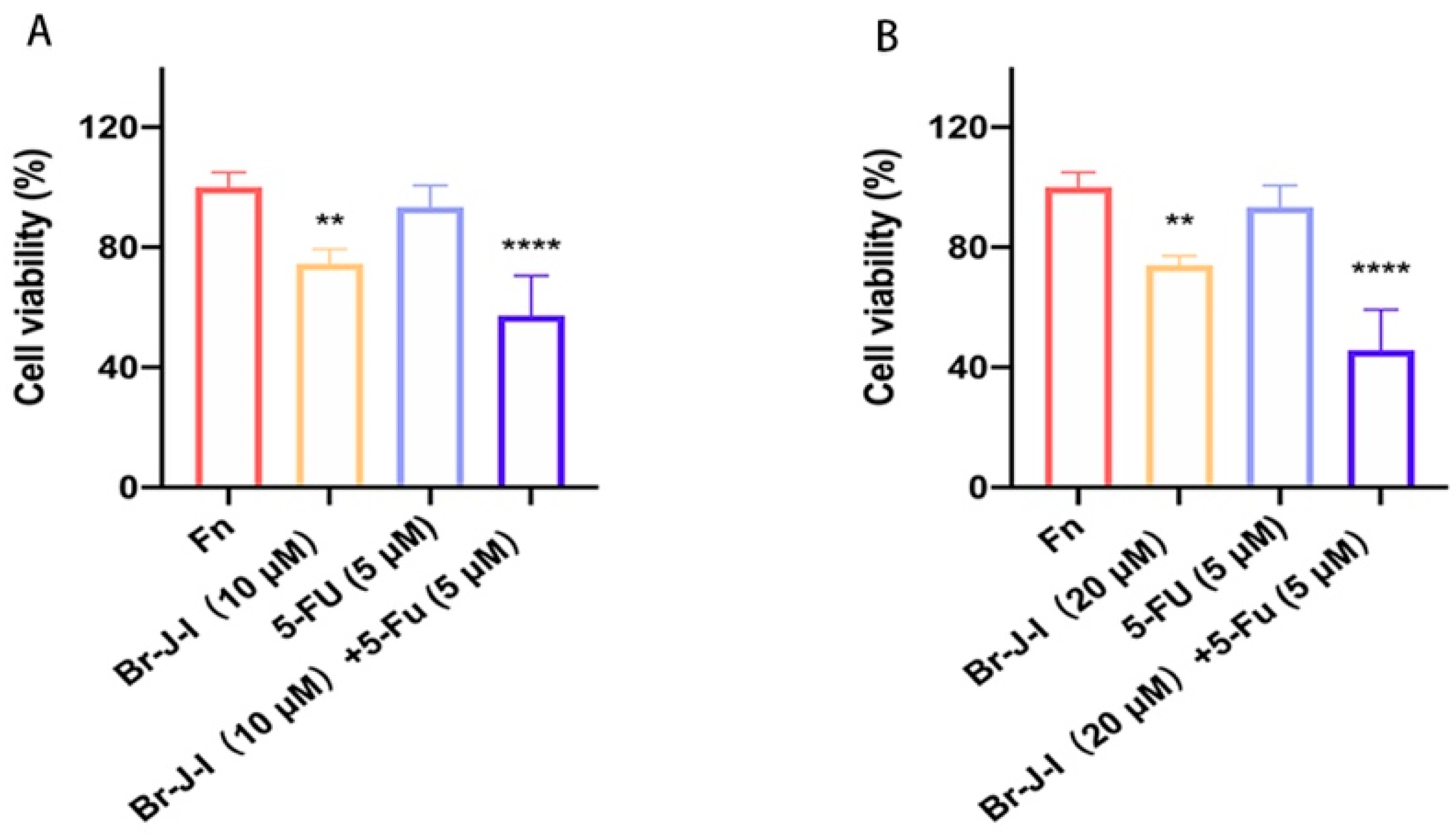
2.8. Br-J-I Synergizes with 5-FU to Exert the Antitumor Effect
3. Discussion
4. Materials and Methods
4.1. Synthesis and Purification of AMPs
4.2. Minimum Inhibitory Concentration (MIC) Assay
4.3. Minimum Bactericidal Concentration (MBC) Assay
4.4. Outer Membrane (OM) Permeability
4.5. Flow Cytometric Analysis
4.6. Laser Scanning Confocal Microscopy (LSCM)
4.7. Binding Mode Prediction
4.8. Cytotoxicity
4.9. Effect of Br-J-I on Cell Proliferation
4.10. Effect of Br-J-I on the Growth of Murine CRC with Fn Colonization
4.11. Quantification of Fn Load in the Tumors and the Expression of the Proinflammatory Cytokines in Tumors and Colon Tissues
4.12. Analysis of Intestinal Permeability of Murine CRC
4.13. Effect of Br-J-I Combined with 5-Fluorouracil
4.14. Statistical Analyses
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
References
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global Cancer Statistics 2020: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef] [PubMed]
- Rawla, P.; Sunkara, T.; Barsouk, A. Epidemiology of colorectal cancer: Incidence, mortality, survival, and risk factors. Prz. Gastroenterol. 2019, 14, 89–103. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.; He, F.; Karin, M. From Liver Fat to Cancer: Perils of the Western Diet. Cancers 2021, 13, 1095. [Google Scholar] [CrossRef] [PubMed]
- Akimoto, N.; Ugai, T.; Zhong, R.; Hamada, T.; Fujiyoshi, K.; Giannakis, M.; Wu, K.; Cao, Y.; Ng, K.; Ogino, S. Rising incidence of early-onset colorectal cancer—A call to action. Nat. Rev. Clin. Oncol. 2021, 18, 230–243. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Probin, V.; Zhou, D. Cancer therapy-induced residual bone marrow injury-Mechanisms of induction and implication for therapy. Curr. Cancer Ther. Rev. 2006, 2, 271–279. [Google Scholar] [CrossRef] [Green Version]
- Chu, D.; Zhang, Z.; Li, Y.; Wu, L.; Zhang, J.; Wang, W.; Zhang, J. Prediction of colorectal cancer relapse and prognosis by tissue mRNA levels of NDRG2. Mol. Cancer Ther. 2011, 10, 47–56. [Google Scholar] [CrossRef] [Green Version]
- Gomes, A.C.; Hoffmann, C.; Mota, J.F. The human gut microbiota: Metabolism and perspective in obesity. Gut Microbes 2018, 9, 308–325. [Google Scholar] [CrossRef] [Green Version]
- Cheng, Y.; Ling, Z.; Li, L. The Intestinal Microbiota and Colorectal Cancer. Front. Immunol. 2020, 11, 615056. [Google Scholar] [CrossRef]
- Huh, J.W.; Roh, T.Y. Opportunistic detection of Fusobacterium nucleatum as a marker for the early gut microbial dysbiosis. BMC Microbiol. 2020, 20, 208. [Google Scholar] [CrossRef]
- Kostic, A.D.; Gevers, D.; Pedamallu, C.S.; Michaud, M.; Duke, F.; Earl, A.M.; Ojesina, A.I.; Jung, J.; Bass, A.J.; Tabernero, J.; et al. Genomic analysis identifies association of Fusobacterium with colorectal carcinoma. Genome Res. 2012, 22, 292–298. [Google Scholar] [CrossRef]
- Mima, K.; Nishihara, R.; Qian, Z.R.; Cao, Y.; Sukawa, Y.; Nowak, J.A.; Yang, J.; Dou, R.; Masugi, Y.; Song, M.; et al. Fusobacterium nucleatum in colorectal carcinoma tissue and patient prognosis. Gut 2016, 65, 1973–1980. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haruki, K.; Kosumi, K.; Hamada, T.; Twombly, T.S.; Vayrynen, J.P.; Kim, S.A.; Masugi, Y.; Qian, Z.R.; Mima, K.; Baba, Y.; et al. Association of autophagy status with amount of Fusobacterium nucleatum in colorectal cancer. J. Pathol. 2020, 250, 397–408. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Chen, Y.; Zhang, J.; Cao, P.; Su, W.; Deng, Y.; Zhan, N.; Fu, X.; Huang, Y.; Dong, W. Fusobacterium nucleatum Promotes Metastasis in Colorectal Cancer by Activating Autophagy Signaling via the Upregulation of CARD3 Expression. Theranostics 2020, 10, 323–339. [Google Scholar] [CrossRef] [PubMed]
- Marmol, I.; Sanchez-de-Diego, C.; Pradilla Dieste, A.; Cerrada, E.; Rodriguez Yoldi, M.J. Colorectal Carcinoma: A General Overview and Future Perspectives in Colorectal Cancer. Int. J. Mol. Sci. 2017, 18, 197. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cochrane, K.; Robinson, A.V.; Holt, R.A.; Allen-Vercoe, E. A survey of Fusobacterium nucleatum genes modulated by host cell infection. Microb Genom. 2020, 6, e000300. [Google Scholar] [CrossRef] [PubMed]
- Hashemi Goradel, N.; Heidarzadeh, S.; Jahangiri, S.; Farhood, B.; Mortezaee, K.; Khanlarkhani, N.; Negahdari, B. Fusobacterium nucleatum and colorectal cancer: A mechanistic overview. J. Cell Physiol. 2019, 234, 2337–2344. [Google Scholar] [CrossRef]
- Kostic, A.D.; Chun, E.; Robertson, L.; Glickman, J.N.; Gallini, C.A.; Michaud, M.; Clancy, T.E.; Chung, D.C.; Lochhead, P.; Hold, G.L.; et al. Fusobacterium nucleatum potentiates intestinal tumorigenesis and modulates the tumor-immune microenvironment. Cell Host Microbe 2013, 14, 207–215. [Google Scholar] [CrossRef] [Green Version]
- Liu, F.; Li, P.; Chen, M.; Luo, Y.; Prabhakar, M.; Zheng, H.; He, Y.; Qi, Q.; Long, H.; Zhang, Y.; et al. Fructooligosaccharide (FOS) and Galactooligosaccharide (GOS) Increase Bifidobacterium but Reduce Butyrate Producing Bacteria with Adverse Glycemic Metabolism in healthy young population. Sci. Rep. 2017, 7, 11789. [Google Scholar] [CrossRef] [Green Version]
- Shridhar, P.B.; Amachawadi, R.G.; Tokach, M.; Patel, I.; Gangiredla, J.; Mammel, M.; Nagaraja, T.G. Whole genome sequence analyses-based assessment of virulence potential and antimicrobial susceptibilities and resistance of Enterococcus faecium strains isolated from commercial swine and cattle probiotic products. J. Anim. Sci. 2022, 100, skac030. [Google Scholar] [CrossRef]
- Hill, C. Balancing the risks and rewards of live biotherapeutics. Nat. Rev. Gastroenterol. Hepatol. 2020, 17, 133–134. [Google Scholar] [CrossRef]
- Rautio, M.; Jousimies-Somer, H.; Kauma, H.; Pietarinen, I.; Saxelin, M.; Tynkkynen, S.; Koskela, M. Liver abscess due to a Lactobacillus rhamnosus strain indistinguishable from L. rhamnosus strain GG. Clin. Infect. Dis. 1999, 28, 1159–1160. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Settle, C.D.; Wilcox, M.H. Review article: Antibiotic-induced Clostridium difficile infection. Aliment Pharmacol. Ther. 1996, 10, 835–841. [Google Scholar] [CrossRef] [PubMed]
- Gleckman, R.A.; Borrego, F. Adverse reactions to antibiotics. Clues for recognizing, understanding, and avoiding them. Postgrad Med. 1997, 101, 97–98, 101–104, 107–108. [Google Scholar] [CrossRef]
- Reddy, K.V.; Yedery, R.D.; Aranha, C. Antimicrobial peptides: Premises and promises. Int. J. Antimicrob. Agents 2004, 24, 536–547. [Google Scholar] [CrossRef] [PubMed]
- Tanhaieian, A.; Sekhavati, M.H.; Ahmadi, F.S.; Mamarabadi, M. Heterologous expression of a broad-spectrum chimeric antimicrobial peptide in Lactococcus lactis: Its safety and molecular modeling evaluation. Microb. Pathog. 2018, 125, 51–59. [Google Scholar] [CrossRef]
- Tanhaeian, A.; Mirzaii, M.; Pirkhezranian, Z.; Sekhavati, M.H. Generation of an engineered food-grade Lactococcus lactis strain for production of an antimicrobial peptide: In vitro and in silico evaluation. BMC Biotechnol. 2020, 20, 19. [Google Scholar] [CrossRef] [Green Version]
- Pirkhezranian, Z.; Tanhaeian, A.; Mirzaii, M.; Sekhavati, M.H. Expression of Enterocin-P in HEK Platform: Evaluation of Its Cytotoxic Effects on Cancer Cell Lines and Its Potency to Interact with Cell-Surface Glycosaminoglycan by Molecular Modeling. Int. J. Pept. Res. Ther. 2020, 26, 1503–1512. [Google Scholar] [CrossRef]
- Tahmoorespur, M.; Azghandi, M.; Javadmanesh, A.; Meshkat, Z.; Sekhavati, M.H. A Novel Chimeric Anti-HCV Peptide Derived from Camel Lactoferrin and Molecular Level Insight on Its Interaction with E2. Int. J. Pept. Res. Ther. 2020, 26, 1593–1605. [Google Scholar] [CrossRef]
- Fontana, R.; Mendes, M.A.; de Souza, B.M.; Konno, K.; Cesar, L.M.; Malaspina, O.; Palma, M.S. Jelleines: A family of antimicrobial peptides from the Royal Jelly of honeybees (Apis mellifera). Peptides 2004, 25, 919–928. [Google Scholar] [CrossRef]
- Jia, F.; Zhang, Y.; Wang, J.; Peng, J.; Zhao, P.; Zhang, L.; Yao, H.; Ni, J.; Wang, K. The effect of halogenation on the antimicrobial activity, antibiofilm activity, cytotoxicity and proteolytic stability of the antimicrobial peptide Jelleine-I. Peptides 2019, 112, 56–66. [Google Scholar] [CrossRef]
- Bahar, A.A.; Ren, D. Antimicrobial peptides. Pharmaceuticals 2013, 6, 1543–1575. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, Y.; Huang, J.; Chen, Y. Alpha-helical cationic antimicrobial peptides: Relationships of structure and function. Protein Cell 2010, 1, 143–152. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Srinivasan, S.; Waghu, F.H.; Idicula-Thomas, S.; Venkatesh, K.V. A steady-state modeling approach for simulation of antimicrobial peptide-cell membrane interaction. Biochim. Biophys. Acta Biomembr. 2020, 1862, 183242. [Google Scholar] [CrossRef]
- Haake, S.K.; Wang, X. Cloning and expression of FomA, the major outer-membrane protein gene from Fusobacterium nucleatum T18. Arch. Oral Biol. 1997, 42, 19–24. [Google Scholar] [CrossRef] [PubMed]
- Cowan, S.W.; Schirmer, T.; Rummel, G.; Steiert, M.; Ghosh, R.; Pauptit, R.A.; Jansonius, J.N.; Rosenbusch, J.P. Crystal structures explain functional properties of two E. coli porins. Nature 1992, 358, 727–733. [Google Scholar] [CrossRef] [PubMed]
- Jumper, J.; Evans, R.; Pritzel, A.; Green, T.; Figurnov, M.; Ronneberger, O.; Tunyasuvunakool, K.; Bates, R.; Zidek, A.; Potapenko, A.; et al. Highly accurate protein structure prediction with AlphaFold. Nature 2021, 596, 583–589. [Google Scholar] [CrossRef] [PubMed]
- Arthur, J.C.; Gharaibeh, R.Z.; Mühlbauer, M.; Perez-Chanona, E.; Uronis, J.M.; McCafferty, J.; Fodor, A.A.; Jobin, C. Microbial genomic analysis reveals the essential role of inflammation in bacteria-induced colorectal cancer. Nat. Commun. 2014, 5, 4724. [Google Scholar] [CrossRef] [Green Version]
- Ray, A.L.; Berggren, K.L.; Restrepo Cruz, S.; Gan, G.N.; Beswick, E.J. Inhibition of MK2 suppresses IL-1β, IL-6, and TNF-α-dependent colorectal cancer growth. Int. J. Cancer 2018, 142, 1702–1711. [Google Scholar] [CrossRef] [Green Version]
- Wang, Z.; Li, R.; Tan, J.; Peng, L.; Wang, P.; Liu, J.; Xiong, H.; Jiang, B.; Chen, Y. Syndecan-1 Acts in Synergy with Tight Junction Through Stat3 Signaling to Maintain Intestinal Mucosal Barrier and Prevent Bacterial Translocation. Inflamm. Bowel Dis. 2015, 21, 1894–1907. [Google Scholar] [CrossRef]
- Cho, Y.H.; Ro, E.J.; Yoon, J.S.; Mizutani, T.; Kang, D.W.; Park, J.C.; Il Kim, T.; Clevers, H.; Choi, K.Y. 5-FU promotes stemness of colorectal cancer via p53-mediated WNT/beta-catenin pathway activation. Nat. Commun. 2020, 11, 5321. [Google Scholar] [CrossRef]
- Yu, T.; Guo, F.; Yu, Y.; Sun, T.; Ma, D.; Han, J.; Qian, Y.; Kryczek, I.; Sun, D.; Nagarsheth, N.; et al. Fusobacterium nucleatum Promotes Chemoresistance to Colorectal Cancer by Modulating Autophagy. Cell 2017, 170, 548–563.e16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, S.; Cai, S.; Ma, Y. Association between Fusobacterium nucleatum and colorectal cancer: Progress and future directions. J. Cancer 2018, 9, 1652–1659. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tew, G.N.; Clements, D.; Tang, H.; Arnt, L.; Scott, R.W. Antimicrobial activity of an abiotic host defense peptide mimic. Biochim. Biophys. Acta 2006, 1758, 1387–1392. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tanhaeian, A.; Jaafari, M.R.; Ahmadi, F.S.; Vakili-Ghartavol, R.; Sekhavati, M.H. Secretory Expression of a Chimeric Peptide in Lactococcus lactis: Assessment of its Cytotoxic Activity and a Deep View on Its Interaction with Cell-Surface Glycosaminoglycans by Molecular Modeling. Probiotics Antimicrob. Proteins 2019, 11, 1034–1041. [Google Scholar] [CrossRef] [PubMed]
- He, F.; Antonucci, L.; Karin, M. NRF2 as a regulator of cell metabolism and inflammation in cancer. Carcinogenesis 2020, 41, 405–416. [Google Scholar] [CrossRef]
- He, F.; Ru, X.; Wen, T. NRF2, a Transcription Factor for Stress Response and Beyond. Int. J. Mol. Sci. 2020, 21, 4777. [Google Scholar] [CrossRef]
- Sethy, C.; Kundu, C.N. 5-Fluorouracil (5-FU) resistance and the new strategy to enhance the sensitivity against cancer: Implication of DNA repair inhibition. Biomed. Pharmacother. 2021, 137, 111285. [Google Scholar] [CrossRef]
- Fields, G.B.; Noble, R.L. Solid phase peptide synthesis utilizing 9-fluorenylmethoxycarbonyl amino acids. Int. J. Pept. Protein Res. 1990, 35, 161–214. [Google Scholar] [CrossRef]
- Jia, F.; Liang, X.; Wang, J.; Zhang, L.; Zhou, J.; He, Y.; Zhang, F.; Yan, W.; Wang, K. Tryptic Stability and Antimicrobial Activity of the Derivatives of Polybia-CP with Fine-Tuning Modification in the Side Chain of Lysine. Int. J. Pept. Res. Ther. 2021, 27, 851–862. [Google Scholar] [CrossRef]
- Chacko, S.; Samanta, S. Novel Thiosemicarbazide Hybrids with Amino Acids and Peptides Against Hepatocellular Carcinoma: A Molecular Designing Approach Towards Multikinase Inhibitor. Curr. Comput. Aided Drug Des. 2015, 11, 279–290. [Google Scholar] [CrossRef]
- Ferreira, L.G.; Dos Santos, R.N.; Oliva, G.; Andricopulo, A.D. Molecular docking and structure-based drug design strategies. Molecules 2015, 20, 13384–13421. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Varadi, M.; Anyango, S.; Deshpande, M.; Nair, S.; Natassia, C.; Yordanova, G.; Yuan, D.; Stroe, O.; Wood, G.; Laydon, A.; et al. AlphaFold Protein Structure Database: Massively expanding the structural coverage of protein-sequence space with high-accuracy models. Nucleic Acids Res. 2022, 50, D439–D444. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Shi, Y.; Chen, Y.; Yu, S.; Hao, J.; Luo, J.; Sha, X.; Fang, X. Enhanced antitumor efficacy by paclitaxel-loaded pluronic P123/F127 mixed micelles against non-small cell lung cancer based on passive tumor targeting and modulation of drug resistance. Eur. J. Pharm. Biopharm. 2010, 75, 341–353. [Google Scholar] [CrossRef] [PubMed]
- Cheng, Y.; Wang, G.; Zhao, L.; Dai, S.; Han, J.; Hu, X.; Zhou, C.; Wang, F.; Ma, H.; Li, B.; et al. Periplocymarin Induced Colorectal Cancer Cells Apoptosis Via Impairing PI3K/AKT Pathway. Front. Oncol. 2021, 11, 753598. [Google Scholar] [CrossRef]
- Guo, P.; Tian, Z.; Kong, X.; Yang, L.; Shan, X.; Dong, B.; Ding, X.; Jing, X.; Jiang, C.; Jiang, N.; et al. FadA promotes DNA damage and progression of Fusobacterium nucleatum-induced colorectal cancer through up-regulation of chk2. J. Exp. Clin. Cancer Res. 2020, 39, 202. [Google Scholar] [CrossRef]
- Rubinstein, M.R.; Wang, X.; Liu, W.; Hao, Y.; Cai, G.; Han, Y.W. Fusobacterium nucleatum promotes colorectal carcinogenesis by modulating E-cadherin/β-catenin signaling via its FadA adhesin. Cell Host Microbe 2013, 14, 195–206. [Google Scholar] [CrossRef] [Green Version]
- Abed, J.; Emgård, J.E.M.; Zamir, G.; Faroja, M.; Almogy, G.; Grenov, A.; Sol, A.; Naor, R.; Pikarsky, E.; Atlan, K.A.; et al. Fap2 Mediates Fusobacterium nucleatum Colorectal Adenocarcinoma Enrichment by Binding to Tumor-Expressed Gal-GalNAc. Cell Host Microbe 2016, 20, 215–225. [Google Scholar] [CrossRef] [Green Version]
- He, F.; Antonucci, L.; Yamachika, S.; Zhang, Z.; Taniguchi, K.; Umemura, A.; Hatzivassiliou, G.; Roose-Girma, M.; Reina-Campos, M.; Duran, A.; et al. NRF2 activates growth factor genes and downstream AKT signaling to induce mouse and human hepatomegaly. J. Hepatol. 2020, 72, 1182–1195. [Google Scholar] [CrossRef] [Green Version]
- Yueh, M.F.; He, F.; Chen, C.; Vu, C.; Tripathi, A.; Knight, R.; Karin, M.; Chen, S.; Tukey, R.H. Triclosan leads to dysregulation of the metabolic regulator FGF21 exacerbating high fat diet-induced nonalcoholic fatty liver disease. Proc. Natl. Acad. Sci. USA 2020, 117, 31259–31266. [Google Scholar] [CrossRef]
- Jin, Z.J. Addition in drug combination (author’s transl). Zhongguo Yao Li Xue Bao 1980, 1, 70–76. [Google Scholar]
- Wang, Y.; Yu, H.; Zhang, D.; Wang, G.; Song, W.; Liu, Y.; Ma, S.; Tang, Z.; Liu, Z.; Sakurai, K.; et al. Co-administration of combretastatin A4 nanoparticles and sorafenib for systemic therapy of hepatocellular carcinoma. Acta Biomater. 2019, 92, 229–240. [Google Scholar] [CrossRef] [PubMed]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | Peptides Sequence |
|---|---|
| J-I | PFKLSLHL-NH2 |
| Br-J-I | PFaKLSLHL-NH2 |
| Cl-J-I | PFbKLSLHL-NH2 |
| I-J-I | PFcKLSLHL-NH2 |
| Name | Br-J-I | Cl-J-I | I-J-I | J-I | MET |
|---|---|---|---|---|---|
| MIC | 5 μM | 10 μM | 10 μM | 160 μM | 0.125 μM |
| MBC | 10 μM | 40 μM | 20 μM | 320 μM | 0.5 μM |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jia, F.; Yu, Q.; Wang, R.; Zhao, L.; Yuan, F.; Guo, H.; Shen, Y.; He, F. Optimized Antimicrobial Peptide Jelleine-I Derivative Br-J-I Inhibits Fusobacterium Nucleatum to Suppress Colorectal Cancer Progression. Int. J. Mol. Sci. 2023, 24, 1469. https://doi.org/10.3390/ijms24021469
Jia F, Yu Q, Wang R, Zhao L, Yuan F, Guo H, Shen Y, He F. Optimized Antimicrobial Peptide Jelleine-I Derivative Br-J-I Inhibits Fusobacterium Nucleatum to Suppress Colorectal Cancer Progression. International Journal of Molecular Sciences. 2023; 24(2):1469. https://doi.org/10.3390/ijms24021469
Chicago/Turabian StyleJia, Fengjing, Qun Yu, Ruolei Wang, Ling Zhao, Fuwen Yuan, Haidong Guo, Yunhui Shen, and Feng He. 2023. "Optimized Antimicrobial Peptide Jelleine-I Derivative Br-J-I Inhibits Fusobacterium Nucleatum to Suppress Colorectal Cancer Progression" International Journal of Molecular Sciences 24, no. 2: 1469. https://doi.org/10.3390/ijms24021469