
Marine Natural Products with Activities against Prostate Cancer: Recent Discoveries

 ,
,  , and
, and 
Abstract
:
1. Introduction
2. Why Marine Organisms and Which Are the Marine-Derived Drugs on the Market?
3. Marine Microorganisms
3.1. Fungi
3.2. Bacteria
3.3. Cyanobacteria
3.4. Microalgae

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
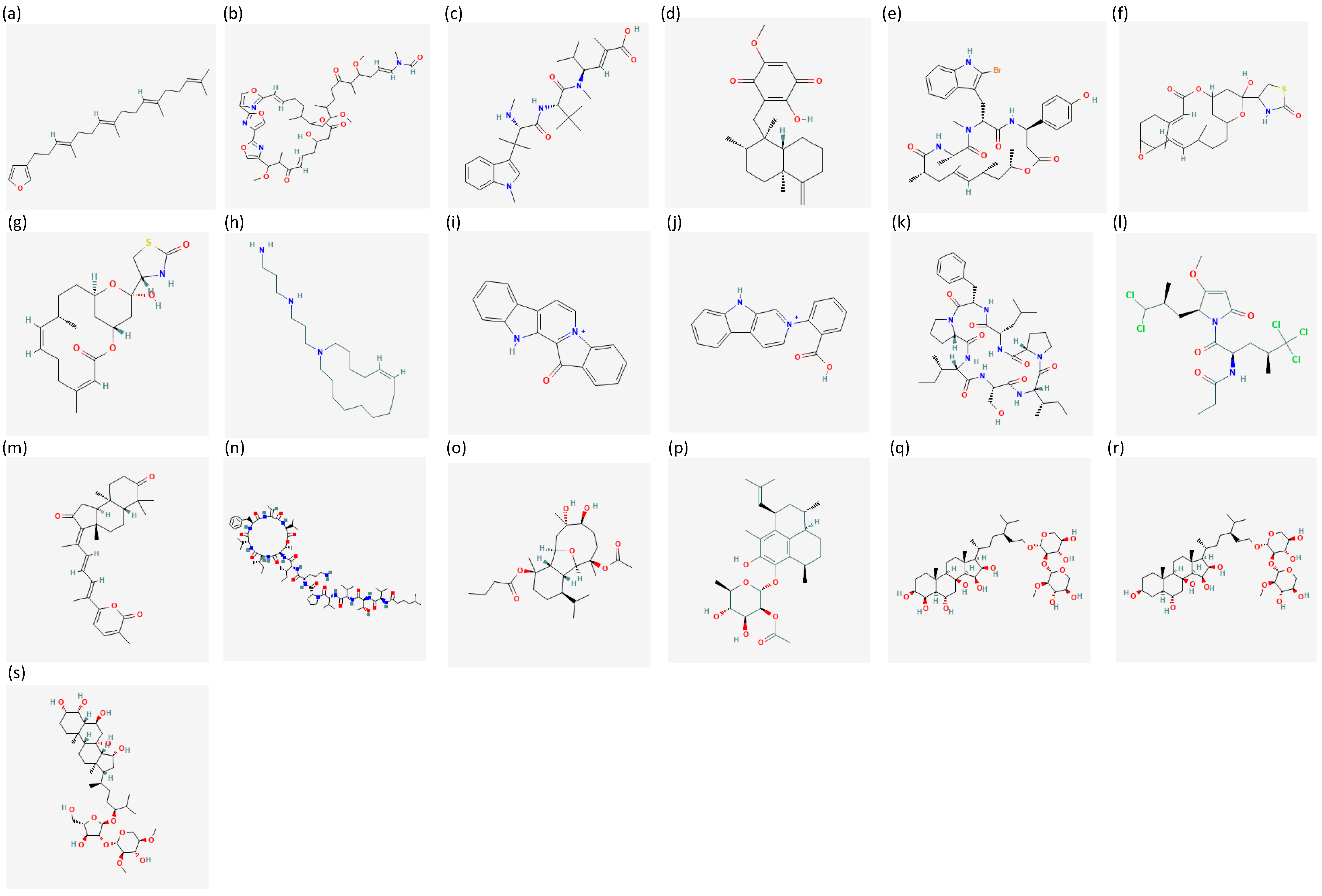{kind=link}
{kind=link}
| Organism | Compound/Extract | Active Concentration | Assay Information | Mechanism of Action | Ref. |
|---|---|---|---|---|---|
| Fungi | |||||
| Originally from the genus Fusarium | Compounds 5 and 11 (N-methylated sansalvamide A) | 10 µM | 72 h exposure; cell counting; PC3 cells | - | [41] |
| Isaria felina KMM 4639 | Isariketide A and compound 3 | 5 μM | 48 h exposure; MTT assay; PC3 cells | Apoptosis induction | [42] |
| Microsporum sp. MFS-YL | Physcion | 25 μM | 72 h exposure; MTT assay; PC3 cells | Apoptosis induction; Downregulation of Ras, Bcl-xL, and Bcl-2 proteins and upregulation of Bax, caspases-3, 8 and 9. | [43] |
| Mangrove-associated fungus from South China Sea | SZ-685C | IC50 3–9.6 μM | 48 h exposure; MTT assay; Annexin V assay | Apoptosis induction; Activation of caspase-8 and caspase-9 | [44] |
| Bacteria | |||||
| Chromobacterium sp. | Chromopeptide A | IC50 2.43 ± 0.02, 2.08 ± 0.16, and 1.75 ± 0.06 nmol/L, respectively | 72 h exposure; by use of sulforhodamine B; PC3, DU145 and LNCaP cells | Increased caspase-3 activity and PARP cleavage in each cell line and promoted G2/M phase arrest in PCa cells by suppressing cdc2 and cdc25C phosphorylation. | [49] |
| Chromobacterium sp. | Chromopeptide A | 1.6, 3.2 mg/kg, once a week for 18 d, in mice | PC3 cells were subcutaneously inoculated | Suppression of tumor growth | [49] |
| 39 Planctomycetes strains | Aqueous extracts | 4% | 72 h exposure; WST-1 assay metabolic assay; PC3 cells | - | [46] |
| 39 Planctomycetes strains | Organic extracts | 1% | 72 h exposure; WST-1 assay metabolic assay; PC3 cells | - | [46] |
| Streptomyces sp. | Lu01-M | IC50 1.03 ± 0.31, 2.12 ± 0.38, and 1.27 ± 0.25 μg/mL, respectively | 72 h exposure; MTT assay; PC3, DU145 and LNCaP cells | Apoptosis, necroptosis, autophagy, cell cycle arrest at the G2/M phase and DNA damage | [36] |
| Bacillus safensis MK541039 | L-asparaginase | IC50 of 38.75 IU/ml | 24 h exposure, by MTT assay, PC3 cells | Catalyzes the hydrolysis of L-asparagine to L-aspartic acid | [50] |
| Streptomyces sp. Mei37 | Mansouramycin C and 3-methyl-7-(methylamino)-5,8-isoquinolinedione | About 0.089 µM for mansouramycin C and 0.24 to 1.11 μM for 3-methyl-7-(methylamino)-5,8-isoquinolinedione | MTT assay; DU-145 cells | - | [47] |
| Cyanobacteria | |||||
| Cyanobacterium Lyngbya majuscule | Aurilide B | GI50 less than 10 nM | 48 h exposure, by MTT assay, panel of cells | - | [54] |
| Cyanobacterium Nostoc sp. GSV 224 | Cryptophycin 52 (LY355703) | 1–10 pM | 48 h of exposure, by cell cucle analysis, DNA ladder formation and detection of cytoplasmic nucleosomes; PC3, LNCaP and DU-145 cells | -Apoptosis induction | [57] |
| Cyanobacterium Lyngbya majuscule | Lagunamide C | 2.6 nM | 72 h exposure; by MTT assay; PC3 cells | - | [55] |
| Microalgae | |||||
| Raw marine microalgal material from Canada | Aqueous extracts | 5 mg/mL | 72 h exposure, by MTT assay; PC3 and DU145 cells | - | [59] |
4. Marine Macroorganisms
4.1. Sponges
4.2. Mollusks
4.3. Anthozoans
4.4. Other Macroorganisms
5. Marine-Derived Drugs in Clinical Trials against Prostate Cancer
6. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- World Health Organization International Agency for Research on Cancer (IARC). GLOBOCAN 2020: Estimated Cancer Incidence, Mortality and Prevalence Worldwide in 2020. Available online: https://Gco.Iarc.Fr/ (accessed on 1 December 2022).
- National Program of Cancer Registries and Surveillance, Epidemiology, and End Results Program SEER*Stat Database: NPCR and SEER Incidence—U.S. Cancer Statistics Public Use Research Database with Prostate Cancer—Cancer Stat Facts—SEER 12 (1992–2022), United States Depart-Ment of Health and Human Services, Centers for Disease Control and Prevention and National Cancer Institute. Available online: https://seer.cancer.gov/statfacts/html/prost.html (accessed on 15 November 2022).
- ECIS—European Cancer Information System. Prostate Cancer Fact Sheet in 2020. Available online: https://ecis.jrc.ec.europa.eu (accessed on 15 November 2022).
- Trama, A.; Foschi, R.; Larrañaga, N.; Sant, M.; Fuentes-Raspall, R.; Serraino, D.; Tavilla, A.; Van Eycken, L.; Nicolai, N.; Hackl, M.; et al. Survival of Male Genital Cancers (Prostate, Testis and Penis) in Europe 1999–2007: Results from the EUROCARE-5 Study. Eur. J. Cancer 2015, 51, 2206–2216. [Google Scholar] [CrossRef] [PubMed]
- Mazhar, D. Prostate Cancer. Postgrad. Med. J. 2002, 78, 590–595. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ashley, T. Using Predictive Value, Sensitivity and Specificity to Interpret Laboratory Tests: PSA for the Diagnosis of Prostate Cancer. J. Insur. Med. 2005, 37, 261–263. [Google Scholar] [PubMed]
- Zamboglou, C.; Schiller, F.; Fechter, T.; Wieser, G.; Jilg, C.A.; Chirindel, A.; Salman, N.; Drendel, V.; Werner, M.; Mix, M.; et al. 68 Ga-HBED-CC-PSMA PET/CT Versus Histopathology In Primary Localized Prostate Cancer: A Voxel-Wise Comparison. Theranostics 2016, 6, 1619–1628. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barakzai, M.A. Prostatic Adenocarcinoma: A Grading from Gleason to the New Grade-Group System: A Historical and Critical Review. Asian Pac. J. Cancer Prev. 2019, 20, 661–666. [Google Scholar] [CrossRef]
- National Cancer Registration & Analysis Service and Cancer Research UK. Chemotherapy, Radiotherapy and Tu-Mour Resections in England: 2013–2014; Workbook; NCRAS: London, UK, 2017.
- Perlmutter, M.A.; Lepor, H. Androgen Deprivation Therapy in the Treatment of Advanced Prostate Cancer. Rev. Urol. 2007, 9 (Suppl. S1), S3–S8. [Google Scholar]
- Fujita, K.; Nonomura, N. Role of Androgen Receptor in Prostate Cancer: A Review. World J. Mens Health 2019, 37, 288. [Google Scholar] [CrossRef]
- Feng, Z.; Graff, J.N. Next-Generation Androgen Receptor-Signaling Inhibitors for Prostate Cancer: Considerations for Older Patients. Drugs Aging 2021, 38, 111–123. [Google Scholar] [CrossRef]
- Baciarello, G.; Delva, R.; Gravis, G.; Tazi, Y.; Beuzeboc, P.; Gross-Goupil, M.; Bompas, E.; Joly, F.; Greilsamer, C.; Hon, T.N.T.; et al. Patient Preference Between Cabazitaxel and Docetaxel for First-Line Chemotherapy in Metastatic Castration-Resistant Prostate Cancer: The CABADOC Trial. Eur. Urol. 2022, 81, 234–240. [Google Scholar] [CrossRef]
- Wang, Y.; Wang, Y.; Ci, X.; Choi, S.Y.C.; Crea, F.; Lin, D.; Wang, Y. Molecular Events in Neuroendocrine Prostate Cancer Development. Nat. Rev. Urol. 2021, 18, 581–596. [Google Scholar] [CrossRef]
- Silvestri, R.; Pucci, P.; Venalainen, E.; Matheou, C.; Mather, R.; Chandler, S.; Aceto, R.; Rigas, S.H.; Wang, Y.; Rietdorf, K.; et al. T-type Calcium Channels Drive the Proliferation of Androgen-receptor Negative Prostate Cancer Cells. Prostate 2019, pros.23879. [Google Scholar] [CrossRef]
- Mather, R.L.; Parolia, A.; Carson, S.E.; Venalainen, E.; Roig-Carles, D.; Jaber, M.; Chu, S.; Alborelli, I.; Wu, R.; Lin, D.; et al. The Evolutionarily Conserved Long Non-coding RNA LINC00261 Drives Neuroendocrine Prostate Cancer Proliferation and Metastasis via Distinct Nuclear and Cytoplasmic Mechanisms. Mol. Oncol. 2021, 15, 1921–1941. [Google Scholar] [CrossRef]
- Pucci, P.; Venalainen, E.; Alborelli, I.; Quagliata, L.; Hawkes, C.; Mather, R.; Romero, I.; Rigas, S.H.; Wang, Y.; Crea, F. LncRNA HORAS5 Promotes Taxane Resistance in Castration-Resistant Prostate Cancer via a BCL2A1-Dependent Mechanism. Epigenomics 2020, 12, 1123–1138. [Google Scholar] [CrossRef]
- Santos, J.D.; Vitorino, I.; Reyes, F.; Vicente, F.; Lage, O.M. From Ocean to Medicine: Pharmaceutical Applications of Metabolites from Marine Bacteria. Antibiotics 2020, 9, 455. [Google Scholar] [CrossRef]
- World Register of Marine Species. Available online: https://www.marinespecies.org/ (accessed on 20 December 2022).
- Available online: http://www.coml.org/index.html (accessed on 20 December 2022).
- MarinLit Dedicated to Marine Natural Products Research. Available online: https://pubs.rsc.org/marinlit (accessed on 20 December 2022).
- Jiménez, C. Marine Natural Products in Medicinal Chemistry. ACS Med. Chem. Lett. 2018, 9, 959–961. [Google Scholar] [CrossRef] [Green Version]
- Carroll, A.R.; Copp, B.R.; Davis, R.A.; Keyzers, R.A.; Prinsep, M.R. Marine Natural Products. Nat. Prod. Rep. 2019, 36, 122–173. [Google Scholar] [CrossRef] [Green Version]
- Martínez, K.A.; Saide, A.; Crespo, G.; Martín, J.; Romano, G.; Reyes, F.; Lauritano, C.; Ianora, A. Promising Antiproliferative Compound From the Green Microalga Dunaliella Tertiolecta Against Human Cancer Cells. Front. Mar. Sci. 2022, 9, 778108. [Google Scholar] [CrossRef]
- Saide, A.; Martínez, K.A.; Ianora, A.; Lauritano, C. Unlocking the Health Potential of Microalgae as Sustainable Sources of Bioactive Compounds. IJMS 2021, 22, 4383. [Google Scholar] [CrossRef]
- Riccio, G.; Nuzzo, G.; Zazo, G.; Coppola, D.; Senese, G.; Romano, L.; Costantini, M.; Ruocco, N.; Bertolino, M.; Fontana, A.; et al. Bioactivity Screening of Antarctic Sponges Reveals Anticancer Activity and Potential Cell Death via Ferroptosis by Mycalols. Mar. Drugs 2021, 19, 459. [Google Scholar] [CrossRef]
- Riccio, G.; Martinez, K.A.; Martín, J.; Reyes, F.; D’Ambra, I.; Lauritano, C. Jellyfish as an Alternative Source of Bioactive Antiproliferative Compounds. Mar. Drugs 2022, 20, 350. [Google Scholar] [CrossRef]
- National Center for Biotechnology Information. PubChem Compound Summary for CID 10395173, Mmaf. 2022. Available online: https://pubchem.ncbi.nlm.nih.gov/compound/10395173#section=2D-Structure (accessed on 30 December 2022).
- National Center for Biotechnology Information. PubChem Compound Summary for CID 11542188, Mmae. 2022. Available online: https://pubchem.ncbi.nlm.nih.gov/compound/11542188#section=2D-Structure (accessed on 30 December 2022).
- National Center for Biotechnology Information. PubChem Compound Summary for CID 11354606, Eribulin. 2022. Available online: https://pubchem.ncbi.nlm.nih.gov/compound/11354606#section=2D-Structure (accessed on 30 December 2022).
- National Center for Biotechnology Information. PubChem Compound Summary for CID 6253, Cytarabine. 2022. Available online: https://pubchem.ncbi.nlm.nih.gov/compound/6253#section=2D-Structure (accessed on 30 December 2022).
- National Center for Biotechnology Information. PubChem Compound Summary for CID 57327016, Lurbinectedin. Available online: https://pubchem.ncbi.nlm.nih.gov/compound/57327016#section=2D-Structure (accessed on 30 December 2022).
- National Center for Biotechnology Information. PubChem Compound Summary for CID 9812534, Plitidepsin. 2022. Available online: https://pubchem.ncbi.nlm.nih.gov/compound/9812534#section=2D-Structure (accessed on 30 December 2022).
- National Center for Biotechnology Information. PubChem Compound Summary for CID 108150, Trabectedin. 2022. Available online: https://pubchem.ncbi.nlm.nih.gov/compound/108150#section=2D-Structure (accessed on 30 December 2022).
- National Center for Biotechnology Information. 2022. Available online: https://pubchem.ncbi.nlm.nih.gov/ (accessed on 30 December 2022).
- Liu, S.N.; Li, C. Clinical Pharmacology Strategies in Supporting Drug Development and Approval of Antibody–Drug Conjugates in Oncology. Cancer Chemother. Pharm. 2021, 87, 743–765. [Google Scholar] [CrossRef]
- Ahmed, S.; Alam, W.; Jeandet, P.; Aschner, M.; Alsharif, K.F.; Saso, L.; Khan, H. Therapeutic Potential of Marine Peptides in Prostate Cancer: Mechanistic Insights. Mar. Drugs 2022, 20, 466. [Google Scholar] [CrossRef] [PubMed]
- Kalimuthu, S.; Venkatesan, J.; Kim, S.-K. Marine Derived Bioactive Compounds for Breast and Prostate Cancer Treatment: A Review. CBC 2014, 10, 62–74. [Google Scholar] [CrossRef]
- Ruiz-Torres, V.; Encinar, J.; Herranz-López, M.; Pérez-Sánchez, A.; Galiano, V.; Barrajón-Catalán, E.; Micol, V. An Updated Review on Marine Anticancer Compounds: The Use of Virtual Screening for the Discovery of Small-Molecule Cancer Drugs. Molecules 2017, 22, 1037. [Google Scholar] [CrossRef] [PubMed]
- Fan, M.; Nath, A.; Tang, Y.; Choi, Y.-J.; Debnath, T.; Choi, E.-J.; Kim, E.-K. Investigation of the Anti-Prostate Cancer Properties of Marine-Derived Compounds. Mar. Drugs 2018, 16, 160. [Google Scholar] [CrossRef] [Green Version]
- Liu, S.; Gu, W.; Lo, D.; Ding, X.-Z.; Ujiki, M.; Adrian, T.E.; Soff, G.A.; Silverman, R.B. N -Methylsansalvamide A Peptide Analogues. Potent New Antitumor Agents. J. Med. Chem. 2005, 48, 3630–3638. [Google Scholar] [CrossRef]
- Smetanina, O.F.; Yurchenko, A.N.; Ivanets, E.V.; Kalinovsky, A.I.; Khudyakova, Y.V.; Dyshlovoy, S.A.; von Amsberg, G.; Yurchenko, E.A.; Afiyatullov, S.S. Unique Prostate Cancer-Toxic Polyketides from Marine Sediment-Derived Fungus Isaria Felina. J. Antibiot. 2017, 70, 856–858. [Google Scholar] [CrossRef]
- Ding, Y.-S.; Kim, W.-S.; Park, S.J.; Kim, S.-K. Apoptotic Effect of Physcion Isolated from Marine Fungus Microsporum Sp. in PC3 Human Prostate Cancer Cells. Fish Aquat. Sci. 2018, 21, 22. [Google Scholar] [CrossRef]
- Xie, G.; Zhu, X.; Li, Q.; Gu, M.; He, Z.; Wu, J.; Li, J.; Lin, Y.; Li, M.; She, Z.; et al. SZ-685C, a Marine Anthraquinone, Is a Potent Inducer of Apoptosis with Anticancer Activity by Suppression of the Akt/FOXO Pathway: SZ-685C Induces Apoptosis and Inhibits Tumour Growth. Br. J. Pharmacol. 2010, 159, 689–697. [Google Scholar] [CrossRef] [Green Version]
- Wei, J.; Gou, Z.; Wen, Y.; Luo, Q.; Huang, Z. Marine Compounds Targeting the PI3K/Akt Signaling Pathway in Cancer Therapy. Biomed. Pharmacother. 2020, 129, 110484. [Google Scholar] [CrossRef]
- Calisto, R.; Sæbø, E.F.; Storesund, J.E.; Øvreås, L.; Herfindal, L.; Lage, O.M. Anticancer Activity in Planctomycetes. Front. Mar. Sci. 2019, 5, 499. [Google Scholar] [CrossRef]
- Hawas, U.W.; Shaaban, M.; Shaaban, K.A.; Speitling, M.; Maier, A.; Kelter, G.; Fiebig, H.H.; Meiners, M.; Helmke, E.; Laatsch, H. Mansouramycins A−D, Cytotoxic Isoquinolinequinones from a Marine Streptomycete. J. Nat. Prod. 2009, 72, 2120–2124. [Google Scholar] [CrossRef]
- Lin, H.-Y.; Lin, Y.-S.; Shih, S.-P.; Lee, S.-B.; El-Shazly, M.; Chang, K.-M.; Yang, Y.-C.S.H.; Lee, Y.-L.; Lu, M.-C. The Anti-Proliferative Activity of Secondary Metabolite from the Marine Streptomyces sp. against Prostate Cancer Cells. Life 2021, 11, 1414. [Google Scholar] [CrossRef]
- Sun, J.; Wang, J.; Wang, X.; Liu, H.; Zhang, M.; Liu, Y.-C.; Zhang, C.; Su, Y.; Shen, Y.; Guo, Y.; et al. Marine-Derived Chromopeptide A, a Novel Class I HDAC Inhibitor, Suppresses Human Prostate Cancer Cell Proliferation and Migration. Acta Pharm. Sin 2017, 38, 551–560. [Google Scholar] [CrossRef] [Green Version]
- Bakeer, W.; Amer, M.; Hozayen, W.; Kotb, N.; Hassan, M.H.A. Isolation of Asparaginase-Producing Microorganisms and Evaluation of the Enzymatic Antitumor Activity. Egypt Pharm. J. 2022, 21, 282. [Google Scholar] [CrossRef]
- Lauritano, C.; De Luca, D.; Ferrarini, A.; Avanzato, C.; Minio, A.; Esposito, F.; Ianora, A. De Novo Transcriptome of the Cosmopolitan Dinoflagellate Amphidinium Carterae to Identify Enzymes with Biotechnological Potential. Sci. Rep. 2017, 7, 11701. [Google Scholar] [CrossRef] [Green Version]
- Dias, F.F.G.; Ruiz, A.L.T.G.; Torre, A.D.; Sato, H.H. Purification, Characterization and Antiproliferative Activity of l-Asparaginase from Aspergillus Oryzae CCT 3940 with No Glutaminase Activity. Asian Pac. J. Trop. Biomed. 2016, 6, 785–794. [Google Scholar] [CrossRef] [Green Version]
- Vingiani, G.M.; De Luca, P.; Ianora, A.; Dobson, A.D.W.; Lauritano, C. Microalgal Enzymes with Biotechnological Applications. Mar. Drugs 2019, 17, 459. [Google Scholar] [CrossRef] [Green Version]
- Han, B.; Gross, H.; Goeger, D.E.; Mooberry, S.L.; Gerwick, W.H. Aurilides B and C, Cancer Cell Toxins from a Papua New Guinea Collection of the Marine Cyanobacterium Lyngbya m Ajuscula. J. Nat. Prod. 2006, 69, 572–575. [Google Scholar] [CrossRef]
- Tripathi, A.; Puddick, J.; Prinsep, M.R.; Rottmann, M.; Chan, K.P.; Chen, D.Y.-K.; Tan, L.T. Lagunamide C, a Cytotoxic Cyclodepsipeptide from the Marine Cyanobacterium Lyngbya Majuscula. Phytochemistry 2011, 72, 2369–2375. [Google Scholar] [CrossRef]
- Trimurtulu, G.; Ohtani, I.; Patterson, G.M.L.; Moore, R.E.; Corbett, T.H.; Valeriote, F.A.; Demchik, L. Total Structures of Cryptophycins, Potent Antitumor Depsipeptides from the Blue-Green Alga Nostoc Sp. Strain GSV 224. J. Am. Chem. Soc. 1994, 116, 4729–4737. [Google Scholar] [CrossRef]
- Drew, L.; Fine, R.L.; Do, T.N.; Douglas, G.P.; Petrylak, D.P. The Novel Antimicrotubule Agent Cryptophycin 52 (LY355703) Induces Apoptosis via Multiple Pathways in Human Prostate Cancer Cells. Clin. Cancer Res. 2002, 8, 3922–3932. [Google Scholar] [PubMed]
- Martínez Andrade, K.; Lauritano, C.; Romano, G.; Ianora, A. Marine Microalgae with Anti-Cancer Properties. Mar. Drugs 2018, 16, 165. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Somasekharan, S.P.; El-Naggar, A.; Sorensen, P.H.; Wang, Y.; Cheng, H. An Aqueous Extract of Marine Microalgae Exhibits Antimetastatic Activity through Preferential Killing of Suspended Cancer Cells and Anticolony Forming Activity. Evid.-Based Complement. Altern. Med. 2016, 2016, 1–8. [Google Scholar] [CrossRef] [Green Version]
- National Center for Biotechnology Information. PubChem Compound Summary for CID 139589440, Isariketide B. 2022. Available online: https://pubchem.ncbi.nlm.nih.gov/compound/139589440#section=2D-Structure (accessed on 2 January 2023).
- National Center for Biotechnology Information. PubChem Compound Summary for CID 10639, Physcion. 2022. Available online: https://pubchem.ncbi.nlm.nih.gov/compound/10639#section=2D-Structure (accessed on 2 January 2023).
- National Center for Biotechnology Information. PubChem Compound Summary for CID 12301973, Haloroquinone. 2022. Available online: https://pubchem.ncbi.nlm.nih.gov/compound/12301973#section=2D-Structure (accessed on 2 January 2023).
- National Center for Biotechnology Information. PubChem Compound Summary for CID 44614386, Mansouramycin C. 2022. Available online: https://pubchem.ncbi.nlm.nih.gov/compound/Mansouramycin-C#section=2D-Structure (accessed on 2 January 2023).
- National Center for Biotechnology Information. PubChem Compound Summary for CID 11721984, Aurilide B. 2022. Available online: https://pubchem.ncbi.nlm.nih.gov/compound/11721984#section=2D-Structure (accessed on 2 January 2023).
- National Center for Biotechnology Information. PubChem Compound Summary for CID 9939639, Cryptophycin 52. 2022. Available online: https://pubchem.ncbi.nlm.nih.gov/compound/9939639#section=2D-Structure (accessed on 2 January 2023).
- National Center for Biotechnology Information. PubChem Compound Summary for CID 56839938, Lagunamide C. 2022. Available online: https://pubchem.ncbi.nlm.nih.gov/compound/56839938#section=2D-Structure (accessed on 2 January 2023).
- Yong, Y.S.; Lim, S.-C.; Lee, P.-C.; Ling, Y.-S. Sponges from North Borneo and Their Bioactivity against Human Colorectal Cancer Cells. Borneo J. Mar. Sci. Aqua 2018, 2, 40–47. [Google Scholar] [CrossRef]
- Dyshlovoy, S.A.; Honecker, F. Marine Compounds and Cancer: Where Do We Stand? Mar. Drugs 2015, 13, 5657–5665. [Google Scholar] [CrossRef]
- Seccatore, J.; de Tomi, G.; Veiga, M. Efficiency as a Road to Sustainability in Small Scale Mining. MSF 2014, 805, 395–402. [Google Scholar] [CrossRef]
- Senderowicz, A.M.J.; Kaur, G.; Sainz, E.; Laing, C.; Inman, W.D.; Rodriguez, J.; Crews, P.; Malspeis, L.; Grever, M.R.; Sausville, E.A.; et al. Jasplakinolide’s Inhibition of the Growth of Prostate Carcinoma Cells In Vitro With Disruption of the Actin Cytoskeleton. JNCI J. Natl. Cancer Inst. 1995, 87, 46–51. [Google Scholar] [CrossRef]
- Roskelley, C.D.; Williams, D.E.; McHardy, L.M.; Leong, K.G.; Troussard, A.; Karsan, A.; Andersen, R.J.; Dedhar, S.; Roberge, M. Inhibition of Tumor Cell Invasion and Angiogenesis by Motuporamines. Cancer Res. 2001, 61, 6788–6794. [Google Scholar]
- Liu, W.K.; Cheung, F.W.K.; Che, C.-T. Stellettin A Induces Oxidative Stress and Apoptosis in HL-60 Human Leukemia and LNCaP Prostate Cancer Cell Lines. J. Nat. Prod. 2006, 69, 934–937. [Google Scholar] [CrossRef]
- Sadar, M.D.; Williams, D.E.; Mawji, N.R.; Patrick, B.O.; Wikanta, T.; Chasanah, E.; Irianto, H.E.; Soest, R.V.; Andersen, R.J. Sintokamides A to E, Chlorinated Peptides from the Sponge Dysidea Sp. That Inhibit Transactivation of the N-Terminus of the Androgen Receptor in Prostate Cancer Cells. Org. Lett. 2008, 10, 4947–4950. [Google Scholar] [CrossRef]
- Sayed, K.A.E.; Khanfar, M.A.; Shallal, H.M.; Muralidharan, A.; Awate, B.; Youssef, D.T.A.; Liu, Y.; Zhou, Y.-D.; Nagle, D.G.; Shah, G. Latrunculin A and Its C-17- O -Carbamates Inhibit Prostate Tumor Cell Invasion and HIF-1 Activation in Breast Tumor Cells. J. Nat. Prod. 2008, 71, 396–402. [Google Scholar] [CrossRef]
- Khanfar, M.A.; Asal, B.A.; Mudit, M.; Kaddoumi, A.; El Sayed, K.A. The Marine Natural-Derived Inhibitors of Glycogen Synthase Kinase-3β Phenylmethylene Hydantoins: In Vitro and in Vivo Activities and Pharmacophore Modeling. Bioorg. Med. Chem. 2009, 17, 6032–6039. [Google Scholar] [CrossRef] [Green Version]
- Shah, G.V.; Muralidharan, A.; Thomas, S.; Gokulgandhi, M.; Mudit, M.; Khanfar, M.; El Sayed, K. Identification of a Small Molecule Class to Enhance Cell-Cell Adhesion and Attenuate Prostate Tumor Growth and Metastasis. Mol. Cancer Ther. 2009, 8, 509–520. [Google Scholar] [CrossRef] [Green Version]
- Mudit, M.; El Sayed, K.A. Optimization of (Phenylmethylidene)-Hydantoins as Prostate Cancer Migration Inhibitors: SAR-Directed Design, Synthesis, and Pharmacophore Modeling. Chem. Biodivers. 2011, 8, 1470–1485. [Google Scholar] [CrossRef]
- Elgoud Said, A.A.; Mahmoud, B.K.; Attia, E.Z.; Abdelmohsen, U.R.; Fouad, M.A. Bioactive Natural Products from Marine Sponges Belonging to Family Hymedesmiidae. RSC Adv. 2021, 11, 16179–16191. [Google Scholar] [CrossRef]
- Williams, P.G. Panning for Chemical Gold: Marine Bacteria as a Source of New Therapeutics. Trends Biotechnol. 2009, 27, 45–52. [Google Scholar] [CrossRef]
- Holland, I.P.; McCluskey, A.; Sakoff, J.A.; Gilbert, J.; Chau, N.; Robinson, P.J.; Motti, C.A.; Wright, A.D.; van Altena, I.A. Steroids from an Australian Sponge Psammoclema Sp. J. Nat. Prod. 2009, 72, 102–106. [Google Scholar] [CrossRef]
- Wang, F.; Ezell, S.J.; Zhang, Y.; Wang, W.; Rayburn, E.R.; Nadkarni, D.H.; Murugesan, S.; Velu, S.E.; Zhang, R. FBA-TPQ, a Novel Marine-Derived Compound as Experimental Therapy for Prostate Cancer. Invest New Drugs 2010, 28, 234–241. [Google Scholar] [CrossRef]
- Kudryavtsev, D.; Spirova, E.; Shelukhina, I.; Son, L.; Makarova, Y.; Utkina, N.; Kasheverov, I.; Tsetlin, V. Makaluvamine G from the Marine Sponge Zyzzia Fuliginosa Inhibits Muscle NAChR by Binding at the Orthosteric and Allosteric Sites. Mar. Drugs 2018, 16, 109. [Google Scholar] [CrossRef] [Green Version]
- Shin, Y.; Kim, G.; Jeon, J.; Shin, J.; Lee, S. Antimetastatic Effect of Halichondramide, a Trisoxazole Macrolide from the Marine Sponge Chondrosia Corticata, on Human Prostate Cancer Cells via Modulation of Epithelial-to-Mesenchymal Transition. Mar. Drugs 2013, 11, 2472–2485. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shin, J.; Jin, M. Potential Immunotherapeutics for Immunosuppression in Sepsis. Biomol. Ther. 2017, 25, 569–577. [Google Scholar] [CrossRef] [PubMed]
- Du, L.; Zhou, Y.-D.; Nagle, D.G. Inducers of Hypoxic Response: Marine Sesquiterpene Quinones Activate HIF-1. J. Nat. Prod. 2013, 76, 1175–1181. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chiu, S.C.; Huang, S.Y.; Chen, S.P.; Su, C.C.; Chiu, T.L.; Pang, C.Y. Tanshinone IIA Inhibits Human Prostate Cancer Cells Growth by Induction of Endoplasmic Reticulum Stress in Vitro and in Vivo. Prostate Cancer Prostatic Dis. 2013, 16, 315–322. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tran, T.; Pham, N.; Fechner, G.; Hooper, J.; Quinn, R. Potent Cytotoxic Peptides from the Australian Marine Sponge Pipestela Candelabra. Mar. Drugs 2014, 12, 3399–3415. [Google Scholar] [CrossRef] [Green Version]
- Khokhar, S.; Feng, Y.; Campitelli, M.R.; Ekins, M.G.; Hooper, J.N.A.; Beattie, K.D.; Sadowski, M.C.; Nelson, C.C.; Davis, R.A. Isolation, Structure Determination and Cytotoxicity Studies of Tryptophan Alkaloids from an Australian Marine Sponge Hyrtios Sp. Bioorg. Med. Chem. Lett. 2014, 24, 3329–3332. [Google Scholar] [CrossRef]
- Arai, M.; Kawachi, T.; Kotoku, N.; Nakata, C.; Kamada, H.; Tsunoda, S.; Tsutsumi, Y.; Endo, H.; Inoue, M.; Sato, H.; et al. Furospinosulin-1, Marine Spongean Furanosesterterpene, Suppresses the Growth of Hypoxia-Adapted Cancer Cells by Binding to Transcriptional Regulators P54 nrb and LEDGF/P75. ChemBioChem 2016, 17, 181–189. [Google Scholar] [CrossRef]
- Murniasih, T.; Putra, M.Y.; Bayu, A.; Wibowo, J.T. A Review on Diversity of Anticancer Compounds Derived from Indonesian Marine Sponges. IOP Conf. Ser.: Mater. Sci. Eng. 2021, 1192, 012012. [Google Scholar] [CrossRef]
- Makarieva, T.N.; Denisenko, V.A.; Stonik, V.A.; Milgrom, Y.M.; Rashkes, Y.V. Rhizochalin, a Novel Secondary Metabolite of Mixed Biosynthesis from the Sponge Rhizochalina Incrustata. Tetrahedron Lett. 1989, 30, 6581–6584. [Google Scholar] [CrossRef]
- Dyshlovoy, S.A.; Otte, K.; Alsdorf, W.H.; Hauschild, J.; Lange, T.; Venz, S.; Bauer, C.K.; Bähring, R.; Amann, K.; Mandanchi, R.; et al. Marine Compound Rhizochalinin Shows High in Vitro and in Vivo Efficacy in Castration Resistant Prostate Cancer. Oncotarget 2016, 7, 69703–69717. [Google Scholar] [CrossRef] [Green Version]
- Palapattu, G.S. Commentary on “AR-V7 and Resistance to Enzalutamide and Abiraterone in Prostate Cancer.” Antonarakis E.S.; Lu C.; Wang H.; Luber B.; Nakazawa M.; Roeser J.C.; Chen Y.; Mohammad T.A.; Chen Y.; Fedor H.L.; Lotan T.L.; Zheng Q.; De Marzo A.M.; Isaacs J.T.; Isaacs W.B.; Nadal R.; Paller C.J.; Denmeade S.R.; Carducci M.A.; Eisenberger M.A.; Luo J.; Division of Urologic Oncology, Department of Urology, University of Michigan, MI. N Engl J Med 2014; 371(11):1028-38. Urol. Oncol. Semin. Orig. Investig. 2016, 34, 520. [Google Scholar] [CrossRef]
- Dal Pra, A.; Locke, J.A.; Borst, G.; Supiot, S.; Bristow, R.G. Mechanistic Insights into Molecular Targeting and Combined Modality Therapy for Aggressive, Localized Prostate Cancer. Front. Oncol. 2016, 6. [Google Scholar] [CrossRef]
- Prevarskaya, N.; Skryma, R.; Bidaux, G.; Flourakis, M.; Shuba, Y. Ion Channels in Death and Differentiation of Prostate Cancer Cells. Cell Death Differ. 2007, 14, 1295–1304. [Google Scholar] [CrossRef]
- Bary, K.; Elamraoui, B.; Laasri, F.E.; El Mzibri, M.; Benbacer, L.; Bamhaoud, T. Cytotoxic Effect of Extracts from the Moroccan Marine Sponge on Human Prostate Cancer Cell Line. Int. J. New Technol. Res. 2018, 4, 74–78. [Google Scholar]
- Jomori, T.; Setiawan, A.; Sasaoka, M.; Arai, M. Cytotoxicity of New Diterpene Alkaloids, Ceylonamides G-I, Isolated From Indonesian Marine Sponge of Spongia Sp. Nat. Prod. Commun. 2019, 14, 1934578X1985729. [Google Scholar] [CrossRef] [Green Version]
- Panggabean, J.A.; Adiguna, S.P.; Murniasih, T.; Rahmawati, S.I.; Bayu, A.; Putra, M.Y. Structure–Activity Relationship of Cytotoxic Natural Products from Indonesian Marine Sponges. Rev. Bras. Farmacogn. 2022, 32, 12–38. [Google Scholar] [CrossRef]
- Shubina, L.K.; Makarieva, T.N.; Denisenko, V.A.; Popov, R.S.; Dyshlovoy, S.A.; Grebnev, B.B.; Dmitrenok, P.S.; von Amsberg, G.; Stonik, V.A. Gracilosulfates A–G, Monosulfated Polyoxygenated Steroids from the Marine Sponge Haliclona Gracilis. Mar. Drugs 2020, 18, 454. [Google Scholar] [CrossRef]
- Dyshlovoy, S.A.; Shubina, L.K.; Makarieva, T.N.; Hauschild, J.; Strewinsky, N.; Guzii, A.G.; Menshov, A.S.; Popov, R.S.; Grebnev, B.B.; Busenbender, T.; et al. New Diterpenes from the Marine Sponge Spongionella Sp. Overcome Drug Resistance in Prostate Cancer by Inhibition of P-Glycoprotein. Sci. Rep. 2022, 12, 13570. [Google Scholar] [CrossRef]
- Yang, Y.C.; Meimetis, L.G.; Tien, A.H.; Mawji, N.R.; Carr, G.; Wang, J.; Andersen, R.J.; Sadar, M.D. Spongian Diterpenoids Inhibit Androgen Receptor Activity. Mol. Cancer Ther. 2013, 12, 621–631. [Google Scholar] [CrossRef] [Green Version]
- Nelson, P.S. Targeting the Androgen Receptor in Prostate Cancer—A Resilient Foe. N. Engl. J. Med. 2014, 371, 1067–1069. [Google Scholar] [CrossRef] [Green Version]
- Suárez, Y.; González, L.; Cuadrado, A.; Berciano, M.; Lafarga, M.; Muñoz, A. Kahalalide F, a New Marine-Derived Compound, Induces Oncosis in Human Prostate and Breast Cancer Cells. Mol. Cancer Ther. 2003, 2, 863–872. [Google Scholar] [PubMed]
- Hegazy, M.-E.F.; Moustfa, A.Y.; Mohamed, A.E.-H.H.; Alhammady, M.A.; Elbehairi, S.E.I.; Ohta, S.; Paré, P.W. New Cytotoxic Halogenated Sesquiterpenes from the Egyptian Sea Hare, Aplysia Oculifera. Tetrahedron Lett. 2014, 55, 1711–1714. [Google Scholar] [CrossRef]
- Hassan, H.M.; Khanfar, M.A.; Elnagar, A.Y.; Mohammed, R.; Shaala, L.A.; Youssef, D.T.A.; Hifnawy, M.S.; El Sayed, K.A. Pachycladins A−E, Prostate Cancer Invasion and Migration Inhibitory Eunicellin-Based Diterpenoids from the Red Sea Soft Coral Cladiella Pachyclados. J. Nat. Prod. 2010, 73, 848–853. [Google Scholar] [CrossRef] [PubMed]
- Bowers, Z.; Famuyiwa, T.; Caraballo, D.; Jebelli, J.; Kumi-Diaka, J.; West, L. Therapeutic Potential of Pseudopterosin H on a Prostate Cancer Cell Line. FASEB J. 2021, 35, fasebj.2021.35.S1.04601. [Google Scholar] [CrossRef]
- Li, X.; Tang, Y.; Yu, F.; Sun, Y.; Huang, F.; Chen, Y.; Yang, Z.; Ding, G. Inhibition of Prostate Cancer DU-145 Cells Proliferation by Anthopleura Anjunae Oligopeptide (YVPGP) via PI3K/AKT/MTOR Signaling Pathway. Mar. Drugs 2018, 16, 325. [Google Scholar] [CrossRef] [Green Version]
- Kapustina, I.I.; Makarieva, T.N.; Guzii, A.G.; Kalinovsky, A.I.; Popov, R.S.; Dyshlovoy, S.A.; Grebnev, B.B.; von Amsberg, G.; Stonik, V.A. Leptogorgins A–C, Humulane Sesquiterpenoids from the Vietnamese Gorgonian Leptogorgia Sp. Mar. Drugs 2020, 18, 310. [Google Scholar] [CrossRef]
- Guha, P.; Kaptan, E.; Bandyopadhyaya, G.; Kaczanowska, S.; Davila, E.; Thompson, K.; Martin, S.S.; Kalvakolanu, D.V.; Vasta, G.R.; Ahmed, H. Cod Glycopeptide with Picomolar Affinity to Galectin-3 Suppresses T-Cell Apoptosis and Prostate Cancer Metastasis. Proc. Natl. Acad. Sci. USA 2013, 110, 5052–5057. [Google Scholar] [CrossRef] [Green Version]
- Ngoan, B.T.; Hanh, T.T.H.; Vien, L.T.; Diep, C.N.; Thao, N.P.; Thao, D.T.; Thanh, N.V.; Cuong, N.X.; Nam, N.H.; Thung, D.C.; et al. Asterosaponins and Glycosylated Polyhydroxysteroids from the Starfish Culcita Novaeguineae and Their Cytotoxic Activities. J. Asian Nat. Prod. Res. 2015, 17, 1010–1017. [Google Scholar] [CrossRef]
- Bjerknes, C.; Framroze, B.; Currie, C.; Pettersen, C.H.H.; Axcrona, K.; Hermansen, E. Salmon Protein Hydrolysate Potentiates the Growth Inhibitory Effect of Bicalutamide on Human Prostate Cancer Cell Lines LNCaP and PC3 by Modulating Iron Homeostasis. Mar. Drugs 2022, 20, 228. [Google Scholar] [CrossRef]
- National Center for Biotechnology Information. PubChem Compound Summary for CID 1593085, Furospinosulin-1. 2022. Available online: https://pubchem.ncbi.nlm.nih.gov/compound/15930585#section=2D-Structure (accessed on 2 January 2023).
- National Center for Biotechnology Information. PubChem Compound Summary for CID: 10010725, Halichondramide. 2022. Available online: https://pubchem.ncbi.nlm.nih.gov/compound/Halichondramide#section=2D-Structure (accessed on 2 January 2023).
- National Center for Biotechnology Information. PubChem Compound Summary for CID 5352092, Hemiasterlin. 2022. Available online: https://pubchem.ncbi.nlm.nih.gov/compound/5352092#section=2D-Structure (accessed on 2 January 2023).
- National Center for Biotechnology Information. PubChem Compound Summary for CID 72291, Ilimaquinone. 2022. Available online: https://pubchem.ncbi.nlm.nih.gov/compound/72291#section=2D-Structure (accessed on 2 January 2023).
- National Center for Biotechnology Information. PubChem Compound Summary for CID 9831636, Jasplakinolide. 2022. Available online: https://pubchem.ncbi.nlm.nih.gov/compound/9831636#section=2D-Structure (accessed on 2 January 2023).
- National Center for Biotechnology Information. PubChem Compound Summary for CID: 6442447, Latrunculins A. 2022. Available online: https://pubchem.ncbi.nlm.nih.gov/compound/6442447#section=2D-Structure (accessed on 2 January 2023).
- National Center for Biotechnology Information. PubChem Compound Summary for CID 6436219, Latrunculin B. 2022. Available online: https://pubchem.ncbi.nlm.nih.gov/compound/6436219#section=2D-Structure (accessed on 2 January 2023).
- National Center for Biotechnology Information. PubChem Compound Summary for CID 9818652, Motuporamine C. 2022. Available online: https://pubchem.ncbi.nlm.nih.gov/compound/9818652#section=2D-Structure (accessed on 2 January 2023).
- National Center for Biotechnology Information. PubChem Compound Summary for CID 73293, Fascaplysin. 2022. Available online: https://pubchem.ncbi.nlm.nih.gov/compound/73293#section=2D-Structure (accessed on 2 January 2023).
- National Center for Biotechnology Information. PubChem Compound Summary for CID: 21777613, Reticulatate. 2022. Available online: https://pubchem.ncbi.nlm.nih.gov/compound/21777613#section=2D-Structure (accessed on 2 January 2023).
- National Center for Biotechnology Information. PubChem Compound Summary for CID: 44233194, Rolloamide B. 2022. Available online: https://pubchem.ncbi.nlm.nih.gov/compound/44233194#section=2D-Structure (accessed on 2 January 2023).
- National Center for Biotechnology Information. PubChem Compound Summary for CID: 25140878, Sintokamide A. 2022. Available online: https://pubchem.ncbi.nlm.nih.gov/compound/25140878#section=2D-Structure (accessed on 2 January 2023).
- National Center for Biotechnology Information. PubChem Compound Summary for CID: 5352083, Stellettin A. 2022. Available online: https://pubchem.ncbi.nlm.nih.gov/compound/5352083#section=2D-Structure (accessed on 2 January 2023).
- National Center for Biotechnology Information. PubChem Compound Summary for CID: 9898671, Kahalalide F. 2022. Available online: https://pubchem.ncbi.nlm.nih.gov/compound/9898671#section=2D-Structure (accessed on 2 January 2023).
- National Center for Biotechnology Information. PubChem Compound Summary for CID: 46832819, Pachycladin A. 2022. Available online: https://pubchem.ncbi.nlm.nih.gov/compound/46832819#section=2D-Structure (accessed on 2 January 2023).
- National Center for Biotechnology Information. PubChem Compound Summary for CID: 102058557, Pseudopterosin H. 2022. Available online: https://pubchem.ncbi.nlm.nih.gov/compound/102058557#section=2D-Structure (accessed on 2 January 2023).
- National Center for Biotechnology Information. PubChem Compound Summary for CID: 21674185, Halityloside A. 2022. Available online: https://pubchem.ncbi.nlm.nih.gov/compound/21674185#section=2D-Structure (accessed on 2 January 2023).
- National Center for Biotechnology Information. PubChem Compound Summary for CID: 13990011, Halityloside B. 2022. Available online: https://pubchem.ncbi.nlm.nih.gov/compound/13990011#section=2D-Structure (accessed on 2 January 2023).
- National Center for Biotechnology Information. PubChem Compound Summary for CID: 505375, Halityloside D. 2022. Available online: https://pubchem.ncbi.nlm.nih.gov/compound/505375#section=2D-Structure (accessed on 2 January 2023).
- Newman, D.; Cragg, G. Marine-Sourced Anti-Cancer and Cancer Pain Control Agents in Clinical and Late Preclinical Development. Mar. Drugs 2014, 12, 255–278. [Google Scholar] [CrossRef] [Green Version]
- Danila, D.C.; Szmulewitz, R.Z.; Vaishampayan, U.; Higano, C.S.; Baron, A.D.; Gilbert, H.N.; Brunstein, F.; Milojic-Blair, M.; Wang, B.; Kabbarah, O.; et al. Phase I Study of DSTP3086S, an Antibody-Drug Conjugate Targeting Six-Transmembrane Epithelial Antigen of Prostate 1, in Metastatic Castration-Resistant Prostate Cancer. JCO 2019, 37, 3518–3527. [Google Scholar] [CrossRef]
- Available online: https://www.midwestern.edu/departments/marinepharmacology/clinical-pipeline (accessed on 8 November 2022).
- Hadaschik, B.A.; Ettinger, S.; Sowery, R.D.; Zoubeidi, A.; Andersen, R.J.; Roberge, M.; Gleave, M.E. Targeting Prostate Cancer with HTI-286, a Synthetic Analog of the Marine Sponge Product Hemiasterlin. Int. J. Cancer 2008, 122, 2368–2376. [Google Scholar] [CrossRef]
- Rui, X.; Pan, H.-F.; Shao, S.-L.; Xu, X.-M. Anti-Tumor and Anti-Angiogenic Effects of Fucoidan on Prostate Cancer: Possible JAK-STAT3 Pathway. BMC Complement. Altern. Med. 2017, 17, 378. [Google Scholar] [CrossRef]
- Choo, G.-S.; Lee, H.-N.; Shin, S.-A.; Kim, H.-J.; Jung, J.-Y. Anticancer Effect of Fucoidan on DU-145 Prostate Cancer Cells through Inhibition of PI3K/Akt and MAPK Pathway Expression. Mar. Drugs 2016, 14, 126. [Google Scholar] [CrossRef] [Green Version]
- Slovin, S.F.; Ragupathi, G.; Fernandez, C.; Jefferson, M.P.; Diani, M.; Wilton, A.S.; Powell, S.; Spassova, M.; Reis, C.; Clausen, H.; et al. A Bivalent Conjugate Vaccine in the Treatment of Biochemically Relapsed Prostate Cancer: A Study of Glycosylated MUC-2-KLH and Globo H-KLH Conjugate Vaccines given with the New Semi-Synthetic Saponin Immunological Adjuvant GPI-0100 OR QS-21. Vaccine 2005, 23, 3114–3122. [Google Scholar] [CrossRef]
- Park, M.-H.; Ill Lee, B.; Byeon, J.-J.; Shin, S.-H.; Choi, J.; Park, J.; Shin, J.G. Pharmacokinetic and Metabolism Studies of Monomethyl Auristatin F via Liquid Chromatography-Quadrupole-Time-of-Flight Mass Spectrometry. Molecules 2019, 24, 2754. [Google Scholar] [CrossRef] [Green Version]
- El-Shorbagi, A.N.; Chaudhary, S.; Chaudhary, A.; Agarwal, G.; Tripathi, P.N.; Dumoga, S.; Aljarad, A.A.; Omer, E.; Mahmoud, F.; Gupta, R.K.; et al. Marine Antineoplastic Templates: Clinical Trials (I–III) and Motifs Carried via Antibodies to Target Specific Cancerous Tissues. Biomed. Pharmacol. J. 2022, 15, 579–603. [Google Scholar] [CrossRef]
- Available online: https://www.genmab.com/pipeline/ (accessed on 8 November 2022).
- Petrylak, D.P.; Vogelzang, N.J.; Chatta, K.; Fleming, M.T.; Smith, D.C.; Appleman, L.J.; Hussain, A.; Modiano, M.; Singh, P.; Tagawa, S.T.; et al. PSMA ADC Monotherapy in Patients with Progressive Metastatic Castration-resistant Prostate Cancer Following Abiraterone and/or Enzalutamide: Efficacy and Safety in Open-label Single-arm Phase 2 Study. Prostate 2020, 80, 99–108. [Google Scholar] [CrossRef]
- Uika Vaishampayan, M.G.; Wei, D.U.; Kraft, A.; Hudes, G.; Wright, J.; Hussain, M. Phase II Study of Dolastatin-10 in Patients with Hormone-Refractory Metastatic Prostate Adenocarcinoma. Clin. Cancer Res. 2000, 4205, 4205–4208. [Google Scholar]
- Nagamine, T.; Kadena, K.; Tomori, M.; Nakajima, K.; Iha, M. Activation of NK Cells in Male Cancer Survivors by Fucoidan Extracted from Cladosiphon Okamuranus. Mol. Clin. Oncol. 2019, 12, 81–88. [Google Scholar] [CrossRef] [Green Version]
- Cragg, G.M.; Pezzuto, J.M. Natural Products as a Vital Source for the Discovery of Cancer Chemotherapeutic and Chemopreventive Agents. Med. Princ. Pract. 2016, 25, 41–59. [Google Scholar] [CrossRef] [PubMed]
- Integration and Application Network. Available online: https://ian.umces.edu/media-library/ (accessed on 17 December 2022).
- National Center for Biotechnology Information. PubChem Compound Summary for CID 9810929, Dolastatin 10. 2022. Available online: https://pubchem.ncbi.nlm.nih.gov/compound/9810929#section=2D-Structure (accessed on 17 December 2022).






| Species | Active Compound | Mechanism of Action | Drug Type | Drug | Cancer Type |
|---|---|---|---|---|---|
| Mollusk/Cyanobacterium | |||||
| Sea Hare (Dolabella auricularia) | Several monomethyl auristatins, derivatives of the original compound, dolastatin | Anti-tubulin agent | Ab-drug conjugate (anti-BCMA Ab conjugated to monomethyl aurostatin F (MMAF) | BLENREPTM (2020) * belantamab mafodotin-blmf | Relapsed/refractory multiple myeloma |
| Ab-drug conjugate (anti-Nectin 4 Ab conjugated to monomethyl auristatin E (MMAE) | PADCEV TM (2019) * enfortumab vedotin | Metastatic urothelial cancer | |||
| Ab-drug conjugate (anti-CD79b Ab conjugated to MMAE) | Polivy TM (2019) * polatuzumab vedotin-piiq | Non-Hodgkin lymphoma, chronic lymphocytic leukaemia, lymphoma, diffuse large B-cell lymphoma | |||
| Ab-drug conjugate (anti-CD30 Ab conjugated to MMAE) | Adcetris® (2011) * brentuximab vedotin | Anaplastic large T-cell systemic malignant lymphoma, Hodgkin’s disease | |||
| Sponges | |||||
| Halichondria okadai | Synthetic derivative from original compound halichondrin B | Anti-tubulin anti-mitotic agent | Antineoplastic chemotherapeutic drug | Halaven® (2010) * eribulin mesylate | Metastatic breast cancer |
| Cryptotethia crypta | Synthetic derivative from original C-nucleoside compounds | DNA alkylating agent | Alkylating chemotherapeutic drug | Cytosar-U® (1969) * cytosine arabinoside (Ara-C) | Leukaemia |
| Tunicates | |||||
| Ecteinascidia turbinata | Derivative of the original compound ecteinascidin (trabectedin) | DNA alkylating agent | Alkylating chemotherapeutic drug | Zepzelca TM (2020) * lurbinectedin | Metastatic Small Cell Lung Cancer |
| Aplidium albicans | Synthetic derivative of original compound ascidian | Inhibitor of eEF1A | Antineoplastic chemotherapeutic drug | Aplidin® ** plitidepsin | Multiple Myeloma, leukaemia, lymphoma |
| Ecteinascidia turbinata | Synthetic derived from original ecteinascidins | DNA alkylating agent | Alkylating chemotherapeutic drug | Yondelis® (2015) * trabectedin | Soft tissue sarcoma and ovarian cancer |
| Organism | Compound/Extract | Active Concentration | Assay Information | Mechanism of Action | Ref. |
|---|---|---|---|---|---|
| Sponges | |||||
| Sponia sp. | Ceylonamide G Ceylonamide F | IC50 of 6.9 µM IC50 of 8.8 µM | MTT assay; DU145 cells | Inhibited cell growth in 2D culture of DU145 | [97] |
| Cliona viridis | Dichloromethane Extract | IC50 of 150 µg/mL | WST 1 assay; PC3 cells | Cytotoxic activity against PC3 cells; inhibits the proliferation of PC3 with in dose-dependent way | [96] |
| Spongiella sp. | 15,16-Dideoxy-15α,17β-Dihydroxy-15,17-Oxidospongian-16-Carboxylate 15,17-Diacetate | IC50 2.51 ± 0.93 µM for PC3 IC50 1.52 ± 0.87 µM for DU145 IC50 1.82 ± 0.85 for LNCaP | 48 h treatment MTT assay; Western blot; Annexin V assay; PCa cells | Cytotoxic effect on PCa Pro-apoptotic effect against PCa | [100] |
| Synthetic (originally from sponges) | FBA-TPQ | PC3 (IC50 of 0.79 μM), LNCaP (IC50 of 0.42 μM and TRAMPC1 (IC50 of 0.26 μM) | MTT assay; Annexin V test; PC3, LNCaP, TRAMPC1 cells | Dose-dependent cytotoxicity on human prostate cancer cells PC3, LNCaP and murine prostate cancer cell TRAMPC1. Induced apoptosis. Inhibited G2/M cell cycle transition | [81] |
| Indonesian marine sponge | Furospinosulin-1 | 1–100 nM. | DU145 cells | Antiproliferative activity on the prostate cancer cell line DU145 in vitro. Suppressed the growth of hypoxia-adapted cancer cells by binding to transcriptional regulators p54(nrb) and LEDGF/p75 | [89] |
| Haliclona gracilis | Gracilosulfate A, Gracilosulfate B, Gracilosulfate D, Gracilosulfate F, Gracilosulfate G | IC50 of 64.4 ± 14.9 µM and IC50 > 100 µM | 24–48 h of treatment; 22Rv1 cells | Inhibited the expression of prostate-specific antigen (PSA) in 22Rv1 human hormone-independent prostate cancer cells | [99] |
| Chondrosia corticata | Halichondramide | IC50 value of 0.81 µM | Sulforhodamine B (SRB) assay; wound-healing test; invasion assays; PC3 cells | Anti-proliferative activities against PC3. Anti-migratory and anti-invasive effects | [83] |
| Pipestela candelabra | Hemiasterlin D | IC50 of 2.20 nM for PC3 and 8.16 nM NFF | PC3 and NFF cells | Cytotoxic activity against PC3 and Inhibiting cells growth. No cytotoxic activity on normal cells NFF | [87] |
| Hippospongia metachromia | Ilimaquinone | - | LNCaP cells | Promoted the hypoxia-inducible factor-1 (HIF-1), induced G1 phase cycle arrest in prostate cancer cells | [85] |
| Jaspis johnstoni | Jasplakinolide | IC50 of 0.3 µM, 0.07 µM, 170 nM respectively | 48 h expos ure; PC3, LNCaP and TSU-Pr1 cells | Reduction of the prostate cancer cell lines PC3, LNCaP and TSU-Pr1 growth and distruption of actin cytoskeleton | [70] |
| Negombata magnofica | Latrunculin A Latrunculin B | 50 nM to 1 µM. | Wound-healing assay; Matrigel assay; PC3M-CT+ cells | Anti-migratory activity against highly metastatic human prostate cancer PC3M-CT+ | [74] |
| Pipestela candelabra | Milnamide E Milnamide F Milnamide G | IC50 of 34.2 nM for PC3 and 123 nM for NFFA IC50 of 2180 nM for PC3 and 5650 for NFF IC50 of 2807 for PC3 and 30% of inhibitions for the NFF at concentration of 10 µM | PC3 and NFF cells | Cytotoxic activity against PC3 and Inhibiting cells growth. No cytotoxic activity on normal cells NFF | [71] |
| Xestospongia exigua | Motuporamine C | - | PC3 cells | Inhibited cell migration in monolayer cultures and inhibited angiogenesis in vitro | [71] |
| Hyrtios sp. | 6-Oxofascaplysin Fascaplysin Reticulatate | IC50 value ranging of 0.54 to 44.9 μM. | LNCaP cells | Cytotoxic activity against the prostate cancer cell line LNCaP | [88] |
| Hemimycale arabica | Phenylmethylene Hydantoin | EC50 of 150 µM | PC3M cells | Anti-invasive and cytotoxic effect against prostate cancer cell line PC3M | [77] |
| Rhizochalina incrustata | Rhizochalinin | 1.8 mg/kg/day. | PC3 and 22Rv1 human tumor xenograft models | Induced caspase-dependent apoptosis, inhibited autophagy, blocks voltage-gated potassium channels. In in vivo experiments, rhizochalinin was administered in PC3 and 22Rv1 human tumor xenograft models at a dose of 1.8 mg/kg/day | [91] |
| Eurypon laughlini | Rolloamide A Rolloamide B | IC50 of 0. 8 µM on LCNaP, 4.7 µM on PC3MM2, 1.4 µM PC3, 0.85 µM DU145 respectively. | MTT assay; LNCaP, PC3MM2, PC3 and DU145 cells | Growth suppression of human prostate cell lines LNCaP, PC3MM2, PC3 and DU145 | [79] |
| Dysidea sp. | Sintokamide A | 5 µg/mL | LNCaP cells | Inhibited the transactivation of N-terminus of the androgen receptor of prostate cancer cell LNCaP | [73] |
| Geodia japonica | Stellettin A | IC50 of 120µg/mL | 48 h treatment; MTT assay; LNCaP cells | Induced cytotoxicity and moderate oxidative stress in the human prostate cancer cell line LNCaP | [72] |
| Spongiella sp. | Spongionellol A | 0.964 ± 0.11 for PC3 0.936 ± 0.39 for DU145 1.02 ± 0.57 for LNCaP | 48 h treatment MTT assay; Western blot; Annexin V assay; PCa cells | Cytotoxic effect on PCa Pro-apoptotic effect against PCa | [100] |
| Psammoclema sp. | Trihydroxysterols Colled Trihydroxysterol 1, Trihydroxysterol 2, Trihydroxysterol 3, Trihydroxysterol 4 | GI50 13 ±1 GI50 27 ±1 GI50 27 ±1 GI50 6.7 ±1 | 72 h of exposure; MTT assay; DU145 cells | Inhibited the cellular growth of human prostate cancer cell DU145 | [80] |
| Mollusks | |||||
| Mollusks | Kahalalide F | PC3 with a IC50 of 0.07 µM DU145 with a IC50 of 0.28 µM | PC3 and DU145 cells. | Induced oncosis, potent cytotoxic activity | [103] |
| Gasteropod Aplysia oculifera | Oculiferane And Epi-Obtusane | IG50 values ranging between 2 and 8 μM | Sulforhodamine-B (SRB) method; PC3 cells | Cytotoxic activity against PC3 cells | [104] |
| Anthozoans | |||||
| Anthopleura anjunae | AAP-H | IC50 of 9.605 mM, 7.910 mM, and 2.298 mM (in vitro) 150 mg/kg/day (in vivo) | 24–72 h of cells treatment; DU145 cells. DU-145 xenografts for in vivo experiments | Cytotoxic effect against DU145 cells. Involved in regulation of the PI3K/AKT/mTOR signaling pathway. Induced the cell S phase arrest, a dose-dependent up-regulation of cleaved-caspase-3 apoptotic protein and a down-regulation of pro-cancer protein p-AKT (Ser473), p-PI3K (p85), and p-mTOR (Ser2448). In in vivo experiments, APP-H reduced tumor weight and slightly increased body weight and quality of life of nude mice. AKT, PI3K, and mTOR levels were reduced in DU-145 xenografts | [107] |
| Leptogorgia sp. | Leptogorgin A Leptogorgin B | - | 72 h of treatment MTT assay; 22Rv1 cells | Moderate cytotoxicity of both compounds against 22Rv1 human drug-resistant prostate cancer cells | [108] |
| Cladiella pachyclados | Pachycladin A Pachycladin D | 50 µM | 24 h treatment; Wound-healing assay; MTT assay; PC3 cells | Anti-migratory and anti-invasive activities against the PC3 cells | [108] |
| Pseudopterogorgia elisabethae | Pseudopterosin H | - | MTT assay; PC3 cells | Anti-neoplastic activity in the PC3 cells | [106] |
| Other Macroorganisms | |||||
| Starfish Culcita novaeguineae | Halityloside A, Halityloside B, Halityloside D Culcitoside C5 | LNCaP with IC50 = 48.59 ± 2.30 mM, IC50 = 39.6 ± 82.65; IC50 = 31.80 ± 1.59, IC50 = 57.0 ± 81.81 | sulforhodamine B (SRB) assay; LNCaP | Moderate cytotoxicity against the human prostate cell line LNCaP | [110] |
| Salmon | Sph | 160 µg/mL | LCNaP and PC3 cells | Anticancer effect on LCNaP and PC3 cells Decreased LNCaP cells’ relative colony survival from 25% to 2% | [111] |
| Pacific cod | Tfd100 | - | PC3 cells. PC3-induced metastasis in mice | Inhibited in vitro adhesion of androgen-independent prostate cancer cells PC3 to endothelial cells, angiogenesis, and gal3-mediated T-cell apoptosis and also prevented PC3-induced metastasis in mice | [109] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Montuori, E.; Hyde, C.A.C.; Crea, F.; Golding, J.; Lauritano, C. Marine Natural Products with Activities against Prostate Cancer: Recent Discoveries. Int. J. Mol. Sci. 2023, 24, 1435. https://doi.org/10.3390/ijms24021435
Montuori E, Hyde CAC, Crea F, Golding J, Lauritano C. Marine Natural Products with Activities against Prostate Cancer: Recent Discoveries. International Journal of Molecular Sciences. 2023; 24(2):1435. https://doi.org/10.3390/ijms24021435
Chicago/Turabian StyleMontuori, Eleonora, Caroline A. C. Hyde, Francesco Crea, Jon Golding, and Chiara Lauritano. 2023. "Marine Natural Products with Activities against Prostate Cancer: Recent Discoveries" International Journal of Molecular Sciences 24, no. 2: 1435. https://doi.org/10.3390/ijms24021435