
Increased Hepatic ATG7 mRNA and ATG7 Protein Expression in Nonalcoholic Steatohepatitis Associated with Obesity

 ,
,  , ,
, ,
Abstract
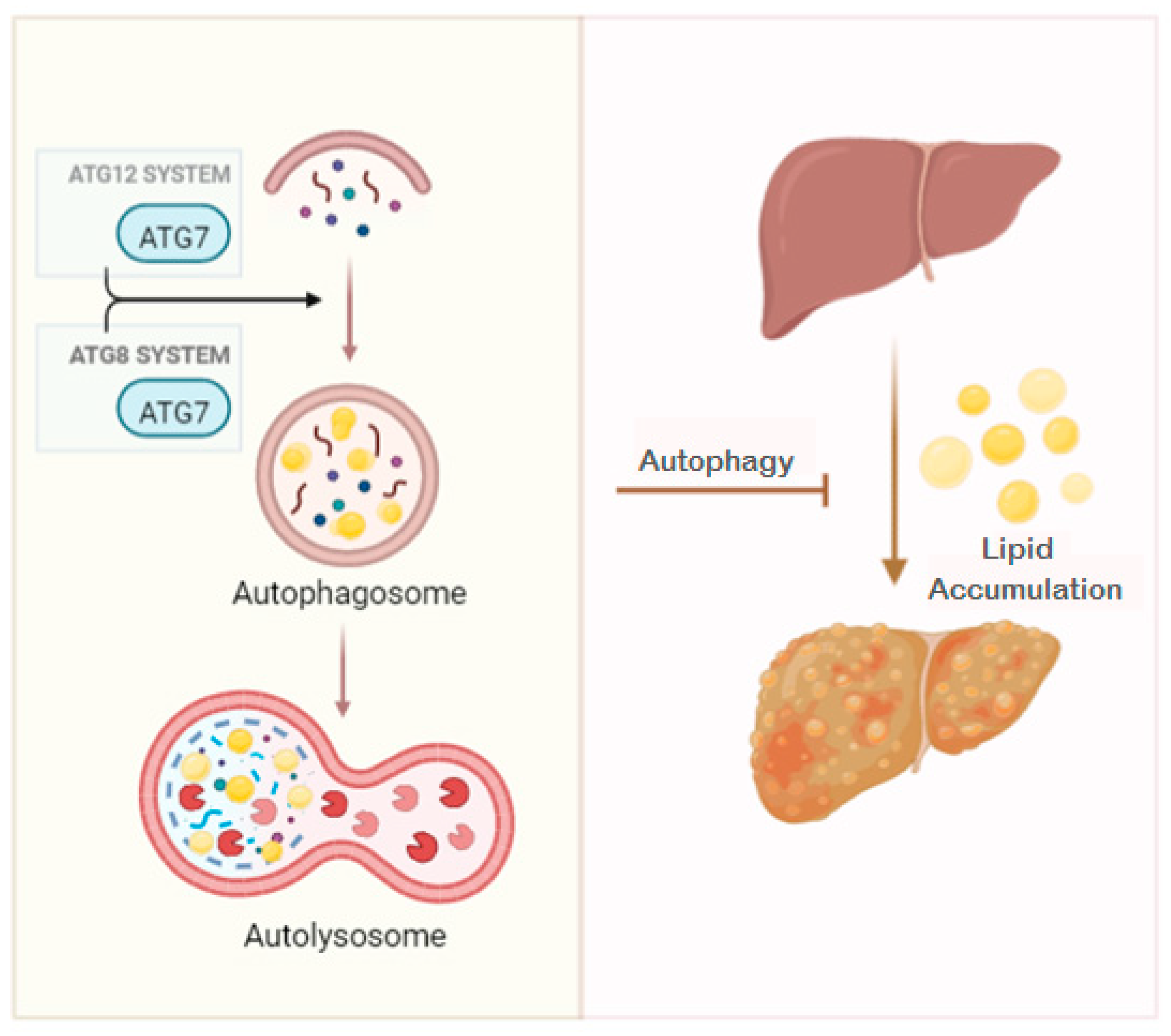
:1. Introduction
2. Results
2.1. Baseline Characteristics of Subjects
2.2. Evaluation of the Relative mRNA and Protein Expressions of Hepatic ATG7 According to BMI
2.3. Evaluation of the Relative mRNA and Protein Expressions of Hepatic ATG7 According to Liver Histology
2.4. Evaluation of the Relative mRNA and Protein Expressions of Hepatic ATG7 According to NASH-Related Parameters
2.5. Evaluation of the Relative mRNA and Protein Expressions of ATG7 According to NAFLD Comorbidities
2.6. Correlations between Relative mRNA and Protein Expressions of Hepatic ATG7, with Clinical and Biochemical-Related Parameters
3. Discussion
4. Material and Methods
4.1. Participants
4.2. Hepatopathological Diagnosis
4.3. Biochemical Analysis
4.4. Hepatic mRNA Expression
4.5. Western Blot Analysis
4.6. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Younossi, Z.M.; Koenig, A.B.; Abdelatif, D.; Fazel, Y.; Henry, L.; Wymer, M. Global Epidemiology of Nonalcoholic Fatty Liver Disease-Meta-Analytic Assessment of Prevalence, Incidence, and Outcomes. Hepatology 2016, 64, 73–84. [Google Scholar] [CrossRef] [Green Version]
- Chalasani, N.; Younossi, Z.; Lavine, J.E.; Charlton, M.; Cusi, K.; Rinella, M.; Harrison, S.A.; Brunt, E.M.; Sanyal, A.J. The Diagnosis and Management of Nonalcoholic Fatty Liver Disease: Practice Guidance from the American Association for the Study of Liver Diseases. Hepatology 2018, 67, 328–357. [Google Scholar] [CrossRef] [Green Version]
- Younossi, Z.; Anstee, Q.M.; Marietti, M.; Hardy, T.; Henry, L.; Eslam, M.; George, J.; Bugianesi, E. Global Burden of NAFLD and NASH: Trends, Predictions, Risk Factors and Prevention. Nat. Rev. Gastroenterol. Hepatol. 2018, 15, 11–20. [Google Scholar] [CrossRef]
- Dai, W.; Ye, L.; Liu, A.; Wen, S.W.; Deng, J.; Wu, X.; Lai, Z. Prevalence of Nonalcoholic Fatty Liver Disease in Patients with Type 2 Diabetes Mellitus: A Meta-Analysis. Medicine 2017, 96, e8179. [Google Scholar] [CrossRef]
- Goldberg, D.; Ditah, I.C.; Saeian, K.; Lalehzari, M.; Aronsohn, A.; Gorospe, E.C.; Charlton, M. Changes in the Prevalence of Hepatitis C Virus Infection, Nonalcoholic Steatohepatitis, and Alcoholic Liver Disease Among Patients With Cirrhosis or Liver Failure on the Waitlist for Liver Transplantation. Gastroenterology 2017, 152, 1090–1099.e1. [Google Scholar] [CrossRef] [Green Version]
- Mittal, S.; El-Serag, H.B.; Sada, Y.H.; Kanwal, F.; Duan, Z.; Temple, S.; May, S.B.; Kramer, J.R.; Richardson, P.A.; Davila, J.A. Hepatocellular Carcinoma in the Absence of Cirrhosis in United States Veterans Is Associated With Nonalcoholic Fatty Liver Disease. Clin. Gastroenterol. Hepatol. 2016, 14, 124–131.e1. [Google Scholar] [CrossRef] [Green Version]
- Maya-Miles, D.; Ampuero, J.; Gallego-Durán, R.; Dingianna, P.; Romero-Gómez, M. Management of NAFLD Patients with Advanced Fibrosis. Liver Int. 2021, 41 (Suppl. 1), 95–104. [Google Scholar] [CrossRef]
- Xu, Z.; Hu, W.; Wang, B.; Xu, T.; Wang, J.; Wei, D. Canagliflozin Ameliorates Nonalcoholic Fatty Liver Disease by Regulating Lipid Metabolism and Inhibiting Inflammation through Induction of Autophagy. Yonsei Med. J. 2022, 63, 619. [Google Scholar] [CrossRef]
- Duarte, N.; Coelho, I.C.; Patarrão, R.S.; Almeida, J.I.; Penha-Gonçalves, C.; Macedo, M.P. How Inflammation Impinges on NAFLD: A Role for Kupffer Cells. BioMed Res. Int. 2015, 2015, 984578. [Google Scholar] [CrossRef] [Green Version]
- Kazankov, K.; Jørgensen, S.M.D.; Thomsen, K.L.; Møller, H.J.; Vilstrup, H.; George, J.; Schuppan, D.; Grønbæk, H. The Role of Macrophages in Nonalcoholic Fatty Liver Disease and Nonalcoholic Steatohepatitis. Nat. Rev. Gastroenterol. Hepatol. 2019, 16, 145–159. [Google Scholar] [CrossRef]
- Luo, X.; Luo, S.-Z.; Xu, Z.-X.; Zhou, C.; Li, Z.-H.; Zhou, X.-Y.; Xu, M.-Y. Lipotoxic Hepatocyte-Derived Exosomal MiR-1297 Promotes Hepatic Stellate Cell Activation through the PTEN Signaling Pathway in Metabolic-Associated Fatty Liver Disease. World J. Gastroenterol. 2021, 27, 1419–1434. [Google Scholar] [CrossRef] [PubMed]
- Fianchi, F.; Liguori, A.; Gasbarrini, A.; Grieco, A.; Miele, L. Nonalcoholic Fatty Liver Disease (NAFLD) as Model of Gut-Liver Axis Interaction: From Pathophysiology to Potential Target of Treatment for Personalized Therapy. Int. J. Mol. Sci. 2021, 22, 6485. [Google Scholar] [CrossRef] [PubMed]
- Wynn, T.A.; Barron, L. Macrophages: Master Regulators of Inflammation and Fibrosis. Semin. Liver Dis. 2010, 30, 245–257. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, J.; Lee, S.; Lee, M.-S. Suppressive Effect of Autocrine FGF21 on Autophagy-Deficient Hepatic Tumorigenesis. Front. Oncol. 2022, 12, 832804. [Google Scholar] [CrossRef] [PubMed]
- Klionsky, D.J.; Petroni, G.; Amaravadi, R.K.; Baehrecke, E.H.; Ballabio, A.; Boya, P.; Bravo-San Pedro, J.M.; Cadwell, K.; Cecconi, F.; Choi, A.M.K.; et al. Autophagy in Major Human Diseases. EMBO J. 2021, 40, e108863. [Google Scholar] [CrossRef] [PubMed]
- Singh, R.; Kaushik, S.; Wang, Y.; Xiang, Y.; Novak, I.; Komatsu, M.; Tanaka, K.; Cuervo, A.M.; Czaja, M.J. Autophagy Regulates Lipid Metabolism. Nature 2009, 458, 1131–1135. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- González-Rodríguez, A.; Mayoral, R.; Agra, N.; Valdecantos, M.P.; Pardo, V.; Miquilena-Colina, M.E.; Vargas-Castrillón, J.; Lo Iacono, O.; Corazzari, M.; Fimia, G.M.; et al. Impaired Autophagic Flux Is Associated with Increased Endoplasmic Reticulum Stress during the Development of NAFLD. Cell Death Dis. 2014, 5, e1179. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tanaka, S.; Hikita, H.; Tatsumi, T.; Sakamori, R.; Nozaki, Y.; Sakane, S.; Shiode, Y.; Nakabori, T.; Saito, Y.; Hiramatsu, N.; et al. Rubicon Inhibits Autophagy and Accelerates Hepatocyte Apoptosis and Lipid Accumulation in Nonalcoholic Fatty Liver Disease in Mice. Hepatology 2016, 64, 1994–2014. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weiskirchen, R.; Tacke, F. Relevance of Autophagy in Parenchymal and Non-Parenchymal Liver Cells for Health and Disease. Cells 2019, 8, 16. [Google Scholar] [CrossRef] [Green Version]
- Zhang, P.; Cheng, X.; Sun, H.; Li, Y.; Mei, W.; Zeng, C. Atractyloside Protect Mice Against Liver Steatosis by Activation of Autophagy via ANT-AMPK-MTORC1 Signaling Pathway. Front. Pharmacol. 2021, 12, 736655. [Google Scholar] [CrossRef]
- Bruinstroop, E.; Dalan, R.; Cao, Y.; Bee, Y.M.; Chandran, K.; Cho, L.W.; Soh, S.B.; Teo, E.K.; Toh, S.-A.; Leow, M.K.S.; et al. Low-Dose Levothyroxine Reduces Intrahepatic Lipid Content in Patients With Type 2 Diabetes Mellitus and NAFLD. J. Clin. Endocrinol. Metab. 2018, 103, 2698–2706. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.; Tripathi, M.; Ho, J.P.; Widjaja, A.A.; Shekeran, S.G.; Camat, M.D.; James, A.; Wu, Y.; Ching, J.; Kovalik, J.-P.; et al. Thyroid Hormone Decreases Hepatic Steatosis, Inflammation, and Fibrosis in a Dietary Mouse Model of Nonalcoholic Steatohepatitis. Thyroid 2022, 32, 725–738. [Google Scholar] [CrossRef] [PubMed]
- Oh, H.-A.; Kwon, S.; Choi, S.; Shin, H.; Yoon, K.H.; Kim, W.J.; Lim, H.J. Uncovering a Role for Endocannabinoid Signaling in Autophagy in Preimplantation Mouse Embryos. MHR Mol. Hum. Reprod. 2013, 19, 93–101. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.-L.; Lin, Y.-C. Autophagy Dysregulation in Metabolic Associated Fatty Liver Disease: A New Therapeutic Target. Int. J. Mol. Sci. 2022, 23, 10055. [Google Scholar] [CrossRef] [PubMed]
- Sukseree, S.; Schwarze, U.Y.; Gruber, R.; Gruber, F.; Quiles Del Rey, M.; Mancias, J.D.; Bartlett, J.D.; Tschachler, E.; Eckhart, L. ATG7 Is Essential for Secretion of Iron from Ameloblasts and Normal Growth of Murine Incisors during Aging. Autophagy 2020, 16, 1851–1857. [Google Scholar] [CrossRef] [PubMed]
- Baselli, G.A.; Jamialahmadi, O.; Pelusi, S.; Ciociola, E.; Malvestiti, F.; Saracino, M.; Santoro, L.; Cherubini, A.; Dongiovanni, P.; Maggioni, M.; et al. Rare ATG7 Genetic Variants Predispose Patients to Severe Fatty Liver Disease. J. Hepatol. 2022, 77, 596–606. [Google Scholar] [CrossRef]
- Kim, K.H.; Jeong, Y.T.; Oh, H.; Kim, S.H.; Cho, J.M.; Kim, Y.-N.; Kim, S.S.; Kim, D.H.; Hur, K.Y.; Kim, H.K.; et al. Autophagy Deficiency Leads to Protection from Obesity and Insulin Resistance by Inducing Fgf21 as a Mitokine. Nat. Med. 2013, 19, 83–92. [Google Scholar] [CrossRef] [Green Version]
- Soussi, H.; Reggio, S.; Alili, R.; Prado, C.; Mutel, S.; Pini, M.; Rouault, C.; Clément, K.; Dugail, I. DAPK2 Downregulation Associates With Attenuated Adipocyte Autophagic Clearance in Human Obesity. Diabetes 2015, 64, 3452–3463. [Google Scholar] [CrossRef] [Green Version]
- Xu, Q.; Mariman, E.C.M.; Roumans, N.J.T.; Vink, R.G.; Goossens, G.H.; Blaak, E.E.; Jocken, J.W.E. Adipose Tissue Autophagy Related Gene Expression Is Associated with Glucometabolic Status in Human Obesity. Adipocyte 2018, 7, 12–19. [Google Scholar] [CrossRef]
- Dong, H.; Czaja, M.J. Regulation of Lipid Droplets by Autophagy. Trends Endocrinol. Metab. 2011, 22, 234–240. [Google Scholar] [CrossRef]
- Ohsaki, Y.; Cheng, J.; Fujita, A.; Tokumoto, T.; Fujimoto, T. Cytoplasmic Lipid Droplets Are Sites of Convergence of Proteasomal and Autophagic Degradation of Apolipoprotein B. Mol. Biol. Cell 2006, 17, 2674–2683. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harada, M.; Hanada, S.; Toivola, D.M.; Ghori, N.; Omary, M.B. Autophagy Activation by Rapamycin Eliminates Mouse Mallory-Denk Bodies and Blocks Their Proteasome Inhibitor-Mediated Formation. Hepatology 2008, 47, 2026–2035. [Google Scholar] [CrossRef] [PubMed]
- Serafim, M.P.; Santo, M.A.; Gadducci, A.V.; Scabim, V.M.; Cecconello, I.; de Cleva, R. Very Low-Calorie Diet in Candidates for Bariatric Surgery: Change in Body Composition during Rapid Weight Loss. Clinics 2019, 74, e560. [Google Scholar] [CrossRef]
- Yamamoto, T.; Takabatake, Y.; Takahashi, A.; Kimura, T.; Namba, T.; Matsuda, J.; Minami, S.; Kaimori, J.; Matsui, I.; Matsusaka, T.; et al. High-Fat Diet–Induced Lysosomal Dysfunction and Impaired Autophagic Flux Contribute to Lipotoxicity in the Kidney. J. Am. Soc. Nephrol. 2017, 28, 1534–1551. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Byrnes, K.; Blessinger, S.; Bailey, N.T.; Scaife, R.; Liu, G.; Khambu, B. Therapeutic Regulation of Autophagy in Hepatic Metabolism. Acta Pharm. Sin. B 2022, 12, 33–49. [Google Scholar] [CrossRef]
- Camargo, A.; Rangel-Zúñiga, O.A.; Alcalá-Díaz, J.; Gomez-Delgado, F.; Delgado-Lista, J.; García-Carpintero, S.; Marín, C.; Almadén, Y.; Yubero-Serrano, E.M.; López-Moreno, J.; et al. Dietary Fat May Modulate Adipose Tissue Homeostasis through the Processes of Autophagy and Apoptosis. Eur. J. Nutr. 2017, 56, 1621–1628. [Google Scholar] [CrossRef]
- Hadavi, M.; Najdegerami, E.H.; Nikoo, M.; Nejati, V. Protective Effect of Protein Hydrolysates from Litopenaeus Vannamei Waste on Oxidative Status, Glucose Regulation, and Autophagy Genes in Non-Alcoholic Fatty Liver Disease in Wistar Rats. Iran J. Basic Med. Sci. 2022, 25, 954–963. [Google Scholar] [CrossRef]
- Rutsatz, K.; Sobkowiak, E.M.; Bienengräber, V.; Held, M. [Cytological changes in the pulp-dentin system following injury due to cavity preparation and the application of phosphate cement]. Zahn Mund Kieferheilkd Zent. 1985, 73, 123–131. [Google Scholar]
- Zhong, Z.; Sanchez-Lopez, E.; Karin, M. Autophagy, Inflammation, and Immunity: A Troika Governing Cancer and Its Treatment. Cell 2016, 166, 288–298. [Google Scholar] [CrossRef] [Green Version]
- Hammoutene, A.; Biquard, L.; Lasselin, J.; Kheloufi, M.; Tanguy, M.; Vion, A.-C.; Mérian, J.; Colnot, N.; Loyer, X.; Tedgui, A.; et al. A Defect in Endothelial Autophagy Occurs in Patients with Non-Alcoholic Steatohepatitis and Promotes Inflammation and Fibrosis. J. Hepatol 2020, 72, 528–538. [Google Scholar] [CrossRef]
- Caballería, L.; Auladell, M.A.; Torán, P.; Miranda, D.; Aznar, J.; Pera, G.; Gil, D.; Muñoz, L.; Planas, J.; Canut, S.; et al. Prevalence and Factors Associated with the Presence of Non Alcoholic Fatty Liver Disease in an Apparently Healthy Adult Population in Primary Care Units. BMC Gastroenterol. 2007, 7, 41. [Google Scholar] [CrossRef] [PubMed]
- Deprince, A.; Haas, J.T.; Staels, B. Dysregulated Lipid Metabolism Links NAFLD to Cardiovascular Disease. Mol. Metab. 2020, 42, 101092. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Li, P.; Fu, S.; Calay, E.S.; Hotamisligil, G.S. Defective Hepatic Autophagy in Obesity Promotes ER Stress and Causes Insulin Resistance. Cell Metab. 2010, 11, 467–478. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martinez-Lopez, N.; Singh, R. Autophagy and Lipid Droplets in the Liver. Annu. Rev. Nutr. 2015, 35, 215–237. [Google Scholar] [CrossRef]
- Parlati, L.; Régnier, M.; Guillou, H.; Postic, C. New Targets for NAFLD. JHEP Rep. 2021, 3, 100346. [Google Scholar] [CrossRef]
- Schlaepfer, I.R.; Joshi, M. CPT1A-Mediated Fat Oxidation, Mechanisms, and Therapeutic Potential. Endocrinology 2020, 161, bqz046. [Google Scholar] [CrossRef]
- Steinhoff, J.S.; Lass, A.; Schupp, M. Biological Functions of RBP4 and Its Relevance for Human Diseases. Front. Physiol. 2021, 12, 659977. [Google Scholar] [CrossRef]
- Joseph, S.B.; Castrillo, A.; Laffitte, B.A.; Mangelsdorf, D.J.; Tontonoz, P. Reciprocal Regulation of Inflammation and Lipid Metabolism by Liver X Receptors. Nat. Med. 2003, 9, 213–219. [Google Scholar] [CrossRef]
- Liu, Y.; Qiu, D.K.; Ma, X. Liver X Receptors Bridge Hepatic Lipid Metabolism and Inflammation: LXR, Hepatic Lipogenesis & Inflammation. J. Dig. Dis. 2012, 13, 69–74. [Google Scholar] [CrossRef]
- Janowski, B.A.; Willy, P.J.; Devi, T.R.; Falck, J.R.; Mangelsdorf, D.J. An Oxysterol Signalling Pathway Mediated by the Nuclear Receptor LXRα. Nature 1996, 383, 728–731. [Google Scholar] [CrossRef]
- Kim, Y.S.; Nam, H.J.; Han, C.Y.; Joo, M.S.; Jang, K.; Jun, D.W.; Kim, S.G. Liver X Receptor Alpha Activation Inhibits Autophagy and Lipophagy in Hepatocytes by Dysregulating Autophagy-Related 4B Cysteine Peptidase and Rab-8B, Reducing Mitochondrial Fuel Oxidation. Hepatology 2021, 73, 1307–1326. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, S.; Sou, Y.-S.; Saito, T.; Kuma, A.; Yabe, T.; Sugiura, Y.; Lee, H.-C.; Suematsu, M.; Yokomizo, T.; Koike, M.; et al. Loss of Autophagy Impairs Physiological Steatosis by Accumulation of NCoR1. Life Sci. Alliance 2020, 3, e201900513. [Google Scholar] [CrossRef] [Green Version]
- Weber, M.; Mera, P.; Casas, J.; Salvador, J.; Rodríguez, A.; Alonso, S.; Sebastián, D.; Soler-Vázquez, M.C.; Montironi, C.; Recalde, S.; et al. Liver CPT1A Gene Therapy Reduces Diet-induced Hepatic Steatosis in Mice and Highlights Potential Lipid Biomarkers for Human NAFLD. FASEB J. 2020, 34, 11816–11837. [Google Scholar] [CrossRef] [PubMed]
- Moore, M.P.; Cunningham, R.P.; Meers, G.M.; Johnson, S.A.; Wheeler, A.A.; Ganga, R.R.; Spencer, N.M.; Pitt, J.B.; Diaz-Arias, A.; Swi, A.I.A.; et al. Compromised Hepatic Mitochondrial Fatty Acid Oxidation and Reduced Markers of Mitochondrial Turnover in Human NAFLD. Hepatology 2022, 76, 1452–1465. [Google Scholar] [CrossRef] [PubMed]
- Yanagitani, A.; Yamada, S.; Yasui, S.; Shimomura, T.; Murai, R.; Murawaki, Y.; Hashiguchi, K.; Kanbe, T.; Saeki, T.; Ichiba, M.; et al. Retinoic Acid Receptor Alpha Dominant Negative Form Causes Steatohepatitis and Liver Tumors in Transgenic Mice. Hepatology 2004, 40, 366–375. [Google Scholar] [CrossRef]
- Blaner, W.S. Vitamin A Signaling and Homeostasis in Obesity, Diabetes, and Metabolic Disorders. Pharmacol. Ther. 2019, 197, 153–178. [Google Scholar] [CrossRef] [PubMed]
- Von Eynatten, M.; Humpert, P.M. Retinol-Binding Protein-4 in Experimental and Clinical Metabolic Disease. Expert Rev. Mol. Diagn. 2008, 8, 289–299. [Google Scholar] [CrossRef]
- Giashuddin, S.; Alawad, M. Histopathological Diagnosis of Nonalcoholic Steatohepatitis (NASH). Methods Mol. Biol. 2022, 2455, 1–18. [Google Scholar] [CrossRef]
- Auguet, T.; Berlanga, A.; Guiu-Jurado, E.; Terra, X.; Martinez, S.; Aguilar, C.; Filiu, E.; Alibalic, A.; Sabench, F.; Hernández, M.; et al. Endocannabinoid Receptors Gene Expression in Morbidly Obese Women with Nonalcoholic Fatty Liver Disease. BioMed Res. Int. 2014, 2014, 502542. [Google Scholar] [CrossRef] [Green Version]
- Julien, B.; Grenard, P.; Teixeira-Clerc, F.; Van Nhieu, J.T.; Li, L.; Karsak, M.; Zimmer, A.; Mallat, A.; Lotersztajn, S. Antifibrogenic Role of the Cannabinoid Receptor CB2 in the Liver. Gastroenterology 2005, 128, 742–755. [Google Scholar] [CrossRef]
- Bátkai, S.; Osei-Hyiaman, D.; Pan, H.; El-Assal, O.; Rajesh, M.; Mukhopadhyay, P.; Hong, F.; Harvey-White, J.; Jafri, A.; Haskó, G.; et al. Cannabinoid-2 Receptor Mediates Protection against Hepatic Ischemia/Reperfusion Injury. FASEB J. 2007, 21, 1788–1800. [Google Scholar] [CrossRef] [PubMed]
- Lotersztajn, S.; Teixeira-Clerc, F.; Julien, B.; Deveaux, V.; Ichigotani, Y.; Manin, S.; Tran-Van-Nhieu, J.; Karsak, M.; Zimmer, A.; Mallat, A. CB2 Receptors as New Therapeutic Targets for Liver Diseases. Br. J. Pharmacol. 2008, 153, 286–289. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Osei-Hyiaman, D.; Liu, J.; Zhou, L.; Godlewski, G.; Harvey-White, J.; Jeong, W.; Bátkai, S.; Marsicano, G.; Lutz, B.; Buettner, C.; et al. Hepatic CB1 Receptor Is Required for Development of Diet-Induced Steatosis, Dyslipidemia, and Insulin and Leptin Resistance in Mice. J. Clin. Investig. 2008, 118, 3160–3169. [Google Scholar] [CrossRef]
- Shabalala, S.C.; Dludla, P.V.; Mabasa, L.; Kappo, A.P.; Basson, A.K.; Pheiffer, C.; Johnson, R. The Effect of Adiponectin in the Pathogenesis of Non-Alcoholic Fatty Liver Disease (NAFLD) and the Potential Role of Polyphenols in the Modulation of Adiponectin Signaling. Biomed. Pharmacother. 2020, 131, 110785. [Google Scholar] [CrossRef] [PubMed]
- Shehzad, A.; Iqbal, W.; Shehzad, O.; Lee, Y.S. Adiponectin: Regulation of Its Production and Its Role in Human Diseases. Hormones 2012, 11, 8–20. [Google Scholar] [CrossRef] [PubMed]
- Kleiner, D.E.; Brunt, E.M.; Van Natta, M.; Behling, C.; Contos, M.J.; Cummings, O.W.; Ferrell, L.D.; Liu, Y.-C.; Torbenson, M.S.; Unalp-Arida, A.; et al. Design and Validation of a Histological Scoring System for Nonalcoholic Fatty Liver Disease. Hepatology 2005, 41, 1313–1321. [Google Scholar] [CrossRef] [PubMed]
- Brunt, E.M.; Janney, C.G.; Di Bisceglie, A.M.; Neuschwander-Tetri, B.A.; Bacon, B.R. Nonalcoholic Steatohepatitis: A Proposal for Grading and Staging The Histological Lesions. Am. J. Gastroenterol. 1999, 94, 2467–2474. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| MO (n = 72) | ||||
|---|---|---|---|---|
| Variables | NW (n = 6) | NL (n = 11) | SS (n = 29) | NASH (n = 32) |
| Weight (kg) | 63.00 (58.00–69.00) | 121.50 (107.00–130.00) * | 124.00 (112.00–136.00) * | 120.00 (109.90–137.20) * |
| BMI (kg/m2) | 22.94 (19.93–25.00) | 44.46 (49.56–42.60) * | 46.36 (43.58–50.95) * | 46.28 (43.26–51.04) * |
| SBP (mmHg) | 110.00 (108.00–128.00) | 125.00 (109.75–138.25) | 130.00 (117.00–141.25) | 130.00 (119.00–144.00) * $ |
| DBP (mmHg) | 70.00 (67.50–72.00) | 68.00 (58.00–78.00) | 70.00 (61.00–80.00) | 73.00 (63.50–86.00) $ # |
| HOMA1-IR | 1.50 (0.99–2.23) | 2.96 (1.92–6.46) | 4.28 (2.83–6.88) * | 3.95 (2.58–10.70) * |
| Glucose (mg/dL) | 84.5 (72.6–96.12) | 97.11 (86.84–116.38) | 108.10 (92.60–139.08) | 132.06 (102.51–187.73) *$ |
| Insulin (mUI/L) | 6.41 (4.67–9.56) | 13.01 (8.60–25.13) * | 14.93 (10.25–27.23) * | 18.91 (8.46–52.87) * |
| HbA1c (%) | 4.45 (4.30–4.90) | 5.20 (4.75–5.63) | 5.60 (5.00–6.15) | 5.30 (5.00–6.55) |
| TG (mg/dL) | 123.50 (74.89–245.25) | 136.00 (104.50–177.75) | 188.00 (144.75–238.50) $ | 153.00 (119.50–197.50) |
| Cholesterol (mg/dL) | 181.00 (134.18–197.60) | 178.80 (145.45–195.75) | 171.35 (154.85–189.55) | 183.80 (153.50–203.00) |
| HDL-C (mg/dL) | 40.50 (35.75–45.75) | 39.50 (31.90–47.50) | 37.10 (33.88–47.00) | 40.00 (33.00–42.00) |
| LDL-C (mg/dL) | 90.45 (76.48–129.40) | 103.50 (86.28–123.75) | 100.50 (79.15–124.55) | 93.50 (83.50–128.60) |
| AST (UI/L) | 27.50 (21.75–46.00) | 36.00 (21.00–45.00) | 26.00 (21.50–37.00) | 40.50 (23.75–57.25) |
| ALT (UI/L) | 19.00 (16.00–70.00) | 33.00 (21.50–49.50) | 29.00 (21.00–41.00) | 34.00 (24.25–67.25) |
| GGT (UI/L) | 37.00 (12.00–131.00) | 19.00 (14.00–29.50) | 22.50 (17.00–33.25) | 26.00 (15.00–68.40) |
| ALP (Ul/L) | 72.00 (68.50–112.50) | 63.50 (50.00–73.50) | 66.50 (54.00–76.00) | 64.00 (55.00–77.00 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Barrientos-Riosalido, A.; Real, M.; Bertran, L.; Aguilar, C.; Martínez, S.; Parada, D.; Vives, M.; Sabench, F.; Riesco, D.; Castillo, D.D.; et al. Increased Hepatic ATG7 mRNA and ATG7 Protein Expression in Nonalcoholic Steatohepatitis Associated with Obesity. Int. J. Mol. Sci. 2023, 24, 1324. https://doi.org/10.3390/ijms24021324
Barrientos-Riosalido A, Real M, Bertran L, Aguilar C, Martínez S, Parada D, Vives M, Sabench F, Riesco D, Castillo DD, et al. Increased Hepatic ATG7 mRNA and ATG7 Protein Expression in Nonalcoholic Steatohepatitis Associated with Obesity. International Journal of Molecular Sciences. 2023; 24(2):1324. https://doi.org/10.3390/ijms24021324
Chicago/Turabian StyleBarrientos-Riosalido, Andrea, Monica Real, Laia Bertran, Carmen Aguilar, Salomé Martínez, David Parada, Margarita Vives, Fàtima Sabench, David Riesco, Daniel Del Castillo, and et al. 2023. "Increased Hepatic ATG7 mRNA and ATG7 Protein Expression in Nonalcoholic Steatohepatitis Associated with Obesity" International Journal of Molecular Sciences 24, no. 2: 1324. https://doi.org/10.3390/ijms24021324