
Pterostilbene-Mediated Inhibition of Cell Proliferation and Cell Death Induction in Amelanotic and Melanotic Melanoma

Abstract
:1. Introduction
2. Results
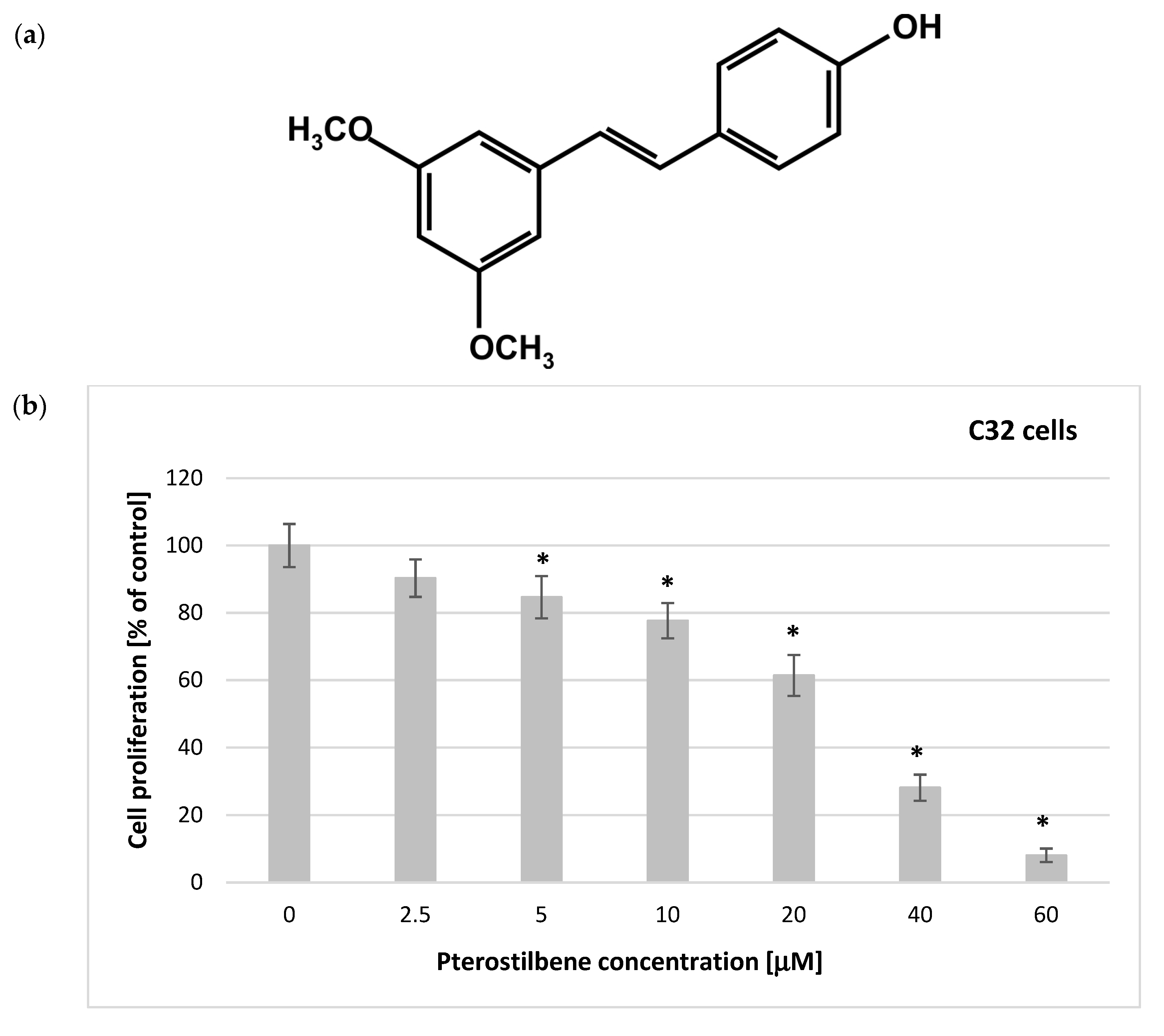
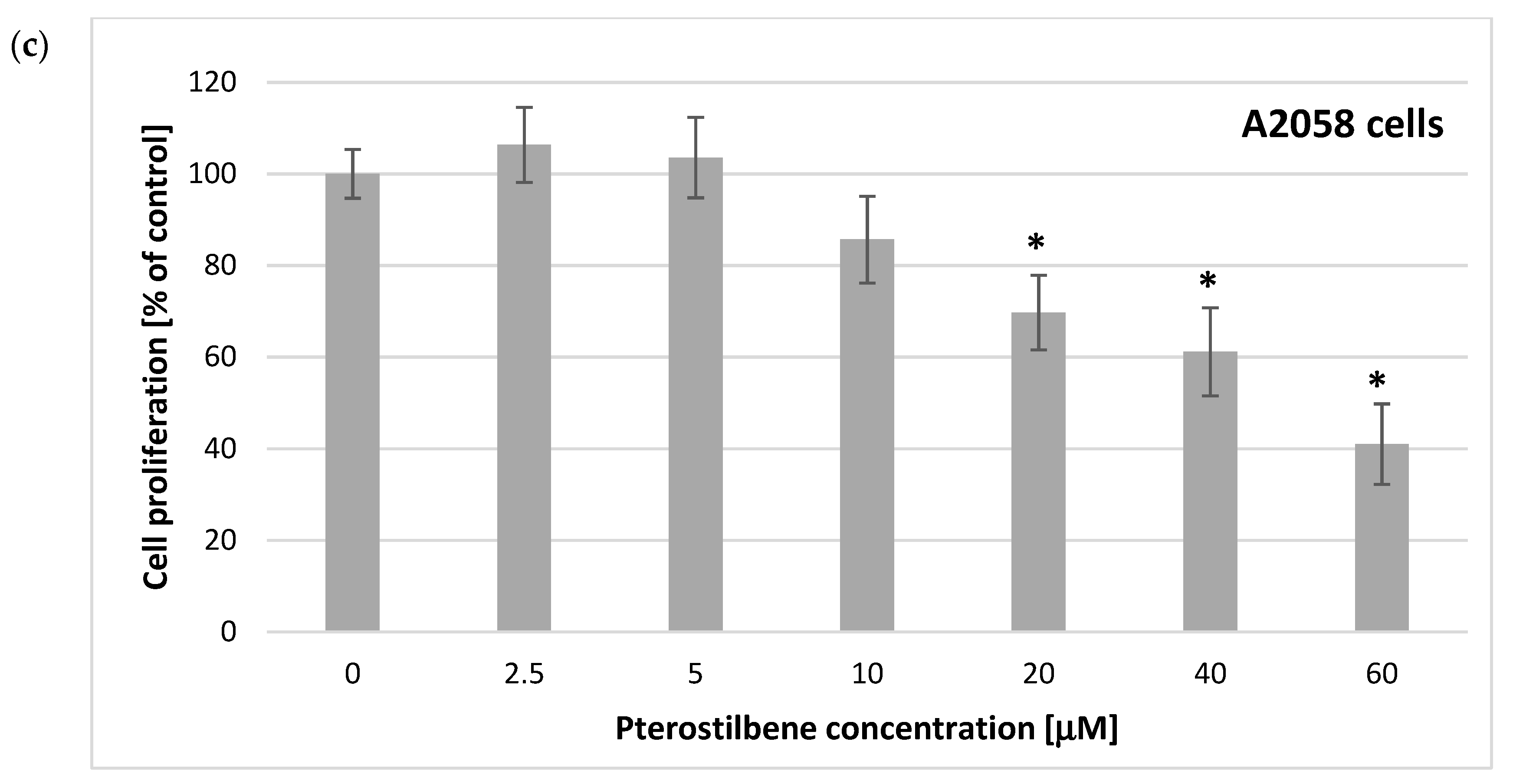
2.1. Antiproliferation Activity of Pterostilbene on C32 and A2058 Melanoma Cells
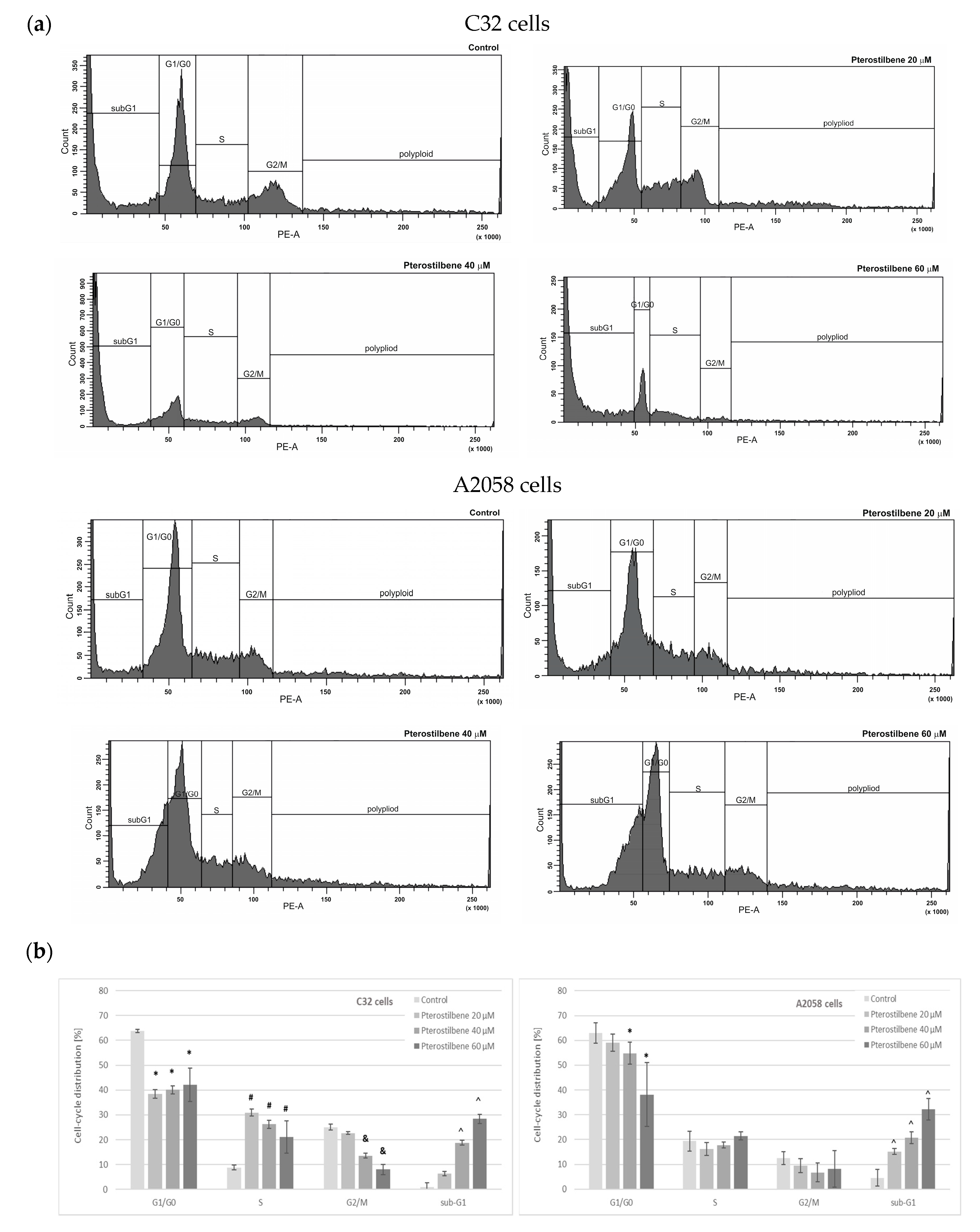
2.2. The Influence of Pterostilbene on Melanoma Cell Cycle
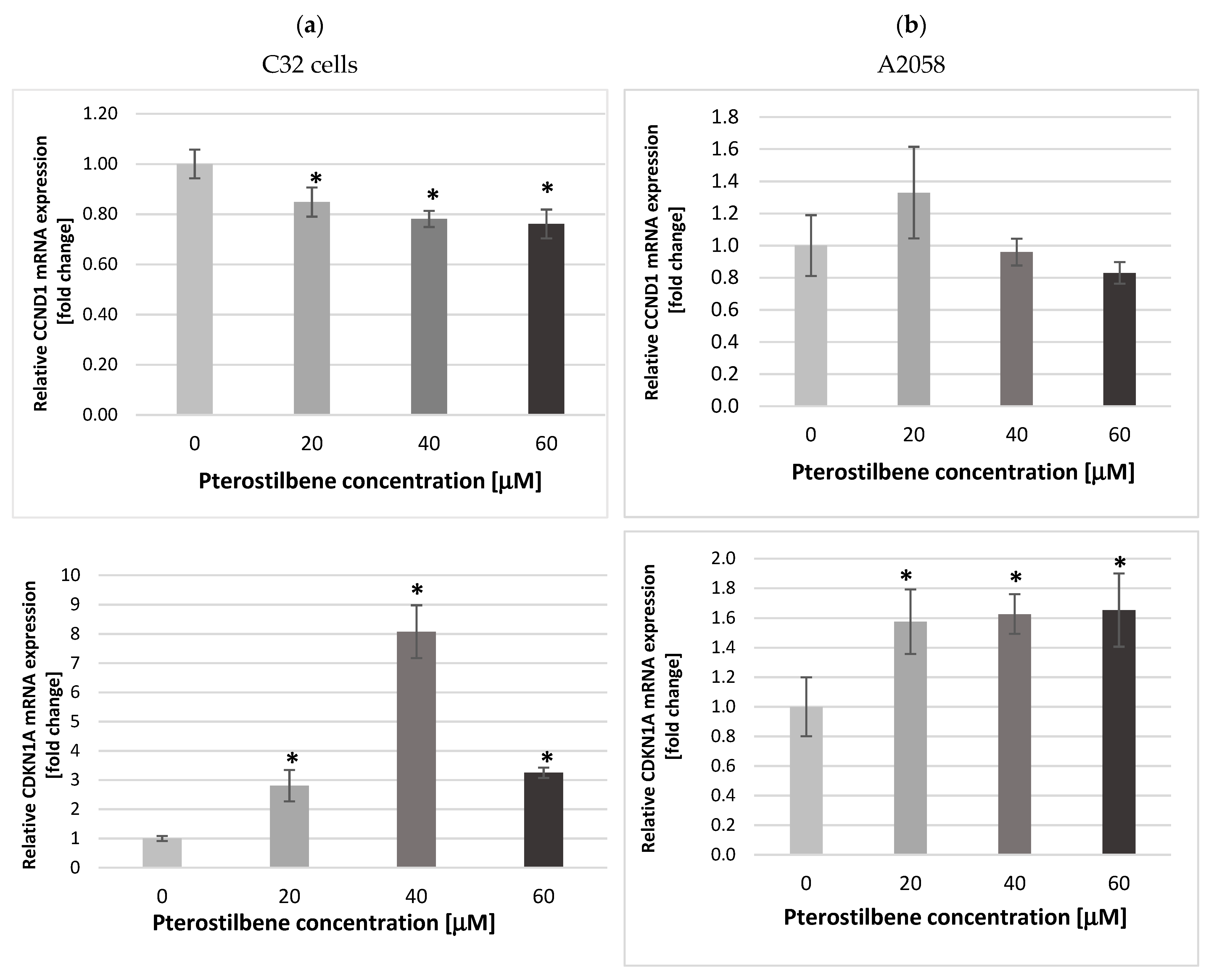
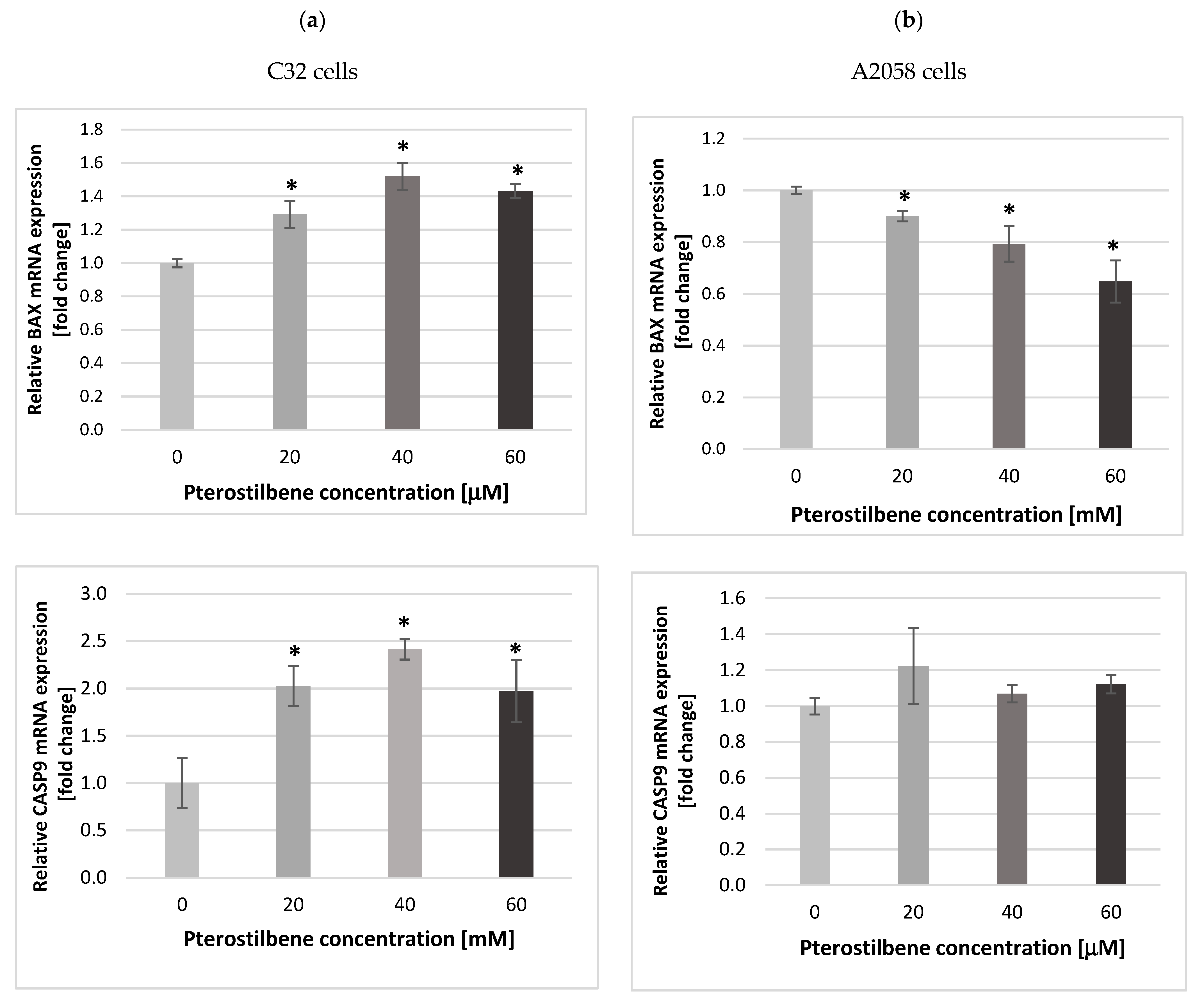
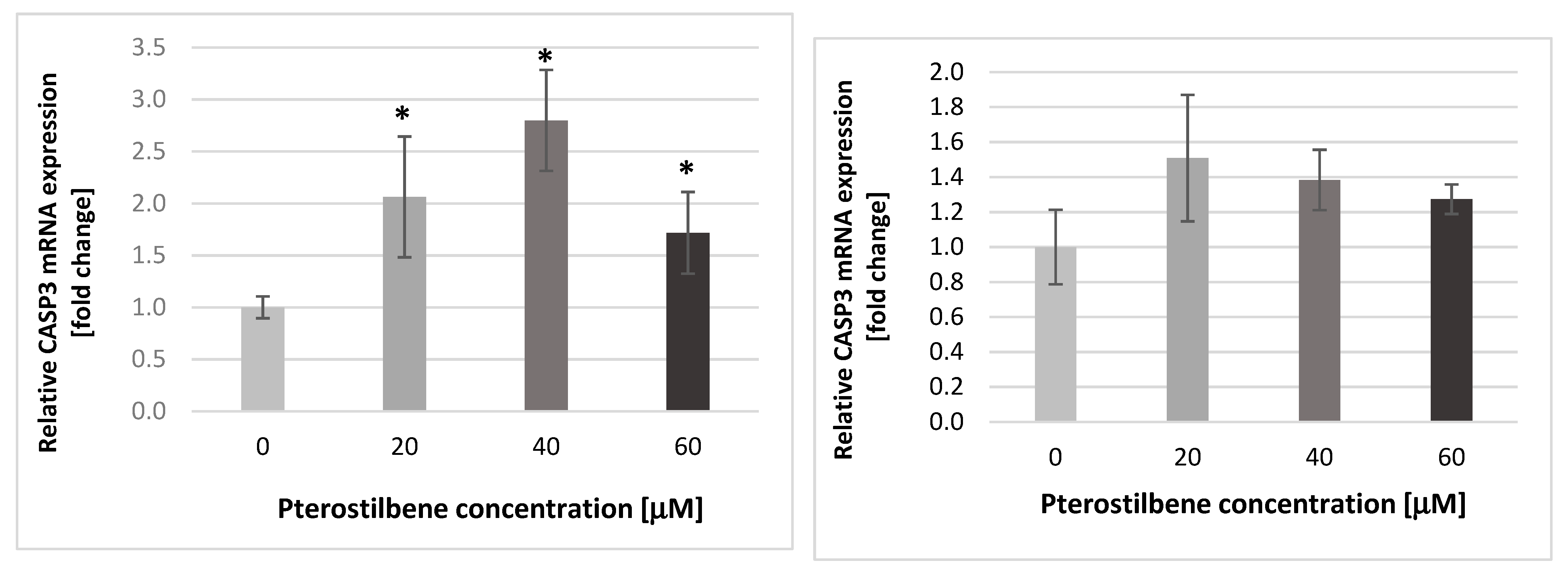
2.3. The Impact of Pterostilbene on Transcriptional Activity of Genes Encoding the Cell Cycle-Regulating Proteins
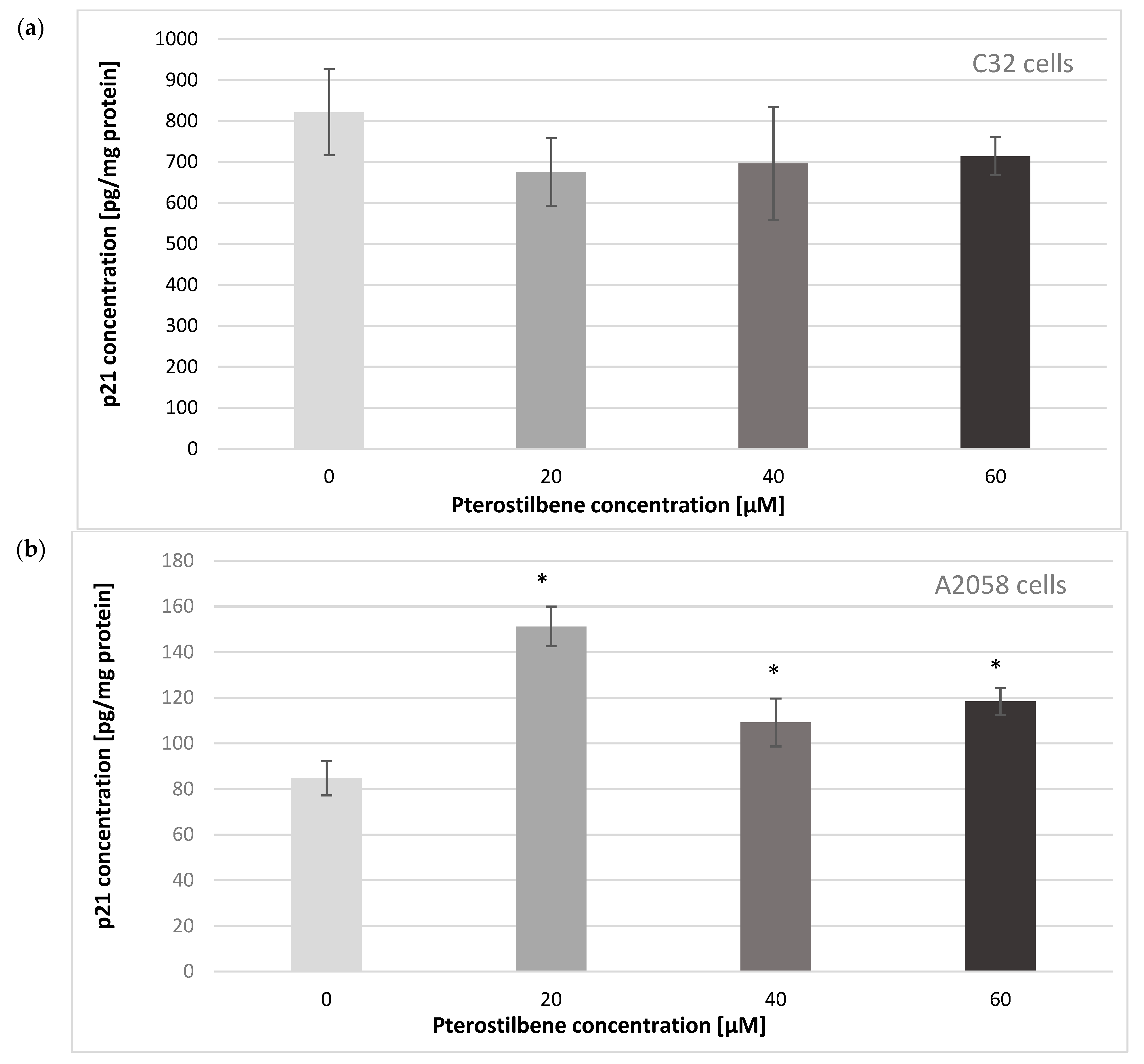
2.4. The Effect of Pterostilbene on the p21Waf1/Cip1 Protein Level
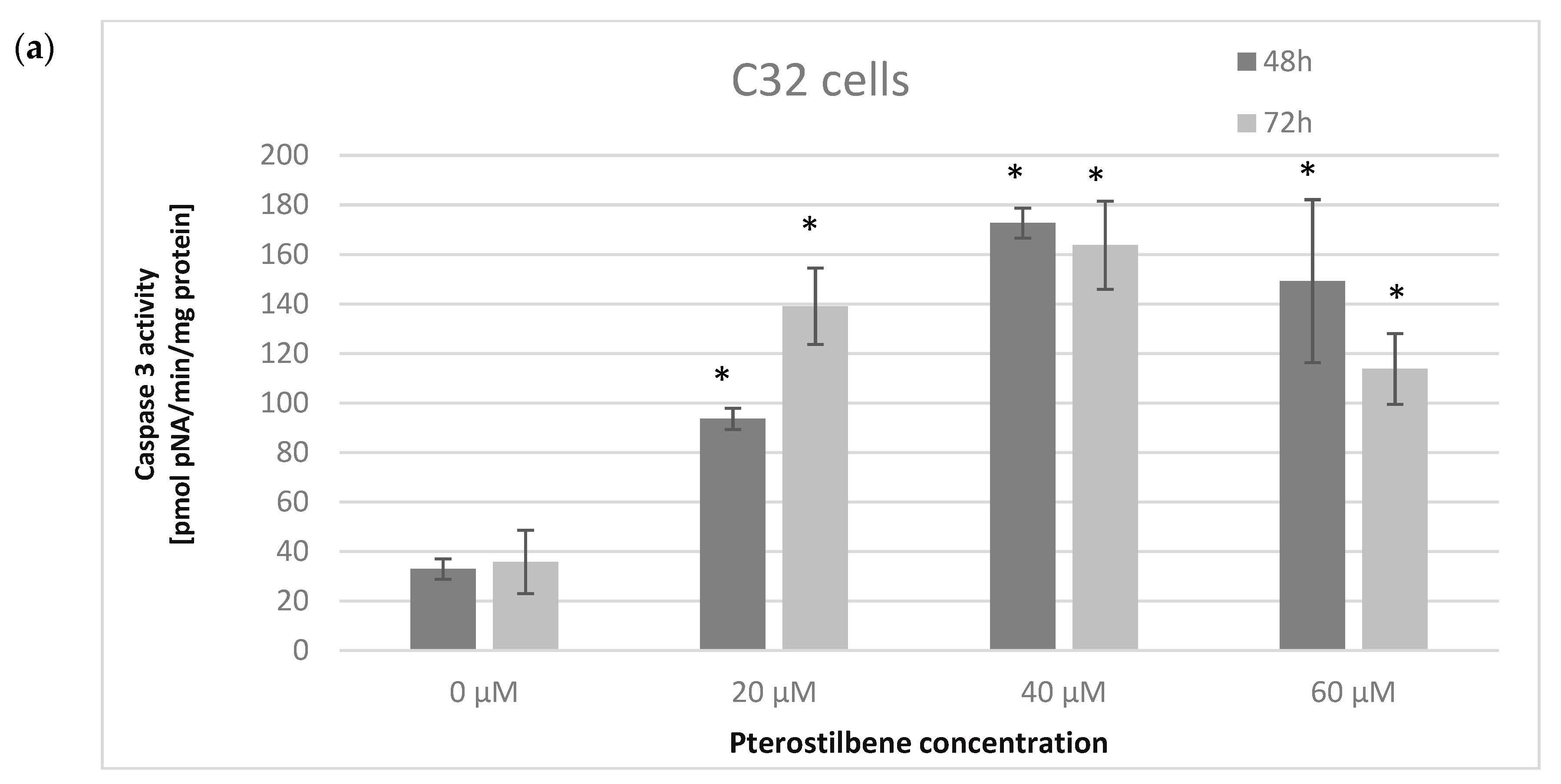
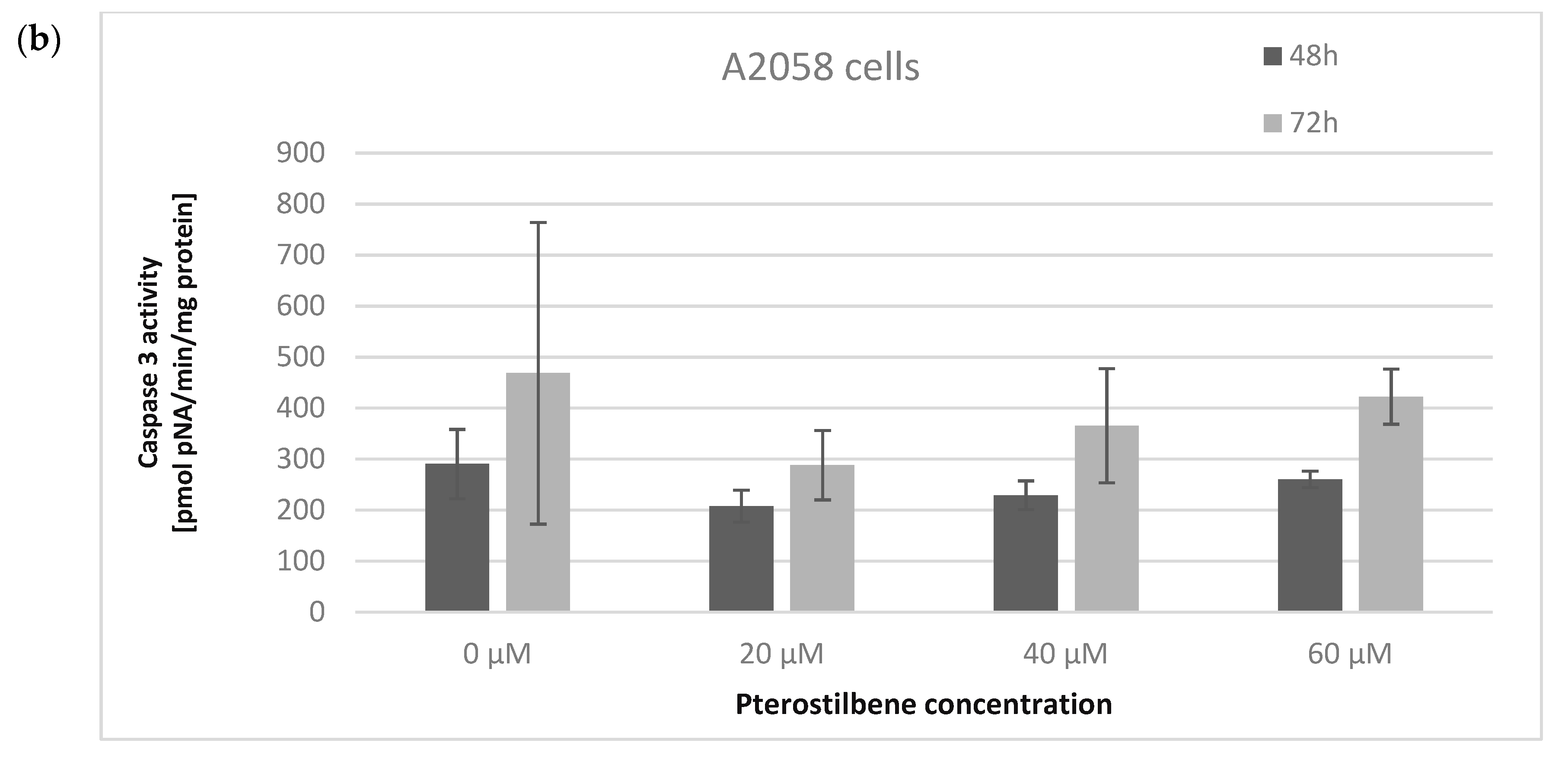
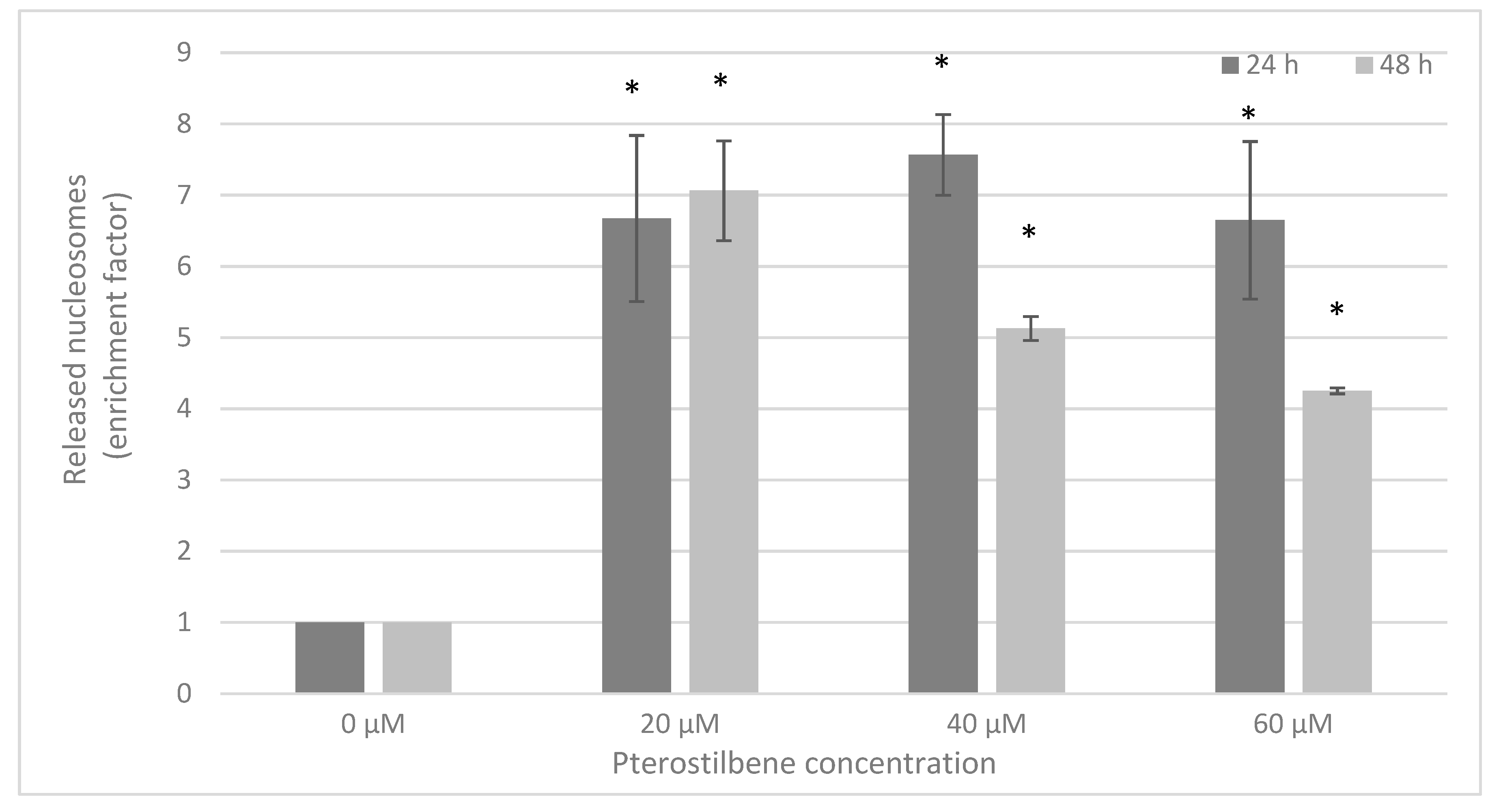
2.5. The Influence of Pterostilbene on Melanoma Cell Apoptosis
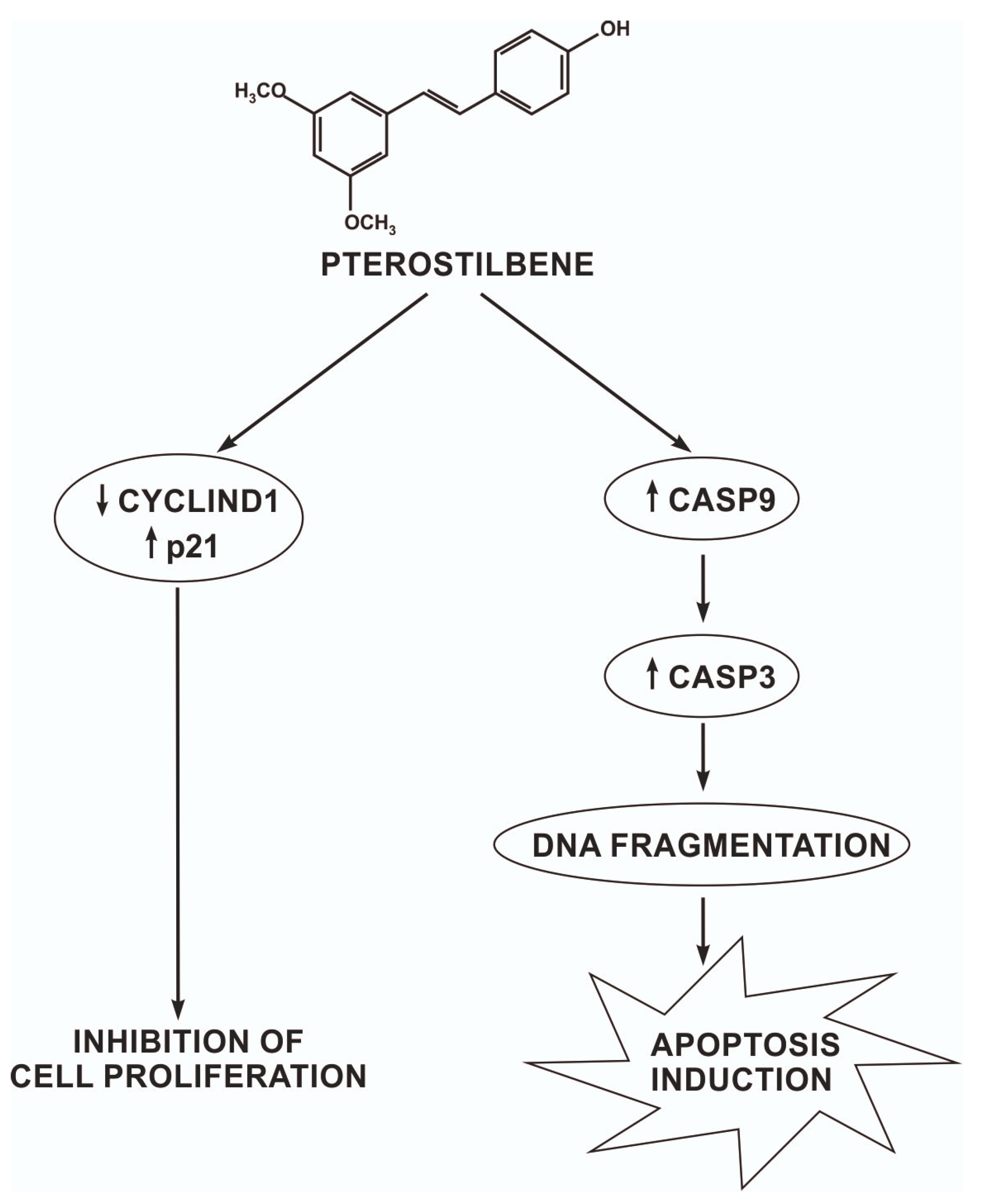
3. Discussion
4. Materials and Methods
4.1. Cell Lines and Cell Culture
4.2. Preparation of Pterostilbene Solution
4.3. Cell Proliferation Analysis
4.4. Cell Cycle Analysis
4.5. DNA Fragmentation Assay
4.6. Total RNA Extraction and Quantitative Real-Time RT-PCR (RT-qPCR)
4.7. Detection of the p21Waf1/Cip1 Protein Level
4.8. Caspase 3 Activity
4.9. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- von Schuckmann, L.A.; Hughes, M.C.B.; Ghiasvand, R.; Malt, M.; van der Pols, J.C.; Beesley, V.L.; Khosrotehrani, K.; Smithers, B.M.; Green, A.C. Risk of Melanoma Recurrence After Diagnosis of a High-Risk Primary Tumor. JAMA Dermatol. 2019, 155, 688. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lopes, J.; Rodrigues, C.M.P.; Gaspar, M.M.; Reis, C.P. Melanoma Management: From Epidemiology to Treatment and Latest Advances. Cancers 2022, 14, 4652. [Google Scholar] [CrossRef] [PubMed]
- Moreau, J.F.; Weissfeld, J.L.; Ferris, L.K. Characteristics and Survival of Patients with Invasive Amelanotic Melanoma in the USA. Melanoma Res. 2013, 23, 408–413. [Google Scholar] [CrossRef]
- Ricci, F.; Fania, L.; Paradisi, A.; Di Lella, G.; Pallotta, S.; Sobrino, L.; Panebianco, A.; Annessi, G.; Abeni, D. Delayed Melanoma Diagnosis in the COVID-19 Era: Increased Breslow Thickness in Primary Melanomas Seen after the COVID-19 Lockdown. J. Eur. Acad. Dermatol. Venereol. 2020, 34, e778–e779. [Google Scholar] [CrossRef] [PubMed]
- Chacón, M.; Pfluger, Y.; Angel, M.; Waisberg, F.; Enrico, D. Uncommon Subtypes of Malignant Melanomas: A Review Based on Clinical and Molecular Perspectives. Cancers 2020, 12, 2362. [Google Scholar] [CrossRef]
- Broussard, L.; Howland, A.; Ryu, S.; Song, K.; Norris, D.; Armstrong, C.A.; Song, P.I. Melanoma Cell Death Mechanisms. Chonnam Med. J. 2018, 54, 135. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mattia, G.; Puglisi, R.; Ascione, B.; Malorni, W.; Carè, A.; Matarrese, P. Cell death-based treatments of melanoma:conventional treatments and new therapeutic strategies. Cell Death Dis. 2018, 9, 112. [Google Scholar] [CrossRef] [Green Version]
- Hamilton, C.; Fox, J.P.; Longley, D.B.; Higgins, C.A. Therapeutics Targeting the Core Apoptotic Machinery. Cancers 2021, 13, 2618. [Google Scholar] [CrossRef]
- Forni, C.; Rossi, M.; Borromeo, I.; Feriotto, G.; Platamone, G.; Tabolacci, C.; Mischiati, C.; Beninati, S. Flavonoids: A Myth or a Reality for Cancer Therapy? Molecules 2021, 26, 3583. [Google Scholar] [CrossRef]
- Miyata, Y.; Shida, Y.; Hakariya, T.; Sakai, H. Anti-Cancer Effects of Green Tea Polyphenols Against Prostate Cancer. Molecules 2019, 24, 193. [Google Scholar] [CrossRef]
- Cháirez-Ramírez, M.H.; de la Cruz-López, K.G.; García-Carrancá, A. Polyphenols as Antitumor Agents Targeting Key Players in Cancer-Driving Signaling Pathways. Front. Pharmacol. 2021, 12, 710304. [Google Scholar] [CrossRef]
- Hazafa, A.; Iqbal, M.O.; Javaid, U.; Tareen, M.B.K.; Amna, D.; Ramzan, A.; Piracha, S.; Naeem, M. Inhibitory Effect of Polyphenols (Phenolic Acids, Lignans, and Stilbenes) on Cancer by Regulating Signal Transduction Pathways: A Review. Clin. Transl. Oncol. 2022, 24, 432–445. [Google Scholar] [CrossRef]
- Maleki Dana, P.; Sadoughi, F.; Asemi, Z.; Yousefi, B. The Role of Polyphenols in Overcoming Cancer Drug Resistance: A Comprehensive Review. Cell. Mol. Biol. Lett. 2022, 27, 10. [Google Scholar] [CrossRef] [PubMed]
- Ma, Z.; Zhang, X.; Xu, L.; Liu, D.; Di, S.; Li, W.; Zhang, J.; Zhang, H.; Li, X.; Han, J.; et al. Pterostilbene: Mechanisms of Its Action as Oncostatic Agent in Cell Models and in Vivo Studies. Pharmacol. Res. 2019, 145, 104265. [Google Scholar] [CrossRef]
- Elsherbini, A.M.; Sheweita, S.A.; Sultan, A.S. Pterostilbene as a Phytochemical Compound Induces Signaling Pathways Involved in the Apoptosis and Death of Mutant P53-Breast Cancer Cell Lines. Nutr. Cancer 2021, 73, 1976–1984. [Google Scholar] [CrossRef] [PubMed]
- Yu, C.L.; Yang, S.F.; Hung, T.W.; Lin, C.L.; Hsieh, Y.H.; Chiou, H.L. Inhibition of eIF2α dephosphorylation accelerates pterostilbene-induced cell death in human hepatocellular carcinoma cells in an ER stress and autophagy-dependent manner. Cell Death Dis. 2019, 10, 418. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, Z.; Yang, Y.; Di, S.; Feng, X.; Liu, D.; Jiang, S.; Hu, W.; Qin, Z.; Li, Y.; Lv, J.; et al. Pterostilbene Exerts Anticancer Activity on Non-Small-Cell Lung Cancer via Activating Endoplasmic Reticulum Stress. Sci. Rep. 2017, 7, 8091. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Surien, O.; Masre, S.F.; Basri, D.F.; Ghazali, A.R. Chemopreventive Effects of Oral Pterostilbene in Multistage Carcinogenesis of Skin Squamous Cell Carcinoma Mouse Model Induced by DMBA/TPA. Biomedicines 2022, 10, 2743. [Google Scholar] [CrossRef]
- Garbe, C.; Peris, K.; Hauschild, A.; Saiag, P.; Middleton, M.; Bastholt, L.; Grob, J.-J.; Malvehy, J.; Newton-Bishop, J.; Stratigos, A.J.; et al. Diagnosis and Treatment of Melanoma. European Consensus-Based Interdisciplinary Guideline—Update 2016. Eur. J. Cancer 2016, 63, 201–217. [Google Scholar] [CrossRef]
- Iqbal, J.; Abbasi, B.A.; Ahmad, R.; Batool, R.; Mahmood, T.; Ali, B.; Khalil, A.T.; Kanwal, S.; Afzal Shah, S.; Alam, M.M.; et al. Potential Phytochemicals in the Fight against Skin Cancer: Current Landscape and Future Perspectives. Biomed. Pharmacother. 2019, 109, 1381–1393. [Google Scholar] [CrossRef]
- Strickland, L.R.; Pal, H.C.; Elmets, C.A.; Afaq, F. Targeting Drivers of Melanoma with Synthetic Small Molecules and Phytochemicals. Cancer Lett. 2015, 359, 20–35. [Google Scholar] [CrossRef] [Green Version]
- Ranjan, A.; Ramachandran, S.; Gupta, N.; Kaushik, I.; Wright, S.; Srivastava, S.; Das, H.; Srivastava, S.; Prasad, S.; Srivastava, S.K. Role of Phytochemicals in Cancer Prevention. Int. J. Mol. Sci. 2019, 20, 4981. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Niles, R.M.; McFarland, M.; Weimer, M.B.; Redkar, A.; Fu, Y.-M.; Meadows, G.G. Resveratrol Is a Potent Inducer of Apoptosis in Human Melanoma Cells. Cancer Lett. 2003, 190, 157–163. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Yu, T.; Zhu, C.; Sun, H.; Qiu, Y.; Zhu, X.; Li, J. Resveratrol Triggers Protective Autophagy Through the Ceramide/Akt/MTOR Pathway in Melanoma B16 Cells. Nutr. Cancer 2014, 66, 435–440. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.-Z.; Zhang, J.; Zeng, J.; Liu, S.; Zhou, F.; Zhang, F.; Giampieri, F.; Cianciosi, D.; Forbes-Hernandez, T.Y.; Ansary, J.; et al. Resveratrol Inhibits the Proliferation of Melanoma Cells by Modulating Cell Cycle. Int. J. Food Sci. Nutr. 2020, 71, 84–93. [Google Scholar] [CrossRef] [PubMed]
- Pannu, N.; Bhatnagar, A. Resveratrol: From Enhanced Biosynthesis and Bioavailability to Multitargeting Chronic Diseases. Biomed. Pharmacother. 2019, 109, 2237–2251. [Google Scholar] [CrossRef]
- Chakraborty, A.; Gupta, N.; Ghosh, K.; Roy, P. In Vitro Evaluation of the Cytotoxic, Anti-Proliferative and Anti-Oxidant Properties of Pterostilbene Isolated from Pterocarpus Marsupium. Toxicol. Vitr. 2010, 24, 1215–1228. [Google Scholar] [CrossRef]
- Pari, L.; Satheesh, M.A. Effect of Pterostilbene on Hepatic Key Enzymes of Glucose Metabolism in Streptozotocin- and Nicotinamide-Induced Diabetic Rats. Life Sci. 2006, 79, 641–645. [Google Scholar] [CrossRef]
- Rimando, A.M.; Nagmani, R.; Feller, D.R.; Yokoyama, W. Pterostilbene, a New Agonist for the Peroxisome Proliferator-Activated Receptor α-Isoform, Lowers Plasma Lipoproteins and Cholesterol in Hypercholesterolemic Hamsters. J. Agric. Food Chem. 2005, 53, 3403–3407. [Google Scholar] [CrossRef]
- Li, Y.-R.; Li, S.; Lin, C.-C. Effect of Resveratrol and Pterostilbene on Aging and Longevity: Effect of Resveratrol and Pterostilbene on Aging and Longevity. BioFactors 2018, 44, 69–82. [Google Scholar] [CrossRef]
- Estrela, J.M.; Ortega, A.; Mena, S.; Rodriguez, M.L.; Asensi, M. Pterostilbene: Biomedical Applications. Crit. Rev. Clin. Lab. Sci. 2013, 50, 65–78. [Google Scholar] [CrossRef] [PubMed]
- Wakimoto, R.; Ono, M.; Takeshima, M.; Higuchi, T.; Nakano, S. Differential Anticancer Activity of Pterostilbene Against Three Subtypes of Human Breast Cancer Cells. Anticancer Res. 2017, 37, 6153–6159. [Google Scholar] [CrossRef] [PubMed]
- McCormack, D.E.; Mannal, P.; McDonald, D.; Tighe, S.; Hanson, J.; McFadden, D. Genomic Analysis of Pterostilbene Predicts Its Antiproliferative Effects Against Pancreatic Cancer In Vitro and In Vivo. J. Gastrointest. Surg. 2012, 16, 1136–1143. [Google Scholar] [CrossRef] [Green Version]
- Paul, S.; Rimando, A.M.; Lee, H.J.; Ji, Y.; Reddy, B.S.; Suh, N. Anti-Inflammatory Action of Pterostilbene Is Mediated through the P38 Mitogen-Activated Protein Kinase Pathway in Colon Cancer Cells. Cancer Prev. Res. 2009, 2, 650–657. [Google Scholar] [CrossRef] [Green Version]
- Wang, Z.; Wang, T.; Chen, X.; Cheng, J.; Wang, L. Pterostilbene Regulates Cell Proliferation and Apoptosis in Non-small-cell Lung Cancer via Targeting COX-2. Biotechnol. Appl. Biochem. 2022. [Google Scholar] [CrossRef] [PubMed]
- Wen, W.; Lowe, G.; Roberts, C.; Finlay, J.; Han, E.; Glackin, C.; Dellinger, T. Pterostilbene Suppresses Ovarian Cancer Growth via Induction of Apoptosis and Blockade of Cell Cycle Progression Involving Inhibition of the STAT3 Pathway. Int. J. Mol. Sci. 2018, 19, 1983. [Google Scholar] [CrossRef] [Green Version]
- Benlloch, M.; Obrador, E.; Valles, S.L.; Rodriguez, M.L.; Sirerol, J.A.; Alcácer, J.; Pellicer, J.A.; Salvador, R.; Cerdá, C.; Sáez, G.T.; et al. Pterostilbene Decreases the Antioxidant Defenses of Aggressive Cancer Cells In Vivo: A Physiological Glucocorticoids- and Nrf2-Dependent Mechanism. Antioxid. Redox Signal. 2016, 24, 974–990. [Google Scholar] [CrossRef] [Green Version]
- Mena, S.; Rodríguez, M.L.; Ponsoda, X.; Estrela, J.M.; Jäättela, M.; Ortega, A.L. Pterostilbene-Induced Tumor Cytotoxicity: A Lysosomal Membrane Permeabilization-Dependent Mechanism. PLoS ONE 2012, 7, e44524. [Google Scholar] [CrossRef]
- Qian, Y.; Liu, Z.; Zhang, Z.; Levenson, A.; Li, K. Pterostilbene Increases PTEN Expression through the Targeted Downregulation of MicroRNA-19a in Hepatocellular Carcinoma. Mol. Med. Rep. 2018, 17, 5193–5201. [Google Scholar] [CrossRef] [Green Version]
- Kong, Y.; Chen, G.; Xu, Z.; Yang, G.; Li, B.; Wu, X.; Xiao, W.; Xie, B.; Hu, L.; Sun, X.; et al. Pterostilbene Induces Apoptosis and Cell Cycle Arrest in Diffuse Large B-Cell Lymphoma Cells. Sci. Rep. 2016, 6, 37417. [Google Scholar] [CrossRef]
- Dai, B.; Zhao, X.F.; Mazan-Mamczarz, K.; Hagner, P.; Corl, S.; Bahassi, E.M.; Lu, S.; Stambrook, P.J.; Shapiro, P.; Gartenhaus, R.B. Functional and Molecular Interactions between ERK and CHK2 in Diffuse Large B-Cell Lymphoma. Nat. Commun. 2011, 2, 402. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, G.; Xu, Z.; Chang, G.; Hou, J.; Hu, L.; Zhang, Y.; Yu, D.; Li, B.; Chang, S.; Xie, Y.; et al. The Blueberry Component Pterostilbene Has Potent Anti-Myeloma Activity in Bortezomib-Resistant Cells. Oncol. Rep. 2017, 38, 488–496. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ko, C.-P.; Lin, C.-W.; Chen, M.-K.; Yang, S.-F.; Chiou, H.-L.; Hsieh, M.-J. Pterostilbene Induce Autophagy on Human Oral Cancer Cells through Modulation of Akt and Mitogen-Activated Protein Kinase Pathway. Oral Oncol. 2015, 51, 593–601. [Google Scholar] [CrossRef] [PubMed]
- Siedlecka-Kroplewska, K.; Jozwik, A.; Boguslawski, W.; Wozniak, M.; Zauszkiewicz-Pawlak, A.; Spodnik, J.H.; Rychlowski, M.; Kmiec, Z. Pterostilbene Induces Accumulation of Autophagic Vacuoles Followed by Cell Death in HL60 Human Leukemia Cells. J. Physiol. Pharmacol. Off. J. Pol. Physiol. Soc. 2013, 64, 545–556. [Google Scholar]
- González-Ruiz, L.; González-Moles, M.Á.; González-Ruiz, I.; Ruiz-Ávila, I.; Ramos-García, P. Prognostic and Clinicopathological Significance of CCND1/Cyclin D1 Upregulation in Melanomas: A Systematic Review and Comprehensive Meta-Analysis. Cancers 2021, 13, 1314. [Google Scholar] [CrossRef]
- Karimian, A.; Ahmadi, Y.; Yousefi, B. Multiple Functions of P21 in Cell Cycle, Apoptosis and Transcriptional Regulation after DNA Damage. DNA Repair 2016, 42, 63–71. [Google Scholar] [CrossRef] [PubMed]
- Piepkorn, M. Melanoma Genetics: An Update with Focus on the CDKN2A(P16)/ARF Tumor Suppressors. J. Am. Acad. Dermatol. 2000, 42, 705–726. [Google Scholar] [CrossRef] [PubMed]
- Xing, F.; Persaud, Y.; Pratilas, C.A.; Taylor, B.S.; Janakiraman, M.; She, Q.-B.; Gallardo, H.; Liu, C.; Merghoub, T.; Hefter, B.; et al. Concurrent Loss of the PTEN and RB1 Tumor Suppressors Attenuates RAF Dependence in Melanomas Harboring V600EBRAF. Oncogene 2012, 31, 446–457. [Google Scholar] [CrossRef] [Green Version]
- Abbas, T.; Dutta, A. P21 in Cancer: Intricate Networks and Multiple Activities. Nat. Rev. Cancer 2009, 9, 400–414. [Google Scholar] [CrossRef]
- Chen, R.-J.; Lee, Y.-H.; Yeh, Y.-L.; Wu, W.-S.; Ho, C.-T.; Li, C.-Y.; Wang, B.-J.; Wang, Y.-J. Autophagy-Inducing Effect of Pterostilbene: A Prospective Therapeutic/Preventive Option for Skin Diseases. J. Food Drug Anal. 2017, 25, 125–133. [Google Scholar] [CrossRef]
- Chen, R.-J.; Kuo, H.-C.; Cheng, L.-H.; Lee, Y.-H.; Chang, W.-T.; Wang, B.-J.; Wang, Y.-J.; Cheng, H.-C. Apoptotic and Nonapoptotic Activities of Pterostilbene against Cancer. Int. J. Mol. Sci. 2018, 19, 287. [Google Scholar] [CrossRef]
- Schneider, J.G.; Alosi, J.A.; McDonald, D.E.; McFadden, D.W. Effects of Pterostilbene on Melanoma Alone and in Synergy with Inositol Hexaphosphate. Am. J. Surg. 2009, 198, 679–684. [Google Scholar] [CrossRef] [PubMed]
- Pfeffer, C.; Singh, A. Apoptosis: A Target for Anticancer Therapy. Int. J. Mol. Sci. 2018, 19, 448. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Soengas, M.S.; Capodieci, P.; Polsky, D.; Mora, J.; Esteller, M.; Opitz-Araya, X.; McCombie, R.; Herman, J.G.; Gerald, W.L.; Lazebnik, Y.A.; et al. Inactivation of the Apoptosis Effector Apaf-1 in Malignant Melanoma. Nature 2001, 409, 207–211. [Google Scholar] [CrossRef]
- Robertson, G.P. Functional and Therapeutic Significance of Akt Deregulation in Malignant Melanoma. Cancer Metastasis Rev. 2005, 24, 273–285. [Google Scholar] [CrossRef] [PubMed]
- Prokhorova, E.A.; Kopeina, G.S.; Lavrik, I.N.; Zhivotovsky, B. Apoptosis Regulation by Subcellular Relocation of Caspases. Sci. Rep. 2018, 8, 12199. [Google Scholar] [CrossRef]
- Nutakul, W.; Sobers, H.S.; Qiu, P.; Dong, P.; Decker, E.A.; McClements, D.J.; Xiao, H. Inhibitory Effects of Resveratrol and Pterostilbene on Human Colon Cancer Cells: A Side-by-Side Comparison. J. Agric. Food Chem. 2011, 59, 10964–10970. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moon, D.; McCormack, D.; McDonald, D.; McFadden, D. Pterostilbene Induces Mitochondrially Derived Apoptosis in Breast Cancer Cells in Vitro. J. Surg. Res. 2013, 180, 208–215. [Google Scholar] [CrossRef] [PubMed]
- Pan, M.-H.; Chang, Y.-H.; Badmaev, V.; Nagabhushanam, K.; Ho, C.-T. Pterostilbene Induces Apoptosis and Cell Cycle Arrest in Human Gastric Carcinoma Cells. J. Agric. Food Chem. 2007, 55, 7777–7785. [Google Scholar] [CrossRef]
- Lin, C.-S.; Lin, W.-S.; Lin, C.-L.; Kao, C.-H. Carvedilol Use Is Associated with Reduced Cancer Risk: A Nationwide Population-Based Cohort Study. Int. J. Cardiol. 2015, 184, 9–13. [Google Scholar] [CrossRef]
- Lin, V.C.-H.; Tsai, Y.-C.; Lin, J.-N.; Fan, L.-L.; Pan, M.-H.; Ho, C.-T.; Wu, J.-Y.; Way, T.-D. Activation of AMPK by Pterostilbene Suppresses Lipogenesis and Cell-Cycle Progression in P53 Positive and Negative Human Prostate Cancer Cells. J. Agric. Food Chem. 2012, 60, 6399–6407. [Google Scholar] [CrossRef]
- Pan, M.-H.; Lin, Y.-T.; Lin, C.-L.; Wei, C.-S.; Ho, C.-T.; Chen, W.-J. Suppression of Heregulin- β 1/HER2-Modulated Invasive and Aggressive Phenotype of Breast Carcinoma by Pterostilbene via Inhibition of Matrix Metalloproteinase-9, P38 Kinase Cascade and Akt Activation. Evid. Based Complement. Altern. Med. 2011, 2011, 562187. [Google Scholar] [CrossRef] [Green Version]
- Chen, R.-J.; Ho, C.-T.; Wang, Y.-J. Pterostilbene Induces Autophagy and Apoptosis in Sensitive and Chemoresistant Human Bladder Cancer Cells. Mol. Nutr. Food Res. 2010, 54, 1819–1832. [Google Scholar] [CrossRef] [PubMed]
- Ferrer, P.; Asensi, M.; Segarra, R.; Ortega, A.; Benlloch, M.; Obrador, E.; Varea, M.T.; Asensio, G.; Jordá, L.; Estrela, J.M. Association between Pterostilbene and Quercetin Inhibits Metastatic Activity of B16 Melanoma. Neoplasia 2005, 7, 37–47. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Su, Y.; Bintz, M.; Yang, Y.; Robert, L.; Ng, A.H.C.; Liu, V.; Ribas, A.; Heath, J.R.; Wei, W. Phenotypic heterogeneity and evolution of melanoma cells associated with targeted therapy resistance. PLoS Comput. Biol. 2019, 15, e1007034. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Su, Y.; Ko, M.E.; Cheng, H.; Zhu, R.; Xue, M.; Wang, J.; Lee, J.W.; Frankiw, L.; Xu, A.; Wong, S.; et al. Multi-Omic Single-Cell Snapshots Reveal Multiple Independent Trajectories to Drug Tolerance in a Melanoma Cell Line. Nat. Commun. 2020, 11, 2345. [Google Scholar] [CrossRef] [PubMed]
- Su, Y.; Shi, Q.; Wei, W. Single cell proteomics in biomedicine: High-dimensional data acquisition, visualization, and analysis. Proteomics 2017, 17, 10. [Google Scholar] [CrossRef] [Green Version]
- Su, Y.; Wei, W.; Robert, L.; Xue, M.; Tsoi, J.; Garcia-Diaz, A.; Homet Moreno, B.; Kim, J.; Ng, R.H.; Lee, J.W.; et al. Single-cell analysis resolves the cell state transition and signaling dynamics associated with melanoma drug-induced resistance. Proc. Natl. Acad. Sci. USA 2017, 114, 13679–13684. [Google Scholar] [CrossRef] [Green Version]
- Du, J.; Su, Y.; Qian, C.; Yuan, D.; Miao, K.; Lee, D.; Ng, A.H.C.; Wijker, R.S.; Ribas, A.; Levine, R.D.; et al. Raman-guided subcellular pharmaco-metabolomics for metastatic melanoma cells. Nat. Commun. 2020, 11, 4830. [Google Scholar] [CrossRef]
- Hachey, S.J.; Boiko, A.D. Therapeutic implications of melanoma heterogeneity. Exp. Dermatol. 2016, 25, 497–500. [Google Scholar] [CrossRef] [Green Version]
- Nawrot-Hadzik, I.; Choromańska, A.; Abel, R.; Preissner, R.; Saczko, J.; Matkowski, A.; Hadzik, J. Cytotoxic Effect of Vanicosides A and B from Reynoutria Sachalinensis against Melanotic and Amelanotic Melanoma Cell Lines and in Silico Evaluation for Inhibition of BRAFV600E and MEK1. Int. J. Mol. Sci. 2020, 21, 4611. [Google Scholar] [CrossRef] [PubMed]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Forward Primer (5′-3′) | Reverse Primer (5′-3′) |
|---|---|---|
| CCND1 | GCCTCTAAGATGAAGGAGAC | CCATTTGCAGCAGCTC |
| CDKN1A | AGGGATTTCTTCTGTTCAGG | GACAAAGTCGAAGTTCCATC |
| CASP3 | GGCCTGCCGTGGTACAGAACTGG | AGCGACTGGATGAACCAGGAGCCA |
| CASP9 | GACCGGAAACACCCAGACCAGTGGA | GCAGTGGCCACAGGGCTCCAT |
| BAX | TCTGAGCAGATCATGAAGAC | TCCATGTTACTGTCCAGTTC |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wawszczyk, J.; Jesse, K.; Kapral, M. Pterostilbene-Mediated Inhibition of Cell Proliferation and Cell Death Induction in Amelanotic and Melanotic Melanoma. Int. J. Mol. Sci. 2023, 24, 1115. https://doi.org/10.3390/ijms24021115
Wawszczyk J, Jesse K, Kapral M. Pterostilbene-Mediated Inhibition of Cell Proliferation and Cell Death Induction in Amelanotic and Melanotic Melanoma. International Journal of Molecular Sciences. 2023; 24(2):1115. https://doi.org/10.3390/ijms24021115
Chicago/Turabian StyleWawszczyk, Joanna, Katarzyna Jesse, and Małgorzata Kapral. 2023. "Pterostilbene-Mediated Inhibition of Cell Proliferation and Cell Death Induction in Amelanotic and Melanotic Melanoma" International Journal of Molecular Sciences 24, no. 2: 1115. https://doi.org/10.3390/ijms24021115