
Sperm DNA Fragmentation and Sperm-Borne miRNAs: Molecular Biomarkers of Embryo Development?
 , , , , ,
, , , , ,
Abstract
:1. Introduction
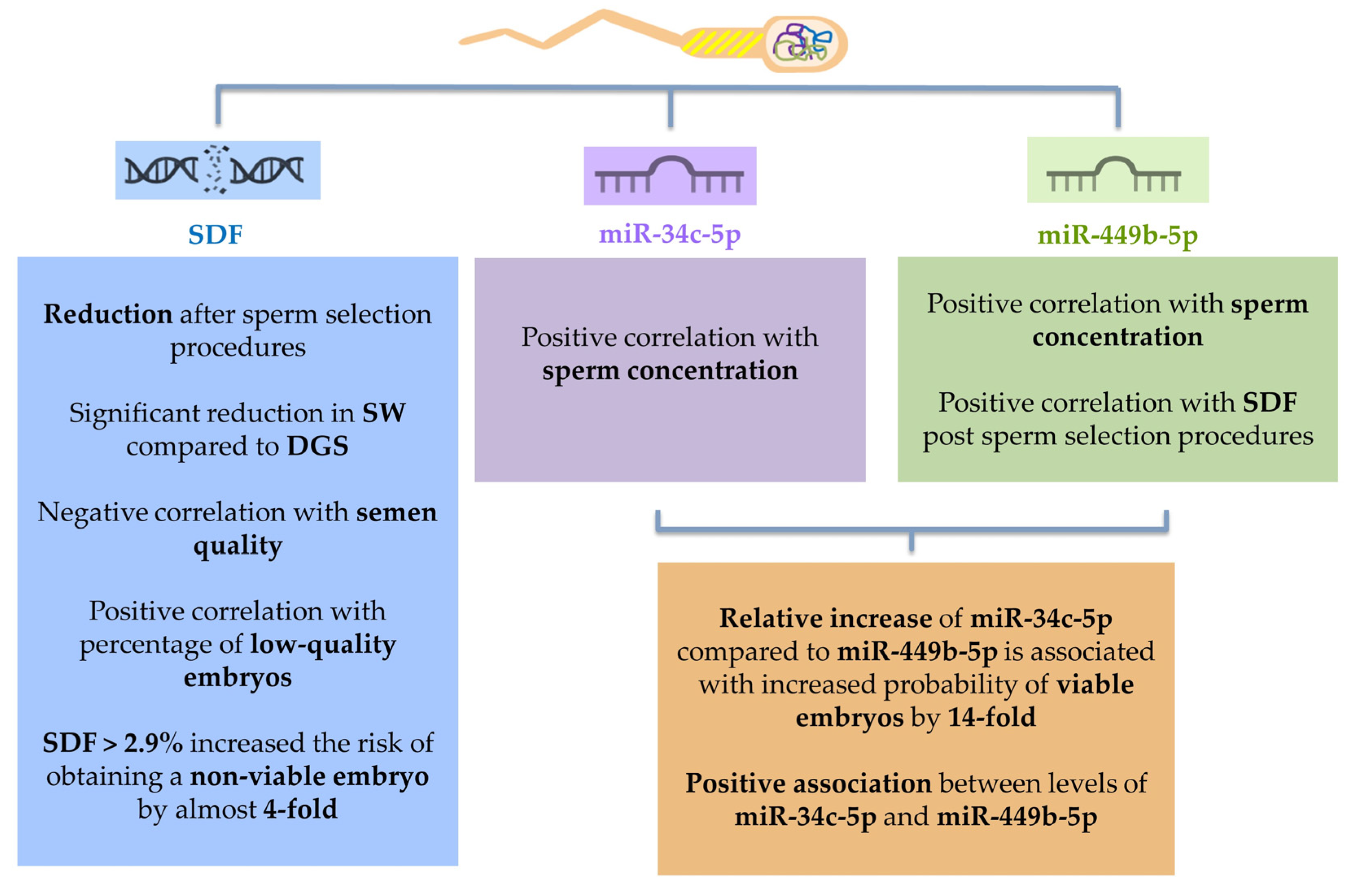
2. Results
2.1. Demographic Characteristics
2.2. Sperm DNA Fragmentation (SDF) Evaluation
2.2.1. Semen Parameters
2.2.2. ICSI Outcome
2.3. Sperm miR-34c-5p and miR-449b-5p
2.3.1. Semen Parameters
2.3.2. ICSI Outcome
3. Discussion
3.1. Sperm DNA Fragmentation (SDF)
3.1.1. Sperm DNA Fragmentation and Seminal Parameters
3.1.2. Sperm DNA Fragmentation and ICSI Outcome
3.2. Sperm-Borne miRNA
3.2.1. Sperm miR-34c-5p and miR-449b-5p and Spermatogenesis
3.2.2. Sperm miR-34c-5p and miR-449b-5p and ICSI Outcome
4. Materials and Methods
4.1. Patients
- age over 18 years and under 50 for male partners;
- age over 18 years and under 43 for female partners;
- presence of mature oocytes suitable for insemination;
- possibility of taking a sufficient aliquot of semen pre and post sperm selection procedures for sperm DNA fragmentation and miRNAs analyses.
- presence of endometriosis diagnosis for female partners;
- presence of recurrent pregnancy loss;
- presence of andrological pathologies (cryptorchidism, clinically relevant varicocele, hypogonadism, etc.) and/or endocrinological diseases which may interfere with semen quality;
- azoospermia (both obstructive and non-obstructive);
- genetic syndromes and/or abnormal karyotype;
- previous treatment(s) with chemo/radiotherapy and/or potentially gonadotoxic drugs for oncological pathology;
- previous treatment(s) with potentially gonadotoxic drugs for non-oncological pathologies.
4.2. Semen Analysis
4.3. Sperm Selection Procedures
- Swim-up (SW): the semen sample was diluted 1:1 with a culture medium (Multipurpose Handling Medium—MHM; Fujifilm, USA) pre-warmed to 37 °C and centrifuged at 1200 rpm for 10 min. At the end of centrifugation, the supernatant was removed and then 100–300 µL of culture medium pre-warmed to 37 °C and containing 10% human albumin was layered on the pellet. After a one-hour incubation at 37 °C in 5% CO2, the upper layer was removed to retrieve the spermatozoa most suitable for insemination.
- Density Gradient Centrifugation (DGC): equal volumes of 80% gradient, 40% gradient (MHM; Fujifilm, USA) and finally the seminal fluid was stratified in a conical test tube. The tube was centrifuged for 10 min at 1500 rpm in order to obtain the sedimentation of sperm with the best microscopic characteristics due to the filtering action of the gradients. The pellet obtained at the end of centrifugation was suspended in culture medium and incubated for one hour at 37 °C in 5% CO2 and then used to inseminate the oocytes.
4.4. Assisted Reproductive Procedures
- Grade 1 embryos: stage-specific cell size, <10% fragmentation and no multinucleation;
- Grade 2 embryos: stage-specific cell size for majority of cells, 10–25% fragmentation and no multinucleation;
- Grade 3 embryos had cell size not stage specific, >35% fragmentation and evidence of multinucleation.
4.5. Sperm DNA Fragmentation (SDF)
4.6. MicroRNA Analysis
4.7. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Krawetz, S.A. Paternal contribution: New insights and future challenges. Nat. Rev. Genet. 2005, 6, 633–642. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization. WHO Laboratory Manual for the Examination and Processing of Human Semen, 6th ed.; World Health Organization: Geneva, Switzerland, 2021. [Google Scholar]
- Paoli, D.; Pallotti, F.; Lenzi, A.; Lombardo, F. Fatherhood and Sperm DNA Damage in Testicular Cancer Patients. Front. Endocrinol. 2018, 9, 506. [Google Scholar] [CrossRef]
- Jodar, M. Sperm and seminal plasma RNAs: What roles do they play beyond fertilization? Reproduction 2019, 158, R113–R123. [Google Scholar] [CrossRef]
- Santiago, J.; Silva, J.V.; Howl, J.; Santos, M.A.S.; Fardilha, M. All you need to know about sperm RNAs. Hum. Reprod. Update 2021, 28, 67–91. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.M.; Pang, R.T.; Chiu, P.C.; Wong, B.P.; Lao, K.; Lee, K.F.; Yeung, W.S. Sperm-borne microRNA-34c is required for the first cleavage division in mouse. Proc. Natl. Acad. Sci. USA 2012, 109, 490–494. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cui, L.; Fang, L.; Shi, B.; Qiu, S.; Ye, Y. Spermatozoa micro ribonucleic acid-34c level is correlated with intracytoplasmic sperm injection outcomes. Fertil. Steril. 2015, 104, 312–317. [Google Scholar] [CrossRef] [Green Version]
- Dorostghoal, M.; Galehdari, H.; Hemadi, M.; Davoodi, E. Sperm miR-34c-5p-5p Transcript Content and Its Association with Sperm Parameters in Unexplained Infertile Men. Reprod. Sci. 2022, 29, 84–90. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Gao, Y.; Qu, P.; Qing, S.; Qiao, F.; Zhang, Y.; Mager, J.; Wang, Y. Sperm-borne miR-449b-5p influences cleavage, epigenetic reprogramming and apoptosis of SCNT embryos in bovine. Sci. Rep. 2017, 7, 13403. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Najafipour, R.; Momeni, A.; Yousefipour, F.; Mousavi, S.; Moghbelinejad, S. Underexpression of hsa-miR-449 family and their promoter hypermethylation in infertile men: A case-control study. Int. J. Reprod. Biomed. 2021, 19, 23–34. [Google Scholar] [CrossRef]
- Llavanera, M.; Delgado-Bermúdez, A.; Ribas-Maynou, J.; Salas-Huetos, A.; Yeste, M. A systematic review identifying fertility biomarkers in semen: A clinical approach through Omics to diagnose male infertility. Fertil. Steril. 2022, 118, 291–313. [Google Scholar] [CrossRef]
- Carlini, T.; Paoli, D.; Pelloni, M.; Faja, F.; Dal Lago, A.; Lombardo, F.; Lenzi, A.; Gandini, L. Sperm DNA fragmentation in Italian couples with recurrent pregnancy loss. Reprod. Biomed. Online 2017, 34, 58–65. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sergerie, M.; Laforest, G.; Bujan, L.; Bissonnette, F.; Bleau, G. Sperm DNA fragmentation: Threshold value in male fertility. Hum. Reprod. 2005, 20, 3446–3451. [Google Scholar] [CrossRef] [PubMed]
- Ribas-Maynou, J.; García-Peiró, A.; Fernández-Encinas, A.; Abad, C.; Amengual, M.J.; Prada, E.; Navarro, J.; Benet, J. Comprehensive analysis of sperm DNA fragmentation by five different assays: TUNEL assay, SCSA, SCD test and alkaline and neutral Comet assay. Andrology 2013, 1, 715–722. [Google Scholar] [CrossRef] [PubMed]
- Hichri, R.; Amor, H.; Khammari, M.; Harzallah, M.; El Fekih, S.; Saad, A.; Ajina, M.; Ben Ali, H. Apoptotic sperm biomarkers and the correlation between conventional sperm parameters and clinical characteristics. Andrologia 2018, 50, e12813. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Muratori, M.; Marchiani, S.; Tamburrino, L.; Baldi, E. Sperm DNA Fragmentation: Mechanisms of Origin. Adv. Exp. Med. Biol. 2019, 1166, 75–85. [Google Scholar] [CrossRef] [PubMed]
- Zini, A. Are sperm chromatin and DNA defects relevant in the clinic? Syst. Biol. Reprod. Med. 2011, 57, 78–85. [Google Scholar] [CrossRef] [PubMed]
- Ruvolo, G.; Fattouh, R.R.; Bosco, L.; Brucculeri, A.M.; Cittadini, E. New molecular markers for the evaluation of gamete quality. J. Assist. Reprod. Genet. 2013, 30, 207–212. [Google Scholar] [CrossRef] [Green Version]
- Borges, E., Jr.; Zanetti, B.F.; Setti, A.S.; Braga, D.P.A.F.; Provenza, R.R.; Iaconelli, A., Jr. Sperm DNA fragmentation is correlated with poor embryo development, lower implantation rate, and higher miscarriage rate in reproductive cycles of non-male factor infertility. Fertil. Steril. 2019, 112, 483–490. [Google Scholar] [CrossRef]
- Tarozzi, N.; Nadalini, M.; Coticchio, G.; Zacà, C.; Lagalla, C.; Borini, A. The paternal toolbox for embryo development and health. Mol. Hum. Reprod. 2021, 27, gaab042. [Google Scholar] [CrossRef]
- Alves, M.B.R.; Celeghini, E.C.C.; Belleannée, C. From Sperm Motility to Sperm-Borne microRNA Signatures: New Approaches to Predict Male Fertility Potential. Front. Cell. Dev. Biol. 2020, 8, 791. [Google Scholar] [CrossRef]
- Ostermeier, G.C.; Dix, D.J.; Miller, D.; Khatri, P.; Krawetz, S.A. Spermatozoal RNA profiles of normal fertile men. Lancet 2002, 360, 9335. [Google Scholar] [CrossRef]
- Engel, M.L.; Chaboud, A.; Dumas, C.; McCormick, S. Sperm cells of Zea mays have a complex complement of mRNAs. T. Plant. J. 2003, 34, 697–707. [Google Scholar] [CrossRef] [Green Version]
- Ostermeier, G.C.; Miller, D.; Huntriss, J.D.; Diamond, M.P.; Krawetz, S.A. Reproductive biology: Delivering spermatozoan RNA to the oocyte. Nature 2004, 429, 6988. [Google Scholar] [CrossRef] [PubMed]
- Johnson, G.D.; Lalancette, C.; Linnemann, A.K.; Leduc, F.; Boissonneault, G.; Krawetz, S.A. The sperm nucleus: Chromatin, RNA, and the nuclear matrix. Reproduction 2011, 141, 21–36. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jodar, M.; Selvaraju, S.; Sendler, E.; Diamond, M.P.; Krawetz, S.A.; Reproductive Medicine Network. The presence, role and clinical use of spermatozoal RNAs. Hum. Reprod. Update 2013, 19, 604–624. [Google Scholar] [CrossRef] [PubMed]
- Ziyyat, A.; Lefèvre, A. Differential gene expression in pre-implantation embryos from mouse oocytes injected with round spermatids or spermatozoa. Hum. Reprod. 2001, 16, 1449–1456. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saunders, C.M.; Larman, M.G.; Parrington, J.; Cox, L.J.; Royse, J.; Blayney, L.M.; Swann, K.; Lai, F.A. PLC zeta: A sperm-specific trigger of Ca2+ oscillations in eggs and embryo development. Development 2002, 129, 3533–3544. [Google Scholar] [CrossRef]
- Hermo, L.; Oko, R.; Morales, C.R. Secretion and endocytosis in the male reproductive tract: A role in sperm maturation. Int. Rev. Cytol. 1994, 154, 106–189. [Google Scholar]
- Yuan, S.; Schuster, A.; Tang, C.; Yu, T.; Ortogero, N.; Bao, J.; Zheng, H.; Yan, W. Sperm-borne miRNAs and endo-siRNAs are important for fertilization and preimplantation embryonic development. Development 2016, 143, 635–647. [Google Scholar] [CrossRef] [Green Version]
- Shyu, A.B.; Wilkinson, M.F.; van Hoof, A. Messenger RNA regulation: To translate or to degrade. EMBO J. 2008, 27, 471–481. [Google Scholar] [CrossRef] [Green Version]
- Kotaja, N. MicroRNAs and spermatogenesis. Fertil. Steril. 2014, 101, 1552–1562. [Google Scholar] [CrossRef]
- Salas-Huetos, A.; Blanco, J.; Vidal, F.; Godo, A.; Grossmann, M.; Pons, M.C.; F-Fernández, S.; Garrido, N.; Anton, E. Spermatozoa from patients with seminal alterations exhibit a differential micro-ribonucleic acid profile. Fertil. Steril. 2015, 104, 591–601. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krawetz, S.A.; Kruger, A.; Lalancette, C.; Tagett, R.; Anton, E.; Draghici, S.; Diamond, M.P. A survey of small RNAs in human sperm. Hum. Reprod. 2011, 26, 3401–3412. [Google Scholar] [CrossRef] [Green Version]
- Krawetz, S.A.; Kruger, A.; Lalancette, C.; Tagett, R.; Anton, E.; Draghici, S.; Diamond, M.P. The effect of sperm miR-34c-5p on human embryonic development kinetics and clinical outcomes. Life Sci. 2020, 256, 117895. [Google Scholar] [CrossRef]
- Pantos, K.; Grigoriadis, S.; Tomara, P.; Louka, I.; Maziotis, E.; Pantou, A.; Nitsos, N.; Vaxevanoglou, T.; Kokkali, G.; Agarwal, A.; et al. Investigating the Role of the microRNA-34/449 Family in Male Infertility: A Critical Analysis and Review of the Literature. Front. Endocrinol. 2021, 12, 709943. [Google Scholar] [CrossRef]
- Xu, H.; Wang, X.; Wang, Z.; Li, J.; Xu, Z.; Miao, M.; Chen, G.; Lei, X.; Wu, J.; Shi, H.; et al. MicroRNA expression profile analysis in sperm reveals hsa-mir-191 as an auspicious omen of in vitro fertilization. BMC Genom. 2020, 21, 165. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mostafa, T.; Rashed, L.A.; Nabil, N.I.; Osman, I.; Mostafa, R.; Farag, M. Seminal miRNA Relationship with Apoptotic Markers and Oxidative Stress in Infertile Men with Varicocele. Biomed. Res. Int. 2016, 2016, 4302754. [Google Scholar] [CrossRef] [PubMed]
- Abu-Halima, M.; Backes, C.; Leidinger, P.; Keller, A.; Lubbad, A.M.; Hammadeh, M.; Meese, E. MicroRNA expression profiles in human testicular tissues of infertile men with different histopathologic patterns. Fertil. Steril. 2014, 101, 78–86.e2. [Google Scholar] [CrossRef] [PubMed]
- Finocchi, F.; Pelloni, M.; Balercia, G.; Pallotti, F.; Radicioni, A.F.; Lenzi, A.; Lombardo, F.; Paoli, D. Seminal plasma miRNAs in Klinefelter syndrome and in obstructive and non-obstructive azoospermia. Mol. Biol. Rep. 2020, 47, 4373–4382. [Google Scholar] [CrossRef]
- Momeni, A.; Najafipour, R.; Hamta, A.; Jahani, S.; Moghbelinejad, S. Expression and Methylation Pattern of hsa-miR-34 Family in Sperm Samples of Infertile Men. Reprod. Sci. 2020, 27, 301–308. [Google Scholar] [CrossRef]
- Wu, J.; Bao, J.; Kim, M.; Yuan, S.; Tang, C.; Zheng, H.; Mastick, G.S.; Xu, C.; Yan, W. Two miRNA clusters, miR-34b/c and miR-449, are essential for normal brain development, motile ciliogenesis, and spermatogenesis. Proc. Natl. Acad. Sci. USA 2014, 111, 28. [Google Scholar] [CrossRef] [PubMed]
- Conine, C.C.; Sun, F.; Song, L.; Rivera-Pérez, J.A.; Rando, O.J. Small RNAs Gained during Epididymal Transit of Sperm Are Essential for Embryonic Development in Mice. Dev. Cell 2018, 46, 470–480.e3. [Google Scholar] [CrossRef] [Green Version]
- Yeh, L.Y.; Lee, R.K.; Lin, M.H.; Huang, C.H.; Li, S.H. Correlation between Sperm Micro Ribonucleic Acid-34b and -34c Levels and Clinical Outcomes of Intracytoplasmic Sperm Injection in Men with Male Factor Infertility. Int. J. Mol. Sci. 2022, 23, 12381. [Google Scholar] [CrossRef] [PubMed]
- Lv, J.; Zhang, Z.; Pan, L.; Zhang, Y. MicroRNA-34/449 family and viral infections. Virus Res. 2019, 260, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Bao, J.; Li, D.; Wang, L.; Wu, J.; Hu, Y.; Wang, Z.; Chen, Y.; Cao, X.; Jiang, C.; Yan, W.; et al. MicroRNA-449 and microRNA-34b/c function redundantly in murine testes by targeting E2F transcription factor-retinoblastoma protein (E2F-pRb) pathway. J. Biol. Chem. 2012, 287, 21686–21698. [Google Scholar] [CrossRef] [Green Version]
- Comazzetto, S.; Di Giacomo, M.; Rasmussen, K.D.; Much, C.; Azzi, C.; Perlas, E.; Morgan, M.; O’Carroll, D. Oligoasthenoteratozoospermia and infertility in mice deficient for miR-34b/c and miR-449 loci. PLoS Genet. 2014, 10, e1004597. [Google Scholar] [CrossRef] [Green Version]
- Song, R.; Walentek, P.; Sponer, N.; Klimke, A.; Lee, J.S.; Dixon, G.; Harland, R.; Wan, Y.; Lishko, P.; Lize, M.; et al. miR-34/449 miRNAs are required for motile ciliogenesis by repressing cp110. Nature 2014, 510, 115–120. [Google Scholar] [CrossRef] [Green Version]
- Yuan, S.; Tang, C.; Zhang, Y.; Wu, J.; Bao, J.; Zheng, H.; Xu, C.; Yan, W. mir-34b/c and mir-449a/b/c are required for spermatogenesis, but not for the first cleavage division in mice. Biol. Open 2015, 4, 212–223. [Google Scholar] [CrossRef] [Green Version]
- Rah, H.; Chung, K.W.; Ko, K.H.; Kim, E.S.; Kim, J.O.; Sakong, J.H.; Kim, J.H.; Lee, W.S.; Kim, N.K. miR-27a and miR-449b-5p polymorphisms associated with a risk of idiopathic recurrent pregnancy loss. PLoS ONE 2017, 12, e0177160. [Google Scholar] [CrossRef] [Green Version]
- Bommer, G.T.; Gerin, I.; Feng, Y.; Kaczorowski, A.J.; Kuick, R.; Love, R.E.; Zhai, Y.; Giordano, T.J.; Qin, Z.S.; Moore, B.B.; et al. p53-mediated activation of miRNA34 candidate tumor-suppressor genes. Curr. Biol. 2007, 17, 1298–1307. [Google Scholar] [CrossRef] [Green Version]
- Hermeking, H. The miR-34 family in cancer and apoptosis. Cell Death Differ. 2010, 17, 193–199. [Google Scholar] [CrossRef]
- Lizé, M.; Klimke, A.; Dobbelstein, M. MicroRNA-449 in cell fate determination. Cell Cycle 2011, 10, 2874–2882. [Google Scholar] [CrossRef] [Green Version]
- World Health Organization. WHO Laboratory Manual for the Examination and Processing of Human Semen, 5th ed.; World Health Organization: Geneva, Switzerland, 2010. [Google Scholar]
- Rienzi, L.; Vajta, G.; Ubaldi, F. Predictive value of oocyte morphology in human IVF: A systematic review of the literature. Hum. Reprod. Update 2011, 17, 34–45. [Google Scholar] [CrossRef] [Green Version]
- Alpha Scientists in Reproductive Medicine and ESHRE Special Interest Group of Embryology. The Istanbul consensus workshop on embryo assessment: Proceedings of an expert meeting. Hum. Reprod. 2011, 26, 1270–1283. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paoli, D.; Pecora, G.; Pallotti, F.; Faja, F.; Pelloni, M.; Lenzi, A.; Lombardo, F. Cytological and molecular aspects of the ageing sperm. Hum. Reprod. 2019, 34, 218–227. [Google Scholar] [CrossRef] [PubMed]
- Paoli, D.; Pelloni, M.; Gallo, M.; Coltrinari, G.; Lombardo, F.; Lenzi, A.; Gandini, L. Sperm glyceraldehyde 3-phosphate dehydrogenase gene expression in asthenozoospermic spermatozoa. Asian. J. Androl. 2017, 19, 409–413. [Google Scholar] [CrossRef] [PubMed]
- Deluao, J.C.; Winstanley, Y.; Robker, R.L.; Pacella-Ince, L.; Gonzalez, M.B.; McPherson, N.O. OXIDATIVE STRESS AND REPRODUCTIVE FUNCTION: Reactive oxygen species in the mammalian pre-implantation embryo. Reproduction 2022, 164, F95–F108. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Age | BMI | Smokers | AMH [ng/mL] | |
|---|---|---|---|---|
| ♀ | 37.0 ± 3.8 37.0 | 22.5 ± 3.6 22.1 | 26/106 (24.5%) | 2.0 ± 1.6 1.6 |
| ♂ | 41.0 ± 6.0 40.0 | 25.7 ± 3.1 25.8 | 33/106 (31.1%) | // |
| Sperm Concentration (×106/mL) | Progressive Motility (%) | Abnormal Form (%) | SDF (%) | Below 5th Percentile WHO 2010 | |
|---|---|---|---|---|---|
| BASELINE 106 pz | 50.9 ± 40.9 40.0 | 39.9 ± 16.5 45.0 | 90.8 ± 3.8 90.0 | 15.5 ± 8.8 14.4 | 22/106 (20.8%) |
| POST DGC 30 pz | 18.9 ± 17.1 13.0 | 68.2 ± 32.9 90.0 | 51.0 ± 21.0 46.5 | 11.1 ± 10.8 a 6.2 | // |
| POST SW 76 pz | 29.9 ± 19.5 30.0 | 89.4 ± 10.2 90.0 | 42.4 ± 10.7 40.0 | 4.5 ± 6.1 a 2.5 | // |
| (a) | ||||
| Pre Sperm Selection Procedures | ||||
| Conc./mL | Progressive Motility (%) | Abnormal Form (%) | ||
| SDF PRE sperm selection procedures | Spearman’s ρ p value | −0.413 <0.001 | −0.508 <0.001 | 0.416 <0.001 |
| (b) | ||||
| Post Sperm Selection Procedures | ||||
| Conc./mL | Progressive Motility (%) | Abnormal Form (%) | ||
| SDF PRE sperm selection procedures | Spearman’s ρ p value | −0.364 <0.001 | −0.223 0.036 | 0.365 <0.001 |
| Sperm Concentration (×106/mL) | Progressive Motility (%) | Abnormal Forms (%) | SDF (%) | Age ♀ | Age ♂ | BMI ♀ | BMI ♂ | |
|---|---|---|---|---|---|---|---|---|
| Fertilization | 26.5 ± 19.5 20.0 | 85.1 ± 18.8 90.0 | 43.8 ± 13.2 40.0 | 5.2 ± 5.8 2.9 | 36.8 ± 3.8 37.0 | 40.5 ± 6.3 40.0 | 22.4 ± 3.6 22.1 | 25.5 ± 3.1 22.5 |
| NO Fertilization | 29.3 ± 19.5 36.0 | 57.1 ± 39.9 80.0 | 60.0 ± 26.8 52.0 | 10.1 ± 11.0 5.6 | 40.3 ± 2.4 41.0 | 48.1 ± 6.7 47.0 | 23.1 ± 3.8 23.5 | 28.7 ± 3.0 29.3 |
| p value | 0.760 | 0.002 | 0.073 | 0.380 | 0.015 | 0.005 | 0.541 | 0.015 |
| Number of Inseminated Oocytes | Number of Fertilised Oocytes | Low-Quality Embryos Ratio * | Viable Embryo | ||
|---|---|---|---|---|---|
| SDF% pre sperm selection procedures | Spearman’ρ p value | −0.043 0.706 | −0.101 0.378 | 0.094 0.472 | −0.132 0.245 |
| SDF% post sperm selection procedures | Spearman’ρ p value | −0.081 0.483 | 0.020 0.865 | 0.230 0.074 | −0.238 0.036 |
| Coeff. | 95% CI | p Value | |
|---|---|---|---|
| SDF pre sperm selection procedures (%) | 0.881 | −0.337–2.099 | 0.153 |
| SDF post sperm selection procedures (%) | −0.742 | −1.292–−0.193 | 0.009 |
| Age ♀ | −0.026 | −0.093–0.040 | 0.430 |
| Age ♂ | 0.026 | −0.016–0.068 | 0.223 |
| Selection technique (DCG vs. SW) | 0.217 | −0.303–0.737 | 0.407 |
| AMH [ng/mL] | 0.163 | 0.026–0.301 | 0.021 |
| OR | 95% CI | SE | p Value | |
|---|---|---|---|---|
| SDF > 14.4% Pre-sperm selection | 1.099 | 0.313–3.852 | 0.640 | 0.883 |
| SDF > 2.9% Post-sperm selection | 3.955 | 1.088–14.379 | 0.659 | 0.037 |
| AMH | 1.143 | 0.777–1.683 | 0.197 | 0.497 |
| Age ♀ | 1.086 | 0.884–1.333 | 0.105 | 0.433 |
| Age ♂ | 1.022 | 0.909–1.149 | 0.060 | 0.714 |
| BMI ♀ | 0.933 | 0.772–1.129 | 0.097 | 0.477 |
| BMI ♂ | 1.012 | 0.827–1.238 | 0.103 | 0.908 |
| Selection technique (DCG vs. SW) | 0.438 | 0.087–2.198 | 0.823 | 0.316 |
| Age | BMI | Smokers [%] | AMH [ng/mL] | |
|---|---|---|---|---|
| ♀ | 36.3 ± 4.6 37.0 | 22.5 ± 3.4 22.2 | 9/38 (23.7%) | 2.2 ± 1.7 1.9 |
| ♂ | 39.8 ± 5.2 40.0 | 25.5 ± 3.5 25.3 | 14/38 (36.8%) | / |
| Sperm Concentration (×106/mL) | Progressive Motility (%) | Abnormal Form (%) | SDF (%) | miR-34c-5p [Copies/µL] | miR-449b-5p [Copies/µL] | |
|---|---|---|---|---|---|---|
| POST sperm selection | 33.1 ± 17.9 37.0 | 90.4 ± 1.8 90.0 | 41.7 ± 10.3 40.0 | 5.0 ± 6.8 2.9 | 1.8 ± 2.7 0.7 | 3.8 ± 5.5 1.6 |
| Sperm Concentration(×106/mL) | Progressive Motility (%) | Abnormal Form (%) | SDF (%) | miR-34c-5p [copies/µL] | miR-449b-5p [copies/µL] | |
|---|---|---|---|---|---|---|
| miR-34c-5p | 0.468 0.003 | 0.099 0.556 | 0.218 0.188 | 0.274 0.117 | // | 0.812 <0.001 |
| miR-449b-5p | 0.514 <0.001 | 0.078 0.652 | 0.235 0.167 | 0.362 0.035 | 0.812 <0.001 | // |
| OR | 95% CI | SE | p Value | |
|---|---|---|---|---|
| miR-34c-5p | 14.266 | 1.328–153.221 | 1.211 | 0.028 |
| miR-449b-5p | 0.038 | 0.002–0.847 | 1.580 | 0.039 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Conflitti, A.C.; Cicolani, G.; Buonacquisto, A.; Pallotti, F.; Faja, F.; Bianchini, S.; Blaconà, G.; Bruno, S.M.; Linari, A.; Lucarelli, M.; et al. Sperm DNA Fragmentation and Sperm-Borne miRNAs: Molecular Biomarkers of Embryo Development? Int. J. Mol. Sci. 2023, 24, 1007. https://doi.org/10.3390/ijms24021007
Conflitti AC, Cicolani G, Buonacquisto A, Pallotti F, Faja F, Bianchini S, Blaconà G, Bruno SM, Linari A, Lucarelli M, et al. Sperm DNA Fragmentation and Sperm-Borne miRNAs: Molecular Biomarkers of Embryo Development? International Journal of Molecular Sciences. 2023; 24(2):1007. https://doi.org/10.3390/ijms24021007
Chicago/Turabian StyleConflitti, Anna Chiara, Gaia Cicolani, Alessandra Buonacquisto, Francesco Pallotti, Fabiana Faja, Serena Bianchini, Giovanna Blaconà, Sabina Maria Bruno, Antonella Linari, Marco Lucarelli, and et al. 2023. "Sperm DNA Fragmentation and Sperm-Borne miRNAs: Molecular Biomarkers of Embryo Development?" International Journal of Molecular Sciences 24, no. 2: 1007. https://doi.org/10.3390/ijms24021007