
Obesity Modifies the Proteomic Profile of the Periodontal Ligament

 , , , , and
, , , , and
Abstract
:1. Introduction
2. Results
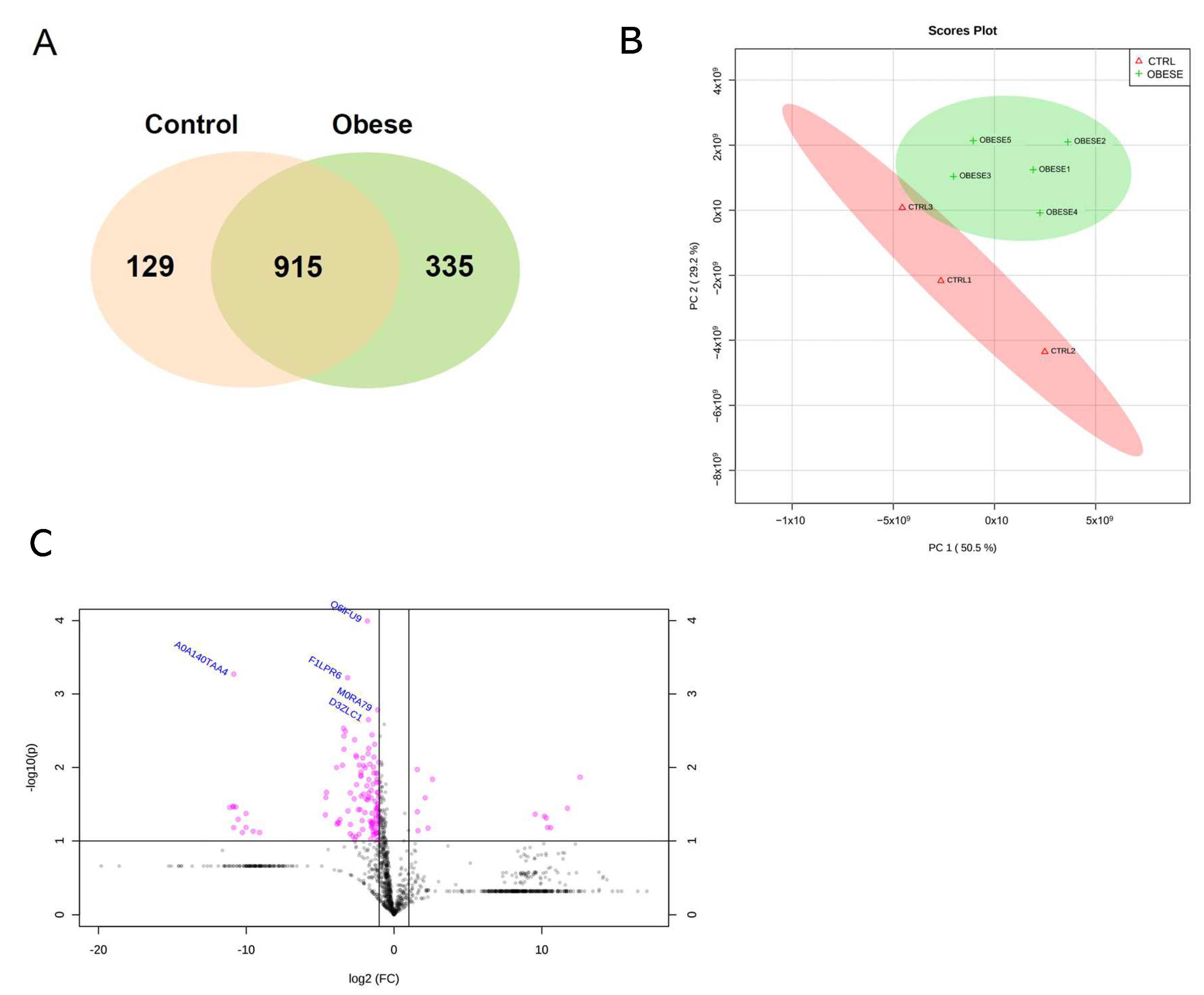
2.1. General Proteomic Profile of PDL in the Control and Obese Groups
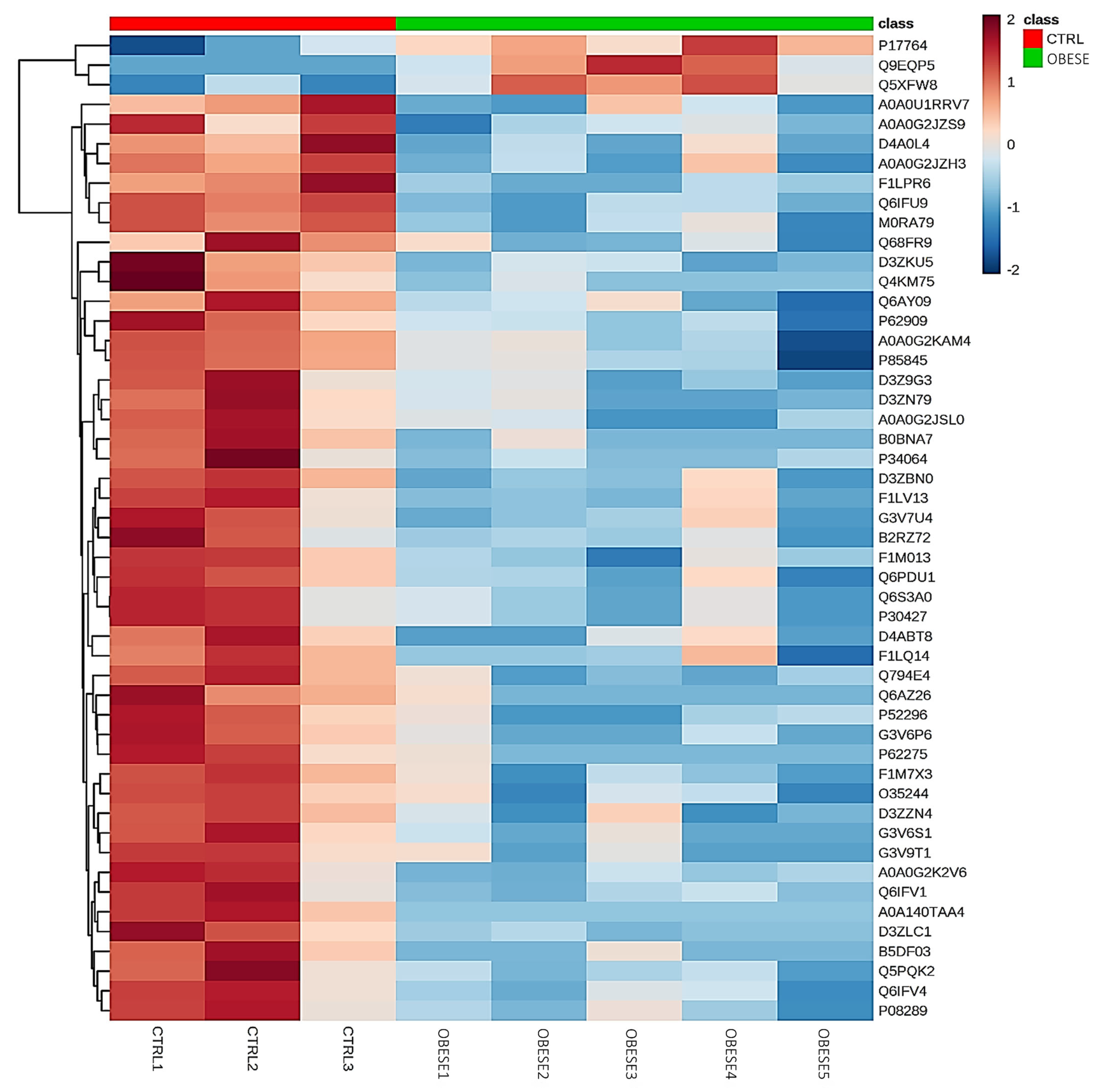
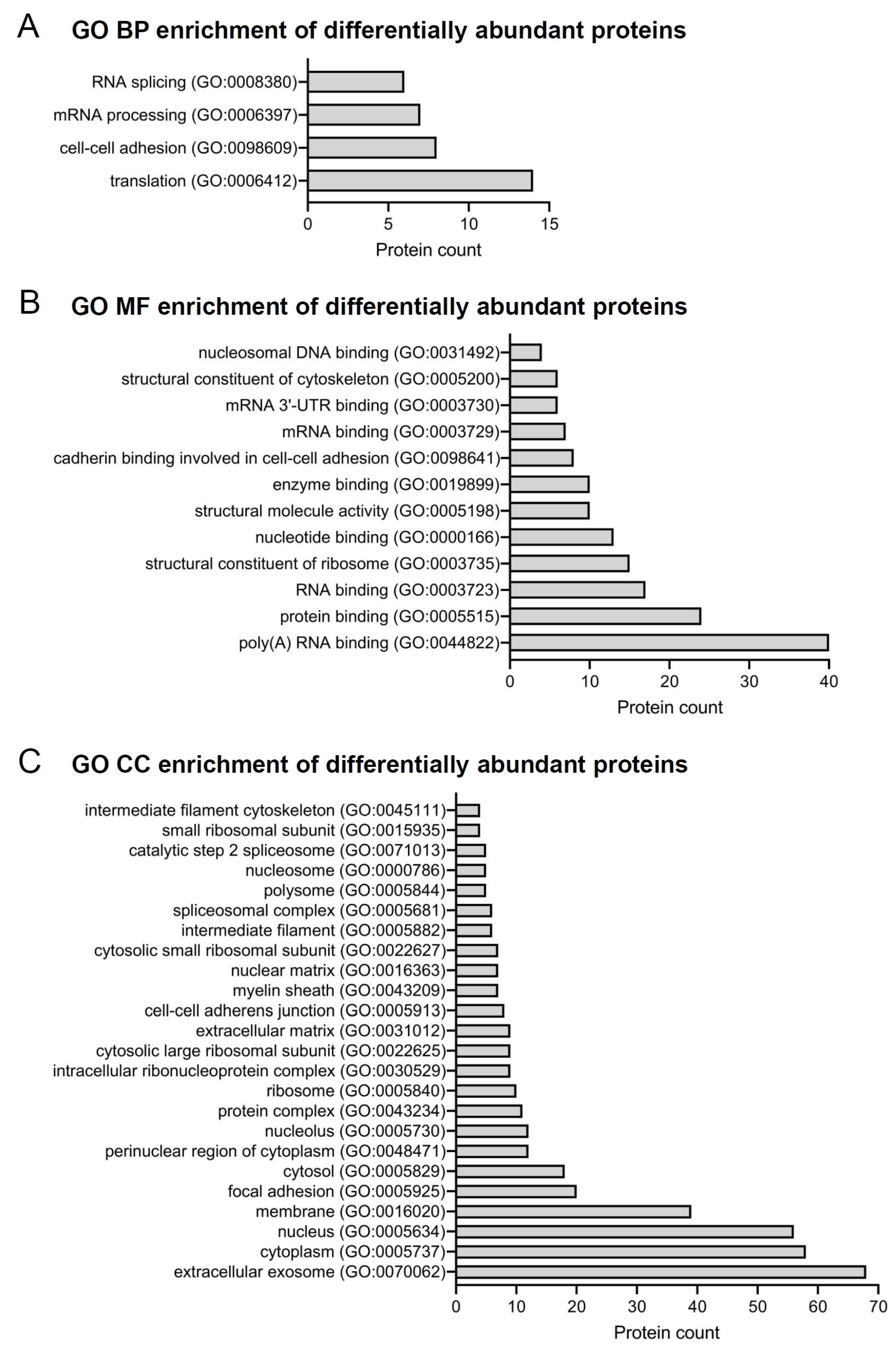
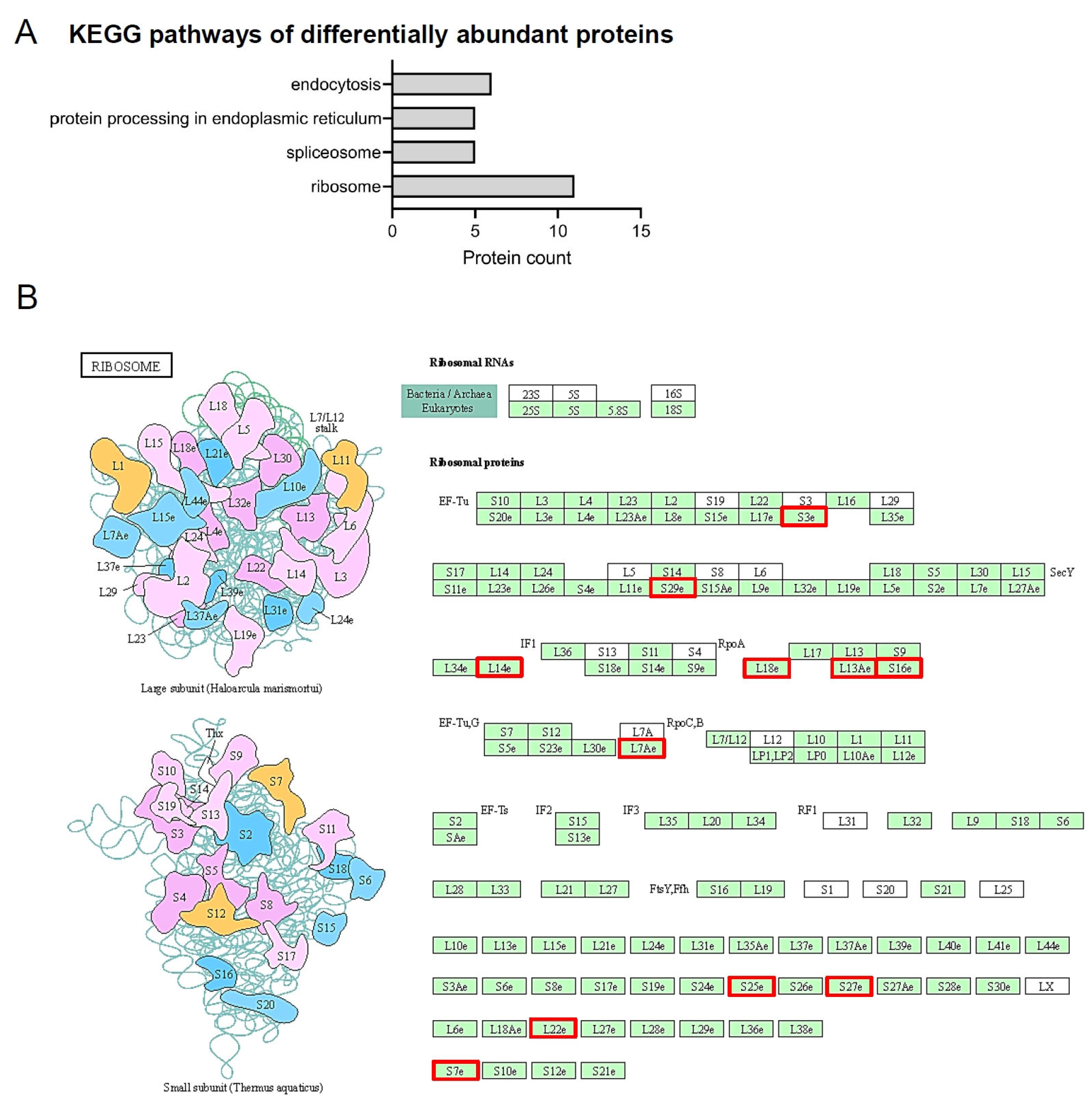
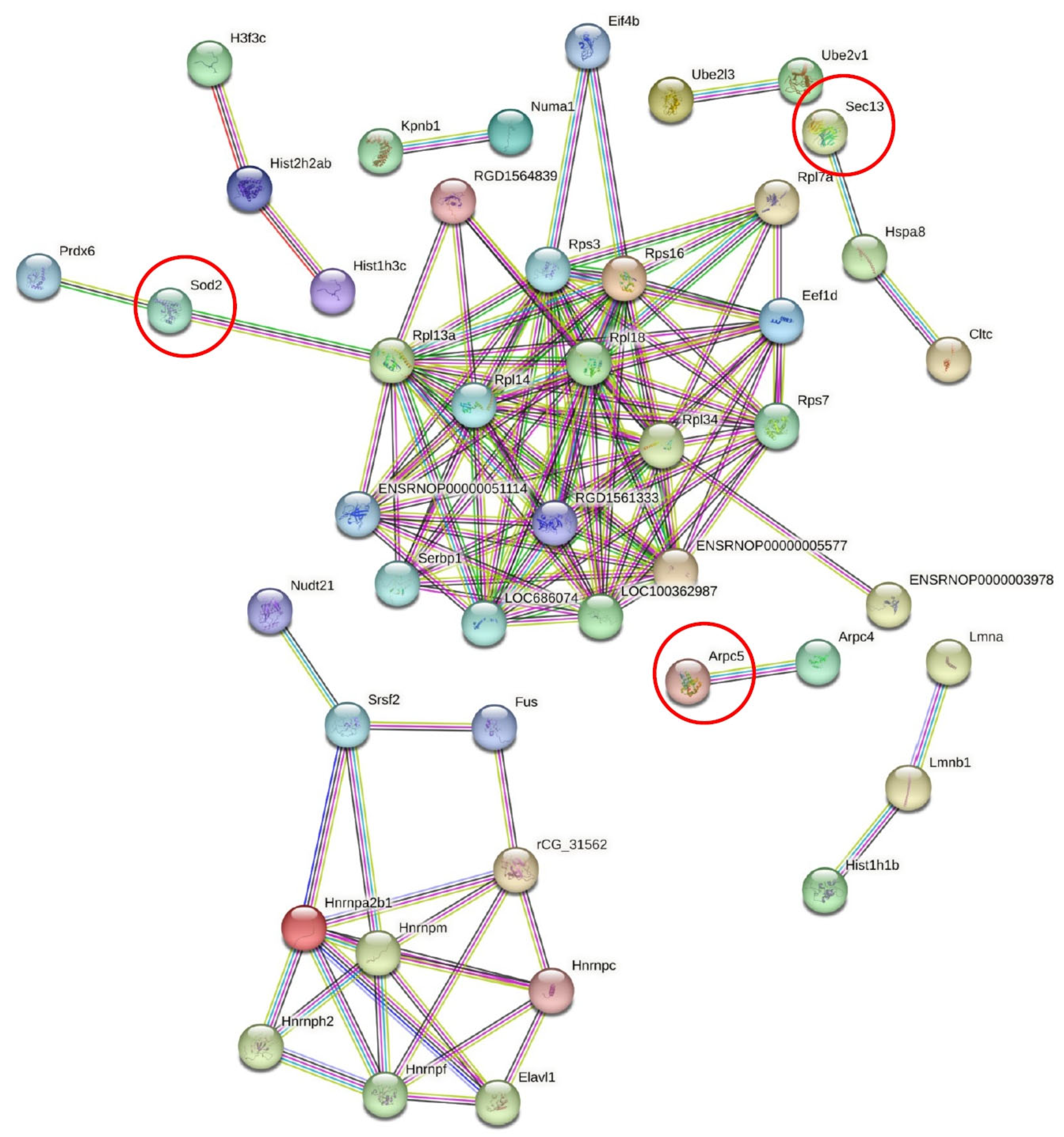
2.2. Differentially Identified Proteins in PDL
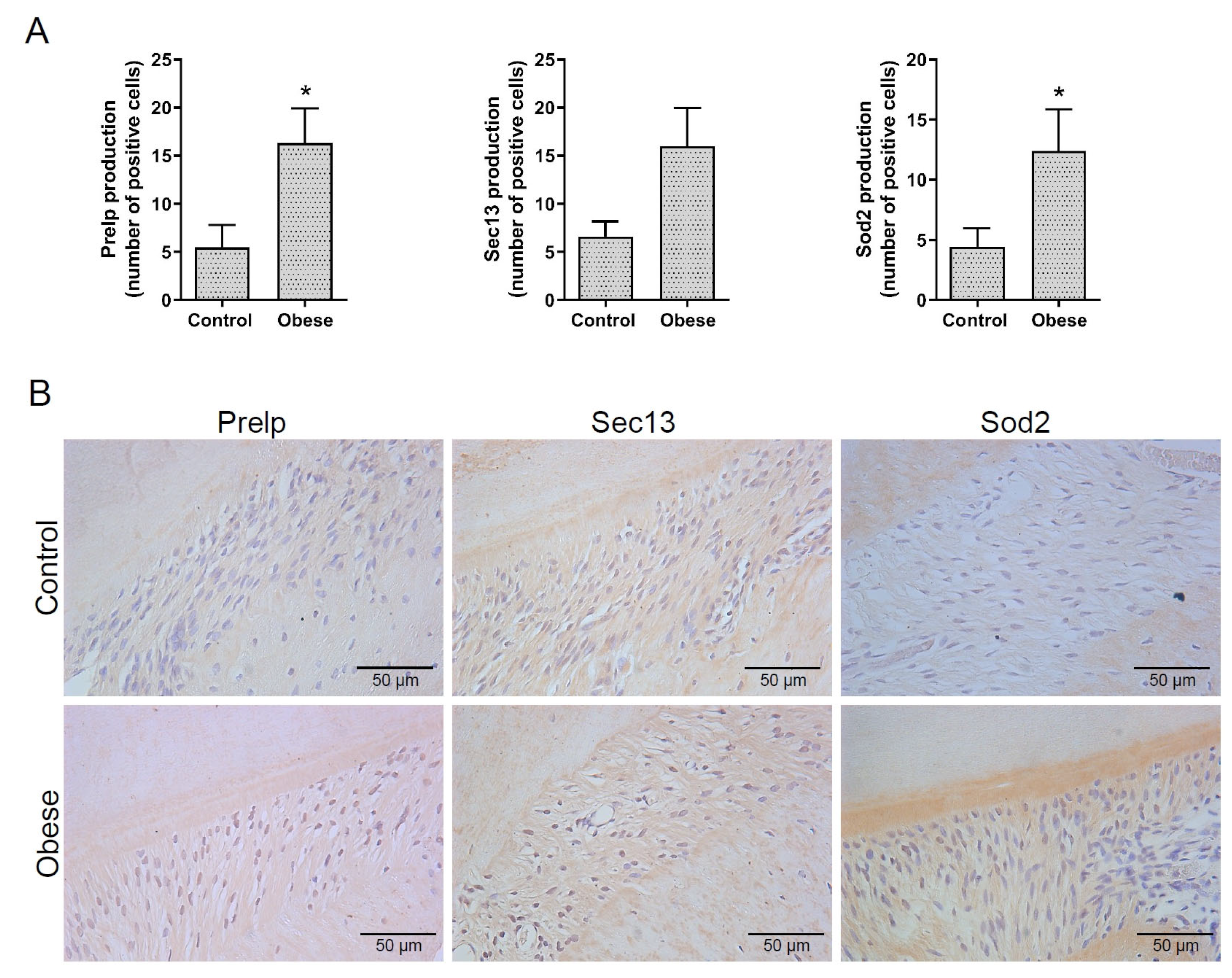
2.3. Immunolocalization of the Selected Proteins Prelp, Sec13, and Sod2 in PDL
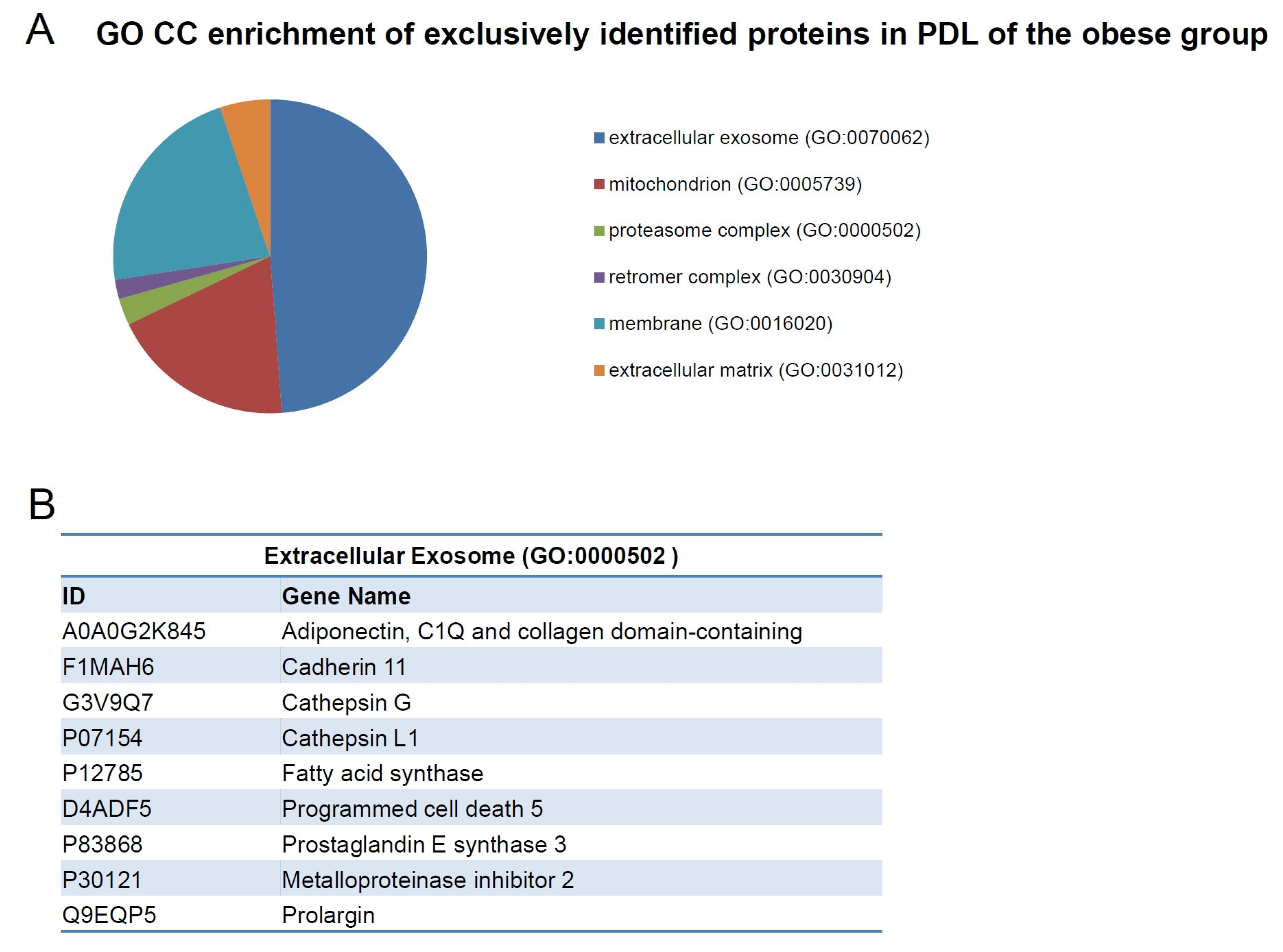
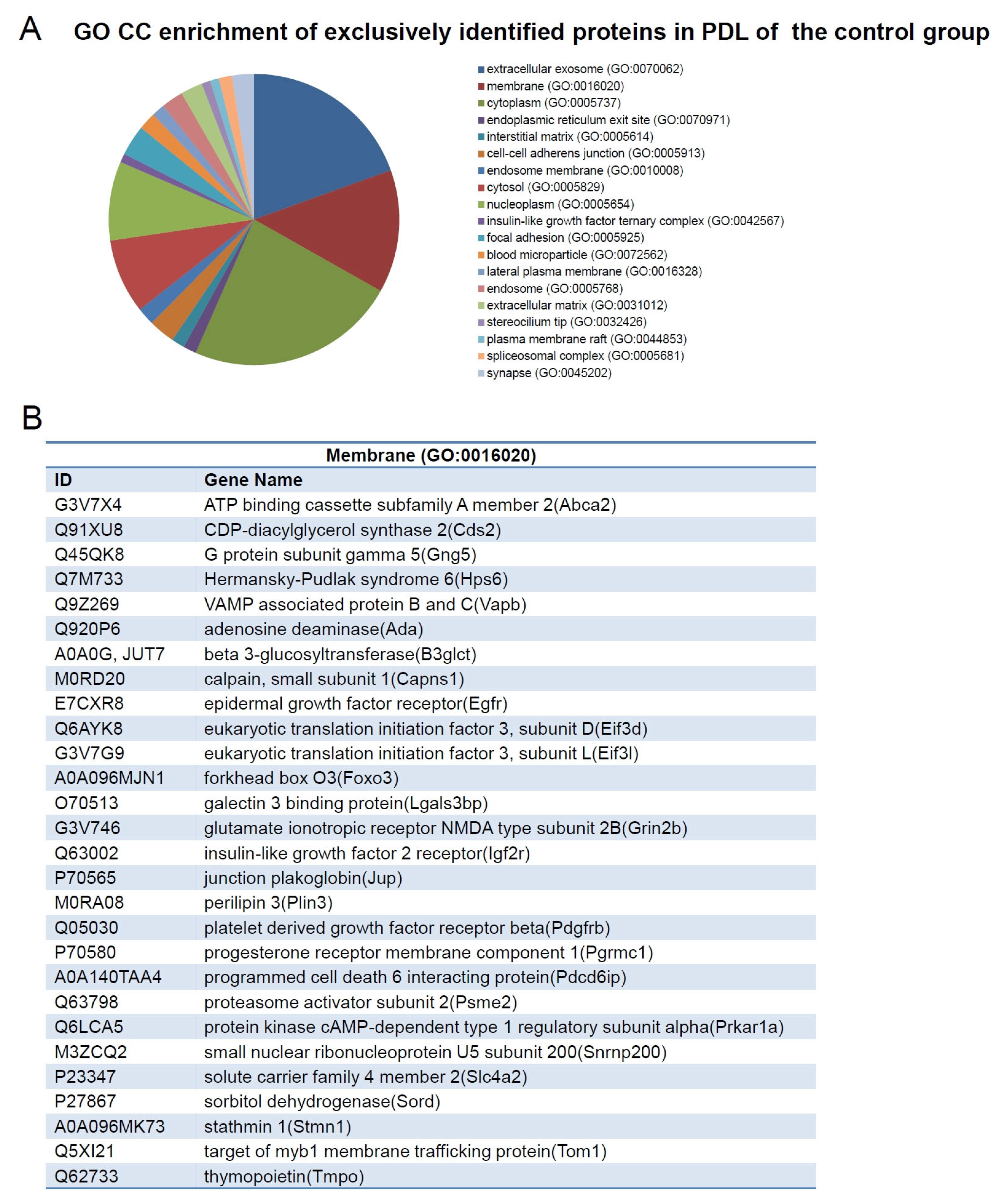
2.4. Exclusively Identified Proteins in PDL
3. Discussion
4. Materials and Methods
4.1. Experimental Protocol
4.2. Laser Capture Microdissection (LCM) and Protein Extraction
4.3. LC-MS/MS and Bioinformatics Analysis
4.4. IHC
4.5. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Jepsen, S.; Suvan, J.; Deschner, J. The Association of Periodontal Diseases with Metabolic Syndrome and Obesity. Periodontol. 2000 2020, 83, 125–153. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization. Global Status Report on Non-Communicable Diseases 2014; World Health Organization: Geneva, Switzerland, 2014. [Google Scholar]
- World Health Organization. Obesity: Preventing and Managing the Global Epidemic. Report of a WHO Consultation; World Health Organization: Geneva, Switzerland, 2000; Volume 894, pp. 1–253. [Google Scholar]
- World Health Organization. Obesity and Overweight. Available online: https://www.who.int/news-room/fact-sheets/detail/obesity-and-overweight (accessed on 1 October 2022).
- World Health Organization. Global Database on Body Mass Index—An Interactive Surveillance Tool for Monitoring Nutrition Transition; World Health Organization: Geneva, Switzerland, 2009. [Google Scholar]
- Blüher, M. Obesity: Global Epidemiology and Pathogenesis. Nat. Rev. Endocrinol. 2019, 15, 288–298. [Google Scholar] [CrossRef] [PubMed]
- Heymsfield, S.B.; Wadden, T.A. Mechanisms, Pathophysiology, and Management of Obesity. N. Engl. J. Med. 2017, 376, 254–266. [Google Scholar] [CrossRef] [PubMed]
- Arroyo-Johnson, C.; Mincey, K.D. Obesity Epidemiology Worldwide. Gastroenterol. Clin. North Am. 2016, 45, 571–579. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jepsen, S.; Caton, J.G.; Albandar, J.M.; Bissada, N.F.; Bouchard, P.; Cortellini, P.; Demirel, K.; de Sanctis, M.; Ercoli, C.; Fan, J.; et al. Periodontal Manifestations of Systemic Diseases and Developmental and Acquired Conditions: Consensus Report of Workgroup 3 of the 2017 World Workshop on the Classification of Periodontal and Peri-Implant Diseases and Conditions. J. Periodontol. 2018, 89 (Suppl. 1), S237–S248. [Google Scholar] [CrossRef]
- Nogueira, A.V.B.; Nokhbehsaim, M.; Eick, S.; Bourauel, C.; Jäger, A.; Jepsen, S.; Cirelli, J.A.; Deschner, J. Regulation of Visfatin by Microbial and Biomechanical Signals in PDL Cells. Clin. Oral Investig. 2014, 18, 171–178. [Google Scholar] [CrossRef]
- Nogueira, A.V.B.; Nokhbehsaim, M.; Tekin, S.; de Molon, R.S.; Spolidorio, L.C.; Memmert, S.; Damanaki, A.; Jäger, A.; Eick, S.; Deschner, J.; et al. Resistin Is Increased in Periodontal Cells and Tissues: In Vitro and In Vivo Studies. Mediat. Inflamm. 2020, 2020, 9817095. [Google Scholar] [CrossRef]
- Nokhbehsaim, M.; Keser, S.; Nogueira, A.V.B.; Jäger, A.; Jepsen, S.; Cirelli, J.A.; Bourauel, C.; Eick, S.; Deschner, J. Leptin Effects on the Regenerative Capacity of Human Periodontal Cells. Int. J. Endocrinol. 2014, 2014, 180304. [Google Scholar] [CrossRef] [Green Version]
- Nokhbehsaim, M.; Keser, S.; Nogueira, A.V.B.; Cirelli, J.A.; Jepsen, S.; Jäger, A.; Eick, S.; Deschner, J. Beneficial Effects of Adiponectin on Periodontal Ligament Cells under Normal and Regenerative Conditions. J. Diabetes Res. 2014, 2014, 796565. [Google Scholar] [CrossRef] [Green Version]
- de Jong, T.; Bakker, A.D.; Everts, V.; Smit, T.H. The Intricate Anatomy of the Periodontal Ligament and Its Development: Lessons for Periodontal Regeneration. J. Periodontal Res. 2017, 52, 965–974. [Google Scholar] [CrossRef]
- Connizzo, B.K.; Sun, L.; Lacin, N.; Gendelman, A.; Solomonov, I.; Sagi, I.; Grodzinsky, A.J.; Naveh, G.R.S. Nonuniformity in Periodontal Ligament: Mechanics and Matrix Composition. J. Dent. Res. 2021, 100, 179–186. [Google Scholar] [CrossRef]
- Liu, J.; Ruan, J.; Weir, M.D.; Ren, K.; Schneider, A.; Wang, P.; Oates, T.W.; Chang, X.; Xu, H.H.K. Periodontal Bone-Ligament-Cementum Regeneration via Scaffolds and Stem Cells. Cells 2019, 8, 537. [Google Scholar] [CrossRef] [Green Version]
- Mangum, J.E.; Veith, P.D.; Reynolds, E.C.; Hubbard, M.J. Towards Second-Generation Proteome Analysis of Murine Enamel-Forming Cells. Eur. J. Oral Sci. 2006, 114 (Suppl. 1), 259–265, discussion 285–286, 382. [Google Scholar] [CrossRef]
- Jágr, M.; Eckhardt, A.; Pataridis, S.; Foltán, R.; Myšák, J.; Mikšík, I. Proteomic Analysis of Human Tooth Pulp Proteomes—Comparison of Caries-Resistant and Caries-Susceptible Persons. J. Proteom. 2016, 145, 127–136. [Google Scholar] [CrossRef]
- Eckhard, U.; Marino, G.; Abbey, S.R.; Matthew, I.; Overall, C.M. TAILS N-Terminomic and Proteomic Datasets of Healthy Human Dental Pulp. Data Br. 2015, 5, 542–548. [Google Scholar] [CrossRef] [Green Version]
- Giovani, P.A.; Martins, L.; Salmon, C.R.; Mofatto, L.S.; Leme, A.F.P.; Puppin-Rontani, R.M.; Kolli, T.N.; Foster, B.L.; Nociti, F.H.; Kantovitz, K.R. Comparative Proteomic Analysis of Dental Cementum from Deciduous and Permanent Teeth. J. Periodontal Res. 2021, 56, 173–185. [Google Scholar] [CrossRef]
- Salmon, C.R.; Giorgetti, A.P.O.; Paes Leme, A.F.; Domingues, R.R.; Sallum, E.A.; Alves, M.C.; Kolli, T.N.; Foster, B.L.; Nociti, F.H. Global Proteome Profiling of Dental Cementum under Experimentally-Induced Apposition. J. Proteom. 2016, 141, 12–23. [Google Scholar] [CrossRef] [Green Version]
- Salmon, C.R.; Tomazela, D.M.; Ruiz, K.G.S.; Foster, B.L.; Paes Leme, A.F.; Sallum, E.A.; Somerman, M.J.; Nociti, F.H. Proteomic Analysis of Human Dental Cementum and Alveolar Bone. J. Proteom. 2013, 91, 544–555. [Google Scholar] [CrossRef] [Green Version]
- Lopes, M.E.S.; Marcantonio, C.C.; de Molon, R.S.; Cerri, P.S.; Salmon, C.R.; Mofatto, L.S.; Nociti Junior, F.H.; Deschner, J.; Cirelli, J.A.; Nogueira, A.V.B. Obesity Influences the Proteome of Periodontal Ligament Tissues Following Periodontitis Induction in Rats. J. Periodontal Res. 2022, 57, 545–557. [Google Scholar] [CrossRef]
- Marcantonio, C.C.; Lopes, M.E.S.; Mofatto, L.S.; Salmon, C.R.; Deschner, J.; Nociti-Junior, F.H.; Cirelli, J.A.; Nogueira, A.V.B. Obesity Affects the Proteome Profile of Periodontal Ligament Submitted to Mechanical Forces Induced by Orthodontic Tooth Movement in Rats. J. Proteom. 2022, 263, 104616. [Google Scholar] [CrossRef]
- Giovani, P.A.; Salmon, C.R.; Martins, L.; Leme, A.F.P.; Puppin-Rontani, R.M.; Mofatto, L.S.; Nociti, F.H.; Kantovitz, K.R. Membrane Proteome Characterization of Periodontal Ligament Cell Sets from Deciduous and Permanent Teeth. J. Periodontol. 2018, 90, 775–787. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Luo, T.; Lu, W.; Yi, X.; Zhao, Z.; Liu, J. Proteomic Analysis of Human Periodontal Ligament Cells under Hypoxia. Proteome Sci. 2019, 17, 3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Onyedibe, K.I.; Elmanfi, S.; Aryal, U.K.; Könönen, E.; Gürsoy, U.K.; Sintim, H.O. Global Proteomics of Fibroblast Cells Treated with Bacterial Cyclic Dinucleotides, c-Di-GMP and c-Di-AMP. J. Oral Microbiol. 2022, 14, 2003617. [Google Scholar] [CrossRef] [PubMed]
- Bao, K.; Li, X.; Kajikawa, T.; Toshiharu, A.; Selevsek, N.; Grossmann, J.; Hajishengallis, G.; Bostanci, N. Pressure Cycling Technology Assisted Mass Spectrometric Quantification of Gingival Tissue Reveals Proteome Dynamics during the Initiation and Progression of Inflammatory Periodontal Disease. Proteomics 2020, 20, e1900253. [Google Scholar] [CrossRef] [PubMed]
- Yaprak, E.; Kasap, M.; Akpınar, G.; Kayaaltı-Yüksek, S.; Sinanoğlu, A.; Guzel, N.; Demirturk Kocasarac, H. The Prominent Proteins Expressed in Healthy Gingiva: A Pilot Exploratory Tissue Proteomics Study. Odontology 2018, 106, 19–28. [Google Scholar] [CrossRef]
- Oliveira, B.P.; Buzalaf, M.A.R.; Silva, N.C.; Ventura, T.M.O.; Toniolo, J.; Rodrigues, J.A. Saliva Proteomic Profile of Early Childhood Caries and Caries-Free Children. Acta Odontol. Scand. 2022, 1–11, Online ahead of print. [Google Scholar] [CrossRef]
- Tsuchida, S.; Satoh, M.; Takiwaki, M.; Nomura, F. Current Status of Proteomic Technologies for Discovering and Identifying Gingival Crevicular Fluid Biomarkers for Periodontal Disease. Int. J. Mol. Sci. 2018, 20, 86. [Google Scholar] [CrossRef] [Green Version]
- Lackey, D.E.; Olefsky, J.M. Regulation of Metabolism by the Innate Immune System. Nat. Rev. Endocrinol. 2016, 12, 15–28. [Google Scholar] [CrossRef]
- Debnath, M.; Agrawal, S.; Agrawal, A.; Dubey, G.P. Metaflammatory Responses during Obesity: Pathomechanism and Treatment. Obes. Res. Clin. Pract. 2016, 10, 103–113. [Google Scholar] [CrossRef]
- Chaffee, B.W.; Weston, S.J. Association between Chronic Periodontal Disease and Obesity: A Systematic Review and Meta-Analysis. J. Periodontol. 2010, 81, 1708–1724. [Google Scholar] [CrossRef]
- Xu, J.; Chang, X.; Zhang, H.; Si, M.; Su, H.; Cao, L.; Li, Y.; Zhai, Y. Exosomes: A Bridge of Periodontitis and Systemic Diseases. Gene Protein Dis. 2022, 1, 99. [Google Scholar] [CrossRef]
- Pardo, F.; Villalobos-Labra, R.; Sobrevia, B.; Toledo, F.; Sobrevia, L. Extracellular Vesicles in Obesity and Diabetes Mellitus. Mol. Asp. Med. 2018, 60, 81–91. [Google Scholar] [CrossRef] [PubMed]
- Gaye, A.; Doumatey, A.P.; Davis, S.K.; Rotimi, C.N.; Gibbons, G.H. Whole-Genome Transcriptomic Insights into Protective Molecular Mechanisms in Metabolically Healthy Obese African Americans. NPJ Genom. Med. 2018, 3, 4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ghosh, S.; Dent, R.; Harper, M.-E.; Gorman, S.A.; Stuart, J.S.; McPherson, R. Gene Expression Profiling in Whole Blood Identifies Distinct Biological Pathways Associated with Obesity. BMC Med. Genom. 2010, 3, 56. [Google Scholar] [CrossRef] [Green Version]
- Campbell, L.E.; Langlais, P.R.; Day, S.E.; Coletta, R.L.; Benjamin, T.R.; De Filippis, E.A.; Madura, J.A.; Mandarino, L.J.; Roust, L.R.; Coletta, D.K. Identification of Novel Changes in Human Skeletal Muscle Proteome After Roux-En-Y Gastric Bypass Surgery. Diabetes 2016, 65, 2724–2731. [Google Scholar] [CrossRef] [Green Version]
- Lu, Z.; Meng, L.; Sun, Z.; Shi, X.; Shao, W.; Zheng, Y.; Yao, X.; Song, J. Differentially Expressed Genes and Enriched Signaling Pathways in the Adipose Tissue of Obese People. Front. Genet. 2021, 12, 620740. [Google Scholar] [CrossRef]
- van der Kolk, B.W.; Saari, S.; Lovric, A.; Arif, M.; Alvarez, M.; Ko, A.; Miao, Z.; Sahebekhtiari, N.; Muniandy, M.; Heinonen, S.; et al. Molecular Pathways behind Acquired Obesity: Adipose Tissue and Skeletal Muscle Multiomics in Monozygotic Twin Pairs Discordant for BMI. Cell Rep. Med. 2021, 2, 100226. [Google Scholar] [CrossRef]
- Rucci, N.; Rufo, A.; Alamanou, M.; Capulli, M.; Del Fattore, A.; Åhrman, E.; Capece, D.; Iansante, V.; Zazzeroni, F.; Alesse, E.; et al. The Glycosaminoglycan-Binding Domain of PRELP Acts as a Cell Type–Specific NF-ΚB Inhibitor That Impairs Osteoclastogenesis. J. Cell Biol. 2009, 187, 669–683. [Google Scholar] [CrossRef] [Green Version]
- Heinegård, D.; Aspberg, A.; Franzén, A.; Lorenzo, P. Glycosylated Matrix Proteins. In Connective Tissue and Its Heritable Disorders; Royce, P.M., Steinmann, B., Eds.; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2002; pp. 271–291. [Google Scholar]
- Grover, J.; Chen, X.-N.; Korenberg, J.R.; Recklies, A.D.; Roughley, P.J. The Gene Organization, Chromosome Location, and Expression of a 55-KDa Matrix Protein (PRELP) of Human Articular Cartilage. Genomics 1996, 38, 109–117. [Google Scholar] [CrossRef]
- Heinegård, D.; Larsson, T.; Sommarin, Y.; Franzén, A.; Paulsson, M.; Hedbom, E. Two Novel Matrix Proteins Isolated from Articular Cartilage Show Wide Distributions among Connective Tissues. J. Biol. Chem. 1986, 261, 13866–13872. [Google Scholar] [CrossRef]
- Tsuru, M.; Soejima, T.; Shiba, N.; Kimura, K.; Sato, K.; Toyama, Y.; Nagata, K. Proline/Arginine-Rich End Leucine-Rich Repeat Protein Converts Stem Cells to Ligament Tissue and Zn(II) Influences Its Nuclear Expression. Stem Cells Dev. 2013, 22, 2057–2070. [Google Scholar] [CrossRef]
- Rucci, N.; Capulli, M.; Ventura, L.; Angelucci, A.; Peruzzi, B.; Tillgren, V.; Muraca, M.; Heinegård, D.; Teti, A. Proline/Arginine-Rich End Leucine-Rich Repeat Protein N-Terminus Is a Novel Osteoclast Antagonist That Counteracts Bone Loss. J. Bone Miner. Res. 2013, 28, 1912–1924. [Google Scholar] [CrossRef]
- Eguchi, J.; Wada, J. Functional Roles of Proline/Arginine-Rich End Leucine-Rich Repeat Protein (PRELP) in Adipocytes. Diabetes 2018, 67 (Suppl. 1), 33-OR. [Google Scholar] [CrossRef]
- Hruska, P.; Kucera, J.; Pekar, M.; Holéczy, P.; Mazur, M.; Buzga, M.; Kuruczova, D.; Lenart, P.; Fialova Kucerova, J.; Potesil, D.; et al. Proteomic Signatures of Human Visceral and Subcutaneous Adipocytes. J. Clin. Endocrinol. Metab. 2022, 107, 755–775. [Google Scholar] [CrossRef]
- Lewandowski, Ł.; Kepinska, M.; Milnerowicz, H. Alterations in Concentration/Activity of Superoxide Dismutases in Context of Obesity and Selected Single Nucleotide Polymorphisms in Genes: SOD1, SOD2, SOD3. Int. J. Mol. Sci. 2020, 21, 5049. [Google Scholar] [CrossRef]
- Rath-Deschner, B.; Nogueira, A.V.B.; Memmert, S.; Nokhbehsaim, M.; Augusto Cirelli, J.; Eick, S.; Miosge, N.; Kirschneck, C.; Kesting, M.; Deschner, J.; et al. Regulation of Anti-Apoptotic SOD2 and BIRC3 in Periodontal Cells and Tissues. Int. J. Mol. Sci. 2021, 22, 591. [Google Scholar] [CrossRef]
- Yoon, Y.; Kim, T.-J.; Lee, J.-M.; Kim, D.-Y. SOD2 Is Upregulated in Periodontitis to Reduce Further Inflammation Progression. Oral Dis. 2018, 24, 1572–1580. [Google Scholar] [CrossRef]
- Xia, Y.; Sun, M.; Xie, Y.; Shu, R. MTOR Inhibition Rejuvenates the Aging Gingival Fibroblasts through Alleviating Oxidative Stress. Oxid. Med. Cell. Longev. 2017, 2017, 6292630. [Google Scholar] [CrossRef] [Green Version]
- Duarte, P.M.; Napimoga, M.H.; Fagnani, E.C.; Santos, V.R.; Bastos, M.F.; Ribeiro, F.V.; Araújo, V.C.; Demasi, A.P.D. The Expression of Antioxidant Enzymes in the Gingivae of Type 2 Diabetics with Chronic Periodontitis. Arch. Oral Biol. 2012, 57, 161–168. [Google Scholar] [CrossRef]
- Knaś, M.; Maciejczyk, M.; Sawicka, K.; Hady, H.R.; Niczyporuk, M.; Ładny, J.R.; Matczuk, J.; Waszkiel, D.; Żendzian-Piotrowska, M.; Zalewska, A. Impact of Morbid Obesity and Bariatric Surgery on Antioxidant/Oxidant Balance of the Unstimulated and Stimulated Human Saliva. J. Oral Pathol. Med. 2016, 45, 455–464. [Google Scholar] [CrossRef]
- Zamora-Mendoza, R.; Rosas-Vargas, H.; Ramos-Cervantes, M.T.; Garcia-Zuniga, P.; Perez-Lorenzana, H.; Mendoza-Lorenzo, P.; Perez-Ortiz, A.C.; Estrada-Mena, F.J.; Miliar-Garcia, A.; Lara-Padilla, E.; et al. Dysregulation of Mitochondrial Function and Biogenesis Modulators in Adipose Tissue of Obese Children. Int. J. Obes. 2018, 42, 618–624. [Google Scholar] [CrossRef]
- Moreira, T.G.; Zhang, L.; Shaulov, L.; Harel, A.; Kuss, S.K.; Williams, J.; Shelton, J.; Somatilaka, B.; Seemann, J.; Yang, J.; et al. Sec13 Regulates Expression of Specific Immune Factors Involved in Inflammation in Vivo. Sci. Rep. 2015, 5, 17655. [Google Scholar] [CrossRef] [Green Version]
- Tang, B.L.; Peter, F.; Krijnse-Locker, J.; Low, S.H.; Griffiths, G.; Hong, W. The Mammalian Homolog of Yeast Sec13p Is Enriched in the Intermediate Compartment and Is Essential for Protein Transport from the Endoplasmic Reticulum to the Golgi Apparatus. Mol. Cell. Biol. 1997, 17, 256–266. [Google Scholar] [CrossRef] [Green Version]
- De Giorgio, M.R.; Yoshioka, M.; St-Amand, J. Feeding Induced Changes in the Hypothalamic Transcriptome. Clin. Chim. Acta. 2009, 406, 103–107. [Google Scholar] [CrossRef] [PubMed]
- Arriga, R.; Pacifici, F.; Capuani, B.; Coppola, A.; Orlandi, A.; Scioli, M.G.; Pastore, D.; Andreadi, A.; Sbraccia, P.; Tesauro, M.; et al. Peroxiredoxin 6 Is a Key Antioxidant Enzyme in Modulating the Link between Glycemic and Lipogenic Metabolism. Oxid. Med. Cell. Longev. 2019, 2019, 9685607. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paula, F.M.M.; Ferreira, S.M.; Boschero, A.C.; Souza, K.L.A. Modulation of the Peroxiredoxin System by Cytokines in Insulin-Producing RINm5F Cells: Down-Regulation of PRDX6 Increases Susceptibility of Beta Cells to Oxidative Stress. Mol. Cell. Endocrinol. 2013, 374, 56–64. [Google Scholar] [CrossRef] [PubMed]
- Foster, B.L.; Ao, M.; Salmon, C.R.; Chavez, M.B.; Kolli, T.N.; Tran, A.B.; Chu, E.Y.; Kantovitz, K.R.; Yadav, M.; Narisawa, S.; et al. Osteopontin Regulates Dentin and Alveolar Bone Development and Mineralization. Bone 2018, 107, 196–207. [Google Scholar] [CrossRef]
- Foster, B.L. Methods for Studying Tooth Root Cementum by Light Microscopy. Int. J. Oral Sci. 2012, 4, 119–128. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Extracellular Exosome (GO:0070062) | ||||||
|---|---|---|---|---|---|---|
| Average Peak Area Values | ||||||
| Protein Name | Uniprot Accession | Protein Symbol | Control | Obese | p-Value | Fold-Change |
| Prolargin | Q9EQP5 | Prelp | 0.00 | 1.20 × 107 | 0.014 * | 1.2 × 107 |
| Protein SEC13 homolog | Q5XFW8 | Sec13 | 7.40 × 105 | 4.60 × 106 | 0.014 * | 6.10 |
| Superoxide dismutase | P07895 | Sod2 | 2.60 × 106 | 7.80 × 106 | 0.040 * | 3.00 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nogueira, A.V.B.; Lopes, M.E.S.; Marcantonio, C.C.; Salmon, C.R.; Mofatto, L.S.; Deschner, J.; Nociti-Junior, F.H.; Cirelli, J.A. Obesity Modifies the Proteomic Profile of the Periodontal Ligament. Int. J. Mol. Sci. 2023, 24, 1003. https://doi.org/10.3390/ijms24021003
Nogueira AVB, Lopes MES, Marcantonio CC, Salmon CR, Mofatto LS, Deschner J, Nociti-Junior FH, Cirelli JA. Obesity Modifies the Proteomic Profile of the Periodontal Ligament. International Journal of Molecular Sciences. 2023; 24(2):1003. https://doi.org/10.3390/ijms24021003
Chicago/Turabian StyleNogueira, Andressa V. B., Maria Eduarda S. Lopes, Camila C. Marcantonio, Cristiane R. Salmon, Luciana S. Mofatto, James Deschner, Francisco H. Nociti-Junior, and Joni A. Cirelli. 2023. "Obesity Modifies the Proteomic Profile of the Periodontal Ligament" International Journal of Molecular Sciences 24, no. 2: 1003. https://doi.org/10.3390/ijms24021003