
Effects of Nanosecond Pulsed Electric Field (nsPEF) on a Multicellular Spheroid Tumor Model: Influence of Pulse Duration, Pulse Repetition Rate, Absorbed Energy, and Temperature

 , , , ,
, , , ,
Abstract
:1. Introduction
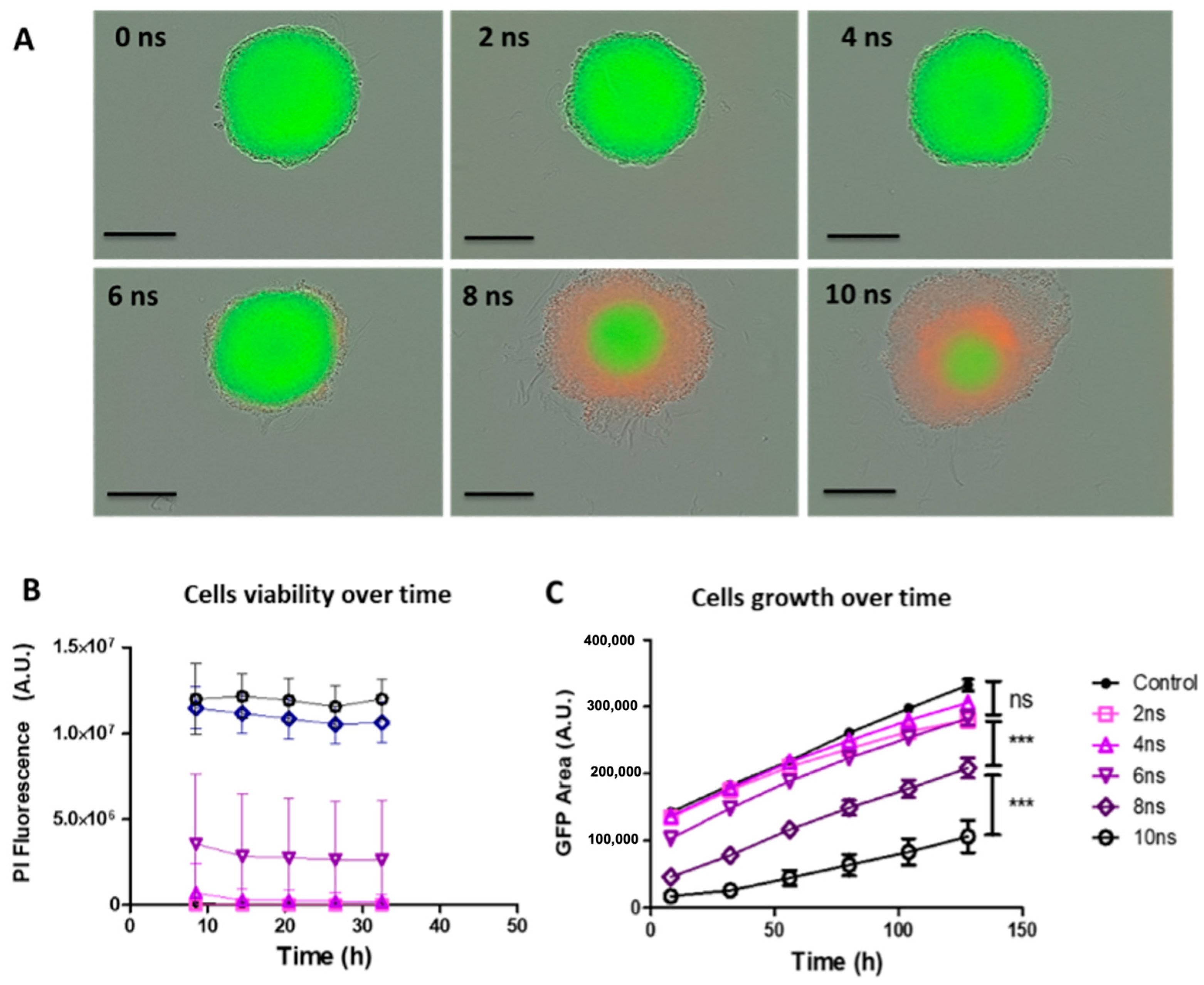
2. Results
2.1. Effects of Pulse Durations in HBSS Buffer at 20 Hz
2.2. Effects of Pulse Durations in HBSS Buffer at 200 Hz
2.3. Effects of Pulse Durations with Constant Cumulative Time Duration and Absorbed Energy Density in HBSS Buffer at 20 Hz and 200 Hz
2.4. Effect of Low-Conductivity Sucrose-Containing (ZAP) Buffer
2.5. Induced Temperature Elevation
2.5.1. Electro-Induced Temperature Elevation
2.5.2. Hyperthermia by Convection
3. Discussion
4. Materials and Methods
4.1. 3D Multicellular Spheroids
4.2. nsPEF Generator
4.3. Pulsing Buffers
4.4. Exposure Protocols
4.4.1. Pulsed Electric Field Protocols
4.4.2. Hyperthermia Protocols
4.5. Electromagnetic and Thermal Dosimetry
4.5.1. Electromagnetic Dosimetry
4.5.2. Thermal Dosimetry
4.6. Microscopy
4.7. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Gudvangen, E.; Mangalanathan, U.; Semenov, I.; Kiester, A.S.; Keppler, M.A.; Ibey, B.L.; Bixler, J.N.; Pakhomov, A.G. Pulsed Electric Field Ablation of Esophageal Malignancies and Mitigating Damage to Smooth Muscle: An In Vitro Study. Int. J. Mol. Sci. 2023, 24, 2854. [Google Scholar] [CrossRef]
- Ford, W.E.; Ren, W.; Blackmore, P.F.; Schoenbach, K.H.; Beebe, S.J. Nanosecond Pulsed Electric Fields Stimulate Apoptosis without Release of Pro-Apoptotic Factors from Mitochondria in B16f10 Melanoma. Arch. Biochem. Biophys. 2010, 497, 82–89. [Google Scholar] [CrossRef]
- Kiełbik, A.; Szlasa, W.; Novickij, V.; Szewczyk, A.; Maciejewska, M.; Saczko, J.; Kulbacka, J. Effects of High-Frequency Nanosecond Pulses on Prostate Cancer Cells. Sci. Rep. 2021, 11, 15835. [Google Scholar] [CrossRef]
- Nuccitelli, R. Application of Pulsed Electric Fields to Cancer Therapy. Bioelectricity 2019, 1, 30–34. [Google Scholar] [CrossRef]
- Nuccitelli, R. Tissue Ablation Using Nanosecond Electric Pulses. In Handbook of Electroporation; Miklavcic, D., Ed.; Springer International Publishing: Cham, Switzerland, 2016; pp. 1–11. ISBN 978-3-319-26779-1. [Google Scholar]
- Nuccitelli, R.; Wood, R.; Kreis, M.; Athos, B.; Huynh, J.; Lui, K.; Nuccitelli, P.; Epstein, E.H. First-in-Human Trial of Nanoelectroablation Therapy for Basal Cell Carcinoma: Proof of Method. Exp. Dermatol. 2014, 23, 135–137. [Google Scholar] [CrossRef]
- Ross, A.S.; Schlesinger, T.; Harmon, C.B.; Moy, R.L.; Rohrer, T.E.; Mehregan, D.R.; Nuccitelli, R.; Johnston, L.J.; Knape, W.A. Multicenter, Prospective Feasibility Study of Nano-Pulse StimulationTM Technology for the Treatment of Both Nodular and Superficial Low-Risk Basal Cell Carcinoma. Front. Oncol. 2022, 12, 1044694. [Google Scholar] [CrossRef]
- Beebe, S.J. UsEPs as a Possible Immunotherapy. In Ultrashort Electric Pulse Effects in Biology and Medicine; Beebe, S.J., Joshi, R., Schoenbach, K.H., Xiao, S., Eds.; Series in BioEngineering; Springer: Singapore, 2021; pp. 419–451. [Google Scholar]
- Beebe, S. UsEP Induce Regulated Cell Death Mechanisms|Request PDF. In Ultrashort Electric Pulse Effects in Biology and Medicine; Springer: Berlin/Heidelberg, Germany, 2021; pp. 227–263. [Google Scholar]
- Nuccitelli, R.; Berridge, J.C.; Mallon, Z.; Kreis, M.; Athos, B.; Nuccitelli, P. Nanoelectroablation of Murine Tumors Triggers a CD8-Dependent Inhibition of Secondary Tumor Growth. PLoS ONE 2015, 10, e0134364. [Google Scholar] [CrossRef]
- Nuccitelli, R.; Pliquett, U.; Chen, X.; Ford, W.; Swanson, R.J.; Beebe, S.J.; Kolb, J.F.; Schoenbach, K.H. Nanosecond Pulsed Electric Fields Cause Melanomas to Self-Destruct. Biochem. Biophys. Res. Commun. 2006, 343, 351–360. [Google Scholar] [CrossRef]
- Nuccitelli, R.; Tran, K.; Lui, K.; Huynh, J.; Athos, B.; Kreis, M.; Nuccitelli, P.; De Fabo, E.C. Non-Thermal Nanoelectroablation of UV-Induced Murine Melanomas Stimulates an Immune Response. Pigment. Cell Melanoma Res. 2012, 25, 618–629. [Google Scholar] [CrossRef]
- McDaniel, A.; Freimark, B.; Navarro, C.; Von Rothstein, K.; Gonzalez, D.; Linder, K.; Nuccitelli, R. Nano-Pulse StimulationTM Therapy (NPSTM) Is Superior to Cryoablation in Clearing Murine Melanoma Tumors. Front. Oncol. 2023, 12, 948472. [Google Scholar] [CrossRef]
- Nuccitelli, R.; Tran, K.; Sheikh, S.; Athos, B.; Kreis, M.; Nuccitelli, P. Optimized Nanosecond Pulsed Electric Field Therapy Can Cause Murine Malignant Melanomas to Self-Destruct with a Single Treatment. Int. J. Cancer 2010, 127, 1727–1736. [Google Scholar] [CrossRef]
- Beebe, S.J.; Lassiter, B.P.; Guo, S. Nanopulse Stimulation (NPS) Induces Tumor Ablation and Immunity in Orthotopic 4T1 Mouse Breast Cancer: A Review. Cancers 2018, 10, 97. [Google Scholar] [CrossRef]
- Guo, S.; Jing, Y.; Burcus, N.I.; Lassiter, B.P.; Tanaz, R.; Heller, R.; Beebe, S.J. Nano-Pulse Stimulation Induces Potent Immune Responses, Eradicating Local Breast Cancer While Reducing Distant Metastases. Int. J. Cancer 2018, 142, 629–640. [Google Scholar] [CrossRef]
- Gudvangen, E.; Kim, V.; Novickij, V.; Battista, F.; Pakhomov, A.G. Electroporation and Cell Killing by Milli- to Nanosecond Pulses and Avoiding Neuromuscular Stimulation in Cancer Ablation. Sci. Rep. 2022, 12, 1763. [Google Scholar] [CrossRef]
- Xiao, S. Pulsed Power Generators. In Ultrashort Electric Pulse Effects in Biology and Medicine; Springer: Berlin/Heidelberg, Germany, 2021; pp. 373–384. [Google Scholar]
- Tabcheh, N.; Orlacchio, R.; Couderc, V.; Leveque, P.; Arnaud-Cormos, D. High Voltage Generator With Adjustable Delay Between Two Nanosecond Pulses. IEEE Photon. Technol. Lett. 2022, 34, 553–556. [Google Scholar] [CrossRef]
- Carr, L.; Bardet, S.M.; Burke, R.C.; Arnaud-Cormos, D.; Leveque, P.; O’Connor, R.P. Calcium-Independent Disruption of Microtubule Dynamics by Nanosecond Pulsed Electric Fields in U87 Human Glioblastoma Cells. Sci. Rep. 2017, 7, 41267. [Google Scholar] [CrossRef]
- Burke, R.C.; Bardet, S.M.; Carr, L.; Romanenko, S.; Arnaud-Cormos, D.; Leveque, P.; O’Connor, R.P. Nanosecond Pulsed Electric Fields Depolarize Transmembrane Potential via Voltage-Gated K+, Ca2+ and TRPM8 Channels in U87 Glioblastoma Cells. Biochim. Biophys. Acta Biomembr. 2017, 1859, 2040–2050. [Google Scholar] [CrossRef]
- Zaklit, J.; Cabrera, A.; Shaw, A.; Aoun, R.; Vernier, P.T.; Leblanc, N.; Craviso, G.L. 5 Ns Electric Pulses Induce Ca2+-Dependent Exocytotic Release of Catecholamine from Adrenal Chromaffin Cells. Bioelectrochemistry 2021, 140, 107830. [Google Scholar] [CrossRef]
- Sözer, E.B.; Vernier, P.T. Modulation of Biological Responses to 2 ns Electrical Stimuli by Field Reversal. Biochim. Biophys. Acta Biomembr. 2019, 1861, 1228–1239. [Google Scholar] [CrossRef]
- Vernier, P.T.; Ziegler, M.J.; Sun, Y.; Gundersen, M.A.; Tieleman, D.P. Nanopore-Facilitated, Voltage-Driven Phosphatidylserine Translocation in Lipid Bilayers—In Cells and in Silico. Phys. Biol. 2006, 3, 233. [Google Scholar] [CrossRef]
- Ning, T.; Guo, J.; Zhang, K.; Li, K.; Zhang, J.; Yang, Z.; Ge, Z. Nanosecond Pulsed Electric Fields Enhanced Chondrogenic Potential of Mesenchymal Stem Cells via JNK/CREB-STAT3 Signaling Pathway. Stem Cell Res. Ther. 2019, 10, 45. [Google Scholar] [CrossRef] [PubMed]
- Carr, L.; Golzio, M.; Orlacchio, R.; Alberola, G.; Kolosnjaj-Tabi, J.; Leveque, P.; Arnaud-Cormos, D.; Rols, M.-P. A Nanosecond Pulsed Electric Field (NsPEF) Can Affect Membrane Permeabilization and Cellular Viability in a 3D Spheroids Tumor Model. Bioelectrochemistry 2021, 141, 107839. [Google Scholar] [CrossRef]
- Nuccitelli, R.; McDaniel, A.; Connolly, R.; Zelickson, B.; Hartman, H. Nano-Pulse Stimulation Induces Changes in the Intracellular Organelles in Rat Liver Tumors Treated In Situ. Lasers Surg. Med. 2020, 52, 882–889. [Google Scholar] [CrossRef] [PubMed]
- Beebe, S.J.; Ravi, J.; Schoenbach, K.; Xiao, S. Ultrashort Electric Pulse Effects in Biology and Medicine; Springer: Berlin/Heidelberg, Germany, 2021. [Google Scholar]
- Ibrahimi, N.; Vallet, L.; Andre, F.M.; Rivaletto, M.; Novac, B.M.; Mir, L.M.; Pécastaing, L. An Overview of Subnanosecond Pulsed Electric Field Biological Effects: Toward Contactless Technologies for Cancer Treatment. Bioelectricity 2023, 5, 76–98. [Google Scholar] [CrossRef]
- Kolosnjaj-Tabi, J.; Gibot, L.; Fourquaux, I.; Golzio, M.; Rols, M.-P. Electric Field-Responsive Nanoparticles and Electric Fields: Physical, Chemical, Biological Mechanisms and Therapeutic Prospects. Adv. Drug Deliv. Rev. 2019, 138, 56–67. [Google Scholar] [CrossRef] [PubMed]
- Habash, R.W.Y.; Bansal, R.; Krewski, D.; Alhafid, H.T. Thermal Therapy, Part 1: An Introduction to Thermal Therapy. Crit. Rev. Biomed. Eng. 2006, 34, 459–489. [Google Scholar] [CrossRef] [PubMed]
- Schoenbach, K.H. Thermal Effects in Bioelectrics. In Ultrashort Electric Pulse Effects in Biology and Medicine; Beebe, S.J., Joshi, R., Schoenbach, K.H., Xiao, S., Eds.; Series in BioEngineering; Springer: Singapore, 2021; pp. 275–299. [Google Scholar]
- Yan, Z.; Yin, L.; Hao, C.; Liu, K.; Qiu, J. Synergistic Effect of Pulsed Electric Fields and Temperature on the Inactivation of Microorganisms. AMB Express 2021, 11, 47. [Google Scholar] [CrossRef]
- Camp, J.; Jing, Y.; Zhuang, J.; Kolb, J.; Beebe, S.; Song, J.; Joshi, R.; Xiao, S.; Schoenbach, K. Cell Death Induced by Subnanosecond Pulsed Electric Fields at Elevated Temperatures. IEEE Trans. Plasma Sci. 2012, 40, 2334–2347. [Google Scholar] [CrossRef]
- Edelblute, C.M.; Hornef, J.; Burcus, N.I.; Norman, T.; Beebe, S.J.; Schoenbach, K.; Heller, R.; Jiang, C.; Guo, S. Controllable Moderate Heating Enhances the Therapeutic Efficacy of Irreversible Electroporation for Pancreatic Cancer. Sci. Rep. 2017, 7, 11767. [Google Scholar] [CrossRef]
- Agnass, P.; Rodermond, H.M.; van Veldhuisen, E.; Vogel, J.A.; ten Cate, R.; van Lienden, K.P.; van Gulik, T.M.; Franken, N.A.P.; Oei, A.L.; Kok, H.P.; et al. Quantitative Analysis of Contribution of Mild and Moderate Hyperthermia to Thermal Ablation and Sensitization of Irreversible Electroporation of Pancreatic Cancer Cells. J. Therm. Biol. 2023, 2023, 103619. [Google Scholar] [CrossRef]
- Hildebrandt, B.; Wust, P.; Ahlers, O.; Dieing, A.; Sreenivasa, G.; Kerner, T.; Felix, R.; Riess, H. The Cellular and Molecular Basis of Hyperthermia. Crit. Rev. Oncol./Hematol. 2002, 43, 33–56. [Google Scholar] [CrossRef] [PubMed]
- Pakhomova, O.N.; Gregory, B.W.; Semenov, I.; Pakhomov, A.G. Two Modes of Cell Death Caused by Exposure to Nanosecond Pulsed Electric Field. PLoS ONE 2013, 8, e70278. [Google Scholar] [CrossRef]
- Morotomi-Yano, K.; Akiyama, H.; Yano, K. Different Involvement of Extracellular Calcium in Two Modes of Cell Death Induced by Nanosecond Pulsed Electric Fields. Arch. Biochem. Biophys. 2014, 555–556, 47–54. [Google Scholar] [CrossRef] [PubMed]
- Kulbacka, J.; Rembiałkowska, N.; Szewczyk, A.; Rossowska, J.; Drąg-Zalesińska, M.; Kulbacki, M.; Choromańska, A. Nanosecond PEF Induces Oxidative Stress and Apoptosis via Proteasomal Activity Inhibition in Gastric Adenocarcinoma Cells with Drug Resistance. Int. J. Mol. Sci. 2022, 23, 12943. [Google Scholar] [CrossRef]
- Pakhomova, O.N.; Gregory, B.; Semenov, I.; Pakhomov, A.G. Calcium-Mediated Pore Expansion and Cell Death Following Nanoelectroporation. Biochim. Biophys. Acta (BBA)—Biomembr. 2014, 1838, 2547–2554. [Google Scholar] [CrossRef]
- Pelofy, S.; Teissié, J.; Golzio, M.; Chabot, S. Chemically Modified Oligonucleotide-Increased Stability Negatively Correlates with Its Efficacy despite Efficient Electrotransfer. J. Membr. Biol. 2012, 245, 565–571. [Google Scholar] [CrossRef]
- Leveque, P.; O’Connor, R.; Arnaud-Cormos, D. Measurement and Characterization of Exposure Systems for High-Frequency, Ultrashort Pulses. In Handbook of Electroporation; Miklavčič, D., Ed.; Springer International Publishing: Cham, Switzerland, 2017; pp. 813–836. ISBN 978-3-319-32886-7. [Google Scholar]
- Arena, C.B.; Garcia, P.A.; Sano, M.B.; Olson, J.D.; Rogers-Cotrone, T.; Rossmeisl, J.H.; Davalos, R.V. Focal Blood-Brain-Barrier Disruption with High-Frequency Pulsed Electric Fields. Technology 2014, 2, 206–213. [Google Scholar] [CrossRef]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Pulse Duration d (ns) | Number of Pulses N | Cumulative Time Duration D (μs) | Total Exposure Duration t (s) | Absorbed Energy Density u (mJ/mm3) in HBSS | Temperature Increase-ΔT (°C) in HBSS | Absorbed Energy Density u (mJ/mm3) in ZAP | Temperature Increase-ΔT (°C) in ZAP | E-Field Intensity in HBSS (kV/cm) | E-field Intensity in ZAP (kV/cm) | Number of Independent Experiments HBSS (N) | |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Protocol 1 | |||||||||||
| (PRR = 20 Hz) | 2 | 500 | 1 | 25 | 140 | 3.8 ± 0.2 | 16 | NA | 100 | 90 | 3 |
| 4 | 500 | 2 | 25 | 280 | 5.7 ± 0.3 | 32 | NA | 100 | 90 | 3 | |
| 6 | 500 | 3 | 25 | 420 | 9.1 ± 1.8 | 47 | NA | 100 | 90 | 3 | |
| 8 | 500 | 4 | 25 | 560 | 10.8 ± 2.2 | 65 | NA | 100 | 90 | 3 | |
| 10 | 500 | 5 | 25 | 700 | 15.1± 2.7 | 81 | 1.1 ± 0.2 | 100 | 90 | 3 | |
| Protocol 2 | |||||||||||
| (PRR = 200 Hz) | 2 | 500 | 1 | 2.5 | 140 | 12.6 ± 0.8 | 16 | 1.2 ± 0.1 | 100 | 90 | 2 |
| 6 | 500 | 3 | 2.5 | 420 | 32.2 ± 0.7 | 47 | 2.5 ± 0.1 | 100 | 90 | 2 | |
| 10 | 500 | 5 | 2.5 | 700 | 46.8 ± 3.9 | 81 | 4.4 ± 0.2 | 100 | 90 | 2 | |
| Protocol 3 | |||||||||||
| (PRR = 20 Hz) | 2 | 2500 | 5 | 125 | 700 | 8.5 ± 1.5 | 81 | NA | 100 | 90 | 3 |
| 6 | 833 | 5 | 41.6 | 700 | 15.2 ± 1.9 | 81 | NA | 100 | 90 | 3 | |
| 10 | 500 | 5 | 25 | 700 | 16.8 ± 2.2 | 81 | 1.1 ± 0.2 | 100 | 90 | 3 | |
| Protocol 4 | |||||||||||
| (PRR = 200 Hz) | 2 | 2500 | 5 | 12.5 | 700 | 22.0 ± 2.2 | 81 | 3.2 ± 0.1 | 100 | 90 | 2 |
| 6 | 833 | 5 | 4.16 | 700 | 42.8 ± 1.4 | 81 | 3.7 ± 0.4 | 100 | 90 | 2 | |
| 10 | 500 | 5 | 2.5 | 700 | 46.8 ± 3.9 | 81 | 4.4 ± 0.2 | 100 | 90 | 2 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Orlacchio, R.; Kolosnjaj-Tabi, J.; Mattei, N.; Lévêque, P.; Rols, M.P.; Arnaud-Cormos, D.; Golzio, M. Effects of Nanosecond Pulsed Electric Field (nsPEF) on a Multicellular Spheroid Tumor Model: Influence of Pulse Duration, Pulse Repetition Rate, Absorbed Energy, and Temperature. Int. J. Mol. Sci. 2023, 24, 14999. https://doi.org/10.3390/ijms241914999
Orlacchio R, Kolosnjaj-Tabi J, Mattei N, Lévêque P, Rols MP, Arnaud-Cormos D, Golzio M. Effects of Nanosecond Pulsed Electric Field (nsPEF) on a Multicellular Spheroid Tumor Model: Influence of Pulse Duration, Pulse Repetition Rate, Absorbed Energy, and Temperature. International Journal of Molecular Sciences. 2023; 24(19):14999. https://doi.org/10.3390/ijms241914999
Chicago/Turabian StyleOrlacchio, Rosa, Jelena Kolosnjaj-Tabi, Nicolas Mattei, Philippe Lévêque, Marie Pierre Rols, Delia Arnaud-Cormos, and Muriel Golzio. 2023. "Effects of Nanosecond Pulsed Electric Field (nsPEF) on a Multicellular Spheroid Tumor Model: Influence of Pulse Duration, Pulse Repetition Rate, Absorbed Energy, and Temperature" International Journal of Molecular Sciences 24, no. 19: 14999. https://doi.org/10.3390/ijms241914999