
Consequences of Disturbing Manganese Homeostasis

 , , , , ,
, , , , ,
Abstract
:1. Introduction
2. Manganese (Mn)
2.1. The Source of Exposure to Mn
2.2. Recommended Dietary Intake of Mn
2.3. Accumulation of Mn in the Brain
2.4. Evaluation of the State of Mn
2.5. Reference Values for Mn Homeostasis
2.6. Association of Mn with Other Metals
2.6.1. Magnesium (Mg)
2.6.2. Iron (Fe)
2.6.3. Zinc (Zn), Copper (Cu), Lead (Pb), and Calcium (Ca)
3. The Role of Mn in the Body
3.1. Mn-Dependent Enzymes
3.2. The Role of Mn in Smooth Muscle Cell Contraction
3.3. Mn Transport
3.4. Oxidative Stress/Inflammation
3.5. Neurotransmitters
3.6. Protein Aggregation
4. Brain Function and Neurodevelopment
4.1. Neurodegenerative Disorders
4.2. Autism Spectrum Disorder (ASD)
4.3. Attention Deficit Hyperactivity Disorder (ADHD)
4.4. Manganism
4.4.1. Parkinson Disease (PD)
4.4.2. Alzheimer’s Disease (AD)
4.4.3. Huntington’s Disease (HD)
4.5. Epilepsy
4.6. Prion Diseases
4.7. Leigh-Like Syndrome
5. Infectious Diseases
6. Genetic Diseases Associated with Mn Imbalance
6.1. Mn Deficiency
6.2. Mn Accumulation
6.3. Mn Deficiency and Carcinogenesis
7. Metabolic Diseases
7.1. Type 2 Diabetes Mellitus/Insulin Resistance
7.2. Osteoporosis
7.3. Obesity
7.4. Atherosclerosis
7.5. Non-Alcoholic Fatty Liver Disease
8. Fertility
8.1. Male Fertility
8.2. Female Fertility
9. Wound Healing
10. Conclusions and Future Prospective
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Skalnaya, M.G.; Skalny, A.V. Essential Trace Elements in Human Health: A Physician’s View; Publishing House of Tomsk State University: Tomsk, Russia, 2018; ISBN 978-5-94621-683-8. [Google Scholar]
- Horowitz, G.L.; Altaie, S.; Boyd, J.; Ceriotti, F.; Gard, U.; Horn, P.; Pesce, A.; Sine, H.; Zakowski, J. Defining, Establishing, and Verifying Reference Intervals in the Clinical Laboratory, 3rd ed.; Approved Guidelines; Clinical and Laboratory Standards Institute document C28-A3; CLSI: Wayne, PA, USA, 2008. [Google Scholar]
- Guo, H.; Li, M.; Liu, H. Selenium-Rich Yeast Peptide Fraction Ameliorates Imiquimod-Induced Psoriasis-like Dermatitis in Mice by Inhibiting Inflammation via MAPK and NF-κB Signaling Pathways. Int. J. Mol. Sci. 2022, 23, 2112. [Google Scholar] [CrossRef] [PubMed]
- Wu, T.; Gagnon, A.; McGourty, K.; DosSantos, R.; Chanetsa, L.; Zhang, B.; Bello, D.; Kelleher, S.L. Zinc Exposure Promotes Commensal-to-Pathogen Transition in Pseudomonas aeruginosa Leading to Mucosal Inflammation and Illness in Mice. Int. J. Mol. Sci. 2021, 22, 13321. [Google Scholar] [CrossRef]
- Cunningham, F.; Cahyadi, S.; Lengyel, I. A Potential New Role for Zinc in Age-Related Macular Degeneration through Regulation of Endothelial Fenestration. Int. J. Mol. Sci. 2021, 22, 11974. [Google Scholar] [CrossRef] [PubMed]
- Xiao, J.; Li, N.; Xiao, S.; Wu, Y.; Liu, H. Comparison of Selenium Nanoparticles and Sodium Selenite on the Alleviation of Early Atherosclerosis by Inhibiting Endothelial Dysfunction and Inflammation in Apolipoprotein E-Deficient Mice. Int. J. Mol. Sci. 2021, 22, 11612. [Google Scholar] [CrossRef]
- Nicolai, M.M.; Weishaupt, A.-K.; Baesler, J.; Brinkmann, V.; Wellenberg, A.; Winkelbeiner, N.; Gremme, A.; Aschner, M.; Fritz, G.; Schwerdtle, T.; et al. Effects of Manganese on Genomic Integrity in the Multicellular Model Organism Caenorhabditis elegans. Int. J. Mol. Sci. 2021, 22, 10905. [Google Scholar] [CrossRef] [PubMed]
- Vogel-González, M.; Musa-Afaneh, D.; Rivera Gil, P.; Vicente, R. Zinc Favors Triple-Negative Breast Cancer’s Microenvironment Modulation and Cell Plasticity. Int. J. Mol. Sci. 2021, 22, 9188. [Google Scholar] [CrossRef] [PubMed]
- Li, K.; Feng, T.; Liu, L.; Liu, H.; Huang, K.; Zhou, J. Hepatic Proteomic Analysis of Selenoprotein T Knockout Mice by TMT: Implications for the Role of Selenoprotein T in Glucose and Lipid Metabolism. Int. J. Mol. Sci. 2021, 22, 8515. [Google Scholar] [CrossRef]
- Skalny, A.V.; Aschner, M.; Lei, X.G.; Gritsenko, V.A.; Santamaria, A.; Alekseenko, S.I.; Prakash, N.T.; Chang, J.-S.; Sizova, E.A.; Chao, J.C.J.; et al. Gut Microbiota as a Mediator of Essential and Toxic Effects of Zinc in the Intestines and Other Tissues. Int. J. Mol. Sci. 2021, 22, 13074. [Google Scholar] [CrossRef]
- Koski, L.; Ronnevi, C.; Berntsson, E.; Wärmländer, S.K.T.S.; Roos, P.M. Metals in ALS TDP-43 Pathology. Int. J. Mol. Sci. 2021, 22, 12193. [Google Scholar] [CrossRef]
- He, Z.; You, G.; Liu, Q.; Li, N. Alzheimer’s Disease and Diabetes Mellitus in Comparison: The Therapeutic Efficacy of the Vanadium Compound. Int. J. Mol. Sci. 2021, 22, 11931. [Google Scholar] [CrossRef]
- Kim, S.J.; Choi, M.C.; Park, J.M.; Chung, A.S. Antitumor Effects of Selenium. Int. J. Mol. Sci. 2021, 22, 11844. [Google Scholar] [CrossRef]
- Ye, R.; Huang, J.; Wang, Z.; Chen, Y.; Dong, Y. Trace Element Selenium Effectively Alleviates Intestinal Diseases. Int. J. Mol. Sci. 2021, 22, 11708. [Google Scholar] [CrossRef]
- Aaseth, J.; Skalny, A.V.; Roos, P.M.; Alexander, J.; Aschner, M.; Tinkov, A.A. Copper, Iron, Selenium and Lipo-Glycemic Dysmetabolism in Alzheimer’s Disease. Int. J. Mol. Sci. 2021, 22, 9461. [Google Scholar] [CrossRef]
- Mertz, W. Review of the scientific basis for establishing the essentiality of trace elements. Biol. Trace Elem. Res. 1998, 66, 185–191. [Google Scholar] [CrossRef] [PubMed]
- Otten, J.J.; Hellwig, J.P.; Meyers, L.D. Dietary Reference Intakes: The Essential Guide to Nutrient Requirements; National Academies Press: Washington, DC, USA, 2006; p. 1344. [Google Scholar]
- Serebryansky, E.P.; Skalny, A.V.; Kuznetzov, V.V. Rapid ICP-OES determination of up to 20 essential and toxic elements in human hair for estimation of human microelemental status. In Proceedings of the 21st Workshop on Macro and Trace Elements, Jena, Germany, 18–19 October 2002. [Google Scholar]
- Skalny, A.V. Bioelementology as an interdisciplinary integrative approach in life sciences: Terminology, classification, perspectives. J. Trace Elem. Med. Biol. 2011, 25S, S3–S10. [Google Scholar] [CrossRef]
- Hope, S.; Daniel, K.; Gleason, K.L.; Comber, S.; Nelson, M.; Powell, J.J. Influence of tea drinking on manganese intake, manganese status and leucocyte expression of MnSOD and cytosolic aminopeptidase P. Eur. J. Clin. Nutr. 2006, 60, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Expert Group on Vitamins and Minerals, 2003. Safe Upper Levels for Vitamins and Minerals. United Kingdom: Food Standards Agency. Available online: www.food.gov.uk/multimedia/pdfs/vitamin2003.pdf (accessed on 25 March 2011).
- Lao, Y.; Dion, L.A.; Gilbert, G.; Bouchard, M.F.; Rocha, G.; Wang, Y.; Leporé, N.; Saint-Amour, D. Mapping the basal ganglia alterations in children chronically exposed to manganese. Sci. Rep. 2017, 7, 41804. [Google Scholar] [CrossRef]
- Haynes, E.N.; Sucharew, H.; Kuhnell, P.; Alden, J.; Barnas, M.; Wright, R.O.; Parsons, P.J.; Aldous, K.M.; Praamsma, M.L.; Beidler, C.; et al. Manganese Exposure and Neurocognitive Outcomes in Rural School-Age Children: The Communities Actively Researching Exposure Study (Ohio, USA). Environ. Health Perspect. 2015, 123, 1066–1071. [Google Scholar] [CrossRef] [PubMed]
- Hsieh, C.T.; Liang, J.S.; Peng, S.S.; Lee, W.T. Seizure associated with total parenteral nutrition-related hypermanganesemia. Pediatr. Neurol. 2007, 36, 181–183. [Google Scholar] [CrossRef]
- Aschner, J.L.; Anderson, A.; Slaughter, J.C.; Aschner, M.; Steele, S.; Beller, A.; Mouvery, A.; Furlong, H.M.; Maitre, N.L. Neuroimaging identifies increased manganese deposition in infants receiving parenteral nutrition. Am. J. Clin. Nutr. 2015, 102, 1482–1489. [Google Scholar] [CrossRef] [PubMed]
- Sikk, K.; Haldre, S.; Aquilonius, S.M.; Taba, P. Manganese-Induced Parkinsonism due to Ephedrone Abuse. Park. Dis. 2011, 2011, 865319. [Google Scholar] [CrossRef]
- Iqbal, M.; Monaghan, T.; Redmond, J. Manganese toxicity with ephedrone abuse manifesting as parkinsonism: A case report. J. Med. Case Rep. 2012, 6, 52. [Google Scholar] [CrossRef]
- Teeguarden, J.G.; Gearhart, J.; Clewell, H.J., 3rd; Covington, T.R.; Nong, A.; Andersen, M.E. Pharmacokinetic Modeling of Manganese. III. Physiologic Approaches Accounting for Background and Tracer Kinetics. J. Toxicol. Environ. Health A 2007, 70, 1515–1526. [Google Scholar] [CrossRef] [PubMed]
- Aschner, M. The transport of manganese across the blood-brain barrier. Neurotoxicology 2006, 27, 311–314. [Google Scholar] [CrossRef] [PubMed]
- Elsner, R.J.; Spangler, J.G. Neurotoxicity of inhaled manganese: Public health danger in the shower? Med. Hypotheses 2005, 65, 607–616. [Google Scholar] [CrossRef] [PubMed]
- Aschner, M. Manganese in the shower: Mere speculation over an invalidated public health danger. Med. Hypotheses 2006, 66, 200–201. [Google Scholar] [CrossRef] [PubMed]
- Yin, Z.; Jiang, H.; Lee, E.S.; Ni, M.; Erikson, K.M.; Milatovic, D.; Bowman, A.B.; Aschner, M. Ferroportin is a manganese-responsive protein that decreases manganese cytotoxicity and accumulation. J. Neurochem. 2010, 112, 1190–1198. [Google Scholar] [CrossRef]
- Finley, J.W.; Davis, C.D. Manganese deficiency and toxicity: Are high or low dietary amounts of manganese cause for concern? Biofactors 1999, 10, 15–24. [Google Scholar] [CrossRef]
- Abboud, S.; Haile, D.J. A novel mammalian iron-regulated protein involved in intracellular iron metabolism. J. Biol. Chem. 2000, 275, 19906–19912. [Google Scholar] [CrossRef]
- Donovan, A.; Lima, C.A.; Pinkus, J.L.; Pinkus, G.S.; Zon, L.I.; Robine, S.; Andrews, N.C. The iron exporter ferroportin/Slc40a1 is essential for iron homeostasis. Cell Metab. 2005, 1, 191–200. [Google Scholar] [CrossRef]
- McKie, A.T.; Marciani, P.; Rolfs, A.; Brennan, K.; Wehr, K.; Barrow, D.; Miret, S.; Bomford, A.; Peters, T.J.; Farzaneh, F.; et al. A novel duodenal iron-regulated transporter, IREG1, implicated in the basolateral transfer of iron to the circulation. Mol. Cell 2000, 5, 299–309. [Google Scholar] [CrossRef] [PubMed]
- Mora, A.M.; Arora, M.; Harley, K.G.; Kogut, K.; Parra, K.; Hernández-Bonilla, D.; Gunier, R.B.; Bradman, A.; Smith, D.R.; Eskenazi, B. Prenatal and postnatal manganese teeth levels and neurodevelopMent at 7, 9, and 10.5 years in the CHAMACOS cohort. Environ. Int. 2015, 84, 39–54. [Google Scholar] [CrossRef]
- Gunier, R.B.; Arora, M.; Jerrett, M.; Bradman, A.; Harley, K.G.; Mora, A.M.; Kogut, K.; Hubbard, A.; Austin, C.; Holland, N.; et al. Manganese in teeth and neurodevelopment in young Mexican-American children. Environ. Res. 2015, 142, 688–695. [Google Scholar] [CrossRef]
- Gulson, B.; Mizon, K.; Taylor, A.; Korsch, M.; Stauber, J.; Davis, J.M.; Louie, H.; Wu, M.; Swan, H. Changes in manganese and lead in the environment and young children associated with the introduction of methylcyclopentadienyl manganese tricarbonyl in gasoline--preliminary results. Environ. Res. 2006, 100, 100–114. [Google Scholar] [CrossRef]
- Filippini, T.; Michalke, B.; Grill, P.; Malagoli, C.; Malavolti, M.; Vescovi, L.; Sieri, S.; Krogh, V.; Cherubini, A.; Maffeis, G.; et al. Determinants of serum manganese levels in an Italian population. Mol. Med. Rep. 2017, 15, 3340–3349. [Google Scholar] [CrossRef] [PubMed]
- Horning, K.J.; Caito, S.W.; Tipps, K.G.; Bowman, A.B.; Aschner, M. Manganese Is Essential for Neuronal Health. Annu. Rev. Nutr. 2015, 35, 71–108. [Google Scholar] [CrossRef] [PubMed]
- National Center for Environmental Assessment. Manganese (CASRN 7439-96-5); Integrated Risk Information System, Environmental Protection Agency: Washington, DC, USA, 2022.
- Environmental Protection Agency. Secondary Drinking Water Standards: Guidance for Nuisance Chemicals; Environmental Protection Agency: Washington, DC, USA, 2022.
- Bevan, R.; Jones, K.; Cocker, J.; Assem, F.L.; Levy, L.S. Reference ranges for key biomarkers of chemical exposure within the UK population. Int. J. Hyg. Environ. Health 2013, 216, 170–174. [Google Scholar] [CrossRef]
- Hoet, P.; Jacquerye, C.; Deumer, G.; Lison, D.; Haufroid, V. Reference values and upper reference limits for 26 trace elements in the urine of adults living in Belgium. Clin. Chem. Lab. Med. 2013, 51, 839–849. [Google Scholar] [CrossRef]
- Du, X.; Graedel, T.E. Global in-use stocks of the rare Earth elements: A first estimate. Environ. Sci. Technol. 2011, 45, 4096–4101. [Google Scholar] [CrossRef]
- White, M.A.; Sabbioni, E. Trace element reference values in tissues from inhabitants of the European Union. X. A study of 13 elements in blood and urine of a United Kingdom population. Sci. Total. Environ. 1998, 216, 253–270. [Google Scholar] [CrossRef]
- Sieniawska, C.E.; Jung, L.C.; Olufadi, R.; Walker, V. Twenty-four-hour urinary trace element excretion: Reference intervals and interpretive issues. Ann. Clin. Biochem. 2012, 49, 341–351. [Google Scholar] [CrossRef]
- Schoeters, G.; Den Hond, E.; Colles, A.; Loots, I.; Morrens, B.; Keune, H.; Bruckers, L.; Nawrot, T.; Sioen, I.; De Coster, S.; et al. Concept of the Flemish human biomonitoring programme. Int. J. Hyg. Environ. Health 2012, 215, 102–108. [Google Scholar] [CrossRef] [PubMed]
- Cerná, M.; Spĕvácková, V.; Batáriová, A.; Smíd, J.; Cejchanová, M.; Ocadlíková, D.; Bavorová, H.; Benes, B.; Kubínová, R. Human biomonitoring system in the Czech Republic. Int. J. Hyg. Environ. Health 2007, 210, 495–499. [Google Scholar] [CrossRef] [PubMed]
- Schulz, C.; Wilhelm, M.; Heudorf, U.; Kolossa-Gehring, M. Human Biomonitoring Commission of the German Federal Environment Agency. Update of the reference and HBM values derived by the German Human Biomonitoring Commission. Int. J. Hyg. Environ. Health 2011, 215, 26–35. [Google Scholar] [CrossRef] [PubMed]
- NHANES, Centers for Disease Control and Prevention, National Health and Nutrition Examination Survey (NHANES) Fourth National Exposure Report, Updated Tables, February 2015. Available online: https://www.cdc.gov/biomonitoring/pdf/fourthreport_updatedtables_feb2015.pdf (accessed on 21 June 2017).
- Morton, J.; Tan, E.; Leese, E.; Cocker, J. Determination of 61 elements in urine samples collected from a non-occupationally exposed UK adult population. Toxicol. Lett. 2014, 231, 179–193. [Google Scholar] [CrossRef]
- Wang, L.D. Comprehensive Report of Chinese National Nutrition and Health Survey in 2002; People’s Medical Publishing House: Beijing, China, 2005. [Google Scholar]
- Leblanc, J.C.; Guérin, T.; Noël, L.; Calamassi-Tran, G.; Volatier, J.L.; Verger, P. Dietary exposure estimates of 18 elements from the 1st French Total Diet Study. Food Addit. Contam. 2005, 22, 624–641. [Google Scholar] [CrossRef]
- Turconi, G.; Minoia, C.; Ronchi, A.; Roggi, C. Dietary exposure estimates of twenty–one trace elements from a Total Diet Study carried out in Pavia, Northern Italy. Br. J. Nutr. 2009, 101, 1200–1208. [Google Scholar] [CrossRef]
- Rubio, C.; Gutiérrez, A.J.; Revert, C.; Reguera, J.I.; Burgos, A.; Hardisson, A. Daily dietary intake of iron, copper, zinc and manganese in a Spanish population. Int. J. Food Sci. Nutr. 2009, 60, 590–600. [Google Scholar] [CrossRef]
- Kim, E.Y.; Bae, Y.J.; Kim, S.J.; Choi, M.K. Estimation of manganese daily intake among adults in Korea. Nutr. Res. Pract. 2008, 2, 22–25. [Google Scholar] [CrossRef]
- Vanek, V.W.; Borum, P.; Buchman, A.; Fessler, T.A.; Howard, L.; Jeejeebhoy, K.; Kochevar, M.; Shenkin, A.; Valentine, C.J. Novel Nutrient Task Force, Parenteral Multi-Vitamin and Multi–Trace Element Working Group; American Society for Parenteral and Enteral Nutrition (A.S.P.E.N.) Board of Directors. A.S.P.E.N. position paper: Recommendations for changes in commercially available parenteral multivitamin and multi-trace element products. Nutr. Clin. Pract. 2012, 27, 440–491. [Google Scholar] [CrossRef]
- Friedman, B.J.; Freeland-Graves, J.H.; Bales, C.W.; Behmardi, F.; Shorey-Kutschke, R.L.; Willis, R.A.; Crosby, J.B.; Trickett, P.C.; Houston, S.D. Manganese balance and clinical observations in young men fed a manganese-deficient diet. J. Nutr. 1987, 117, 133–143. [Google Scholar] [CrossRef]
- Aschner, M.; Erikson, K.M.; Dorman, D.C. Manganese dosimetry: Species differences and implications for neurotoxicity. Crit. Rev. Toxicol. 2005, 35, 1–32. [Google Scholar] [CrossRef]
- Lehmann, T.; Lämmle, B. IFNalpha treatment in systemic mastocytosis. Ann. Hematol. 1999, 78, 483–484. [Google Scholar] [CrossRef] [PubMed]
- Das, A., Jr.; Hammad, T.A. Efficacy of a combination of FCHG49 glucosamine hydrochloride, TRH122 low molecular weight sodium chondroitin sulfate and manganese ascorbate in the management of knee osteoarthritis. Osteoarthr. Cartil. 2000, 8, 343–350. [Google Scholar] [CrossRef] [PubMed]
- National Academy of Sciences. National Research Council Recommended Daily Allowances, 9th ed.; National Academy of Sciences: Washington, DC, USA, 1980. [Google Scholar]
- World Health Organization. Manganese in Drinking-Water: Background Document for Development of WHO Guidelines for Drinking Water Quality; WHO Press: Geneva, Switzerland, 2011; pp. 5–6. [Google Scholar]
- Santamaria, A.B. Manganese exposure, essentiality and toxicity. Indian J. Med. Res. 2008, 128, 484–500. [Google Scholar] [PubMed]
- Skalny, A.V.; Rudakov, I.A. Bioelements in Medicine; Publishing House “ONIX 21st Century”, Mir: Moscow, Russia, 2004; p. 271. (In Russian) [Google Scholar]
- Skalny, A.V. Chemical Elements in Human Physiology and Ecology; Publishing House “ONIX 21st Century”, Mir: Moscow, Russia, 2004; p. 216. (In Russian) [Google Scholar]
- Finley, J.W.; Johnson, P.E.; Johnson, L.K. Sex affects manganese absorption and retention by humans from a diet adequate in manganese. Am. J. Clin. Nutr. 1994, 60, 949–955. [Google Scholar] [CrossRef]
- Ngueta, G.; Kengne, A.P. Low-Level Environmental Lead Exposure and Dysglycemia in Adult Individuals: Results from the Canadian Health and Measure Survey 2007–2011. Biol. Trace Elem. Res. 2017, 175, 278–286. [Google Scholar] [CrossRef]
- Bocca, B.; Madeddu, R.; Asara, Y.; Tolu, P.; Marchal, J.A.; Forte, G. Assessment of reference ranges for blood Cu, Mn, Se and Zn in a selected Italian population. J. Trace Elem. Med. Biol. 2011, 25, 19–26. [Google Scholar] [CrossRef]
- Clark, N.A.; Teschke, K.; Rideout, K.; Copes, R. Trace element levels in adults from the west coast of Canada and associations with age, gender, diet, activities, and levels of other trace elements. Chemosphere 2007, 70, 155–164. [Google Scholar] [CrossRef]
- Olsén, L.; Lind, P.M.; Lind, L. Gender differences for associations between circulating levels of metals and coronary risk in the elderly. Int. J. Hyg. Environ. Health 2012, 215, 411–417. [Google Scholar] [CrossRef]
- Lee, J.W.; Lee, C.K.; Moon, C.S.; Choi, I.J.; Lee, K.J.; Yi, S.M.; Jang, B.K.; Yoon, B.J.; Kim, D.S.; Peak, D.; et al. Korea National Survey for Environmental Pollutants in the Human Body 2008: Heavy metals in the blood or urine of the Korean population. Int. J. Hyg. Environ. Health 2012, 215, 449–457. [Google Scholar] [CrossRef] [PubMed]
- Oulhote, Y.; Mergler, D.; Bouchard, M.F. Sex- and age-differences in blood manganese levels in the U.S. general population: National health and nutrition examination survey 2011–2012. Environ. Health 2014, 13, 87. [Google Scholar] [CrossRef] [PubMed]
- Bock, N.A.; Paiva, F.F.; Nascimento, G.C.; Newman, J.D.; Silva, A.C. Cerebrospinal fluid to brain transport of manganese in a non-human primate revealed by MRI. Brain Res. 2008, 1198, 160–170. [Google Scholar] [CrossRef]
- Ma, R.E.; Ward, E.J.; Yeh, C.L.; Snyder, S.; Long, Z.; Gokalp Yavuz, F.; Zauber, S.E.; Dydak, U. Thalamic GABA levels and occupational manganese neurotoxicity: Association with exposure levels and brain MRI. Neurotoxicology 2018, 64, 30–42. [Google Scholar] [CrossRef]
- Olanow, C.W. Manganese-induced parkinsonism and Parkinson’s disease. Ann. N. Y. Acad. Sci. 2004, 1012, 209–223. [Google Scholar] [CrossRef]
- Criswell, S.R.; Perlmutter, J.S.; Huan, J.L.; Golchin, N.; Flores, H.P.; Hobson, A.; Aschner, M.; Erikson, K.M.; Checkoway, H.; Racette, B.A. Basal ganglia intensity indices and diffusion weighted imaging in manganese-exposed welders. Occup. Environ. Med. 2012, 69, 437–443. [Google Scholar] [CrossRef] [PubMed]
- Elder, A.; Gelein, R.; Silva, V.; Feikert, T.; Opanashuk, L.; Carter, J.; Potter, R.; Maynard, A.; Ito, Y.; Finkelstein, J.; et al. Translocation of inhaled ultrafine manganese oxide particles to the central nervous system. Environ. Health Perspect 2006, 114, 1172–1178. [Google Scholar] [CrossRef]
- Williams, M.; Todd, G.D.; Roney, N.; Crawford, J.; Coles, C.; McClure, P.R.; Garey, J.D.; Zaccaria, K.; Citra, M. Toxicological Profile for Manganese; Agency for Toxic Substances and Disease Registry: Atlanta, GA, USA, 2012. Available online: https://www.ncbi.nlm.nih.gov/books/NBK158872/ (accessed on 9 February 2016).
- Crossgrove, J.; Zheng, W. Manganese toxicity upon overexposure. NMR Biomed. 2004, 17, 544–553. [Google Scholar] [CrossRef]
- Takeda, A. Manganese action in brain function. Brain Res. Brain Res. Rev. 2003, 41, 79–87. [Google Scholar] [CrossRef]
- Baj, J.; Kowalska, B.; Flieger, W.; Radzikowska-Büchner, E.; Forma, A.; Czeczelewski, M.; Kędzierawski, P.; Karakuła, K.; Flieger, M.; Majerek, D.; et al. Assessment of the Concentration of 51 Elements in the Liver and in Various Parts of the Human Brain—Profiling of the Mineral Status. Nutrients 2023, 15, 2799. [Google Scholar] [CrossRef]
- Baj, J.; Form, A.; Kowalska, B.; Teresinski, G.; Buszewicz, G.; Majerek, D.; Flieger, W.; Maciejewski, R.; Karakula, K.; Flieger, M.; et al. Multi-Elemental Analysis of Human Optic Chiasm—A New Perspective to Reveal the Pathochanism of Nerve Fibers’ Degeneration. Int. J. Environ. Res. Public Health 2022, 19, 4420. [Google Scholar] [CrossRef] [PubMed]
- Baj, J.; Teresiński, G.; Kowalska, B.; Krajka, T.; Buszewicz, G.; Forma, A.; Flieger, W.; Karakuła, K.H.; Kędzierawski, P.; Cywka, T.; et al. ICP-MS Multi-Elemental Analysis of the Human Meninges Collected from Sudden Death Victims in South-Eastern Poland. Molecules 2022, 27, 1911. [Google Scholar] [CrossRef] [PubMed]
- Michalke, B.; Aslanoglou, L.; Ochsenkühn-Petropoulou, M.; Bergström, B.; Berthele, A.; Vinceti, M.; Lucio, M.; Lidén, G. An approach for manganese biomonitoring using a manganese carrier switch in serum from transferrin to citrate at slightly elevated manganese concentration. J. Trace Elem. Med. Biol. 2015, 32, 145–154. [Google Scholar] [CrossRef]
- Michalke, B.; Fernsebner, K. New insights into manganese toxicity and speciation. J. Trace Elem. Med. Biol. 2014, 28, 106–116. [Google Scholar] [CrossRef]
- Horowitz, G.L.; Altaie, S.; Boyd, J.C.; Ceriotti, F.; Garg, U.; Horn, P.; Pesce, A.; Sine, H.E.; Zakowski, J. CLSI/IFCC Guideline C28-A3c. Defining, Establishing, and Verifying Reference Intervals in the Clinical Laboratory, 3rd ed.; Approved Guideline; CLSI: Wayne, PA, USA, 2008. [Google Scholar]
- Ozarda, Y. Reference intervals: Current status, recent developments and future considerations. Biochem. Med. 2016, 26, 5–16. [Google Scholar] [CrossRef] [PubMed]
- Baker, M.G.; Simpson, C.D.; Sheppard, L.; Stover, B.; Morton, J.; Cocker, J.; Seixas, N. Variance components of short-term biomarkers of manganese exposure in an inception cohort of welding trainees. J. Trace Elem. Med. Biol. 2015, 29, 123–129. [Google Scholar] [CrossRef]
- Underwood, E.J. Trace Elements in Human and Animal Nutrition, 4th ed.; Academic Press: New York, NY, USA, 1977. [Google Scholar]
- Rükgauer, M.; Klein, J.; Kruse-Jarres, J.D. Reference values for the trace elements copper, manganese, selenium, and zinc in the serum/plasma of children, adolescents, and adults. J. Trace Elem. Med. Biol. 1997, 11, 92–98. [Google Scholar] [CrossRef]
- Wilson, D.C.; Tubman, R.; Bell, N.; Halliday, H.L.; McMaster, D. Plasma manganese, selenium and glutathione peroxidase levels in the mother and newborn infant. Early Hum. Dev. 1991, 26, 223–226. [Google Scholar] [CrossRef]
- Li, X.; Xie, J.; Lu, L.; Zhang, L.; Zhang, L.; Zou, Y.; Wang, Q.; Luo, X.; Li, S. Kinetics of manganese transport and gene expressions of manganese transport carriers in Caco-2 cell monolayers. Biometals 2013, 26, 941–953. [Google Scholar] [CrossRef]
- Nemeth, E.; Tuttle, M.S.; Powelson, J.; Vaughn, M.B.; Donovan, A.; Ward, D.M.; Ganz, T.; Kaplan, J. Hepcidin regulates cellular iron efflux by binding to ferroportin and inducing its internalization. Science 2004, 306, 2090–2093. [Google Scholar] [CrossRef]
- Mukhopadhyay, S.; Linstedt, A.D. Identification of a gain-of-function mutation in a Golgi P-type ATPase that enhances Mn2+ efflux and protects against toxicity. Proc. Natl. Acad. Sci. USA 2011, 108, 858–863. [Google Scholar] [CrossRef]
- He, W.; Hu, Z. The role of the Golgi-resident SPCA Ca2+/Mn2+pump in ionic homeostasis and neural function. Neurochem. Res. 2012, 37, 455–468. [Google Scholar] [CrossRef]
- Culotta, V.C.; Yang, M.; Hall, M.D. Manganese Transport and Trafficking:Lessons Learned from Saccharomyces cerevisiae. Eukaryot Cell 2005, 4, 1159–1165. [Google Scholar] [CrossRef] [PubMed]
- Fujishiro, H. Roles of ZIP8, ZIP14, and DMT1 in transport of cadmium and manganese in mouse kidney proximal tubule cells. Metallomics 2012, 4, 700–708. [Google Scholar] [CrossRef]
- Baker, M.G.; Simpson, C.D.; Stover, B.; Sheppard, L.; Checkoway, H.; Racette, B.A.; Seixas, N.S. Blood manganese as an exposure biomarker: State of the evidence. J. Occup. Environ. Hyg. 2014, 11, 210–217. [Google Scholar] [CrossRef] [PubMed]
- O’Neal, S.L.; Zheng, W. Manganese Toxicity Upon Overexposure: A Decade in Review. Curr. Environ. Health Rep. 2015, 2, 315–328. [Google Scholar] [CrossRef]
- Smith, D.; Gwiazda, R.; Bowler, R.; Roels, H.; Park, R.; Taicher, C.; Lucchini, R. Biomarkers of Mn exposure in humans. Am. J. Ind. Med. 2007, 50, 801–811. [Google Scholar] [CrossRef]
- Chojnacka, K.; Zielińska, A.; Górecka, H.; Dobrzański, Z.; Górecki, H. Reference values for hair minerals of Polish students. Environ. Toxicol. Pharmacol. 2010, 29, 314–319. [Google Scholar] [CrossRef]
- Skalny, A.V.; Skalnaya, M.G.; Tinkov, A.A.; Serebryansky, E.P.; Demidov, V.A.; Lobanova, Y.N.; Grabeklis, A.R.; Berezkina, E.S.; Gryazeva, I.V.; Skalny, A.A.; et al. Reference values of hair toxic trace elements content in occupationally non-exposed Russian population. Environ. Toxicol. Pharmacol. 2015, 40, 18–21. [Google Scholar] [CrossRef] [PubMed]
- Tamburo, E.; Dongarrà, G.; Varrica, D.; D’Andrea, D. Trace elements in hair of urban schoolboys: A diagnostic tool in environmental risk assessment. Geophys. Res. Abstr. 2011, 13, 1157. [Google Scholar]
- Razagui, I.B.; Ghribi, I. Maternal and neonatal scalp hair concentrations of zinc, copper, cadmium, and lead: Relationship to some lifestyle factors. Biol. Trace Elem. Res. 2005, 106, 1–28. [Google Scholar] [CrossRef]
- Torres-Agustín, R.; Rodríguez-Agudelo, Y.; Schilmann, A.; Solís-Vivanco, R.; Montes, S.; Riojas-Rodríguez, H.; Cortez-Lugo, M.; Ríos, C. Effect of environmental manganese exposure on verbal learning and memory in Mexican children. Environ. Res. 2013, 121, 39–44. [Google Scholar] [CrossRef]
- Carvalho, C.F.; Menezes-Filho, J.A.; de Matos, V.P.; Bessa, J.R.; Coelho-Santos, J.; Viana, G.F.; Argollo, N.; Abreu, N. Elevated airborne manganese and low executive function in school-aged children in Brazil. Neurotoxicology 2014, 45, 301–308. [Google Scholar] [CrossRef]
- Hassani, H.; Golbabaei, F.; Shirkhanloo, H.; Tehrani-Doust, M. Relations of biomarkers of manganese exposure and neuropsychological effects among welders and ferroalloy smelters. Ind. Health 2016, 54, 79–86. [Google Scholar] [CrossRef]
- Laohaudomchok, W.; Lin, X.; Herrick, R.F.; Fang, S.C.; Cavallari, J.M.; Christiani, D.C.; Weisskopf, M.G. Toenail, blood, and urine as biomarkers of manganese exposure. J. Occup. Environ. Med. 2011, 53, 506–510. [Google Scholar] [CrossRef] [PubMed]
- Toxicological Profile for Manganese. U.S. Department of Health and Human Services, Public Health Service Agency for Toxic Substances and Disease Registry, September 2012. Available online: https://www.atsdr.cdc.gov/toxprofiles/tp151.pdf (accessed on 15 March 2023).
- Versieck, J.; Cornelis, R.; Lemey, G.; De Rudder, J. Determination of manganese in whole blood and serum. Clin. Chem. 1980, 26, 531–532. [Google Scholar] [CrossRef]
- Heydorn, K.; Nørgård, K. Analysis of precision of activation-analysis methods. Talanta 1973, 20, 835–842. [Google Scholar] [CrossRef]
- Kanabrocki, E.L.; Fields, T.; Decker, C.F.; Case, L.F.; Miller, E.B.; Kaplan, E.; Oester, Y.T. Neutron activation studies of bio-logical fluids: Manganese and copper. Int. J. Appl. Radiat. Isot. 1964, 15, 175–190. [Google Scholar] [CrossRef] [PubMed]
- Mayo Clinic Laboratories. Test Definition: MNB. Available online: https://www.mayocliniclabs.com/api/sitecore/TestCatalog/DownloadTestCatalog?testId=89120 (accessed on 5 January 2023).
- Goullé, J.P.; Mahieu, L.; Castermant, J.; Neveu, N.; Bonneau, L.; Lainé, G.; Bouige, D.; Lacroix, C. Metal and metalloid multi-elementary ICP-MS validation in whole blood, plasma, urine and hair. Reference values. Forensic Sci. Int. 2005, 153, 39–44. [Google Scholar] [CrossRef] [PubMed]
- Goullé, J.-P.; Saussereau, E.; Mahieu, L.; Guerbet, M. Current role of ICP–MS in clinical toxicology and forensic toxicology: A metallic profile. Bioanalysis 2014, 6, 2245–2259. [Google Scholar] [CrossRef]
- Zheng, W.; Fu, S.X.; Dydak, U.; Cowan, D.M. Biomarkers of manganese intoxication. Neurotoxicology 2011, 32, 1–8. [Google Scholar] [CrossRef]
- Marreilha Dos Santos, A.P.; Lopes Santos, M.; Batoréu, M.C.; Aschner, M. Prolactin is a peripheral marker of manganese neurotoxicity. Brain Res. 2011, 1382, 282–290. [Google Scholar] [CrossRef]
- Chavan, S.; Bhat, R.; Nandoskar, A.; Bhat, P.; Rokade, S.R.; Mohire, R.R.; Patil, S.N. Determination of Manganese in Serum Using GFAAS: Serum Reference Values for Manganese in the Adolescent Girls of the DERVAN Cohort. Indian J. Clin. Biochem. 2022, 37, 487–493. [Google Scholar] [CrossRef]
- Tuakashikila, Y.M.; Mata, H.M.; Kabamba, M.M.; Malumba, A.M.; Tuakuila, J.K. Reference intervals for cd, hg, Mn and Pb in the general children population (3–14 years) of Kinshasa, Democratic Republic of Congo (DRC) between June 2019 and June 2020. Arch. Public Health 2023, 81, 40. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.; Lee, B.K. Iron deficiency increases blood manganese level in the Korean general population according to KNHANES 2008. Neurotoxicology 2011, 32, 247–254. [Google Scholar] [CrossRef]
- National Center for Environmental Health (U.S.); Division of Laboratory Sciences; National Center for Health Statistics (U.S.); National Health and Nutrition Examination Survey (U.S.). Fourth National Report on Human Exposure to Environmental Chemicals. Updated Tables, March 2021. Available online: https://stacks.cdc.gov/view/cdc/105344 (accessed on 30 March 2021).
- Baldwin, M.; Mergler, D.; Larribe, F.; Bélanger, S.; Tardif, R.; Bilodeau, L.; Hudnell, K. Bioindicator and exposure data for a population based study of manganese. Neurotoxicology 1999, 20, 343–353. [Google Scholar] [PubMed]
- Lee, B.K.; Kim, Y. Effects of menopause on blood manganese levels in women: Analysis of 2008–2009 Korean National Health and Nutrition Examination Survey data. Neurotoxicology 2012, 33, 401–405. [Google Scholar] [CrossRef] [PubMed]
- Spencer, A. Whole blood manganese levels in pregnancy and the neonate. Nutrition 1999, 15, 731–734. [Google Scholar] [CrossRef] [PubMed]
- Zota, A.R.; Ettinger, A.S.; Bouchard, M.; Amarasiriwardena, C.J.; Schwartz, J.; Hu, H.; Wright, R.O. Maternal blood manganese levels and infant birth weight. Epidemiology 2009, 20, 367–373. [Google Scholar] [CrossRef]
- Freire, C.; Koifman, R.J.; Fujimoto, D.; de Oliveira Souza, V.C.; Barbosa, F., Jr.; Koifman, S. Reference values of cadmium, arsenic and manganese in blood and factors associated with exposure levels among adult population of Rio Branco, Acre, Brazil. Chemosphere 2015, 128, 70–78. [Google Scholar] [CrossRef]
- Batyrova, G.; Tlegenova, Z.; Kononets, V.; Umarova, G.; Bazargaliyev, Y.; Taskozhina, G.; Kereyeva, N.; Umarov, Y. Content of Essential Trace Elements in the Hair of Residents of the Caspian Region of the Republic of Kazakhstan Who Recovered from COVID-19. Diagnostics 2022, 12, 2734. [Google Scholar] [CrossRef] [PubMed]
- Pleban, P.A.; Pearson, K.H. Determination of manganese in whole blood and serum. Clin. Chem. 1979, 25, 1915–1918. [Google Scholar] [CrossRef] [PubMed]
- Skalny, A.V.; Skalnaya, M.G.; Tinkov, A.A.; Serebryansky, E.P.; Demidov, V.A.; Lobanova, Y.N.; Grabeklis, A.R.; Berezkina, E.S.; Gryazeva, I.V.; Skalny, A.A.; et al. Hair concentration of essential trace elements in adult non-exposed Russian population. Environ. Monit. Assess 2015, 187, 677. [Google Scholar] [CrossRef]
- Goullé, J.P.; Le Roux, P.; Castanet, M.; Mahieu, L.; Guyet-Job, S.; Guerbet, M. Metallic Profile of Whole Blood and Plasma in a Series of 99 Healthy Children. J. Anal. Toxicol. 2015, 39, 707–713. [Google Scholar] [CrossRef]
- Cesbron, A.; Saussereau, E.; Mahieu, L.; Couland, I.; Guerbet, M.; Goullé, J.P. Metallic profile of whole blood and plasma in a series of 106 healthy volunteers. J. Anal. Toxicol. 2013, 37, 401–405. [Google Scholar] [CrossRef] [PubMed]
- Heitland, P.; Köster, H.D. Biomonitoring of 37 trace elements in blood samples from inhabitants of northern Germany by ICP-MS. J. Trace Elem. Med. Biol. 2006, 20, 253–262. [Google Scholar] [CrossRef]
- Schultze, B.; Lind, P.M.; Larsson, A.; Lind, L. Whole blood and serum concentrations of metals in a Swedish population-based sample. Scand. J. Clin. Lab. Investig. 2013, 74, 143–148. [Google Scholar] [CrossRef]
- Dressler, V.L.; Pozebon, D.; Mesko, M.F.; Matusch, A.; Kumtabtim, U.; Wu, B.; Sabine Becker, J. Biomonitoring of essential and toxic metals in single hair using on-line solution-based calibration in laser ablation inductively coupled plasma mass spectrometry. Talanta 2010, 82, 1770–1777. [Google Scholar] [CrossRef]
- Nunes, J.A.; Batista, B.L.; Rodrigues, J.L.; Caldas, N.M.; Neto, J.A.; Barbosa, F., Jr. A simple method based on ICP-MS for estimation of background levels of arsenic, cadmium, copper, manganese, nickel, lead, and selenium in blood of the Brazilian population. J. Toxicol. Environ. Health A 2010, 73, 878–887. [Google Scholar] [CrossRef]
- Tuakuila, J.; Kabamba, M.; Mata, H.; Mata, G. Toxic and essential elements in children’s blood (<6 years) from Kinshasa, DRC (the Democratic Republic of Congo). J. Trace Elem. Med. Biol. 2014, 28, 45–49. [Google Scholar] [CrossRef] [PubMed]
- Moreno, M.E.; Acosta-Saavedra, L.C.; Meza-Figueroa, D.; Vera, E.; Cebrian, M.E.; Ostrosky-Wegman, P.; Calderon-Aranda, E.S. Biomonitoring of metal in children living in a mine tailings zone in Southern Mexico: A pilot study. Int. J. Hyg. Environ. Health 2010, 213, 252–258. [Google Scholar] [CrossRef] [PubMed]
- Knerr, I.; Blessing, H.; Seyferth, S.; Watling, R.J.; Chaudhri, M.A. Evaluation of plasma trace element and mineral status in children and adolescents with phenylketonuria using data from inductively-coupled-plasma atomic emission and mass spectrometric analysis. Ann. Nutr. Metab. 2013, 63, 168–173. [Google Scholar] [CrossRef] [PubMed]
- Kimura, M.; Ujihara, M.; Yokoi, K. Tissue manganese levels and liver pyruvate carboxylase activity in magnesium-deficient rats. Biol. Trace Elem. Res. 1996, 52, 171–179. [Google Scholar] [CrossRef]
- Kies, C. Bioavailability of Manganese. In Manganese in Health and Disease; Klimis-Tavantzis, D., Ed.; CRC Press, Inc.: Boca Raton, FL, USA, 1994; pp. 39–58. [Google Scholar]
- Erikson, K.M.; Syversen, T.; Steinnes, E.; Aschner, M. Globus pallidus: A target brain region for divalent metal accumulation associated with dietary iron deficiency. J. Nutr. Biochem. 2004, 15, 335–341. [Google Scholar] [CrossRef] [PubMed]
- Gunshin, H.; Allerson, C.R.; Polycarpou-Schwarz, M.; Rofts, A.; Rogers, J.T.; Kishi, F.; Hentze, M.W.; Rouault, T.A.; Andrews, N.C.; Hediger, M.A. Iron-dependent regulation of the divalent metal ion transporter. FEBS Lett. 2001, 509, 309–316. [Google Scholar] [CrossRef] [PubMed]
- Hurley, L.S.; Keen, C.L. Manganese. In Trace Elements in Human Health and Animal Nutrition; Underwood, E., Mertz, W., Eds.; Academic Press: New York, NY, USA, 1987; pp. 185–223. [Google Scholar]
- Mackenzie, B.; Garrick, M.D. Iron Imports. II. Iron uptake at the apical membrane in the intestine. Am. J. Physiol. Gastrointest. Liver Physiol. 2005, 289, G981–G986. [Google Scholar] [CrossRef]
- Davis, C.D.; Wolf, T.L.; Greger, J.L. Varying levels of manganese and iron affect absorption and gut endogenous losses of manganese by rats. J. Nutr. 1992, 122, 1300–1308. [Google Scholar] [CrossRef]
- Mena, I.; Horiuchi, K.; Burke, K.; Cotzias, G.C. Chronic manganese poisoning. Individual susceptibility and absorption of iron. Neurology 1969, 19, 1000–1006. [Google Scholar] [CrossRef]
- Finley, J.W. Manganese absorption and retention by young women is associated with serum ferritin concentration. Am. J. Clin. Nutr. 1999, 70, 37–43. [Google Scholar] [CrossRef]
- Kim, Y.; Park, J.K.; Choi, Y.; Yoo, C.I.; Lee, C.R.; Lee, H.; Lee, J.H.; Kim, S.R.; Jeong, T.H.; Yoon, C.S.; et al. Blood manganese concentration is elevated in iron deficiency anemia patients, whereas globus pallidus signal intensity is minimally affected. Neurotoxicology 2005, 26, 107–111. [Google Scholar] [CrossRef]
- Meltzer, H.M.; Brantsaeter, A.L.; Borch-Iohnsen, B.; Ellingsen, D.G.; Alexander, J.; Thomassen, Y.; Stigum, H.; Ydersbond, T.A. Low iron stores are related to higher blood concentrations of manganese, cobalt and cadmium in non-smoking, Norwegian women in the HUNT 2 study. Environ. Res. 2010, 110, 497–504. [Google Scholar] [CrossRef]
- Park, S.; Sim, C.S.; Lee, H.; Kim, Y. Blood manganese concentration is elevated in infants with iron deficiency. Biol. Trace Elem. Res. 2013, 155, 184–189. [Google Scholar] [CrossRef] [PubMed]
- Ye, Q.; Park, J.E.; Gugnani, K.; Betharia, S.; Pino-Figueroa, A.; Kim, J. Influence of iron metabolism on manganese transport and toxicity. Metallomics 2017, 9, 1028–1046. [Google Scholar] [CrossRef]
- Davis, C.D.; Greger, J.L. Longitudinal changes of manganese-dependent superoxide dismutase and other indexes of manganese and iron status in women. Am. J. Clin. Nutr. 1992, 55, 747–752. [Google Scholar] [CrossRef] [PubMed]
- Chen, P.; Chakraborty, S.; Mukhopadhyay, S.; Lee, E.; Paoliello, M.M.; Bowman, A.B.; Aschner, M. Manganese homeostasis in the nervous system. J. Neurochem. 2015, 134, 601–610. [Google Scholar] [CrossRef]
- Kim, Y.; Lobdell, D.T.; Wright, C.W.; Gocheva, V.V.; Hudgens, E.; Bowler, R.M. Blood Metal Concentrations of Manganese, Lead, and Cadmium in Relation to Serum Ferritin Levels in Ohio Residents. Biol. Trace Elem. Res. 2015, 165, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Rahman, M.A.; Rahman, B.; Ahmed, N. High blood manganese in iron-deficient children in Karachi. Public Health Nutr. 2013, 16, 1677–1683. [Google Scholar] [CrossRef]
- Lee, B.K.; Kim, Y. Sex-specific Profiles of Blood Metal Levels Associated with Metal-Iron Interactions. Saf. Health Work 2014, 5, 113–117. [Google Scholar] [CrossRef]
- Takser, L.; Lafond, J.; Bouchard, M.; St-Amour, G.; Mergler, D. Manganese levels during pregnancy and at birth: Relation to environmental factors and smoking in a Southwest Quebec population. Environ. Res. 2004, 95, 119–125. [Google Scholar] [CrossRef]
- Aschner, M.; Dorman, D.C. Manganese: Pharmacokinetics and molecular mechanisms of brain uptake. Toxicol. Rev. 2006, 25, 147–154. [Google Scholar] [CrossRef]
- Diessl, J.; Berndtsson, J.; Broeskamp, F.; Habernig, L.; Kohler, V.; Vazquez-Calvo, C.; Nandy, A.; Peselj, C.; Drobysheva, S.; Pelosi, L.; et al. Manganese-driven CoQ deficiency. Nat. Commun. 2022, 13, 6061. [Google Scholar] [CrossRef] [PubMed]
- Sandstead, H.H. Requirements and toxicity of essential trace elements, illustrated by zinc and copper. Am. J. Clin. Nutr. 1995, 61, 621S–624S. [Google Scholar] [CrossRef] [PubMed]
- Keen, C.L.; Ensunsa, J.L.; Clegg, M.S. Manganese metabolism in animals and humans including the toxicity of manganese. Met. Ions Biol. Syst. 2000, 37, 89–121. [Google Scholar] [PubMed]
- Kwik-Uribe, C.L.; Gietzen, D.; German, J.B.; Golub, M.S.; Keen, C.L. Chronic marginal iron intakes during early development in mice result in persistent changes in dopamine metabolism and myelin composition. J. Nutr. 2000, 130, 2821–2830. [Google Scholar] [CrossRef]
- Wasserman, G.A.; Liu, X.; Parvez, F.; Ahsan, H.; Levy, D.; Factor-Litvak, P.; Kline, J.; van Geen, A.; Slavkovich, V.; LoIacono, N.J.; et al. Water manganese exposure and children’s intellectual function in Araihazar, Bangladesh. Environ. Health Perspect. 2006, 114, 124–129. [Google Scholar] [CrossRef]
- Guilarte, T.R. Manganese and Parkinson’s disease: A critical review and new findings. Environ. Health Perspect. 2010, 118, 1071–1080. [Google Scholar] [CrossRef]
- Johnson, P.; Lykken, G. Manganese and calcium absorption and balance in young women fed diets with varying amounts of manganese and calcium. J. Trace Elem. Exp. Med. 1991, 4, 19–35. [Google Scholar]
- Macmillan-Crow, L.A.; Cruthirds, D.L. Invited review: Manganese superoxide dismutase in disease. Free Radic. Res. 2001, 34, 325–336. [Google Scholar] [CrossRef]
- Ilies, M.; Di Costanzo, L.; North, M.L.; Scott, J.A.; Christianson, D.W. 2-aminoimidazole amino acids as inhibitors of the binuclear manganese metalloenzyme human arginase I. J. Med. Chem. 2010, 53, 4266–4276. [Google Scholar] [CrossRef]
- Hamberg, M.; Su, C.; Oliw, E. Manganese lipoxygenase. Discovery of a bisallylic hydroperoxide as product and intermediate in a lipoxygenase reaction. J. Biol. Chem. 1998, 273, 13080–13088. [Google Scholar] [CrossRef]
- Spodenkiewicz, M.; Spodenkiewicz, M.; Cleary, M.; Massier, M.; Fitsialos, G.; Cottin, V.; Jouret, G.; Poirsier, C.; Doco-Fenzy, M.; Lèbre, A.-S. Clinical Genetics of Prolidase Deficiency: An Updated Review. Biology 2020, 9, 108. [Google Scholar] [CrossRef] [PubMed]
- Nordhøy, W.; Anthonsen, H.W.; Bruvold, M.; Jynge, P.; Krane, J.; Brurok, H. Manganese ions as intracellular contrast agents: Proton relaxation and calcium interactions in rat myocardium. NMR Biomed. 2003, 16, 82–95. [Google Scholar] [CrossRef] [PubMed]
- Kalea, A.Z.; Harris, P.D.; Klimis-Zacas, D.J. Dietary manganese suppresses alpha1 adrenergic receptor-mediated vascular contraction. J. Nutr. Biochem. 2005, 16, 44–49. [Google Scholar] [CrossRef] [PubMed]
- Au, C.; Benedetto, A.; Aschner, M. Manganese transport in eukaryotes: The role of DMT1. Neurotoxicology 2008, 29, 569–576. [Google Scholar] [CrossRef] [PubMed]
- Aschner, M.; Gannon, M. Manganese (Mn) transport across the rat blood-brain barrier: Saturable and transferrin-dependent transport mechanisms. Brain Res. Bull. 1994, 33, 345–349. [Google Scholar] [CrossRef]
- Davidsson, L.; Lönnerdal, B.; Sandström, B.; Kunz, C.; Keen, C.L. Identification of transferrin as the major plasma carrier protein for manganese introduced orally or intravenously or after in vitro addition in the rat. J. Nutr. 1989, 119, 1461–1464. [Google Scholar] [CrossRef]
- Garrick, M.D.; Kuo, H.C.; Vargas, F.; Singleton, S.; Zhao, L.; Smith, J.J.; Paradkar, P.; Roth, J.A.; Garrick, L.M. Comparison of mammalian cell lines expressing distinct isoforms of divalent metal transporter 1 in a tetracycline-regulated fashion. Biochem. J. 2006, 398, 539–546. [Google Scholar] [CrossRef]
- Gunshin, H.; Mackenzie, B.; Berger, U.V.; Gunshin, Y.; Romero, M.F.; Boron, W.F.; Nussberger, S.; Gollan, J.L.; Hediger, M.A. Cloning and characterization of a mammalian proton-coupled metal-ion transporter. Nature 1997, 388, 482–488. [Google Scholar] [CrossRef] [PubMed]
- He, L.; Girijashanker, K.; Dalton, T.P.; Reed, J.; Li, H.; Soleimani, M.; Nebert, D.W. ZIP8, member of the solute-carrier-39 (SLC39) metal-transporter family: Characterization of transporter properties. Mol. Pharmacol. 2006, 70, 171–180. [Google Scholar] [CrossRef]
- Lucaciu, C.M.; Dragu, C.; Copăescu, L.; Morariu, V.V. Manganese transport through human erythrocyte membranes—An EPR Study. Biochim. Biophys. Acta. 1997, 1328, 90–98. [Google Scholar] [CrossRef]
- Huang, E.; Ong, W.Y.; Connor, J.R. Distribution of divalent metal transporter-1 in the monkey basal ganglia. Neuroscience 2004, 128, 487–496. [Google Scholar] [CrossRef]
- Fu, S.; O’Neal, S.; Hong, L.; Jiang, W.; Zheng, W. Elevated adult neurogenesis in brain subventricular zone following in vivo manganese exposure: Roles of copper and DMT1. Toxicol. Sci. 2015, 143, 482–498. [Google Scholar] [CrossRef]
- Wang, X.S.; Ong, W.Y.; Connor, J.R. A light and electron microscopic study of the iron transporter protein DMT-1 in the monkey cerebral neocortex and hippocampus. J. Neurocytol. 2001, 30, 353–360. [Google Scholar] [CrossRef]
- Thompson, K.; Molina, R.M.; Donaghey, T.; Schwob, J.E.; Brain, J.D.; Wessling-Resnick, M. Olfactory uptake of manganese requires DMT1 and is enhanced by anemia. FASEB J. 2007, 21, 223–230. [Google Scholar] [CrossRef]
- Scheiber, I.F.; Alarcon, N.O.; Zhao, N. Manganese Uptake by A549 Cells is Mediated by Both ZIP8 and ZIP14. Nutrients 2019, 11, 1473. [Google Scholar] [CrossRef] [PubMed]
- Steimle, B.L.; Smith, F.M.; Kosman, D.J. The solute carriers ZIP8 and ZIP14 regulate manganese accumulation in brain microvascular endothelial cells and control brain manganese levels. J. Biol. Chem. 2019, 294, 19197–19208. [Google Scholar] [CrossRef] [PubMed]
- Tuschl, K.; Meyer, E.; Valdivia, L.E.; Zhao, N.; Dadswell, C.; Abdul-Sada, A.; Hung, C.Y.; Simpson, M.A.; Chong, W.K.; Jacques, T.S.; et al. Mutations in SLC39A14 disrupt manganese homeostasis and cause childhood-onset parkinsonism-dystonia. Nat. Commun. 2016, 7, 11601. [Google Scholar] [CrossRef]
- Jenkitkasemwong, S.; Akinyode, A.; Paulus, E.; Weiskirchen, R.; Hojyo, S.; Fukada, T.; Giraldo, G.; Schrier, J.; Garcia, A.; Janus, C.; et al. SLC39A14 deficiency alters manganese homeostasis and excretion resulting in brain manganese accumulation and motor deficits in mice. Proc. Natl. Acad. Sci. USA 2018, 115, E1769–E1778. [Google Scholar] [CrossRef] [PubMed]
- Marti-Sanchez, L.; Ortigoza-Escobar, J.D.; Darling, A.; Villaronga, M.; Baide, H.; Molero-Luis, M.; Batllori, M.; Vanegas, M.I.; Muchart, J.; Aquino, L.; et al. Hypermanganesemia due to mutations in SLC39A14: Further insights into Mn deposition in the central nervous system. Orphanet. J. Rare Dis. 2018, 13, 28. [Google Scholar] [CrossRef]
- Choi, E.K.; Nguyen, T.T.; Iwase, S.; Seo, Y.A. Ferroportin disease mutations influence manganese accumulation and cytotoxicity. FASEB J. 2019, 33, 2228–2240. [Google Scholar] [CrossRef] [PubMed]
- Tuschl, K.; Mills, P.B.; Parsons, H.; Malone, M.; Fowler, D.; Bitner-Glindzicz, M.; Clayton, P.T. Hepatic cirrhosis, dystonia, polycythaemia and hypermanganesaemia—A new metabolic disorder. J. Inherit. Metab. Dis. 2008, 31, 151–163. [Google Scholar] [CrossRef]
- Lechpammer, M.; Clegg, M.S.; Muzar, Z.; Huebner, P.A.; Jin, L.; Gospe, S.M. Pathology of inherited manganese transporter deficiency. Ann. Neurol. 2014, 75, 608–612. [Google Scholar] [CrossRef]
- Quadri, M.; Federico, A.; Zhao, T.; Breedveld, G.J.; Battisti, C.; Delnooz, C.; Severijnen, L.; Di Toro Mammarella, L.; Mignarri, A.; Monti, L. Mutations in SLC30A10 cause parkinsonism and dystonia with hypermanganesemia, polycythemia, and chronic liver disease. Am. J. Hum. Genet. 2012, 90, 467–477. [Google Scholar] [CrossRef]
- Tuschl, K.; Clayton, P.T.; Gospe, S.M., Jr.; Gulab, S.; Ibrahim, S.; Singhi, P.; Aulakh, R.; Ribeiro, R.T.; Barsottini, O.G.; Zaki, M.S. Syndrome of hepatic cirrhosis, dystonia, polycythemia, and hypermanganesemia caused by mutations in SLC30A10, a manganese transporter in man. Am. J. Hum. Genet. 2012, 90, 457–466. [Google Scholar] [CrossRef]
- Murphy, K.E.; Cottle, L.; Gysbers, A.M.; Cooper, A.A.; Halliday, G.M. ATP13A2 (PARK9) protein levels are reduced in brain tissue of cases with Lewy bodies. Acta Neuropathol. Commun. 2013, 1, 11. [Google Scholar] [CrossRef]
- Robison, G.; Sullivan, B.; Cannon, J.R.; Pushkar, Y. Identification of dopaminergic neurons of the substantia nigra pars compacta as a target of manganese accumulation. Metallomics 2015, 7, 748–755. [Google Scholar] [CrossRef]
- Carmona, A.; Roudeau, S.; Perrin, L.; Veronesi, G.; Ortega, R. Environmental manganese compounds accumulate as Mn(II) within the Golgi apparatus of dopamine cells: Relationship between speciation, subcellular distribution, and cytotoxicity. Metallomics 2014, 6, 822–832. [Google Scholar] [CrossRef]
- Kalia, K.; Jiang, W.; Zheng, W. Manganese accumulates primarily in nuclei of cultured brain cells. Neurotoxicology 2008, 29, 466–470. [Google Scholar] [CrossRef]
- Jensen, L.T.; Carroll, M.C.; Hall, M.D.; Harvey, C.J.; Beese, S.E.; Culotta, V.C. Down-regulation of a manganese transporter in the face of metal toxicity. Mol. Biol. Cell. 2009, 20, 2810–2819. [Google Scholar] [CrossRef]
- Au, C.; Benedetto, A.; Anderson, J.; Labrousse, A.; Erikson, K.; Ewbank, J.J.; Aschner, M. SMF-1, SMF-2 and SMF-3 DMT1 orthologues regulate and are regulated differentially by manganese levels in C. elegans. PLoS ONE 2009, 4, e7792. [Google Scholar] [CrossRef]
- Bélanger, M.; Allaman, I.; Magistretti, P.J. Brain energy metabolism: Focus on astrocyte-neuron metabolic cooperation. Cell Metab. 2011, 14, 724–738. [Google Scholar] [CrossRef]
- Cobley, J.N.; Fiorello, M.L.; Bailey, D.M. 13 reasons why the brain is susceptible to oxidative stress. Redox Biol. 2018, 15, 490–503. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Zhao, B. Oxidative stress and the pathogenesis of Alzheimer’s disease. Oxid. Med. Cell Longev. 2013, 2013, 316523. [Google Scholar] [CrossRef]
- Chtourou, Y.; Trabelsi, K.; Fetoui, H.; Mkannez, G.; Kallel, H.; Zeghal, N. Manganese induces oxidative stress, redox state unbalance and disrupts membrane bound ATPases on murine neuroblastoma cells in vitro: Protective role of silymarin. Neurochem. Res. 2011, 36, 1546–1557. [Google Scholar] [CrossRef]
- Omar, R.A.; Chyan, Y.J.; Andorn, A.C.; Poeggeler, B.; Robakis, N.K.; Pappolla, M.A. Increased Expression but Reduced Activity of Antioxidant Enzymes in Alzheimer’s Disease. J. Alzheimers Dis. 1999, 1, 139–145. [Google Scholar] [CrossRef] [PubMed]
- Spranger, M.; Schwab, S.; Desiderato, S.; Bonmann, E.; Krieger, D.; Fandrey, J. Manganese augments nitric oxide synthesis in murine astrocytes: A new pathogenetic mechanism in manganism? Exp. Neurol. 1998, 149, 277–283. [Google Scholar] [CrossRef] [PubMed]
- Li, S.J.; Qin, W.X.; Peng, D.J.; Yuan, Z.X.; He, S.N.; Luo, Y.N.; Aschner, M.; Jiang, Y.M.; Liang, D.Y.; Xie, B.Y.; et al. Sodium P-aminosalicylic acid inhibits sub-chronic manganese-induced neuroinflammation in rats by modulating MAPK and COX-2. Neurotoxicology 2018, 64, 219–229. [Google Scholar] [CrossRef]
- Popichak, K.A.; Afzali, M.F.; Kirkley, K.S.; Tjalkens, R.B. Glial-neuronal signaling mechanisms underlying the neuroinflammatory effects of manganese. J. Neuroinflammation 2018, 15, 324. [Google Scholar] [CrossRef]
- Park, E.; Chun, H.S. Melatonin Attenuates Manganese and Lipopolysaccharide-Induced Inflammatory Activation of BV2 Microglia. Neurochem. Res. 2017, 42, 656–666. [Google Scholar] [CrossRef]
- Barhoumi, R.; Faske, J.; Liu, X.; Tjalkens, R.B. Manganese potentiates lipopolysaccharide-induced expression of NOS2 in C6 glioma cells through mitochondrial-dependent activation of nuclear factor kappaB. Brain Res. Mol. Brain Res. 2004, 122, 167–179. [Google Scholar] [CrossRef]
- Filipov, N.M.; Seegal, R.F.; Lawrence, D.A. Manganese potentiates in vitro production of proinflammatory cytokines and nitric oxide by microglia through a nuclear factor kappa B-dependent mechanism. Toxicol. Sci. 2005, 84, 139–148. [Google Scholar] [CrossRef]
- Moreno, J.A.; Streifel, K.M.; Sullivan, K.A.; Hanneman, W.H.; Tjalkens, R.B. Manganese-induced NF-kappaB activation and nitrosative stress is decreased by estrogen in juvenile mice. Toxicol. Sci. 2011, 122, 121–133. [Google Scholar] [CrossRef]
- Desole, M.S.; Serra, P.A.; Esposito, G.; Delogu, M.R.; Migheli, R.; Fresu, L.; Rocchitta, G.; Miele, M. Glutathione deficiency potentiates manganese-induced increases in compounds associated with high-energy phosphate degradation in discrete brain areas of young and aged rats. Aging 2000, 12, 470–477. [Google Scholar] [CrossRef]
- Roth, J.A.; Li, Z.; Sridhar, S.; Khoshbouei, H. The effect of manganese on dopamine toxicity and dopamine transporter (DAT) in control and DAT transfected HEK cells. Neurotoxicology 2013, 35, 121–128. [Google Scholar] [CrossRef]
- Benedetto, A.; Au, C.; Avila, D.S.; Milatovic, D.; Aschner, M. Extracellular dopamine potentiates mn-induced oxidative stress, lifespan reduction, and dopaminergic neurodegeneration in a BLI-3-dependent manner in Caenorhabditis elegans. PLoS Genet. 2010, 6, e1001084. [Google Scholar] [CrossRef]
- Milatovic, D.; Zaja-Milatovic, S.; Gupta, R.C.; Yu, Y.; Aschner, M. Oxidative damage and neurodegeneration in manganese-induced neurotoxicity. Toxicol. Appl. Pharmacol. 2009, 240, 219–225. [Google Scholar] [CrossRef]
- Milatovic, D.; Yin, Z.; Gupta, R.C.; Sidoryk, M.; Albrecht, J.; Aschner, J.L.; Aschner, M. Manganese induces oxidative impairment in cultured rat astrocytes. Toxicol. Sci. 2007, 98, 198–205. [Google Scholar] [CrossRef]
- Aluru, N.; Karchner, S.I.; Franks, D.G.; Nacci, D.; Champlin, D.; Hahn, M.E. Targeted mutagenesis of aryl hydrocarbon receptor 2a and 2b genes in Atlantic killifish (Fundulus heteroclitus). Aquat. Toxicol. 2015, 158, 192–201. [Google Scholar] [CrossRef] [PubMed]
- Stephenson, A.P.; Schneider, J.A.; Nelson, B.C.; Atha, D.H.; Jain, A.; Soliman, K.F.; Aschner, M.; Mazzio, E.; Renee Reams, R. Manganese-induced oxidative DNA damage in neuronal SH-SY5Y cells: Attenuation of thymine base lesions by glutathione and N-acetylcysteine. Toxicol. Lett. 2013, 218, 299–307. [Google Scholar] [CrossRef] [PubMed]
- Wedler, F.C.; Denman, R.B. Glutamine synthetase: The major Mn(II) enzyme in mammalian brain. Curr. Top. Cell Regul. 1984, 24, 153–169. [Google Scholar] [CrossRef] [PubMed]
- Sidoryk-Wegrzynowicz, M.; Aschner, M. Role of astrocytes in manganese mediated neurotoxicity. BMC Pharmacol. Toxicol. 2013, 14, 23. [Google Scholar] [CrossRef]
- Harischandra, D.S.; Ghaisas, S.; Zenitsky, G.; Jin, H.; Kanthasamy, A.; Anantharam, V.; Kanthasamy, A.G. Manganese-Induced Neurotoxicity: New Insights Into the Triad of Protein Misfolding, Mitochondrial Impairment, and Neuroinflammation. Front. Neurosci. 2019, 13, 654. [Google Scholar] [CrossRef]
- Erikson, K.M.; Aschner, M. Increased manganese uptake by primary astrocyte cultures with altered iron status is mediated primarily by divalent metal transporter. Neurotoxicology 2006, 27, 125–130. [Google Scholar] [CrossRef] [PubMed]
- Huang, C.C. Parkinsonism induced by chronic manganese intoxication--an experience in Taiwan. Chang Gung Med. J. 2007, 30, 385–395. [Google Scholar]
- Perl, D.P.; Olanow, C.W. The neuropathology of manganese-induced Parkinsonism. J. Neuropathol. Exp. Neurol. 2007, 66, 675–682. [Google Scholar] [CrossRef]
- Moreno, J.A.; Streifel, K.M.; Sullivan, K.A.; Legare, M.E.; Tjalkens, R.B. Developmental exposure to manganese increases adult susceptibility to inflammatory activation of glia and neuronal protein nitration. Toxicol. Sci. 2009, 112, 405–415. [Google Scholar] [CrossRef] [PubMed]
- Zhang, P.; Hatter, A.; Liu, B. Manganese chloride stimulates rat microglia to release hydrogen peroxide. Toxicol. Lett. 2007, 173, 88–100. [Google Scholar] [CrossRef] [PubMed]
- Crittenden, P.L.; Filipov, N.M. Manganese-induced potentiation of _in vitro_ proinflammatory cytokine production by activated microglial cells is associated with persistent activation of p38 MAPK. Toxicol. Vitr. 2008, 22, 18–27. [Google Scholar] [CrossRef] [PubMed]
- Kirkley, K.S.; Popichak, K.A.; Afzali, M.F.; Legare, M.E.; Tjalkens, R.B. Microglia amplify inflammatory activation of astrocytes in manganese neurotoxicity. J. Neuroinflammation 2017, 14, 99. [Google Scholar] [CrossRef]
- Harischandra, D.S.; Rokad, D.; Neal, M.L.; Ghaisas, S.; Manne, S.; Sarkar, S.; Panicker, N.; Zenitsky, G.; Jin, H.; Lewis, M.; et al. Manganese promotes the aggregation and prion-like cell-to-cell exosomal transmission of α-synuclein. Sci. Signal. 2019, 12, eaau4543. [Google Scholar] [CrossRef]
- Chen, J.; Su, P.; Luo, W.; Chen, J. Role of LRRK2 in manganese-induced neuroinflammation and microglial autophagy. Biochem. Biophys. Res. Commun. 2018, 498, 171–177. [Google Scholar] [CrossRef] [PubMed]
- Sarkar, S.; Rokad, D.; Malovic, E.; Luo, J.; Harischandra, D.S.; Jin, H.; Anantharam, V.; Huang, X.; Lewis, M.; Kanthasamy, A.; et al. Manganese activates NLRP3 inflammasome signaling and propagates exosomal release of ASC in microglial cells. Sci. Signal. 2019, 12, eaat9900. [Google Scholar] [CrossRef]
- Rudgalvyte, M.; Peltonen, J.; Lakso, M.; Nass, R.; Wong, G. RNA-Seq Reveals Acute Manganese Exposure Increases Endoplasmic Reticulum Related and Lipocalin mRNAs in Caenorhabditis elegans. J. Biochem. Mol. Toxicol. 2016, 30, 97–105. [Google Scholar] [CrossRef]
- Wang, T.; Li, X.; Yang, D.; Zhang, H.; Zhao, P.; Fu, J.; Yao, B.; Zhou, Z. ER stress and ER stress-mediated apoptosis are involved in manganese-induced neurotoxicity in the rat striatum in vivo. Neurotoxicology 2015, 48, 109–119. [Google Scholar] [CrossRef]
- Chun, H.S.; Lee, H.; Son, J.H. Manganese induces endoplasmic reticulum (ER) stress and activates multiple caspases in nigral dopaminergic neuronal cells, SN4741. Neurosci. Lett. 2001, 316, 5–8. [Google Scholar] [CrossRef] [PubMed]
- Tran, T.T.; Chowanadisai, W.; Crinella, F.M.; Chicz-DeMet, A.; Lönnerdal, B. Effect of high dietary manganese intake of neonatal rats on tissue mineral accumulation, striatal dopamine levels, and neurodevelopmental status. Neurotoxicology 2002, 23, 635–643. [Google Scholar] [CrossRef]
- Seth, P.K.; Chandra, S.V. Neurotransmitters and neurotransmitter receptors in developing and adult rats during manganese poisoning. Neurotoxicology 1984, 5, 67–76. [Google Scholar]
- Amos-Kroohs, R.M.; Davenport, L.L.; Gutierrez, A.; Hufgard, J.R.; Vorhees, C.V.; Williams, M.T. Developmental manganese exposure in combination with developmental stress and iron deficiency: Effects on behavior and monoamines. Neurotoxicol. Teratol. 2016, 56, 55–67. [Google Scholar] [CrossRef] [PubMed]
- Khalid, M.; Aoun, R.A.; Mathews, T.A. Altered striatal dopamine release following a sub-acute exposure to manganese. J. Neurosci. Methods 2011, 202, 182–191. [Google Scholar] [CrossRef]
- Vezér, T.; Kurunczi, A.; Náray, M.; Papp, A.; Nagymajtényi, L. Behavioral effects of subchronic inorganic manganese exposure in rats. Am. J. Ind. Med. 2007, 50, 841–852. [Google Scholar] [CrossRef]
- Chen, M.K.; Lee, J.S.; McGlothan, J.L.; Furukawa, E.; Adams, R.J.; Alexander, M.; Wong, D.F.; Guilarte, T.R. Acute manganese administration alters dopamine transporter levels in the non-human primate striatum. Neurotoxicology 2006, 27, 229–236. [Google Scholar] [CrossRef]
- Johnson, J., Jr.; Pajarillo, E.; Karki, P.; Kim, J.; Son, D.S.; Aschner, M.; Lee, E. Valproic acid attenuates manganese-induced reduction in expression of GLT-1 and GLAST with concomitant changes in murine dopaminergic neurotoxicity. Neurotoxicology 2018, 67, 112–120. [Google Scholar] [CrossRef]
- Pajarillo, E.; Rizor, A.; Son, D.S.; Aschner, M.; Lee, E. The transcription factor REST up-regulates tyrosine hydroxylase and antiapoptotic genes and protects dopaminergic neurons against manganese toxicity. J. Biol. Chem. 2020, 295, 3040–3054. [Google Scholar] [CrossRef] [PubMed]
- Pajarillo, E.; Johnson, J., Jr.; Kim, J.; Karki, P.; Son, D.S.; Aschner, M.; Lee, E. 17beta-estradiol and tamoxifen protect mice from manganese-induced dopaminergic neurotoxicity. Neurotoxicology 2018, 65, 280–288. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Fu, H.; Liu, B.; Liu, X.; Chen, W.; Yu, X. The effect of postnatal manganese exposure on the NMDA receptor signaling pathway in rat hippocampus. J. Biochem. Mol. Toxicol. 2017, 31, e21969. [Google Scholar] [CrossRef]
- Ma, Z.; Liu, K.; Li, X.R.; Wang, C.; Liu, C.; Yan, D.Y.; Deng, Y.; Liu, W.; Xu, B. Alpha-synuclein is involved in manganese-induced spatial memory and synaptic plasticity impairments via TrkB/Akt/Fyn-mediated phosphorylation of NMDA receptors. Cell Death Dis. 2020, 11, 834. [Google Scholar] [CrossRef]
- Karki, P.; Kim, C.; Smith, K.; Son, D.S.; Aschner, M.; Lee, E. Transcriptional Regulation of the Astrocytic Excitatory Amino Acid Transporter 1 (EAAT1) via NF-kappaB and Yin Yang 1 (YY1). J. Biol. Chem. 2015, 290, 23725–23737. [Google Scholar] [CrossRef]
- Karki, P.; Webb, A.; Smith, K.; Johnson, J., Jr.; Lee, K.; Son, D.S.; Aschner, M.; Lee, E. Yin Yang 1 is a repressor of glutamate transporter EAAT2, and it mediates manganese-induced decrease of EAAT2 expression in astrocytes. Mol. Cell Biol. 2014, 34, 1280–1289. [Google Scholar] [CrossRef] [PubMed]
- Danbolt, N.C. Glutamate uptake. Prog. Neurobiol. 2001, 65, 1–105. [Google Scholar] [CrossRef]
- Brouillet, E.P.; Shinobu, L.; McGarvey, U.; Hochberg, F.; Beal, M.F. Manganese injection into the rat striatum produces excitotoxic lesions by impairing energy metabolism. Exp. Neurol. 1993, 120, 89–94. [Google Scholar] [CrossRef]
- Lai, J.C.; Leung, T.K.; Lim, L. Dierences in the neurotoxic effects of manganese during development and aging: Some observations on brain regional neurotransmitter and non-neurotransmitter metabolism in a developmental rat model of chronic manganese encephalopathy. Neurotoxicology 1984, 5, 37–47. [Google Scholar]
- Santos, D.; Milatovic, D.; Andrade, V.; Batoreu, M.C.; Aschner, M.; Marreilha dos Santos, A.P. The inhibitory effect of manganese on acetylcholinesterase activity enhances oxidative stress and neuroinflammation in the rat brain. Toxicology 2012, 292, 90–98. [Google Scholar] [CrossRef]
- Lai, J.C.; Leung, T.K.; Lim, L. Brain regional distribution of glutamic acid decarboxylase, choline acetyltransferase, and acetylcholinesterase in the rat: Effects of chronic manganese chloride administration after two years. J. Neurochem. 1981, 36, 1443–1448. [Google Scholar] [CrossRef] [PubMed]
- Eriksson, H.; Morath, C.; Heilbronn, E. Effects of manganese on the nervous system. Acta Neurol. Scand. Suppl. 1984, 100, 89–93. [Google Scholar] [PubMed]
- Ye, Q.; Kim, J. Loss of hfe function reverses impaired recognition memory caused by olfactory manganese exposure in mice. Toxicol. Res. 2015, 31, 17–23. [Google Scholar] [CrossRef]
- Ali, M.M.; Murthy, R.C.; Saxena, D.K.; Chandra, S.V. Effect of low protein diet on manganese neurotoxicity: II. Brain GABA and seizure susceptibility. Neurobehav. Toxicol. Teratol. 1983, 5, 385–389. [Google Scholar]
- Wong, P.C.; Lai, J.C.; Lim, L.; Davison, A.N. Selective inhibition of L-glutamate and gammaaminobutyrate transport in nerve ending particles by aluminum, manganese, and cadmium chloride. J. Inorg. Biochem. 1981, 14, 253–260. [Google Scholar] [CrossRef] [PubMed]
- Anderson, J.G.; Fordahl, S.C.; Cooney, P.T.; Weaver, T.L.; Colyer, C.L.; Erikson, K.M. Manganese exposure alters extracellular GABA, GABA receptor and transporter protein and mRNA levels in the developing rat brain. Neurotoxicology 2008, 29, 1044–1053. [Google Scholar] [CrossRef]
- Dydak, U.; Jiang, Y.M.; Long, L.L.; Zhu, H.; Chen, J.; Li, W.M.; Edden, R.A.; Hu, S.; Fu, X.; Long, Z.; et al. In vivo measurement of brain GABA concentrations by magnetic resonance spectroscopy in smelters occupationally exposed to manganese. Environ. Health Perspect. 2011, 119, 219–224. [Google Scholar] [CrossRef]
- Uversky, V.N.; Li, J.; Fink, A.L. Metal-triggered structural transformations, aggregation, and fibrillation of human alpha-synuclein. A possible molecular NK between Parkinson’s disease and heavy metal exposure. J. Biol. Chem. 2001, 276, 44284–44296. [Google Scholar] [CrossRef]
- Liu, G.; Aliaga, L.; Cai, H. α-synuclein, LRRK2 and their interplay in Parkinson’s disease. Future Neurol. 2012, 7, 145–153. [Google Scholar] [CrossRef] [PubMed]
- Harischandra, D.S.; Jin, H.; Anantharam, V.; Kanthasamy, A.; Kanthasamy, A.G. alpha-Synuclein protects against manganese neurotoxic insult during the early stages of exposure in a dopaminergic cell model of Parkinson’s disease. Toxicol. Sci. 2015, 143, 454–468. [Google Scholar] [CrossRef]
- Harischandra, D.S.; Ghaisas, S.; Rokad, D.; Zamanian, M.; Jin, H.; Anantharam, V.; Kimber, M.; Kanthasamy, A.; Kanthasamy, A.G. Environmental neurotoxicant manganese regulates exosome-mediated extracellular miRNAs in cell culture model of Parkinson’s disease: Relevance to α-synuclein misfolding in metal neurotoxicity. Neurotoxicology 2018, 64, 267–277. [Google Scholar] [CrossRef] [PubMed]
- Tong, Y.; Yang, H.; Tian, X.; Wang, H.; Zhou, T.; Zhang, S.; Yu, J.; Zhang, T.; Fan, D.; Guo, X.; et al. High manganese, a risk for Alzheimer’s disease: High manganese induces amyloid-β related cognitive impairment. J. Alzheimers Dis. 2014, 42, 865–878. [Google Scholar] [CrossRef] [PubMed]
- Srivastava, R.A.; Jain, J.C. Scavenger receptor class B type I expression and elemental analysis in cerebellum and parietal cortex regions of the Alzheimer’s disease brain. J. Neurol. Sci. 2002, 196, 45–52. [Google Scholar] [CrossRef] [PubMed]
- Moyano, P.; Garcia, J.M.; Garcia, J.; Anadon, M.J.; Naval, M.V.; Frejo, M.T.; Sola, E.; Pelayo, A.; Pino, J.D. Manganese increases Aβ and Tau protein levels through proteasome 20S and heat shock proteins 90 and 70 alteration, leading to SN56 cholinergic cell death following single and repeated treatment. Ecotoxicol. Environ. Saf. 2020, 203, 110975. [Google Scholar] [CrossRef] [PubMed]
- Guilarte, T.R. APLP1, Alzheimer’s-like pathology and neurodegeneration in the frontal cortex of manganese-exposed non-human primates. Neurotoxicology 2010, 31, 572–574. [Google Scholar] [CrossRef] [PubMed]
- Lin, G.; Li, X.; Cheng, X.; Zhao, N.; Zheng, W. Manganese Exposure Aggravates β-Amyloid Pathology by Microglial Activation. Front. Aging Neurosci. 2020, 12, 556008. [Google Scholar] [CrossRef]
- Chen, X.; Xing, L.; Li, X.; Chen, N.; Liu, L.; Wang, J.; Zhou, X.; Liu, S. Manganese Ion-Induced Amyloid Fibrillation Kinetics of Hen Egg White-Lysozyme in Thermal and Acidic Conditions. ACS Omega 2023, 8, 16439–16449. [Google Scholar] [CrossRef]
- Nyarko-Danquah, I.; Pajarillo, E.; Digman, A.; Soliman, K.F.A.; Aschner, M.; Lee, E. Manganese Accumulation in the Brain via Various Transporters and Its Neurotoxicity Mechanisms. Molecules 2020, 25, 5880. [Google Scholar] [CrossRef]
- Aschner, J.L.; Aschner, M. Nutritional aspects of manganese homeostasis. Mol. Asp. Med. 2005, 26, 353–362. [Google Scholar] [CrossRef]
- Vollet, K.; Haynes, E.N.; Dietrich, K.N. Manganese exposure and cognition across the lifespan: Contemporary review and argument for biphasic dose-response health effects. Curr. Environ. Health Rep. 2016, 3, 392–404. [Google Scholar] [CrossRef]
- Bryan, M.R.; Bowman, A.B. Manganese and the Insulin-IGF Signaling Network in Huntington’s Disease and Other Neurodegenerative Disorders. Adv. Neurobiol. 2017, 18, 113–142. [Google Scholar] [CrossRef]
- Pfalzer, A.C.; Bowman, A.B. Relationships Between Essential Manganese Biology and Manganese Toxicity in Neurological Disease. Curr. Environ. Health Rep. 2017, 4, 223–228. [Google Scholar] [CrossRef]
- Aschner, M.; Guilarte, T.R.; Schneider, J.S.; Zheng, W. Manganese: Recent advances in understanding its transport and neurotoxicity. Toxicol. Appl. Pharmacol. 2007, 221, 131–147. [Google Scholar] [CrossRef]
- Just, N.; Cudalbu, C.; Lei, H.; Gruetter, R. Effect of manganese chloride on the neurochemical profile of the rat hypothalamus. J. Cereb. Blood Flow Metab. 2011, 31, 2324–2333. [Google Scholar] [CrossRef]
- Carl, G.F.; Blackwell, L.K.; Barnett, F.C.; Thompson, L.A.; Rissinger, C.J.; Olin, K.L.; Critchfield, J.W.; Keen, C.L.; Gallagher, B.B. Manganese and epilepsy: Brain glutamine synthetase and liver arginase activities in genetically epilepsy prone and chronically seizured rats. Epilepsia 1993, 34, 441–446. [Google Scholar] [CrossRef] [PubMed]
- Deskin, R.; Bursain, S.J.; Edens, F.W. Neurochemical alterations induced by manganese chloride in neonatal rats. Neurotoxicology 1981, 2, 65–73. [Google Scholar] [PubMed]
- Li, L.; Yang, X. The Essential Element Manganese, Oxidative Stress, and Metabolic Diseases: Links and Interactions. Oxid. Med. Cell Longev. 2018, 2018, 7580707. [Google Scholar] [CrossRef]
- Nasr, P.; Ignatova, S.; Lundberg, P.; Kechagias, S.; Ekstedt, M. Low hepatic manganese concentrations in patients with hepatic steatosis–A cohort study of copper, iron and manganese in liver biopsies. J. Trace Elem. Med. Biol. 2021, 67, 126772. [Google Scholar] [CrossRef] [PubMed]
- Woo, S.; Noh, Y.; Koh, S.B.; Lee, S.K.; Il Lee, J.; Kim, H.H.; Kim, S.Y.; Cho, J.; Kim, C. Associations of ambient manganese exposure with brain gray matter thickness and white matter hyperintensities. Hypertens. Res. 2023, 46, 1870–1879. [Google Scholar] [CrossRef] [PubMed]
- Caito, S.; Aschner, M. Neurotoxicity of metals. Handb. Clin. Neurol. 2015, 131, 169–189. [Google Scholar] [CrossRef] [PubMed]
- Martins, A.C., Jr.; Morcillo, P.; Ijomone, O.M.; Venkataramani, V.; Harrison, F.E.; Lee, E.; Bowman, A.B.; Aschner, M. New Insights on the Role of Manganese in Alzheimer’s Disease and Parkinson’s Disease. Int. J. Environ. Res. Public Health 2019, 16, 3546. [Google Scholar] [CrossRef]
- Gonzalez-Cuyar, L.F.; Nelson, G.; Criswell, S.R.; Ho, P.; Lonzanida, J.A.; Checkoway, H.; Seixas, N.; Gelman, B.B.; Evanoff, B.A.; Murray, J.; et al. Quantitative neuropathology associated with chronic manganese exposure in South African mine workers. Neurotoxicology 2014, 45, 260–266. [Google Scholar] [CrossRef] [PubMed]
- Racette, B.A.; Tabbal, S.D.; Jennings, D.; Good, L.; Perlmutter, J.S.; Evanoff, B. Prevalence of parkinsonism and relationship to exposure in a large sample of Alabama welders. Neurology 2005, 64, 230–235. [Google Scholar] [CrossRef] [PubMed]
- Nicolai, M.M.; Pirritano, M.; Gasparoni, G.; Aschner, M.; Simon, M.; Bornhorst, J. Manganese-Induced Toxicity in C. elegans: What Can We Learn from the Transcriptome? Int. J. Mol. Sci. 2022, 23, 10748. [Google Scholar] [CrossRef]
- Tinkov, A.A.; Paoliello, M.M.B.; Mazilina, A.N.; Skalny, A.V.; Martins, A.C.; Voskresenskaya, O.N.; Aaseth, J.; Santamaria, A.; Notova, S.V.; Tsatsakis, A.; et al. Molecular Targets of Manganese-Induced Neurotoxicity: A Five-Year Update. Int. J. Mol. Sci. 2021, 22, 4646. [Google Scholar] [CrossRef]
- Fitsanakis, V.A.; Au, C.; Erikson, K.M.; Aschner, M. The effects of manganese on glutamate, dopamine and γ-aminobutyric acid regulation. Neurochem. Int. 2006, 48, 426–433. [Google Scholar] [CrossRef]
- Kwik-Uribe, C.; Smith, D.R. Temporal responses in the disruption of iron regulation by manganese. J. Neurosci. Res. 2006, 83, 1601–1610. [Google Scholar] [CrossRef]
- Tamm, C.; Sabri, F.; Ceccatelli, S. Mitochondrial-mediated apoptosis in neural stem cells exposed to manganese. Toxicol. Sci. 2008, 101, 310–320. [Google Scholar] [CrossRef]
- Tjalkens, R.B.; Popichak, K.A.; Kirkley, K.A. Inflammatory activation of microglia and astrocytes in manganese neurotoxicity. Adv. Neurobiol. 2017, 18, 159–181. [Google Scholar] [CrossRef] [PubMed]
- Filipov, N.M.; Dodd, C.A. Role of glial cells in manganese neurotoxicity. J. Appl. Toxicol. 2012, 32, 310–317. [Google Scholar] [CrossRef]
- Brown, D.R. Brain proteins that mind metals: A neurodegenerative perspective. Dalton Trans. 2009, 21, 4069–4076. [Google Scholar] [CrossRef] [PubMed]
- Williams, B.B.; Kwakye, G.F.; Wegrzynowicz, M.; Li, D.; Aschner, M.; Erikson, K.M.; Bowman, A.B. Altered manganese homeostasis and manganese toxicity in a Huntington’s disease striatal cell model are not explained by defects in the iron transport system. Toxicol. Sci. 2010, 117, 169–179. [Google Scholar] [CrossRef]
- Laohaudomchok, W.; Lin, X.; Herrick, R.F.; Fang, S.C.; Cavallari, J.M.; Shrairman, R.; Landau, A.; Christiani, D.C.; Weisskopf, M.G. Neuropsychological effects of low-level manganese exposure in welders. Neurotoxicology 2011, 32, 171–179. [Google Scholar] [CrossRef] [PubMed]
- Oulhote, Y.; Mergler, D.; Barbeau, B.; Bellinger, D.C.; Bouffard, T.; Brodeur, M.E.; Saint-Amour, D.; Legrand, M.; Sauvé, S.; Bouchard, M.F. Neurobehavioral function in school-age children exposed to manganese in drinking water. Environ. Health Perspect. 2014, 122, 1343–1350. [Google Scholar] [CrossRef]
- Rugless, F.; Bhattacharya, A.; Succop, P.; Dietrich, K.N.; Cox, C.; Alden, J.; Kuhnell, P.; Barnas, M.; Wright, R.; Parsons, P.J.; et al. Childhood exposure to manganese and postural instability in children living near a ferromanganese refinery in Southeastern Ohio. Neurotoxicol. Teratol. 2014, 41, 71–79. [Google Scholar] [CrossRef] [PubMed]
- Coetzee, D.J.; McGovern, P.M.; Rao, R.; Harnack, L.J.; Georgieff, M.K.; Stepanov, I. Measuring the impact of manganese exposure on children’s neurodevelopment: Advances and research gaps in biomarker-based approaches. Environ. Health 2016, 15, 91. [Google Scholar] [CrossRef] [PubMed]
- Haynes, E.N.; Ryan, P.; Chen, A.; Brown, D.; Roda, S.; Kuhnell, P.; Wittberg, D.; Terrell, M.; Reponen, T. Assessment of personal exposure to manganese in children living near a ferromanganese refinery. Sci. Total Environ. 2012, 427–428, 19–25. [Google Scholar] [CrossRef]
- Yu, X.D.; Zhang, J.; Yan, C.H.; Shen, X.M. Prenatal exposure to manganese at environment relevant level and neonatal neurobehavioral development. Environ. Res. 2014, 133, 232–238. [Google Scholar] [CrossRef]
- Henn, B.C.; Ettinger, A.S.; Schwartz, J.; Téllez-Rojo, M.M.; Lamadrid-Figueroa, H.; Hernández-Avila, M.; Schnaas, L.; Amarasiriwardena, C.; Bellinger, D.C.; Hu, H.; et al. Early postnatal blood manganese levels and children’s neurodevelopment. Epidemiology 2010, 21, 433–439. [Google Scholar] [CrossRef] [PubMed]
- Lin, C.C.; Chen, Y.C.; Su, F.C.; Lin, C.M.; Liao, H.F.; Hwang, Y.H.; Hsieh, W.S.; Jeng, S.F.; Su, Y.N.; Chen, P.C. In utero exposure to environmental lead and manganese and neurodevel-opment at 2 years of age. Environ. Res. 2013, 123, 52–57. [Google Scholar] [CrossRef] [PubMed]
- Bhang, S.Y.; Cho, S.C.; Kim, J.W.; Hong, Y.C.; Shin, M.S.; Yoo, H.J.; Cho, I.H.; Kim, Y.; Kim, B.N. Relationship between blood manganese levels and children’s attention, cognition, behavior, and academic performance--a nationwide cross-sectional study. Environ. Res. 2013, 126, 9–16. [Google Scholar] [CrossRef] [PubMed]
- Rink, S.M.; Ardoino, G.; Queirolo, E.I.; Cicariello, D.; Mañay, N.; Kordas, K. Associations between hair manganese levels and cognitive, language, and motor development in preschool children from Montevideo, Uruguay. Arch. Environ. Occup. Health 2014, 69, 46–54. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues, E.G.; Bellinger, D.C.; Valeri, L.; Hasan, M.O.; Quamruzzaman, Q.; Golam, M.; Kile, M.L.; Christiani, D.C.; Wright, R.O.; Mazumdar, M. Neurodevelopmental outcomes among 2- to 3-year-old children in Bangladesh with elevated blood lead and exposure to arsenic and manganese in drinking water. Environ. Health 2016, 15, 44. [Google Scholar] [CrossRef]
- Lucchini, R.G.; Zoni, S.; Guazzetti, S.; Bontempi, E.; Micheletti, S.; Broberg, K.; Parrinello, G.; Smith, D.R. Inverse association of intellectual function with very low blood lead but not with manganese exposure in Italian adolescents. Environ. Res. 2012, 118, 65–71. [Google Scholar] [CrossRef]
- Rahman, S.M.; Kippler, M.; Tofail, F.; Bölte, S.; Hamadani, J.D.; Vahter, M. Manganese in Drinking Water and Cognitive Abilities and Behavior at 10 Years of Age: A Prospective Cohort Study. Environ. Health Perspect. 2017, 125, 057003. [Google Scholar] [CrossRef]
- Nascimento, S.; Baierle, M.; Göethel, G.; Barth, A.; Brucker, N.; Charão, M.; Sauer, E.; Gauer, B.; Arbo, M.D.; Altknecht, L.; et al. Associations among environmental exposure to manganese, neuropsychological performance, oxidative damage and kidney biomarkers in children. Environ. Res. 2016, 147, 32–43. [Google Scholar] [CrossRef]
- Khan, K.; Wasserman, G.A.; Liu, X.; Ahmed, E.; Parvez, F.; Slavkovic, V.; Levy, D.; Mey, J.; van Geen, A.; Graziano, J.H.; et al. Manganese exposure from drinking water and children’s academic achievement. Neurotoxicology 2012, 33, 91–97. [Google Scholar] [CrossRef]
- Hernández-Bonilla, D.; Schilmann, A.; Montes, S.; Rodríguez-Agudelo, Y.; Rodríguez-Dozal, S.; Solís-Vivanco, R.; Ríos, C.; Riojas-Rodríguez, H. Environmental exposure to manganese and motor function of children in Mexico. Neurotoxicology 2011, 32, 615–621. [Google Scholar] [CrossRef]
- Sanders, A.P.; Claus Henn, B.; Wright, R.O. Perinatal and Childhood Exposure to Cadmium, Manganese, and Metal Mixtures and Effects on Cognition and Behavior: A Review of Recent Literature. Curr. Environ. Health Rep. 2015, 2, 284–294. [Google Scholar] [CrossRef]
- Yousef, S.; Adem, A.; Zoubeidi, T.; Kosanovic, M.; Mabrouk, A.A.; Eapen, V. Attention deficit hyperactivity disorder and environmental toxic metal exposure in the United Arab Emirates. J. Trop. Pediatr. 2011, 57, 457–460. [Google Scholar] [CrossRef] [PubMed]
- Abdullah, M.M.; Ly, A.R.; Goldberg, W.A.; Clarke-Stewart, K.A.; Dudgeon, J.V.; Mull, C.G.; Chan, T.J.; Kent, E.E.; Mason, A.Z.; Ericson, J.E. Heavy metal in children’s tooth enamel: Related to autism and disruptive behaviors? J. Autism Dev. Disord. 2012, 42, 929–936. [Google Scholar] [CrossRef] [PubMed]
- Khan, K.; Factor-Litvak, P.; Wasserman, G.A.; Liu, X.; Ahmed, E.; Parvez, F.; Slavkovich, V.; Levy, D.; Mey, J.; van Geen, A.; et al. Manganese exposure from drinking water and children’s classroom behavior in Bangladesh. Environ. Health Perspect. 2011, 119, 1501–1506. [Google Scholar] [CrossRef]
- Dring, J.C.; Forma, A.; Chilimoniuk, Z.; Dobosz, M.; Teresinski, G.; Buszewicz, G.; Flieger, J.; Cywka, T.; Januszewski, J.; Baj, J. Essentiality of Trace Elements in Pregnancy, Fertility, and Gynecologic Cancers-A State-of-the-Art Review. Nutrients 2022, 14, 185. [Google Scholar] [CrossRef] [PubMed]
- Skogheim, T.S.; Weyde, K.V.F.; Engel, S.M.; Aase, H.; Suren, P.; Oie, M.G.; Biele, G.; Reichborn-Kjennerud, T.; Caspersen, I.H.; Hornig, M.; et al. Metal and essential element concentrations during pregnancy and associations with autism spectrum disorder and attention-deficit/hyperactivity disorder in children. Environ. Int. 2021, 152, 106468. [Google Scholar] [CrossRef]
- Bjørklund, G.; Meguid, N.A.; El-Bana, M.A.; Tinkov, A.A.; Saad, K.; Dadar, M.; Hemimi, M.; Skalny, A.V.; Hosnedlová, B.; Kizek, R.; et al. Oxidative Stress in Autism Spectrum Disorder. Mol. Neurobiol. 2020, 57, 2314–2332. [Google Scholar] [CrossRef]
- Manivasagam, T.; Arunadevi, S.; Essa, M.M.; SaravanaBabu, C.; Borah, A.; Thenmozhi, A.J.; Qoronfleh, M.W. Role of Oxidative Stress and Antioxidants in Autism. Adv. Neurobiol. 2020, 24, 193–206. [Google Scholar] [CrossRef]
- Pangrazzi, L.; Balasco, L.; Bozzi, Y. Oxidative Stress and Immune System Dysfunction in Autism Spectrum Disorders. Int. J. Mol. Sci. 2020, 21, 3293. [Google Scholar] [CrossRef]
- Ye, J.; Wang, H.; Cui, L.; Chu, S.; Chen, N. The progress of chemokines and chemokine receptors in autism spectrum disorders. Brain Res. Bull. 2021, 174, 268–280. [Google Scholar] [CrossRef]
- Robinson-Agramonte, M.L.A.; Noris García, E.; Fraga Guerra, J.; Vega Hurtado, Y.; Antonucci, N.; Semprún-Hernández, N.; Schultz, S.; Siniscalco, D. Immune Dysregulation in Autism Spectrum Disorder: What Do We Know about It? Int. J. Mol. Sci. 2022, 23, 3033. [Google Scholar] [CrossRef] [PubMed]
- Bjørklund, G.; Doşa, M.D.; Maes, M.; Dadar, M.; Frye, R.E.; Peana, M.; Chirumbolo, S. The impact of glutathione metabolism in autism spectrum disorder. Pharmacol. Res. 2021, 166, 105437. [Google Scholar] [CrossRef] [PubMed]
- Bjørklund, G.; Tinkov, A.A.; Hosnedlová, B.; Kizek, R.; Ajsuvakova, O.P.; Chirumbolo, S.; Skalnaya, M.G.; Peana, M.; Dadar, M.; El-Ansary, A.; et al. The role of glutathione redox imbalance in autism spectrum disorder: A review. Free Radic. Biol. Med. 2020, 160, 149–162. [Google Scholar] [CrossRef]
- Yui, K.; Imataka, G.; Sasaki, H.; Shiroki, R. The role of lipid peroxidation in individuals with autism spectrum disorders. Metab. Brain Dis. 2020, 35, 1101–1108. [Google Scholar] [CrossRef] [PubMed]
- Yui, K.; Imataka, G.; Sasaki, H.; Shiroki, R.; Koshiba, M. Lipid Peroxidation With Implication of Organic Pollution in Autistic Behaviors. Cureus 2021, 13, e14188. [Google Scholar] [CrossRef]
- Bijl, N.; Thys, C.; Wittevrongel, C.; De la Marche, W.; Devriendt, K.; Peeters, H.; Van Geet, C.; Freson, K. Platelet studies in autism spectrum disorder patients and first-degree relatives. Mol. Autism 2015, 6, 57. [Google Scholar] [CrossRef]
- Jagadapillai, R.; Qiu, X.; Ojha, K.; Li, Z.; El-Baz, A.; Zou, S.; Gozal, E.; Barnes, G.N. Potential Cross Talk between Autism Risk Genes and Neurovascular Molecules: A Pilot Study on Impact of Blood Brain Barrier Integrity. Cells 2022, 11, 2211. [Google Scholar] [CrossRef]
- Mostafa, G.A.; Bjørklund, G.; Urbina, M.A.; Al-Ayadhi, L.Y. The positive association between elevated blood lead levels and brain-specific autoantibodies in autistic children from low lead-polluted areas. Metab. Brain Dis. 2016, 31, 1047–1054. [Google Scholar] [CrossRef]
- Ouisselsat, M.; Maidoumi, S.; Elmaouaki, A.; Lekouch, N.; Pineau, A.; Sedki, A. Hair Trace Elements and Mineral Content in Moroccan Children with Autism Spectrum Disorder: A Case-Control Study. Biol. Trace Elem. Res. 2023, 201, 2701–2710. [Google Scholar] [CrossRef]
- Baj, J.; Flieger, W.; Flieger, M.; Forma, A.; Sitarz, E.; Skórzyńska-Dziduszko, K.; Grochowski, C.; Maciejewski, R.; Karakuła-Juchnowicz, H. Autism spectrum disorder: Trace elements imbalances and the pathogenesis and severity of autistic symptoms. Neurosci. Biobehav. Rev. 2021, 129, 117–132. [Google Scholar] [CrossRef]
- Ding, M.; Shi, S.; Qie, S.; Li, J.; Xi, X. Association between heavy metals exposure (cadmium, lead, arsenic, mercury) and child autistic disorder: A systematic review and meta-analysis. Front. Pediatr. 2023, 11, 1169733. [Google Scholar] [CrossRef]
- Rahbar, M.H.; Samms-Vaughan, M.; Ardjomand-Hessabi, M.; Loveland, K.A.; Dickerson, A.S.; Chen, Z.; Bressler, J.; Shakespeare-Pellington, S.; Grove, M.L.; Bloom, K.; et al. The role of drinking water sources, consumption of vegetables and seafood in relation to blood arsenic concentrations of Jamaican children with and without Autism Spectrum Disorders. Sci. Total Environ. 2012, 433, 362–370. [Google Scholar] [CrossRef] [PubMed]
- Rahbar, M.H.; Samms-Vaughan, M.; Lee, M.; Zhang, J.; Hessabi, M.; Bressler, J.; Bach, M.A.; Grove, M.L.; Shakespeare-Pellington, S.; Beecher, C.; et al. Interaction between a Mixture of Heavy Metals (Lead, Mercury, Arsenic, Cadmium, Manganese, Aluminum) and GSTP1, GSTT1, and GSTM1 in Relation to Autism Spectrum Disorder. Res. Autism Spectr. Disord. 2020, 79, 101681. [Google Scholar] [CrossRef] [PubMed]
- Rahbar, M.H.; Samms-Vaughan, M.; Ma, J.; Bressler, J.; Dickerson, A.S.; Hessabi, M.; Loveland, K.A.; Grove, M.L.; Shakespeare-Pellington, S.; Beecher, C.; et al. Synergic effect of GSTP1 and blood manganese concentrations in Autism Spectrum disorder. Res. Autism Spectr. Disord. 2015, 18, 73–82. [Google Scholar] [CrossRef]
- Rahbar, M.H.; Samms-Vaughan, M.; Dickerson, A.S.; Loveland, K.A.; Ardjomand-Hessabi, M.; Bressler, J.; Lee, M.; Shakespeare-Pellington, S.; Grove, M.L.; Pearson, D.A.; et al. Role of fruits, grains, and seafood consumption in blood cadmium concentrations of Jamaican children with and without Autism Spectrum disorder. Res. Autism Spectr. Disord. 2014, 8, 1134–1145. [Google Scholar] [CrossRef]
- Yoon, M.; Schroeter, J.D.; Nong, A.; Taylor, M.D.; Dorman, D.C.; Andersen, M.E.; Clewell, H.J. Part III. Physiologically based pharmacokinetic modeling of fetal and neonatal manganese exposure in humans: Describing manganese homeostasis during development. Toxicol. Sci. 2011, 122, 297–316. [Google Scholar] [CrossRef]
- Rodríguez-Barranco, M.; Lacasaña, M.; Aguilar-Garduño, C.; Alguacil, J.; Gil, F.; González-Alzaga, B. Association of arsenic, cadmium and manganese exposure with neurodevelopment and behavioural disorders in children: A systematic review and meta-analysis. Sci. Total Environ. 2013, 454–455, 562–577. [Google Scholar] [CrossRef]
- Eastman, R.R.; Jursa, T.P.; Benedetti, C.; Lucchini, R.G.; Smith, D.R. Hair as a biomarker of environmental manganese exposure. Environ. Sci. Technol. 2013, 47, 1629–1637. [Google Scholar] [CrossRef]
- Chung, S.E.; Cheong, H.K.; Ha, E.H.; Kim, B.N.; Ha, M.; Kim, Y.; Hong, Y.C.; Park, H.; Oh, S.Y. Maternal Blood Manganese and Early Neurodevelopment: The Mothers and Children’s Environmental Health (MOCEH) Study. Environ. Health Perspect. 2015, 123, 717–722. [Google Scholar] [CrossRef]
- Takser, L.; Mergler, D.; Hellier, G.; Sahuquillo, J.; Huel, G. Manganese, monoamine metabolite levels at birth, and child psychomotor development. Neurotoxicology 2003, 24, 667–674. [Google Scholar] [CrossRef] [PubMed]
- Erikson, K.M.; Thompson, K.; Aschner, J.; Aschner, M. Manganese neurotoxicity: A focus on the neonate. Pharmacol. Ther. 2007, 113, 369–377. [Google Scholar] [CrossRef] [PubMed]
- Claus Henn, B.; Ettinger, A.S.; Schwartz, J.; Téllez-Rojo, M.M.; Lamadrid-Figueroa, H.; Hernández-Avila, M.; Schnaas, L.; Amarasiriwardena, C.; Bellinger, D.C.; Hu, H. Association of prenatal and childhood blood lead concentrations with criminal arrests in early adulthood. PLoS Med. 2014, 11, e1001651. [Google Scholar] [CrossRef]
- Kalkbrenner, A.E.; Schmidt, R.J.; Penlesky, A.C.; Bennett, D.H. Environmental chemical exposures and autism spectrum disorder: A review of the epidemiological evidence. Curr. Probl. Pediatr. Adolesc. Health Care 2019, 49, 89–124. [Google Scholar] [CrossRef] [PubMed]
- Menezes-Filho, J.A.; Novaes, C.D.; Moreira, J.C.; Sarcinelli, P.N.; Mergler, D. Elevated manganese and cognitive performance in school-aged children and their mothers. Environ. Res. 2011, 111, 156–163. [Google Scholar] [CrossRef]
- Takser, L.; Mergler, D.; Baldwin, M.; de Grosbois, S.B.; Smargiassi, A.; Lafond, J.; Nakhaei, M. Manganese and child neurodevelopment. NeuroToxicology 2009, 30, 515–521. [Google Scholar] [CrossRef]
- Kim, Y.; Kim, B.-N.; Hong, Y.-C.; Shin, M.-S.; Yoo, H.-J.; Kim, J.-W.; Bhang, S.-Y.; Cho, S.-C. Co-exposure to environmental lead and manganese affects the intelligence of school-aged children. Neurotoxicology 2009, 30, 564–571. [Google Scholar] [CrossRef]
- Riojas-Rodriguez, H.; Solis-Vivanco, R.; Schilmann, A.; Montes, S.; Rodriguez, S.; Rios, C.; Rodríguez-Agudelo, Y. Intellectual Function in Mexican Children Living in a Mining Area and Environmentally Exposed to Manganese. Environ. Health Perspect. 2010, 118, 1465–1470. [Google Scholar] [CrossRef]
- Bouchard, M.F.; Sauve, S.; Barbeau, B.; Legrand, M.; Brodeur, M.E.; Bouffard, T.; Limoges, E.; Bellinger, D.C.; Mergler, D. Intellectual Impairment in School-Age Children Exposed to Manganese from Drinking Water. Environ. Health Perspect. 2011, 119, 138–143. [Google Scholar] [CrossRef]
- Wasserman, G.A.; Liu, X.; Parvez, F.; Factor-Litvak, P.; Ahsan, H.; Levy, D.; Kline, J.; van Geen, A.; Mey, J.; Slavkovich, V.; et al. Arsenic and manganese exposure and children’s intellectual function. Neurotoxicology 2011, 32, 450–457. [Google Scholar] [CrossRef]
- Gunier, R.B.; Mora, A.M.; Smith, D.; Arora, M.; Austin, C.; Eskenazi, B.; Bradman, A. Biomarkers of manganese exposure in pregnant women and children living in an agricultural community in California. Environ. Sci. Technol. 2014, 48, 14695–14702. [Google Scholar] [CrossRef] [PubMed]
- Qin, Y.Y.; Jian, B.; Wu, C.; Jiang, C.Z.; Kang, Y.; Zhou, J.X.; Yang, F.; Liang, Y. A comparison of blood metal levels in autism spectrum disorder and unaffected children in Shenzhen of China and factors involved in bioaccumulation of metals. Environ. Sci. Pollut. Res. Int. 2018, 25, 17950–17956. [Google Scholar] [CrossRef] [PubMed]
- Skalny, A.V.; Simashkova, N.V.; Klyushnik, T.P.; Grabeklis, A.R.; Radysh, I.V.; Skalnaya, M.G.; Nikonorov, A.A.; Tinkov, A.A. Assessment of serum trace elements and electrolytes in children with childhood and atypical autism. J. Trace Elem. Med. Biol. 2017, 43, 9–14. [Google Scholar] [CrossRef]
- Balachandran, R.C.; Mukhopadhyay, S.; McBride, D.; Veevers, J.; Harrison, F.E.; Aschner, M.; Haynes, E.N.; Bowman, A.B. Brain manganese and the balance between essential roles and neurotoxicity. J. Biol. Chem. 2020, 295, 6312–6329. [Google Scholar] [CrossRef] [PubMed]
- Tuschl, K.; Mills, P.B.; Clayton, P.T. Manganese and the brain. Int. Rev. Neurobiol. 2013, 110, 277–312. [Google Scholar] [CrossRef]
- Peres, T.V.; Schettinger, M.R.; Chen, P.; Carvalho, F.; Avila, D.S.; Bowman, A.B.; Aschner, M. Manganese-induced neurotoxicity: A review of its behavioral consequences and neuroprotective strategies. BMC Pharmacol. Toxicol. 2016, 17, 57. [Google Scholar] [CrossRef]
- Takeda, A. Essential trace metals and brain function. Yakugaku Zasshi 2004, 124, 577–585. [Google Scholar] [CrossRef]
- Lucchini, R.; Placidi, D.; Cagna, G.; Fedrighi, C.; Oppini, M.; Peli, M.; Zoni, S. Manganese and Developmental Neurotoxicity. Adv. Neurobiol. 2017, 18, 13–34. [Google Scholar] [CrossRef]
- Tarnacka, B.; Jopowicz, A.; Maślińska, M. Copper, Iron, and Manganese Toxicity in Neuropsychiatric Conditions. Int. J. Mol. Sci. 2021, 22, 7820. [Google Scholar] [CrossRef] [PubMed]
- Hong, S.B.; Kim, J.W.; Choi, B.S.; Hong, Y.C.; Park, E.J.; Shin, M.S.; Kim, B.N.; Yoo, H.J.; Cho, I.H.; Bhang, S.Y.; et al. Blood manganese levels in relation to comorbid behavioral and emotional problems in children with attentiondeficit/hyperactivity disorder. Psychiatry Res. 2014, 220, 418–425. [Google Scholar] [CrossRef]
- Farias, A.C. Manganese in children with attention-deficit/hyperactivity disorder: Relationship with methylphenidate exposure. J. Child Adolesc. Psychopharmacol. 2010, 20, 113–118. [Google Scholar] [CrossRef]
- Beaudin, S.A.; Strupp, B.J.; Lasley, S.M.; Fornal, C.A.; Mandal, S.; Smith, D.R. Oral methylphenidate alleviates the fine motor dysfunction caused by chronic postnatal manganese exposure in adult rats. Toxicol. Sci. 2015, 144, 318–327. [Google Scholar] [CrossRef] [PubMed]
- Lee, M.-J.; Chou, M.-C.; Chou, W.-J.; Huang, C.-W.; Kuo, H.-C.; Lee, S.-Y.; Wang, L.-J. Heavy Metals’ Effect on Susceptibility to Attention-Deficit/Hyperactivity Disorder: Implication of Lead, Cadmium, and Antimony. Int. J. Environ. Res. Public Health 2018, 15, 1221. [Google Scholar] [CrossRef] [PubMed]
- Calne, D.B.; Chu, N.S.; Huang, C.C.; Lu, C.S.; Olanow, W. Manganism and idiopathic parkinsonism: Similarities and differences. Neurology 1994, 44, 1583–1586. [Google Scholar] [CrossRef]
- Cordova, F.M.; Aguiar, A.S., Jr.; Peres, T.V.; Lopes, M.W.; Gonçalves, F.M.; Pedro, D.Z.; Lopes, S.C.; Pilati, C.; Prediger, R.D.; Farina, M.; et al. Manganese-exposed developing rats display motor deficits and striatal oxidative stress that are reversed by Trolox. Arch. Toxicol. 2013, 87, 1231–1244. [Google Scholar] [CrossRef]
- Di, J.W.; Bi, S.P. UV-Vis spectroscopic study of aluminum-manganese cooperative effect on the melanin formation from DOPA oxidation. Guang Pu Xue Yu Guang Pu Fen Xi. 2005, 25, 83–85. [Google Scholar]
- Slominski, A.; Tobin, D.J.; Shibahara, S.; Wortsman, J. Melanin pigmentation in mammalian skin and its hormonal regulation. Physiol. Rev. 2004, 84, 1155–1228. [Google Scholar] [CrossRef] [PubMed]
- Mergler, D.; Baldwin, M. Early manifestations of manganese neurotoxicity in humans: An update. Environ. Res. 1997, 73, 92–100. [Google Scholar] [CrossRef] [PubMed]
- Parmalee, N.L.; Aschner, M. Manganese and aging. NeuroToxicology 2016, 56, 262–268. [Google Scholar] [CrossRef]
- WHO. Recommended Health-Based Limits in Occupational Exposure to Heavy Metals, Report of a WHO Study Group; Technical Report Series; WHO: Geneva, Switzerland, 1980; Volume 647. [Google Scholar]
- Guilarte, T.R.; Gonzales, K.K. Manganese-Induced Parkinsonism Is Not Idiopathic Parkinson’s Disease: Environmental and Genetic Evidence. Toxicol. Sci. 2015, 146, 204–212. [Google Scholar] [CrossRef]
- Kim, Y.; Kim, J.; Kim, H.; Kim, J.W.; Chung, K.H.; Park, E. Manganese-induced parkinsonism and Parkinson’s disease: Shared and distinguishable features. Int. J. Environ. Res. Public Health 2019, 16, 341. [Google Scholar] [CrossRef]
- Racette, B.A.; Criswell, S.R.; Lundin, J.I.; Hobson, A.; Seixas, N.; Kotzbauer, P.T.; Evanoff, B.A.; Perlmutter, J.S.; Zhang, J.; Sheppard, L. Increased risk of parkinsonism associated with welding exposure. Neurotoxicology 2012, 33, 1356–1361. [Google Scholar] [CrossRef] [PubMed]
- Benedetto, A.; Au, C.; Aschner, M. Manganese-induced dopaminergic neurodegeneration: Insights into mechanisms and genetics shared with Parkinson’s disease. Chem. Rev. 2009, 109, 4862–4884. [Google Scholar] [CrossRef]
- Lucchini, R.; Albini, E.; Benedetti, L.; Borghesi, S.; Coccaglio, R.; Malara, E.C.; Parrinello, G.; Garattini, S.; Resola, S.; Alessio, L. High prevalence of Parkinsonian disorders associated to manganese exposure in the vicinities of ferroalloy industries. Am. J. Ind. Med. 2007, 50, 788–800. [Google Scholar] [CrossRef]
- Olanow, C.W.; Good, P.F.; Shinotoh, H.; Hewitt, K.A.; Vingerhoets, F.J.; Snow, B.J. Manganese intoxication in the rhesus monkey: A clinical, imaging, pathologic, and biochemical study. Neurology 1996, 46, 492–498. [Google Scholar] [CrossRef] [PubMed]
- van der Mark, M.; Vermeulen, R.; Nijssen, P.C.G.; Mulleners, W.M.; Sas, A.M.G.; van Laar, T.; Anke, H.; Kromhout, H. Occupational exposure to solvents and metals and risk of Parkinson’s disease. Park. Relat. Disord. 2021, 87, 9–16. [Google Scholar] [CrossRef]
- Kwakye, G.F.; Paoliello, M.M.; Mukhopadhyay, S.; Bowman, A.B.; Aschner, M. Manganese-Induced Parkinsonism and Parkinson’s Disease: Shared and Distinguishable Features. Int. J. Environ. Res. Public Health 2015, 12, 7519–7540. [Google Scholar] [CrossRef]
- Cheng, H.; Yang, B.; Ke, T.; Li, S.; Yang, X.; Aschner, M.; Chen, P. Mechanisms of Metal-Induced Mitochondrial Dysfunction in Neurological Disorders. Toxics 2021, 9, 142. [Google Scholar] [CrossRef]
- Hisahara, S.; Shimohama, S. Dopamine receptors and Parkinson’s disease. Int. J. Med. Chem. 2011, 2011, 403039. [Google Scholar] [CrossRef]
- Dharmadhikari, S.; Ma, R.; Yeh, C.L.; Stock, A.K.; Snyder, S.; Zauber, S.E.; Dydak, U.; Beste, C. Striatal and thalamic GABA level concentrations play differential roles for the modulation of response selection processes by proprioceptive information. Neuroimage 2015, 120, 36–42. [Google Scholar] [CrossRef]
- Emir, U.E.; Tuite, P.J.; Oz, G. Elevated pontine and putamenal GABA levels in mild-moderate Parkinson disease detected by 7 tesla proton MRS. PLoS ONE 2012, 7, e30918. [Google Scholar] [CrossRef]
- Casjens, S.; Dydak, U.; Dharmadhikari, S.; Lotz, A.; Lehnert, M.; Quetscher, C.; Stewig, C.; Glaubitz, B.; Schmidt-Wilcke, T.; Edmondson, D.; et al. Association of exposure to manganese and iron with striatal and thalamic GABA and other neurometabolites—Neuroimaging results from the WELDOX II study. Neurotoxicology 2018, 64, 60–66. [Google Scholar] [CrossRef] [PubMed]
- Sanchez-Betancourt, J.; Anaya-Martinez, V.; Gutierrez-Valdez, A.L.; Ordonez-Librado, J.L.; Montiel-Flores, E.; Espinosa-Villanueva, J.; Reynoso-Erazo, L.; Avila-Costa, M.R. Manganese mixture inhalation is a reliable Parkinson disease model in rats. Neurotoxicology 2012, 33, 1346–1355. [Google Scholar] [CrossRef] [PubMed]
- Wolff, N.A.; Garrick, M.D.; Zhao, L.; Garrick, L.M.; Ghio, A.J.; Thevenod, F. A role for divalent metal transporter (DMT1) in mitochondrial uptake of iron and manganese. Sci. Rep. 2018, 8, 211. [Google Scholar] [CrossRef]
- Guilarte, T.R.; Yeh, C.L.; McGlothan, J.L.; Perez, J.; Finley, P.; Zhou, Y.; Wong, D.F.; Dydak, U.; Schneider, J.S. PET imaging of dopamine release in the frontal cortex of manganese-exposed non-human primates. J. Neurochem. 2019, 150, 188–201. [Google Scholar] [CrossRef] [PubMed]
- Criswell, S.R.; Warden, M.N.; Searles Nielsen, S.; Perlmutter, J.S.; Moerlein, S.M.; Sheppard, L.; Lenox-Krug, J.; Checkoway, H.; Racette, B.A. Selective D2 receptor PET in manganese-exposed workers. Neurology 2018, 91, e1022–e1030. [Google Scholar] [CrossRef]
- Cai, T.; Yao, T.; Zheng, G.; Chen, Y.; Du, K.; Cao, Y.; Shen, X.; Chen, J.; Luo, W. Manganese induces the overexpression of alpha-synuclein in PC12 cells via ERK activation. Brain Res. 2010, 1359, 201–207. [Google Scholar] [CrossRef]
- Li, Y.; Jiao, Q.; Xu, H.; Du, X.; Shi, L.; Jia, F.; Jiang, H. Biometal Dyshomeostasis and Toxic Metal Accumulations in the Development of Alzheimer’s Disease. Front. Mol. Neurosci. 2017, 10, 339. [Google Scholar] [CrossRef]
- Szabo, S.T.; Harry, G.J.; Hayden, K.M.; Szabo, D.T.; Birnbaum, L. Comparison of Metal Levels between Postmortem Brain and Ventricular Fluid in Alzheimer’s Disease and Nondemented Elderly Controls. Toxicol Sci. 2016, 150, 292–300. [Google Scholar] [CrossRef]
- Bonda, D.J.; Lee, H.G.; Blair, J.A.; Zhu, X.; Perry, G.; Smith, M.A. Role of metal dyshomeostasis in Alzheimer’s disease. Metallomics 2011, 3, 267–270. [Google Scholar] [CrossRef]
- Ward, R.J.; Zucca, F.A.; Duyn, J.H.; Crichton, R.R.; Zecca, L. The role of iron in brain ageing and neurodegenerative disorders. Lancet Neurol. 2014, 13, 1045–1060. [Google Scholar] [CrossRef]
- DeWitt, M.R.; Chen, P.; Aschner, M. Manganese efflux in Parkinsonism: Insights from newly characterized SLC30A10 mutation. Biochem. Biophys. Res. Commun. 2013, 432, 1–4. [Google Scholar] [CrossRef]
- Avila, D.S.; Puntel, R.L.; Aschner, M. Manganese in health and disease. Met. Ions Life Sci. 2013, 13, 199–227. [Google Scholar] [CrossRef]
- Gunter, T.E.; Gerstner, B.; Lester, T.; Wojtovich, A.P.; Malecki, J.; Swarts, S.G.; Brookes, P.S.; Gavin, C.E.; Gunter, K.K. An analysis of the effects of Mn2+ on oxidative phosphorylation in liver, brain, and heart mitochondria using state 3 oxidation rate assays. Toxicol. Appl. Pharmacol. 2010, 249, 65–75. [Google Scholar] [CrossRef]
- Ammal Kaidery, N.; Thomas, B. Current perspective of mitochondrial biology in Parkinson’s disease. Neurochem. Int. 2018, 117, 91–113. [Google Scholar] [CrossRef]
- Helley, M.P.; Pinnell, J.; Sportelli, C.; Tieu, K. Mitochondria: A Common Target for Genetic Mutations and Environmental Toxicants in Parkinson’s Disease. Front. Genet. 2017, 8, 177. [Google Scholar] [CrossRef]
- Ijomone, O.M.; Aluko, O.M.; Okoh, C.O.A.; Martins, A.C., Jr.; Aschner, M. Role for calcium signaling in manganese neurotoxicity. J. Trace Elem. Med. Biol. 2019, 56, 146–155. [Google Scholar] [CrossRef] [PubMed]
- Neely, M.D.; Davison, C.A.; Aschner, M.; Bowman, A.B. From the Cover: Manganese and Rotenone-Induced Oxidative Stress Signatures Differ in iPSC-Derived Human Dopamine Neurons. Toxicol. Sci. 2017, 159, 366–379. [Google Scholar] [CrossRef]
- Du, K.; Liu, M.; Pan, Y.; Zhong, X.; Wei, M. Association of Serum Manganese Levels with Alzheimer’s Disease and Mild Cognitive Impairment: A Systematic Review and Meta-Analysis. Nutrients 2017, 9, 231. [Google Scholar] [CrossRef] [PubMed]
- Marcus, D.L.; Strafaci, J.A.; Freedman, M.L. Differential neuronal expression of manganese superoxide dismutase in Alzheimer’s disease. Med. Sci. Monit. 2006, 12, BR8–BR14. [Google Scholar]
- Hare, D.J.; Faux, N.G.; Roberts, B.R.; Volitakis, I.; Martins, R.N.; Bush, A.I. Lead and manganese levels in serum and erythrocytes in Alzheimer’s disease and mild cognitive impairment: Results from the Australian Imaging, Biomarkers and Lifestyle Flagship Study of Ageing. Metallomics 2016, 8, 628–632. [Google Scholar] [CrossRef] [PubMed]
- Balmuș, I.M.; Strungaru, S.A.; Ciobica, A.; Nicoara, M.N.; Dobrin, R.; Plavan, G.; Ștefănescu, C. Preliminary Data on the Interaction between Some Biometals and Oxidative Stress Status in Mild Cognitive Impairment and Alzheimer’s Disease Patients. Oxid. Med. Cell Longev. 2017, 2017, 7156928. [Google Scholar] [CrossRef] [PubMed]
- Calderon-Garciduenas, L.; D’Angiulli, A.; Kulesza, R.J.; Torres-Jardon, R.; Osnaya, N.; Romero, L.; Keefe, S.; Herritt, L.; Brooks, D.M.; Avila-Ramirez, J.; et al. Air pollution is associated with brainstem auditory nuclei pathology and delayed brainstem auditory evoked potentials. Int. J. Dev. Neurosci. 2011, 29, 365–375. [Google Scholar] [CrossRef] [PubMed]
- Guilarte, T.R.; Burton, N.C.; Verina, T.; Prabhu, V.V.; Becker, K.G.; Syversen, T.; Schneider, J.S. Increased APLP1 expression and neurodegeneration in the frontal cortex of manganese-exposed non-human primates. J. Neurochem. 2008, 105, 1948–1959. [Google Scholar] [CrossRef] [PubMed]
- Benskey, M.J.; Perez, R.G.; Manfredsson, F.P. The contribution of alpha synuclein to neuronal survival and function—Implications for Parkinson’s disease. J. Neurochem. 2016, 137, 331–359. [Google Scholar] [CrossRef] [PubMed]
- Binolfi, A.; Rasia, R.M.; Bertoncini, C.W.; Ceolin, M.; Zweckstetter, M.; Griesinger, C.; Jovin, T.M.; Fernandez, C.O. Interaction of alpha-synuclein with divalent metal ions reveals key differences: A link between structure, binding specificity and fibrillation enhancement. J. Am. Chem. Soc. 2006, 128, 9893–9901. [Google Scholar] [CrossRef] [PubMed]
- Xu, B.; Jin, C.H.; Deng, Y.; Liu, W.; Yang, T.Y.; Feng, S.; Xu, Z.F. Alpha-synuclein oligomerization in manganese-induced nerve cell injury in brain slices: A role of NO-mediated S-nitrosylation of protein disulfide isomerase. Mol. Neurobiol. 2014, 50, 1098–1110. [Google Scholar] [CrossRef]
- Mesulam, M.M.; Lalehzari, N.; Rahmani, F.; Ohm, D.; Shahidehpour, R.; Kim, G.; Gefen, T.; Weintraub, S.; Bigio, E.; Geula, C. Cortical cholinergic denervation in primary progressive aphasia with Alzheimer pathology. Neurology 2019, 92, e1580–e1588. [Google Scholar] [CrossRef]
- Fernsebner, K.; Zorn, J.; Kanawati, B.; Walker, A.; Michalke, B. Manganese leads to an increase in markers of oxidative stress as well as to a shift in the ratio of Fe(II)/(III) in rat brain tissue. Metallomics 2014, 6, 921–931. [Google Scholar] [CrossRef] [PubMed]
- Chtourou, Y.; Fetoui, H.; Garoui, E.M.; Boudawara, T.; Zeghal, N. Improvement of cerebellum redox states and cholinergic functions contribute to the beneficial effects of silymarin against manganese-induced neurotoxicity. Neurochem. Res. 2012, 37, 469–479. [Google Scholar] [CrossRef]
- Hardy, J.; Selkoe, D.J. The amyloid hypothesis of Alzheimer’s disease: Progress and problems on the road to therapeutics. Science 2002, 297, 353–356. [Google Scholar] [CrossRef]
- Wallin, C.; Kulkarni, Y.S.; Abelein, A.; Jarvet, J.; Liao, Q.; Strodel, B.; Olsson, L.; Luo, J.; Abrahams, J.P.; Sholts, S.B.; et al. Characterization of Mn(II) ion binding to the amyloid-beta peptide in Alzheimer’s disease. J. Trace Elem. Med. Biol. 2016, 38, 183–193. [Google Scholar] [CrossRef]
- Ling, J.; Yang, S.; Huang, Y.; Wei, D.; Cheng, W. Identifying key genes, pathways and screening therapeutic agents for manganese-induced Alzheimer disease using bioinformatics analysis. Medicine 2018, 97, e10775. [Google Scholar] [CrossRef]
- Walker, F.O. Huntington’s disease. Lancet 2007, 369, 218–228. [Google Scholar] [CrossRef]
- Damiano, M.; Galvan, L.; Déglon, N.; Brouillet, E. Mitochondria in Huntington’s disease. Biochim. Biophys. Acta 2010, 1802, 52–61. [Google Scholar] [CrossRef]
- Heng, M.Y.; Detloff, P.J.; Wang, P.L.; Tsien, J.Z.; Albin, R.L. In vivo evidence for NMDA receptor-mediated excitotoxicity in a murine genetic model of Huntington disease. J. Neurosci. 2009, 29, 3200–3205. [Google Scholar] [CrossRef]
- Strand, A.D.; Baquet, Z.C.; Aragaki, A.K.; Holmans, P.; Yang, L.; Cleren, C.; Beal, M.; Jones, L.; Kooperberg, C.; Olson, J.M.; et al. Expression profiling of Huntington’s disease models suggests that brain-derived neurotrophic factor depletion plays a major role in striatal degeneration. J. Neurosci. 2007, 27, 11758–11768. [Google Scholar] [CrossRef] [PubMed]
- Wright, R.O.; Baccarelli, A. Metals and neurotoxicology. J. Nutr. 2007, 137, 2809–2813. [Google Scholar] [CrossRef] [PubMed]
- Williams, B.B.; Li, D.; Wegrzynowicz, M.; Vadodaria, B.K.; Anderson, J.G.; Kwakye, G.F.; Aschner, M.; Erikson, K.M.; Bowman, A.B. Disease-toxicant screen reveals a neuroprotective interaction between Huntington’s disease and manganese exposure. J. Neurochem. 2010, 112, 227–237. [Google Scholar] [CrossRef]
- Trettel, F.; Rigamonti, D.; Hilditch-Maguire, P.; Wheeler, V.C.; Sharp, A.H.; Persichetti, F.; Cattaneo, E.; MacDonald, M.E. Dominant phenotypes produced by the HD mutation in STHdh(Q111) striatal cells. Hum. Mol. Genet. 2000, 9, 2799–2809. [Google Scholar] [CrossRef] [PubMed]
- Bartzokis, G.; Cummings, J.; Perlman, S.; Hance, D.B.; Mintz, J. Increased basal ganglia iron levels in Huntington disease. Arch. Neurol. 1999, 56, 569–574. [Google Scholar] [CrossRef] [PubMed]
- Fitsanakis, V.A.; Zhang, N.; Anderson, J.G.; Erikson, K.M.; Avison, M.J.; Gore, J.C.; Aschner, M. Measuring brain manganese and iron accumulation in rats following 14 weeks of low-dose manganese treatment using atomic absorption spectroscopy and magnetic resonance imaging. Toxicol. Sci. 2008, 103, 116–124. [Google Scholar] [CrossRef] [PubMed]
- Liu, M.; Sun, X.; Chen, B.; Dai, R.; Xi, Z.; Xu, H. Insights into Manganese Superoxide Dismutase and Human Diseases. Int. J. Mol. Sci. 2022, 23, 15893. [Google Scholar] [CrossRef]
- Pfalzer, A.C.; Wilcox, J.M.; Codreanu, S.G.; Totten, M.; Bichell, T.J.V.; Halbesma, T.; Umashanker, P.; Yang, K.L.; Parmalee, N.L.; Sherrod, S.D.; et al. Huntington’s disease genotype suppresses global manganese-responsive processes in pre-manifest and manifest YAC128 mice. Metallomics 2020, 12, 1118–1130. [Google Scholar] [CrossRef]
- Griffiths, G.M.; Fox, J.T. Rhythm in epilepsy. Lancet 1938, 232, 409–416. [Google Scholar] [CrossRef]
- Gonzalez-Reyes, R.E.; Gutierrez-Alvarez, A.M.; Moreno, C.B. Manganese and epilepsy: A systematic review of the literature. Brain Res. Rev. 2007, 53, 332–336. [Google Scholar] [CrossRef] [PubMed]
- Carl, G.F.; Keen, C.L.; Gallagher, B.B.; Clegg, M.S.; Littleton, W.H.; Flannery, D.B.; Hurley, L.S. Association of low blood manganese concentrations with epilepsy. Neurology 1986, 36, 1584–1587. [Google Scholar] [CrossRef] [PubMed]
- Carl, G.; Gallagher, B. Manganese and epilepsy. In Manganese in Health and Disease; Klimis-Tavantzis, D., Ed.; CRC Press, Inc.: Boca Raton, FL, USA, 1994; pp. 133–157. [Google Scholar]
- Choi, C.J.; Anantharam, V.; Martin, D.P.; Nicholson, E.M.; Richt, J.A.; Kanthasamy, A.; Kanthasamy, A.G. Manganese upregulates cellular prion protein and contributes to altered stabilization and proteolysis: Relevance to role of metals in pathogenesis of prion disease. Toxicol. Sci. 2010, 115, 535–546. [Google Scholar] [CrossRef]
- Martins, A.C., Jr.; Gubert, P.; Villas Boas, G.R.; Meirelles Paes, M.; Santamaría, A.; Lee, E.; Tinkov, A.A.; Bowman, A.B.; Aschner, M. Manganese-induced neurodegenerative diseases and possible therapeutic approaches. Expert. Rev. Neurother. 2020, 20, 1109–1121. [Google Scholar] [CrossRef]
- Choi, E.K.; Nguyen, T.T.; Gupta, N.; Iwase, S.; Seo, Y.A. Functional analysis of SLC39A8 mutations and their implications for manganese deficiency and mitochondrial disorders. Sci. Rep. 2018, 8, 3163. [Google Scholar] [CrossRef]
- Maret, W. Metalloproteomics, metalloproteomes, and the annotation of metalloproteins. Metallomics 2010, 2, 117–125. [Google Scholar] [CrossRef]
- Hood, M.I.; Skaar, E.P. Nutritional immunity: Transition metals at the pathogen-host interface. Nat. Rev. Microbiol. 2012, 10, 525–537. [Google Scholar] [CrossRef]
- Haider, B.A.; Bhutta, Z.A. The effect of therapeutic zinc supplementation among young children with selected infections: A review of the evidence. Food Nutr. Bull. 2009, 30 (Suppl. S1), S41–S59. [Google Scholar] [CrossRef]
- Kumar, V.; Choudhry, V.P. Iron deficiency and infection. Indian J. Pediatr. 2010, 77, 789–793. [Google Scholar] [CrossRef] [PubMed]
- Lassi, Z.S.; Haider, B.A.; Bhutta, Z.A. Zinc supplementation for the prevention of pneumonia in children aged 2 months to 59 months. Cochrane Database Syst. Rev. 2010, 12, CD005978. [Google Scholar] [CrossRef]
- Lisher, J.P.; Giedroc, D.P. Manganese acquisition and homeostasis at the host-pathogen interface. Front. Cell. Infect. Microbiol. 2013, 3, 91. [Google Scholar] [CrossRef] [PubMed]
- Jacobsen, F.E.; Kazmierczak, K.M.; Lisher, J.P.; Winkler, M.E.; Giedroc, D.P. Interplay between manganese and zinc homeostasis in the human pathogen Streptococcus pneumoniae. Metallomics 2011, 3, 38–41. [Google Scholar] [CrossRef]
- Veyrier, F.J.; Boneca, I.G.; Cellier, M.F.; Taha, M.K. A novel metal transporter mediating manganese export (MntX) regulates the Mn to Fe intracellular ratio and Neisseria meningitidis virulence. PLoS Pathog. 2011, 7, e1002261. [Google Scholar] [CrossRef]
- Ogunniyi, A.D.; Mahdi, L.K.; Jennings, M.P.; McEwan, A.G.; McDevitt, C.A.; Van der Hoek, M.B.; Bagley, C.J.; Hoffmann, P.; Gould, K.A.; Paton, J.C. Central role of manganese in regulation of stress responses, physiology, and metabolism in Streptococcus pneumoniae. J. Bacteriol. 2010, 192, 4489–4497. [Google Scholar] [CrossRef] [PubMed]
- Perry, R.D.; Craig, S.K.; Abney, J.; Bobrov, A.G.; Kirillina, O.; Mier, I.; Truszczynska, H.; Fetherston, J.D. Manganese transporters Yfe and MntH are Fur-regulated and important for the virulence of Yersinia pestis. Microbiology 2012, 158, 804–815. [Google Scholar] [CrossRef]
- Wichgers Schreur, P.J.; Rebel, J.M.; Smits, M.A.; van Putten, J.P.; Smith, H.E. TroA of Streptococcus suis is required for manganese acquisition and full virulence. J. Bacteriol. 2011, 193, 5073–5080. [Google Scholar] [CrossRef]
- Corbin, B.D.; Seeley, E.H.; Raab, A.; Feldmann, J.; Miller, M.R.; Torres, V.J.; Anderson, K.L.; Dattilo, B.M.; Dunman, P.M.; Gerads, R.; et al. Metal chelation and inhibition of bacterial growth in tissue abscesses. Science 2008, 319, 962–965. [Google Scholar] [CrossRef]
- Forbes, J.R.; Gros, P. Divalent-metal transport by NRAMP proteins at the interface of host-pathogen interactions. Trends Microbiol. 2001, 9, 397–403. [Google Scholar] [CrossRef]
- Boycott, K.M.; Beaulieu, C.L.; Kernohan, K.D.; Gebril, O.H.; Mhanni, A.; Chudley, A.E.; Redl, D.; Qin, W.; Hampson, S.; Küry, S.; et al. Autosomal-Recessive Intellectual Disability with Cerebellar Atrophy Syndrome Caused by Mutation of the Manganese and Zinc Transporter Gene SLC39A8. Am. J. Hum. Genet. 2015, 97, 886–893. [Google Scholar] [CrossRef]
- Riley, L.G.; Cowley, M.J.; Gayevskiy, V.; Roscioli, T.; Thorburn, D.R.; Prelog, K.; Bahlo, M.; Sue, C.M.; Balasubramaniam, S.; Christodoulou, J. A SLC39A8 variant causes manganese deficiency, and glycosylation and mitochondrial disorders. J. Inherit. Metab. Dis. 2017, 40, 261–269. [Google Scholar] [CrossRef]
- Nebert, D.W.; Liu, Z. SLC39A8 gene encoding a metal ion transporter: Discovery and bench to bedside. Hum. Genomics 2019, 13 (Suppl. S1), 51. [Google Scholar] [CrossRef]
- Lin, W.; Vann, D.R.; Doulias, P.T.; Wang, T.; Landesberg, G.; Li, X.; Ricciotti, E.; Scalia, R.; He, M.; Hand, N.J.; et al. Hepatic metal ion transporter ZIP8 regulates manganese homeostasis and manganese-dependent enzyme activity. J. Clin. Investig. 2017, 127, 2407–2417. [Google Scholar] [CrossRef] [PubMed]
- Gomes Martins, E.; Santos Silva, E.; Vilarinho, S.; Saudubray, J.M.; Vilarinho, L. Neonatal cholestasis: An uncommon presentation of hyperargininemia. J. Inherit. Metab. Dis. 2010, 33 (Suppl. S3), S503–S506. [Google Scholar] [CrossRef]
- Rossignol, F.; Wang, H.; Ferreira, C. Prolidase Deficiency. In GeneReviews® [Internet]; Adam, M.P., Everman, D.B., Mirzaa, G.M., Eds.; University of Washington: Seattle, WA, USA, 1993–2023. Available online: https://www.ncbi.nlm.nih.gov/books/NBK299584/ (accessed on 7 July 2022).
- Besio, R.; Gioia, R.; Cossu, F.; Monzani, E.; Nicolis, S.; Cucca, L.; Profumo, A.; Casella, L.; Tenni, R.; Bolognesi, M.; et al. Kinetic and Structural Evidences on Human Prolidase Pathological Mutants Suggest Strategies for Enzyme Functional Rescue. PLoS ONE 2013, 8, 58792. [Google Scholar] [CrossRef] [PubMed]
- Larrèque, M.; Charpentier, C.; Laidet, B.; Lambert, M.; Bressieux, J.M.; Prigent, F.; Canuel, C.; Tanzer, J. Déficit en prolidase et en manganèse. A propos d’une observation: Diagnostic et traitement [Prolidase and manganese deficiency. Apropos of a case: Diagnosis and treatment. Ann. Dermatol. Venereol. 1982, 109, 667–678. [Google Scholar]
- García-Rodríguez, N.; Díaz de la Loza Mdel, C.; Andreson, B.; Monje-Casas, F.; Rothstein, R.; Wellinger, R.E. Impaired manganese metabolism causes mitotic misregulation. Biol.Chem. 2012, 287, 18717–18729. [Google Scholar] [CrossRef] [PubMed]
- Tan, J.; Zhang, T.; Jiang, L.; Chi, J.; Hu, D.; Pan, Q.; Wang, D.; Zhang, Z. Regulation of intracellular manganese homeostasis by Kufor-Rakebsyndrome-associatedATP13A2protein. J. Biol. Chem. 2011, 286, 29654–29662. [Google Scholar] [CrossRef]
- Soldin, O.P.; Aschner, M. Effects of manganese on thyroid hormone homeostasis. Neurotoxicology 2007, 28, 951–956. [Google Scholar] [CrossRef] [PubMed]
- Garrick, M.D.; Dolan, K.G.; Horbinski, C.; Ghio, A.J.; Higgins, D.; Porubcin, M.; Moore, E.G.; Hainsworth, L.N.; Umbreit, J.N.; Conrad, M.E.; et al. DMT1: A mammalian transporter for multiple metals. Biometals 2003, 16, 41–54. [Google Scholar] [CrossRef] [PubMed]
- Holley, A.K.; Dhar, S.K.; Xu, Y.; St Clair, D.K. Manganese superoxide dismutase: Beyond life and death. Amino Acids 2012, 42, 139–158. [Google Scholar] [CrossRef]
- Bai, S.P.; Lu, L.; Wang, R.L.; Xi, L.; Zhang, L.Y.; Luo, X.G. Manganese source affects manganese transport and gene expression of divalent metal transporter 1 in the small intestine of broilers. Br. J. Nutr. 2012, 108, 267–276. [Google Scholar] [CrossRef]
- Cai, Q.; Shu, X.O.; Wen, W.; Cheng, J.R.; Dai, Q.; Gao, Y.T.; Zheng, W. Genetic polymorphism in the manganese superoxide dismutase gene, antioxidant intake, and breast cancer risk: Results from the Shanghai Breast Cancer Study. Breast Cancer Res. 2004, 6, R647–R655. [Google Scholar] [CrossRef]
- Liu, G.; Sun, G.; Wang, Y.; Wang, D.; Hu, W.; Zhang, J. Association between manganese superoxide dismutase gene polymorphism and breast cancer risk: A meta-analysis of 17,842 subjects. Mol. Med. Rep. 2012, 6, 797–804. [Google Scholar] [CrossRef]
- Hart, P.C.; Mao, M.; de Abreu, A.L.P.; Ansenberger-Fricano, K.; Ekoue, D.N.; Ganini, D.; Kajdacsy-Balla, A.; Diamond, A.M.; Minshall, R.D.; Consolaro, M.E.L.; et al. MnSOD upregulation sustains the Warburg effect via mitochondrial ROS and AMPK-dependent signalling in cancer. Nat. Commun. 2015, 6, 6053. [Google Scholar] [CrossRef]
- Becuwe, P.; Ennen, M.; Klotz, R.; Barbieux, C.; Grandemange, S. Manganese superoxide dismutase in breast cancer: From molecular mechanisms of gene regulation to biological and clinical signifi-cance. Free Radic. Biol. Med. 2014, 77, 139–151. [Google Scholar] [CrossRef] [PubMed]
- Juliachs, M.; Pujals, M.; Bellio, C.; Meo-Evoli, N.; Duran, J.M.; Zamora, E.; Parés, M.; Suñol, A.; Méndez, O.; Sánchez-Pla, A.; et al. Circulating SOD2 Is a Candidate Response Biomarker for Neoadjuvant Therapy in Breast Cancer. Cancers 2022, 14, 3858. [Google Scholar] [CrossRef]
- Kattan, Z.; Minig, V.; Leroy, P.; Dauça, M.; Becuwe, P. Role of manganese superoxide dismutase on growth and invasive properties of human estrogen-independent breast cancer cells. Breast Cancer Res. Treat. 2008, 108, 203–215. [Google Scholar] [CrossRef]
- Matsuoka, A.; Kizuka, F.; Lee, L.; Tamura, I.; Taniguchi, K.; Asada, H.; Taketani, T.; Tamura, H.; Sugino, N. Progesterone increases manganese superoxide dismutase expression via a cAMP-dependent signaling mediated by noncanonical Wnt5a pathway in human endometrial stromal cells. J. Clin. Endocrinol. Metab. 2010, 95, E291–E299. [Google Scholar] [CrossRef] [PubMed]
- Sugino, N.; Takiguchi, S.; Kashida, S.; Karube, A.; Nakamura, Y.; Kato, H. Superoxide dismutase expression in the human corpus luteum during the menstrual cycle and in early pregnancy. Mol. Hum. Reprod. 2000, 6, 19–25. [Google Scholar] [CrossRef] [PubMed]
- Sugino, N.; Hirosawa-Takamori, M.; Zhong, L.; Telleria, C.M.; Shiota, K.; Gibori, G. Hormonal regulation of copper-zinc superoxide dismutase and manganese superoxide dismutase messenger ribonucleic acid in the rat corpus luteum: Induction by prolactin and placental lactogens. Biol. Reprod. 1998, 59, 599–605. [Google Scholar] [CrossRef]
- Unfer, T.C.; Figueiredo, C.G.; Zanchi, M.M.; Maurer, L.H.; Kemerich, D.M.; Duarte, M.M.; Konopka, C.K.; Emanuelli, T. Estrogen plus progestin increase superoxide dismutase and total antioxidant capacity in postmenopausal women. Climacteric 2015, 18, 379–388. [Google Scholar] [CrossRef] [PubMed]
- Bureau, I.; Anderson, R.A.; Arnaud, J.; Raysiguier, Y.; Favier, A.E.; Roussel, A.M. Trace mineral status in post menopausal women: Impact of hormonal replacement therapy. J. Trace Elem. Med. Biol. 2002, 16, 9–13. [Google Scholar] [CrossRef] [PubMed]
- Rotter, I.; Kosik-Bogacka, D.; Dołęgowska, B.; Safranow, K.; Lubkowska, A.; Laszczyńska, M. Relationship between the concentrations of heavy metals and bioelements in aging men with metabolic syndrome. Int. J. Environ. Res. Public Health 2015, 12, 3944–3961. [Google Scholar] [CrossRef]
- Liu, D.; Liu, L.; Hu, Z.; Song, Z.; Wang, Y.; Chen, Z. Evaluation of the oxidative stress-related genes ALOX5, ALOX5AP, GPX1, GPX3 and MPO for contribution to the risk of type 2 diabetes mellitus in the Han Chinese population. Diab. Vasc. Dis. Res. 2018, 15, 336–339. [Google Scholar] [CrossRef] [PubMed]
- Zhou, B.; Su, X.; Su, D.; Zeng, F.; Wang, M.H.; Huang, L.; Huang, E.; Zhu, Y.; Zhao, D.; He, D.; et al. Dietary intake of manganese and the risk of the metabolic syndrome in a Chinese population. Br. J. Nutr. 2016, 116, 853–863. [Google Scholar] [CrossRef]
- Rhee, S.Y.; Hwang, Y.C.; Woo, J.T.; Sinn, D.H.; Chin, S.O.; Chon, S.; Kim, Y.S. Blood lead is significantly associated with metabolic syndrome in Korean adults: An analysis based on the Korea National Health and Nutrition Examination Survey (KNHANES), 2008. Cardiovasc. Diabetol. 2013, 12, 9. [Google Scholar] [CrossRef]
- Wong, M.M.H.; Chan, K.Y.; Lo, K. Manganese Exposure and Metabolic Syndrome: A Systematic Review and Meta-Analysis. Nutrients 2022, 14, 825. [Google Scholar] [CrossRef]
- Zimmet, P.; Alberti, K.G.; Shaw, J. Global and societal implications of the diabetes epidemic. Nature 2001, 414, 782–787. [Google Scholar] [CrossRef]
- Johns, M.; Fyalka, R.; Shea, J.A.; Neumann, W.L.; Rausaria, S.; Msengi, E.N.; Imani-Nejad, M.; Zollars, H.; McPherson, T.; Schober, J.; et al. SR-135, a peroxynitrite decomposing catalyst, enhances β-cell function and survival in B6D2F1 mice fed a high fat diet. Arch. Biochem. Biophys. 2015, 577–578, 49–59. [Google Scholar] [CrossRef] [PubMed]
- Nishikawa, T.; Edelstein, D.; Du, X.L.; Yamagishi, S.; Matsumura, T.; Kaneda, Y.; Yorek, M.A.; Beebe, D.; Oates, P.J.; Hammes, H.P.; et al. Normalizing mitochondrial superoxide production blocks three pathways of hyperglycaemic damage. Nature 2000, 404, 787–790. [Google Scholar] [CrossRef] [PubMed]
- Lowell, B.B.; Shulman, G.I. Mitochondrial dysfunction and type 2 diabetes. Science 2005, 307, 384–387. [Google Scholar] [CrossRef]
- Mousavi, Z.; Hassanpourezatti, M.; Najafizadeh, P.; Rezagholian, S.; Rhamanifar, M.S.; Nosrati, N. Effects of Subcutaneous Injection MnO2 Micro- and Nanoparticles on Blood Glucose Level and Lipid Profile in Rat. Iran J. Med. Sci. 2016, 41, 518–524. [Google Scholar]
- Burlet, E.; Jain, S.K. Manganese supplementation increases adiponectin and lowers ICAM-1 and creatinine blood levels in Zucker type 2 diabetic rats, and downregulates ICAM-1 by upregulating adiponectin multimerization protein (DsbA-L) in endothelial cells. Mol. Cell Biochem. 2017, 429, 1–10. [Google Scholar] [CrossRef]
- Flores, C.R.; Puga, M.P.; Wrobel, K.; Garay Sevilla, M.E.; Wrobel, K. Trace elements status in diabetes mellitus type 2: Possible role of the interaction between molybdenum and copper in the progress of typical complications. Diabetes Res. Clin. Pract. 2011, 91, 333–341. [Google Scholar] [CrossRef]
- Badran, M.; Morsy, R.; Soliman, H.; Elnimr, T. Assessment of trace elements levels in patients with Type 2 diabetes using multivariate statistical analysis. J. Trace Elem. Med. Biol. 2016, 33, 114–119. [Google Scholar] [CrossRef] [PubMed]
- Rambousková, J.; Krsková, A.; Slavíková, M.; Cejchanová, M.; Wranová, K.; Procházka, B.; Cerná, M. Trace elements in the blood of institutionalized elderly in the Czech Republic. Arch. Gerontol. Geriatr. 2013, 56, 389–394. [Google Scholar] [CrossRef]
- Rodríguez-Rodríguez, E.; Bermejo, L.M.; López-Sobaler, A.M.; Ortega, R.M. Una ingesta inadecuada de manganeso podría contribuir a la aparición de resistencia a la insulina en niñas [An inadequate intake of manganese may favour insulin resistance in girls]. Nutr. Hosp. 2011, 26, 965–970. (In Spanish) [Google Scholar]
- Koh, E.S.; Kim, S.J.; Yoon, H.E.; Chung, J.H.; Chung, S.; Park, C.W.; Chang, Y.S.; Shin, S.J. Association of blood manganese level with diabetes and renal dysfunction: A cross-sectional study of the Korean general population. BMC Endocr. Disord. 2014, 14, 24. [Google Scholar] [CrossRef]
- Wang, X.; Zhang, M.; Lui, G.; Chang, H.; Zhang, M.; Liu, W.; Li, Z.; Liu, Y.; Huang, G. Associations of Serum Manganese Levels with Prediabetes and Diabetes among ≥60-Year-Old Chinese Adults: A Population-Based Cross-Sectional Analysis. Nutrients 2016, 8, 497. [Google Scholar] [CrossRef]
- Kosenko, L.G. Concentration of trace elements in blood of patients with diabetes mellitus. Fed. Proc. Transl. Suppl. 1965, 24, 237–238. [Google Scholar] [PubMed]
- Walter, R.M., Jr.; Uriu–Hare, J.Y.; Olin, K.L.; Oster, M.H.; Anawalt, B.D.; Critchfield, J.W.; Keen, C.L. Copper, zinc, manganese, and magnesium status and complications of diabetes mellitus. Diabetes Care 1991, 14, 1050–1056. [Google Scholar] [CrossRef]
- Serdar, M.A.; Bakir, F.; Haşimi, A.; Celik, T.; Akin, O.; Kenar, L.; Aykut, O.; Yildirimkaya, M. Trace and toxic element patterns in nonsmoker patients with noninsulin-dependent diabetes mellitus, impaired glucose tolerance, and fasting glucose. Int. J. Diabetes Dev. Ctries. 2009, 29, 35–40. [Google Scholar] [CrossRef] [PubMed]
- Ekmekcioglu, C.; Prohaska, C.; Pomazal, K.; Steffan, I.; Schernthaner, G.; Marktl, W. Concentrations of seven trace elements in different hematological matrices in patients with type 2 diabetes as compared to healthy controls. Biol. Trace Elem. Res. 2001, 79, 205–219. [Google Scholar] [CrossRef]
- Kazi, T.G.; Afridi, H.I.; Kazi, N.; Jamali, M.K.; Arain, M.B.; Jalbani, N.; Kandhro, G.A. Copper, chromium, manganese, iron, nickel, and zinc levels in biological samples of diabetes mellitus patients. Biol. Trace Elem. Res. 2008, 122, 1–18. [Google Scholar] [CrossRef]
- Forte, G.; Bocca, B.; Peruzzu, A.; Tolu, F.; Asara, Y.; Farace, C.; Oggiano, R.; Madeddu, R. Blood metals concentration in type 1 and type 2 diabetics. Biol. Trace Elem. Res. 2013, 156, 79–90. [Google Scholar] [CrossRef] [PubMed]
- Adewumi, M.T.; Njoku, C.H.; Saidu, Y.; Abubakar, M.K.; Shehu, R.A.; Bilbis, L.S. Serum chromium, copper and manganese levels of diabetic subjects in Katsina, Nigeria. Asian J. Biochem. 2007, 2, 284–288. [Google Scholar] [CrossRef]
- Berry, C.; Tardif, J.C.; Bourassa, M.G. Coronary heart disease in patients with diabetes: Part I: Recent advances in prevention and noninvasive management. J. Am. Coll. Cardiol. 2007, 49, 631–642. [Google Scholar] [CrossRef] [PubMed]
- Lisun-Lobanova, V.P. Trace elements (manganese, copper, zinc) in patients with diabetes mellitus. Zdr. Beloruss. 1963, 9, 49–53. [Google Scholar]
- Ekin, S.; Mert, N.; Gunduz, H.; Meral, I. Serum sialic acid levels and selected mineral status in patients with type 2 diabetes mellitus. Biol. Trace Elem. Res. 2003, 94, 193–201. [Google Scholar] [CrossRef]
- Besio, R.; Baratto, M.C.; Gioia, R.; Monzani, E.; Nicolis, S.; Cucca, L.; Profumo, A.; Casella, L.; Basosi, R.; Tenni, R.; et al. A Mn(II)-Mn(II) center in human prolidase. Biochim. Biophys. Acta 2013, 1834, 197–204. [Google Scholar] [CrossRef] [PubMed]
- Eshak, E.S.; Muraki, I.; Imano, H.; Yamagishi, K.; Tamakoshi, A.; Iso, H. Manganese intake from foods and beverages is associated with a reduced risk of type 2 diabetes. Maturitas 2021, 143, 127–131. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Cui, Z.; Lu, W.; Wang, P.; Wang, J.; Zhou, Z.; Zhang, N.; Wang, Z.; Lin, T.; Song, Y.; et al. Association between serum manganese levels and diabetes in Chinese adults with hypertension. J. Clin. Hypertens. 2022, 24, 918–927. [Google Scholar] [CrossRef]
- Shan, Z.; Chen, S.; Sun, T.; Luo, C.; Guo, Y.; Yu, X.; Yang, W.; Hu, F.B.; Liu, L. U-Shaped Association between Plasma Manganese Levels and Type 2 Diabetes. Environ. Health Perspect. 2016, 124, 1876–1881. [Google Scholar] [CrossRef]
- Afridi, H.I.; Kazi, T.G.; Kazi, N.; Baig, J.A.; Jamali, M.K.; Arain, M.B.; Sarfraz, R.A.; Sheikh, H.U.; Kandhro, G.A.; Shah, A.Q. Status of essential trace metals in biological samples of diabetic mother and their neonates. Arch. Gynecol. Obstet. 2009, 280, 415–423. [Google Scholar] [CrossRef]
- Cancarini, A.; Fostinelli, J.; Napoli, L.; Gilberti, M.E.; Apostoli, P.; Semeraro, F. Trace elements and diabetes: Assessment of levels in tears and serum. Exp. Eye Res. 2017, 154, 47–52. [Google Scholar] [CrossRef]
- Clegg, M.S.; Donovan, S.M.; Monaco, M.H.; Baly, D.L.; Ensunsa, J.L.; Keen, C.L. The influence of manganese deficiency on serum IGF-1 and IGF binding proteins in the male rat. Proc. Soc. Exp. Biol. Med. 1998, 219, 41–47. [Google Scholar] [CrossRef]
- Baly, D.L.; Schneiderman, J.S.; Garcia-Welsh, A.L. Effect of manganese deficiency on insulin binding, glucose transport and metabolism in rat adipocytes. J. Nutr. 1990, 120, 1075–1079. [Google Scholar] [CrossRef] [PubMed]
- Strause, L.; Saltman, P. Role of manganese in bone metabolism. In Nutritional Bioavailability of Manganese; Keys, C., Ed.; American Chemical Society: Washington, DC, USA, 1987; ISBN 0841214336. [Google Scholar]
- Pepa, G.D.; Brandi, M.L. Microelements for bone boost: The last but not the least. Clin. Cases Miner. Bone Metab. 2016, 13, 181–185. [Google Scholar] [CrossRef] [PubMed]
- Botre, C.; Shahu, A.; Adkar, N.; Shouche, Y.; Ghaskadbi, S.; Ashma, R. Superoxide Dismutase 2 Polymorphisms and Osteoporosis in Asian Indians: A Genetic Association Analysis. Cell. Mol. Biol. Lett. 2015, 20, 685–697. [Google Scholar] [CrossRef] [PubMed]
- Culotta, V.C.; Yang, M.; O’Halloran, T.V. Activation of superoxide dismutases: Putting the metal to the pedal. Biochim. Biophys. Acta. 2006, 1763, 747–758. [Google Scholar] [CrossRef]
- Guo, T.; Zhang, L.; Konermann, A.; Zhou, H.; Jin, F.; Liu, W. Manganese superoxide dismutase is required to maintain osteoclast differentiation and function under static force. Sci. Rep. 2015, 5, 8016. [Google Scholar] [CrossRef]
- Powers, K.M.; Oberley, L.W.; Domann, F.E. The Adventures of Superoxide Dismutase in Health and Disease: Superoxide in the Balance. In Oxidants in Biology; Valacchi, G., Davis, P.A., Eds.; Springer: Dordrecht, The Netherlands, 2008. [Google Scholar] [CrossRef]
- Fernandez-Twinn, D.S.; Ozanne, S.E. Early life nutrition and metabolic programming. Ann. N. Y. Acad. Sci. 2010, 1212, 78–96. [Google Scholar] [CrossRef]
- Pires, K.M.; Ilkun, O.; Valente, M.; Boudina, S. Treatment with a SOD mimetic reduces visceral adiposity, adipocyte death, and adipose tissue inflammation in high fat-fed mice. Obesity 2014, 22, 178–187. [Google Scholar] [CrossRef]
- Krautbauer, S.; Eisinger, K.; Neumeier, M.; Hader, Y.; Buettner, R.; Schmid, P.M.; Aslanidis, C.; Buechler, C. Free fatty acids, lipopolysaccharide and IL-1α induce adipocyte manganese superoxide dismutase which is increased in visceral adipose tissues of obese rodents. PLoS ONE 2014, 9, e86866. [Google Scholar] [CrossRef]
- Han, Y.H.; Buffolo, M.; Pires, K.M.; Pei, S.; Scherer, P.E.; Boudina, S. Adipocyte-Specific Deletion of Manganese Superoxide Dismutase Protects From Diet-Induced Obesity Through Increased Mitochondrial Uncoupling and Biogenesis. Diabetes 2016, 65, 2639–2651. [Google Scholar] [CrossRef]
- Fan, Y.; Zhang, C.; Bu, J. Relationship between Selected Serum Metallic Elements and Obesity in Children and Adolescent in the U.S. Nutrients 2017, 9, 104. [Google Scholar] [CrossRef]
- Glover, M.; Hebert, V.Y.; Nichols, K.; Xue, S.Y.; Thibeaux, T.M.; Zavecz, J.A.; Dugas, T.R. Overexpression of mitochondrial antioxidant manganese superoxide dismutase (MnSOD) provides protection against AZT- or 3TC-induced endothelial dysfunction. Antivir. Res. 2014, 111, 136–142. [Google Scholar] [CrossRef]
- Steppan, J.; Nyhan, D.; Berkowitz, D.E. Development of novel arginase inhibitors for therapy of endothelial dysfunction. Front. Immunol. 2013, 4, 278. [Google Scholar] [CrossRef] [PubMed]
- Hegyi, L.; Hardwick, S.J.; Siow, R.C.; Skepper, J.N. Macrophage death and the role of apoptosis in human atherosclerosis. J. Hematother. Stem Cell Res. 2001, 10, 27–42. [Google Scholar] [CrossRef] [PubMed]
- Ohashi, M.; Runge, M.S.; Faraci, F.M.; Heistad, D.D. MnSOD deficiency increases endothelial dysfunction in ApoE-deficient mice. Arterioscler. Thromb. Vasc. Biol. 2006, 26, 2331–2336. [Google Scholar] [CrossRef]
- Zhou, Q.; Einert, M.; Schmitt, H.; Wang, Z.; Pankratz, F.; Olivier, C.B.; Bode, C.; Liao, J.K.; Moser, M. MnTBAP increases BMPR-II expression in endothelial cells and attenuates vascular inflammation. Vascul. Pharmacol. 2016, 84, 67–73. [Google Scholar] [CrossRef]
- Mendis, S. Magnesium, zinc, and manganese in atherosclerosis of the aorta. Biol. Trace Elem. Res. 1989, 22, 251–256. [Google Scholar] [CrossRef]
- Ilyas, A.; Shah, M.H. Multivariate statistical evaluation of trace metal levels in the blood of atherosclerosis patients in comparison with healthy subjects. Heliyon 2016, 2, e00054. [Google Scholar] [CrossRef]
- Feldstein, A.E.; Canbay, A.; Guicciardi, M.E.; Higuchi, H.; Bronk, S.F.; Gores, G.J. Diet-induced hepatic steatosis sensitises Fas-mediated liver injury in mice. J. Hepatol. 2003, 39, 978–983. [Google Scholar] [CrossRef]
- Jaeschke, H.; Gores, G.J.; Cederbaum, A.I.; Hinson, J.A.; Pessayre, D.; Lemasters, J.J. Mechanisms of hepatotoxicity. Toxicol. Sci. 2002, 65, 166–176. [Google Scholar] [CrossRef] [PubMed]
- Fromenty, B.; Robin, M.A.; Igoudjil, A.; Mansouri, A.; Pessayre, D. The ins and outs of mitochondrial dysfunction in NASH. Diabetes Metab. 2004, 30, 121–138. [Google Scholar] [CrossRef]
- Wang, S.; Kuang, X.; Fang, Z.; Huang, Z.; Shi, P. Effect of oleic acid on the levels of eight metal ions in human hepatoma SMMC-7721 cells. Biol. Trace Elem. Res. 2014, 159, 445–450. [Google Scholar] [CrossRef] [PubMed]
- Rezazadeh, A.; Yazdanparast, R. Prevention of non-alcoholic steatohepatitis in rats by two manganese-salen complexes. Iran Biomed. J. 2014, 18, 41–48. [Google Scholar] [CrossRef] [PubMed]
- Srivastava, V.K.; Hiney, J.K.; Dees, W.L. Early life manganese exposure upregulates tumor-associated genes in the hypothalamus of female rats: Relationship to manganese-induced precocious puberty. Toxicol. Sci. 2013, 136, 373–381. [Google Scholar] [CrossRef] [PubMed]
- Lee, B.; Pine, M.; Johnson, L.; Rettori, V.; Hiney, J.K.; Dees, W.L. Manganese acts centrally to activate reproductive hormone secretion and pubertal development in male rats. Reprod. Toxicol. 2006, 22, 580–585. [Google Scholar] [CrossRef] [PubMed]
- Xie, J.; Tian, C.; Zhu, Y.; Zhang, L.; Lu, L.; Luo, X. Effects of inorganic and organic manganese supplementation on gonadotropin-releasing hormone-I and follicle-stimulating hormone expression and reproductive performance of broiler breeder hens. Poult. Sci. 2014, 93, 959–969. [Google Scholar] [CrossRef]
- Maciejewski, R.; Radzikowska-Büchner, E.; Flieger, W.; Kulczycka, K.; Baj, J.; Forma, A.; Flieger, J. An Overview of Essential Microelements and Common Metallic Nanoparticles and Their Effects on Male Fertility. Int. J. Environ. Res. Public Health 2022, 19, 11066. [Google Scholar] [CrossRef]
- Cheng, J.; Fu, J.L.; Zhou, Z.C. The inhibitory effects of manganese on steroidogenesis in rat primary Leydig cells by disrupting steroidogenic acute regulatory (StAR) protein expression. Toxicology 2003, 187, 139–148. [Google Scholar] [CrossRef] [PubMed]
- Skalnaya, M.G.; Serebryansky, E.P.; Yurasov, V.V.; Tinkov, A.A.; Demidov, V.A.; Skalny, A.V. Association between semen quality and level of 20 essential and toxic metals in ejaculate. Trace Elem. Electrolytes 2015, 32, 126–132. [Google Scholar] [CrossRef]
- Barber, S.J.; Parker, H.M.; McDaniel, C.D. Broiler breeder semen quality as affected by trace minerals in vitro. Poult. Sci. 2005, 84, 100–105. [Google Scholar] [CrossRef]
- Migliaccio, O.; Castellano, I.; Cirino, P.; Romano, G.; Palumbo, A. Maternal Exposure to Cadmium and Manganese Impairs Reproduction and Progeny Fitness in the Sea Urchin Paracentrotus lividus. PLoS ONE 2015, 10, e0131815. [Google Scholar] [CrossRef]
- Cheema, R.S.; Bansal, A.K.; Bilaspuri, G.S. Manganese Provides Antioxidant Protection for Sperm Cryopreservation that May Offer New Consideration for Clinical Fertility. Oxidative Med. Cell. Longev. 2009, 2, 152–159. [Google Scholar] [CrossRef]
- Tajaddini, S.; Ebrahimi, S.; Behnam, B.; Bakhtiyari, M.; Joghataei, M.T.; Abbasi, M.; Amini, M.; Amanpour, S.; Koruji, M. Antioxidant effect of manganese on the testis structure and sperm parameters of formalin-treated mice. Andrologia 2014, 46, 246–253. [Google Scholar] [CrossRef]
- Laskey, J.W.; Rehnberg, G.L.; Hein, J.F.; Carter, S.D. Effects of chronic manganese (Mn3O4) exposure on selected reproductive parameters in rats. J. Toxicol. Environ. Health Part A 1982, 9, 677–687. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Wu, J.; Zhou, W.; Gao, E. Effects of manganese on routine semen quality parameters: Results from a population-based study in China. BMC Public Health 2012, 12, 919. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.-X.; Sun, Y.; Huang, Z.; Wang, P.; Feng, W.; Li, J.; Yang, P.; Wang, M.; Sun, L.; Chen, Y.-J.; et al. Associations of urinary metal levels with serum hormones, spermatozoa apoptosis and sperm DNA damage in a Chinese population. Environ. Int. 2016, 94, 177–188. [Google Scholar] [CrossRef]
- Wirth, J.J.; Rossano, M.G.; Daly, D.C.; Paneth, N.; Puscheck, E.; Potter, R.C.; Diamond, M.P. Ambient Manganese Exposure is Negatively Associated with Human Sperm Motility and Concentration. Epidemiology 2007, 18, 270–273. [Google Scholar] [CrossRef]
- Emara, A.M.; El-Ghawabi, S.H.; Madkour, O.I.; El-Samra, G.H. Chronic manganese poisoning in the dry battery industry. Br. J. Ind. Med. 1971, 28, 78–82. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.; Wang, J.; Yang, X.; Wu, F.; Qi, Z.; Xu, B.; Liu, W.; Deng, Y. Occupational manganese exposure, reproductive hormones, and semen quality in male workers: A cross-sectional study. Toxicol. Ind. Health 2019, 35, 53–62. [Google Scholar] [CrossRef]
- Kim, K.; Wactawski-Wende, J.; Michels, K.A.; Schliep, K.C.; Plowden, T.C.; Chaljub, E.N.; Mumford, S.L. Dietary minerals, reproductive hormone levels and sporadic anovulation: Associations in healthy women with regular menstrual cycles. Br. J. Nutr. 2018, 120, 81–89. [Google Scholar] [CrossRef]
- Skalnaya, M.G.; Tinkov, A.A.; Lobanova, Y.N.; Chang, J.S.; Skalny, A.V. Serum levels of copper, iron, and manganese in women with pregnancy, miscarriage, and primary infertility. J. Trace Elem. Med. Biol. 2019, 56, 124–130. [Google Scholar] [CrossRef]
- Johns Hopkins University Bloomberg School of Public Health. (2020, July 7). ScienceDaily. Available online: www.sciencedaily.com/releases/2020/07/200707113245.htm (accessed on 26 July 2023).
- Nandakumaran, M.; Al-Sannan, B.; Al-Sarraf, H.; Al-Shammari, M. Maternal-fetal transport kinetics of manganese in perfused human placental lobule in vitro. J. Matern. Neonatal Med. 2016, 29, 274–278. [Google Scholar] [CrossRef]
- Studer, J.M.; Schweer, W.P.; Gabler, N.K.; Ross, J.W. Functions of manganese in reproduction. Anim. Reprod. Sci. 2022, 238, 106924. [Google Scholar] [CrossRef] [PubMed]
- Hostetler, C.E.; Kincaid, R.L.; Mirando, M.A. The role of essential trace elements in embryonic and fetal development in livestock. Vet. J. 2003, 166, 125–139. [Google Scholar] [CrossRef] [PubMed]
- Studer, J.M.; Kiefer, Z.E.; Goetz, B.M.; Keating, A.F.; Baumgard, L.H.; Rambo, Z.J.; Schweer, W.P.; Wilson, M.E.; Rapp, C.; Ross, J.W. Impact of manganese amino acid complex on tissue-specific trace mineral distribution and corpus luteum function in gilts. J. Anim. Sci. 2021, 99, skab155. [Google Scholar] [CrossRef] [PubMed]
- Guo, M. Human Milk Biochemistry and Infant Formula Manufacturing Technology, 2nd ed.; Woodhead Publishing: Sawston, UK, 2020. [Google Scholar]
- Tenaud, I.; Sainte-Marie, I.; Jumbou, O.; Litoux, P.; Dréno, B. In vitro modulation of keratinocyte wound healing integrins by zinc, copper and manganese. Br. J. Dermatol. 1999, 140, 26–34. [Google Scholar] [CrossRef]
- Tenaud, I.; Saiagh, I.; Dreno, B. Addition of zinc and manganese to a biological dressing. J. Dermatolog. Treat. 2009, 20, 90–93. [Google Scholar] [CrossRef]
- Chebassier, N.; Ouijja, E.H.; Viegas, I.; Dreno, B. Stimulatory effect of boron and manganese salts on keratinocyte migration. Acta Derm. Venereol. 2004, 84, 191–194. [Google Scholar] [CrossRef]
- Ma, H.; Zhou, Q.; Chang, J.; Wu, C. Grape Seed-Inspired Smart Hydrogel Scaffolds for Melanoma Therapy and Wound Healing. ACS Nano 2019, 13, 4302–4311. [Google Scholar] [CrossRef]
- Chen, J.; Zhang, W.J.; Guo, Z.; Wang, H.B.; Wang, D.D.; Zhou, J.J.; Chen, Q.W. pH-responsive iron manganese silicate nanoparticles as T1-T2* dual-modal imaging probes for tumor diagnosis. ACS Appl. Mater. Interfaces 2015, 7, 5373–5383. [Google Scholar] [CrossRef]
- Ma, W.; Zhang, H.; Ma, H.; Wu, C. Iron manganese silicate incorporated bioactive hydrogels for therapy of skin tumor. Prog. Nat. Sci. Mater. Int. 2022, 32, 171–178. [Google Scholar] [CrossRef]
- Lu, M.; Li, S.; Xiong, X.; Huang, Z.; Xu, B.; Liu, Y.; Wu, Q.; Wu, N.; Liu, H.; Zhou, D. A Photo-Responsive Hollow Manganese/Carbon Hybrid Nanosphere for Wound Disinfection and Healing. Adv. Funct. Mater. 2022, 32, 2208061. [Google Scholar] [CrossRef]
- Shahriari-Khalaji, M.; Hong, S.; Hu, G.; Ji, Y.; Hong, F.F. Bacterial Nanocellulose-Enhanced Alginate Double-Network Hydrogels Cross-Linked with Six Metal Cations for Antibacterial Wound Dressing. Polymers 2020, 12, 2683. [Google Scholar] [CrossRef] [PubMed]
- Matta, F.V.; Xiong, J.; Lila, M.A.; Ward, N.I.; Felipe-Sotelo, M.; Esposito, D. Chemical Composition and Bioactive Properties of Commercial and Non-Commercial Purple and White Açaí Berries. Foods 2020, 9, 1481. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Zhang, J.; Wu, L.; Chen, L.; Niu, P.; Li, J. Glutamine supplementation reverses manganese neurotoxicity by eliciting the mitochondrial unfolded protein response. iScience 2023, 26, 107136. [Google Scholar] [CrossRef]






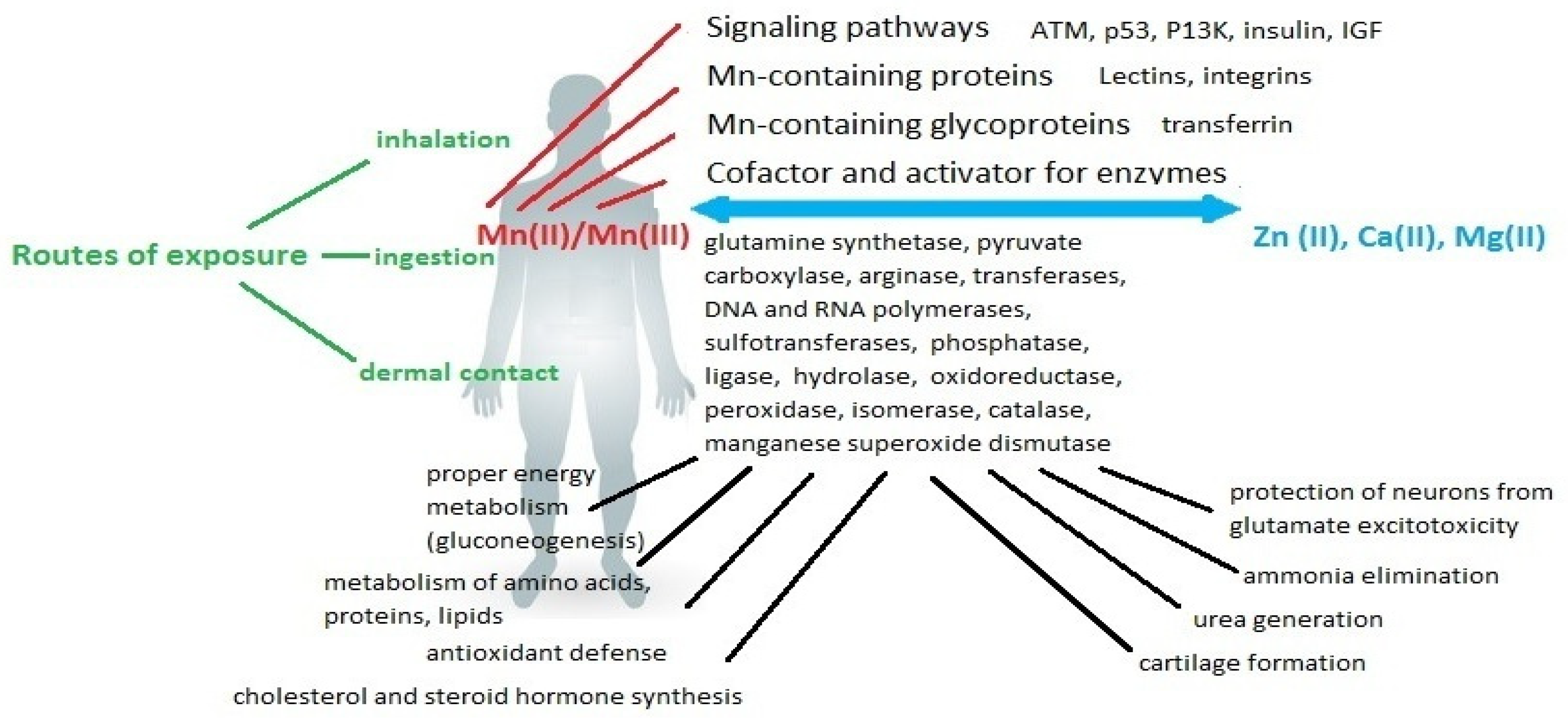{kind=link}
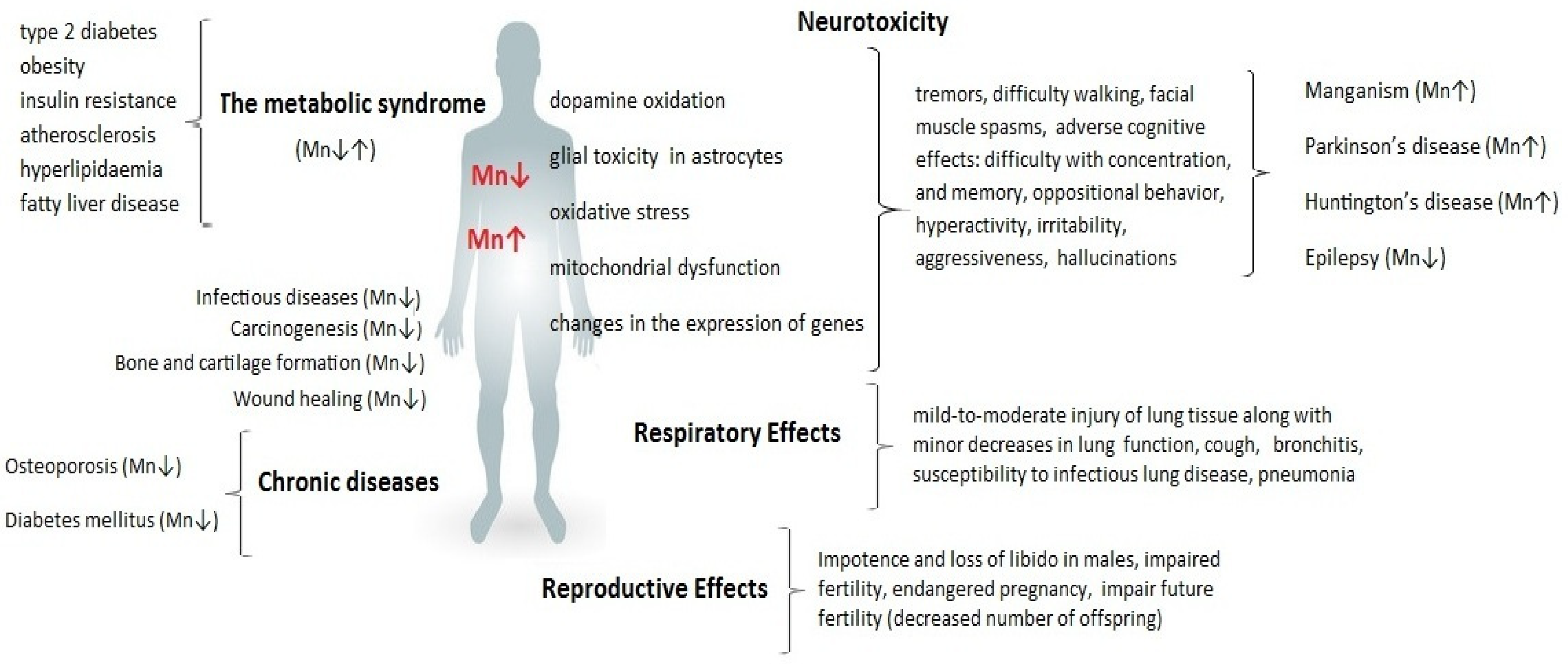{kind=link}
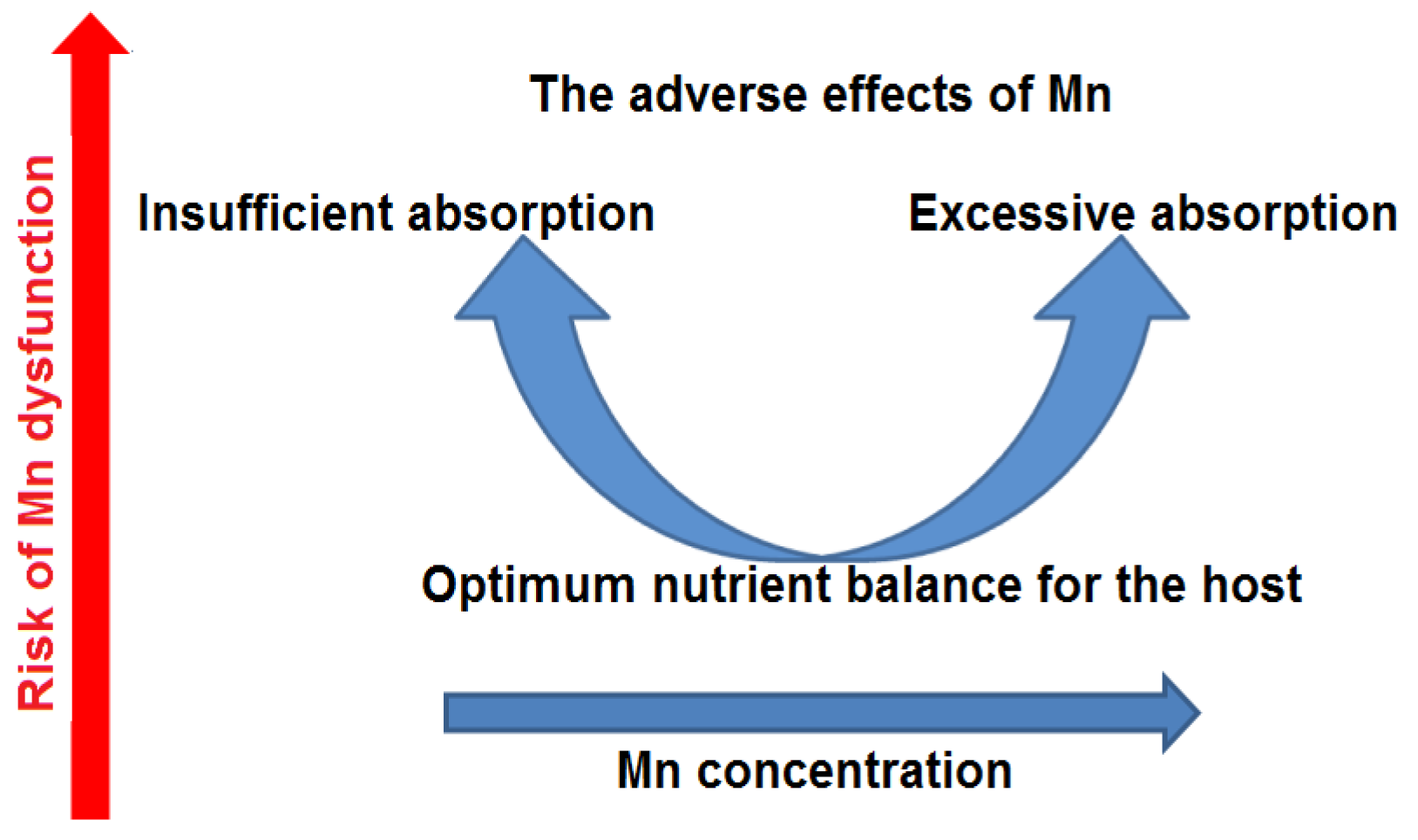{kind=link}
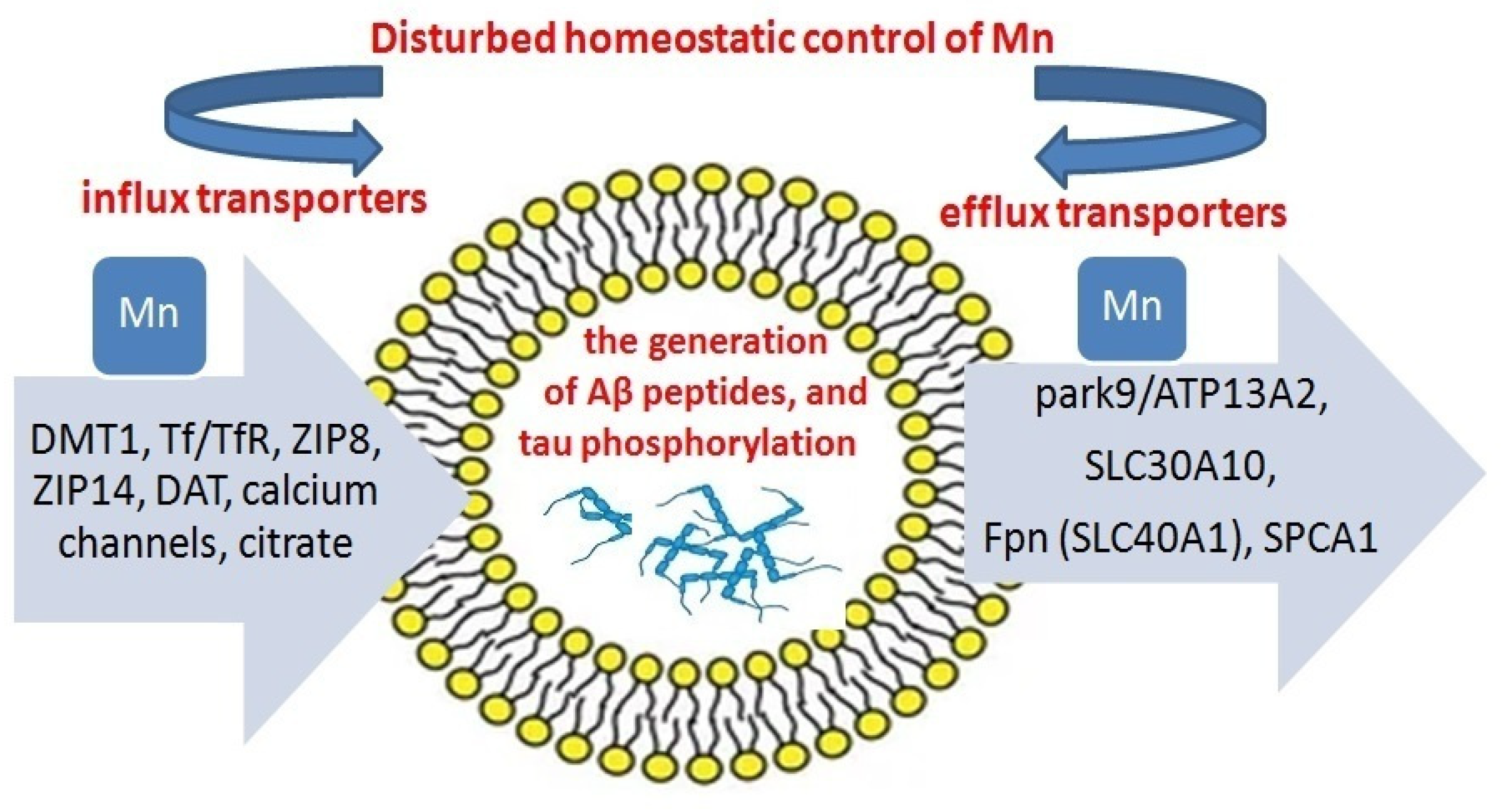{kind=link}
{kind=link}
| Matrix | Mn Concentration | Population | Ages | Location | Method | Ref. |
|---|---|---|---|---|---|---|
| plasma | mean: 14.3 ± 11.4 nmol L−1 range: 3–27.5 nmol L−1 | 68 | 22–75 y | Germany | ETAAS | [91] |
| mean: 25.5 ± 22.8 nmol L−1 range: 2.7–48.3 nmol L−1 | 129 | 1 mth–18 y | ||||
| serum | range: 4.7–215 nmol L−1 | 584 | 16–18 y | India | GFAAS | [121] |
| serum | 2.36 µg L−1 | 50 | 35–70 y | Italy | SEC-ICP-DRC-MS | [40] |
| blood | mean: 10.240 ± 2.834 range: 6.597–15.740 μg L−1 | 200 (97 f, 103 m) | 3–14 y | Kinshasa Congo | ICP-MS | [122] |
| urine | mean: 0.510 ± 1.643 range: 0.07–1.703 μg L−1; 0.948 ± 5.254 μg g−1 of creatinine | |||||
| blood | female: 1.403 µg/dL male: 1.192 µg/dL | 2005 | >20 y | Korea | GFAAS | [123] |
| blood, urine | blood: 10.5–17.9 μg L−1 urine: 0.133–0.410 μg L−1 | 3–11 y | USA | [124] | ||
| blood, scalp hair, fingernails | blood: 9.95 µg L−1 scalp hair: 380.98 ng g−1 fingernails: 936.71 ng g−1 | 130 (95 f, 35 m) | 20–71 y | Spain | ICP-MS | [125] |
| blood | pre- menopausal: 14.41 µg L−1 post-: 12.92 µg L−1 | 1826 f | 20–60 y | Korea | GFAAS | [126] |
| blood | 10–20 wk: 150.4 ± 53.4 nmol L−1 25 wk: 171.6 ± 59.7 nmol L−1 34 wk: 230.0 ± 67.8 nmol L−1 neonate: 737.7 ± 209.7 nmol L−1 | 34 pregnant f (wk gestation), 34 neonate | pregnancy period, 3–4 day old | Australia | GFAAS | [127] |
| blood | pregnant: 2.4 µg/dL nonpregnant: 0.8–1.2 µg/dL | 470 | 14–36 y | Canada | DRC–ICP-MS | [128] |
| serum | 29.32 μg L−1 | 1183 (890 m, 293 f) | 18–65 y | Brazil | AAS | [129] |
| hair | mean: 0.320 mg g−1 range: 0.239–0.789 mg g−1 | 260 (122 f, 138 m) | 30–49 y | Kazakhstan | ICP-MS | [130] |
| blood, serum | blood: 9.03 ± 2.25µg L−1 serum: 1.82 ± 0.64µg L−1 | 80 (34 f, 26 m) | 18–64 y | USA | ZAAS, NAA | [131] |
| hair | males: 0.59 ± 0.61 µg g−1 females: 1.09 ± 1.43 µg g−1 mean: 0.95 ± 1.27µg g−1 range: 0.29–1.76 µg g−1 | 7256 (5161 f, 2095 m) | 20–60 y | Russia | ICP-MS | [132] |
| blood | range: 1.5–22 µg L−1 mean: 7.40µg L−1 | 206 | 16–70 y | United Kingdom | ETAAS, ICP-MS | [47] |
| urine | range: 0.09–7.8µg L−1 mean: 0.3 µg/L | 188 | ||||
| blood | range: 7.0–14.1 µg L−1 median: 9.8 µg L−1 | 99 | <5 y (n = 17) 5–15 (n = 31) 15–18 (n = 20) | France | ICP-MS | [133] |
| plasma | range: 0.53–2.21µg L−1 median: 0.97µg L−1 | 96 | ||||
| blood | range: 5.9–13.3 µg L−1 | 106 | 25–55 y | France | ICP-MS | [134] |
| plasma | range: 0.35–1.08µg L−1 | |||||
| plasma | range: 3.0–27.5 nmol L−1 mean: 14.3 nmol L−1 | 68 | 22–75 y | Germany | ETAAS | [93] |
| range: 3.0–53 nmol L−1 mean: 25.5 nmol/L | 129 | 1 mth-18 y | ||||
| blood | range: 4.8–18 µg L−1; mean: 9.0 µg L−1 | 130 (80 f, 50 m) | 18–70 y | Germany | ICP–MS | [135] |
| blood, serum | blood: 144 ± 43.6 nmol L−1 serum: 28.4 ± 21.0 nm L−1 | 1016 | 70 y | Sweden | ICP-SFMS | [136] |
| hair | 0.33 ± 0.16 µg g−1 | 2 | - | Brazil, Thailand | LA-ICP-MS, ICP-MS | [137] |
| urine | 0.46 µg L−1 | 132 (82 m, 50 f) | 18–66 y | UK | ICP-MS | [53] |
| blood | range: 6.9–18.4 µg L−1 mean: 9.6 µg L−1 | 1125 (506 m, 619 f) | 18–60 y | Brazil | ICP-MS, ETAAS | [138] |
| blood | median: 10.4 μg L−1 | 100 (64 f, 36 m) | 36 mths | Congo | ICP-MS | [139] |
| urine | 5.2 ± 0.7µg L−1 | 35 | 6–11 y | Mexico | ICP-OES | [140] |
| plasma | range: 0.63–2.26 µg L−1 | 40 (9 m, 11 f) | 5–18 y | Germany | ICP-MS, ICP-AES | [141] |
| urine | range: 0.427–0.761 µg L−1 | 1022 (460 m, 541 f) | 18–80 y | Belgium | ICP-MS | [45] |
| blood | range: 5–12.8 µg L−1 median: 7.6 µg L−1 | 100 | - | Canada | ICP-MS | [117] |
| urine | range: 0.11–1.32 µg L−1 median: 0.31 µg L−1 | 100 | ||||
| plasma | range: 0.63–2.26 µg L−1 median: 1.12 µg L−1 | 100 | ||||
| hair | range: 0.016–0.57 ng mg−1 median: 0.067 ng mg−1 | 45 |
| Neurodevelopmental Disturbances | Biomarkers | Population | Major Conclusion | Ref. |
|---|---|---|---|---|
| IQ Decrement | hair, blood | infants and toddlers | higher hair Mn concentrations are associated with lower IQ scores; blood biomarkers give inconsistent findings | [299] |
| Cognitive Functions | blood, hair, air | perinatal and childhood exposure (38 children from USA (near the Ohio River) | Mn exposure has negative impacts on cognition and behavior | [300] |
| umbilical cord serum | 933 mother–newborn pairs in Shanghai, China | above 5 μg L−1 in three-day-olds showed cognitive deficits in NBNA test | [301] | |
| blood | 6 to 12 months, 448 children born in Mexico City | U-shaped association between Mn level and mental development scores on the BSID | [302] | |
| cord blood | 2-year-old children in Taiwan (230 pairs) | Mn level was associated with decrements in cognitive and language subscales of CDIIT | [303] | |
| Intellectual Ability | blood and hair | children aged 7–9 years, (n = 404) from Marietta and Cambridge, Ohio | both low and high Mn concentrations in blood and hair were negatively associated with the total IQ scores | [23] |
| blood | children 8–11 years, 1089 children living in in South Korea | high Mn is associated with lower scores in thinking, reading, calculations, and LQ in the LDES and a higher commission error in the CPT; low Mn is associated with lower color scores in the Stroop test | [304] | |
| hair | children aged 14–45 months (n = 60) from Pennsylvania, USA | no evidence of an association between Mn concentrations and BSID scores in a region of low-level Mn exposure | [305] | |
| blood, drinking water | prenatal and early childhood (2–3 year old), 524 children from the Sirajdikhan and Pabna Districts of Bangladesh | Mn content in water was associated with fine motor scores in an inverse U-relationship, the adverse effects of Mn was observed in the case of lower Pb level | [306] | |
| umbilical cord blood samples | in utero exposure, 2 years of age (230) from Taiwan | Mn above the 75th percentile had a significant adverse association with the overall, cognitive, and language quotients of the CDIIT | [303] | |
| blood | adolescents aged 11–14 years (n = 299) from USA | Mn was not associated with cognitive and behavioral outcomes at the low exposure levels | [307] | |
| drinking water | from fetal life to school age (5, 10 years), 1265 children in rural Bangladesh | none of the Mn exposures was associated with the children’s cognitive abilities; prenatal Mn exposure was positively associated with cognitive function in girls, boys were unaffected | [308] | |
| Executive Function | blood, hair, drinking water | children aged 6–12 years old (n = 63) from Brasil | blood Mn was associated with visual attention, negatively associated with visual perception and phonological awareness; hair Mn was inversely associated with working memory; Mn from drinking water associated with inhibition of written language and executive functions | [309] |
| hair | 7- and 12-year-old children (n = 70) living near a ferromanganese alloy plant | airborne Mn exposure may be associated with lower IQ and neuropsychological performance in tests of executive function of inhibition responses, strategic visual formation, and verbal working memory in TAVIS-3R, WCST, WISC-III Digit Span subtest and Corsi Block | [109] | |
| Memory | hair and blood | children aged 7–11 years, 174 children living in the Molango State of Mexico | Mn levels showed a negative association with the CAVLT, WRAML scores | [108] |
| Academic Achievement | drinking water | children aged 8–11 years, 840 children living in Araihazar, Bangladesh | no significant relation was observed | [310] |
| Effects on Motor Functions | drinking water | children aged 20–40 months, 524 children, in the Sirajdikhan and Pabna Districts of Bangladesh | Mn < 400 μg/L, Mn is beneficial to fine motor development, whereas at Mn > 400 μg/L, Mn exposure is detrimental for motor function | [306] |
| blood | children ages 7–9 years, 55 children residing in Marietta, Ohio, USA | Mn exposure was significantly associated with poor postural balance | [298] | |
| hair, blood | 7 and 11 years old, 195 children (100 exposed and 95 unexposed) from Mexico | negative association of Mn exposure on motor speed and coordination was shown | [311] | |
| Behavior | blood | children/adolescent | association between early life exposure to Mn and children/adolescent behavior | [312] |
| blood | 5–15 years, 92 children from rom Al Ain Educational Zone, UAE | Mn was significantly associated with ADHD | [313] | |
| prenatal and postnatal enamel regions of deciduous teeth | 84 (aged 9–14 years) Caucasian children | no significant differences in Mn level for children with ASDs compared with TD children, children with ASDs have slightly lower Mn levels, no significant differences between children with HDB and TD children | [314] | |
| drinking water | 8–11 years (201 children) in Bangladesh | positive dose–response relationship between Mn level and CBCL-TRF total scores | [315] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Baj, J.; Flieger, W.; Barbachowska, A.; Kowalska, B.; Flieger, M.; Forma, A.; Teresiński, G.; Portincasa, P.; Buszewicz, G.; Radzikowska-Büchner, E.; et al. Consequences of Disturbing Manganese Homeostasis. Int. J. Mol. Sci. 2023, 24, 14959. https://doi.org/10.3390/ijms241914959
Baj J, Flieger W, Barbachowska A, Kowalska B, Flieger M, Forma A, Teresiński G, Portincasa P, Buszewicz G, Radzikowska-Büchner E, et al. Consequences of Disturbing Manganese Homeostasis. International Journal of Molecular Sciences. 2023; 24(19):14959. https://doi.org/10.3390/ijms241914959
Chicago/Turabian StyleBaj, Jacek, Wojciech Flieger, Aleksandra Barbachowska, Beata Kowalska, Michał Flieger, Alicja Forma, Grzegorz Teresiński, Piero Portincasa, Grzegorz Buszewicz, Elżbieta Radzikowska-Büchner, and et al. 2023. "Consequences of Disturbing Manganese Homeostasis" International Journal of Molecular Sciences 24, no. 19: 14959. https://doi.org/10.3390/ijms241914959