
The Late Embryogenesis Abundant Proteins in Soybean: Identification, Expression Analysis, and the Roles of GmLEA4_19 in Drought Stress

 ,
,  and
and 
Abstract
:1. Introduction
2. Results
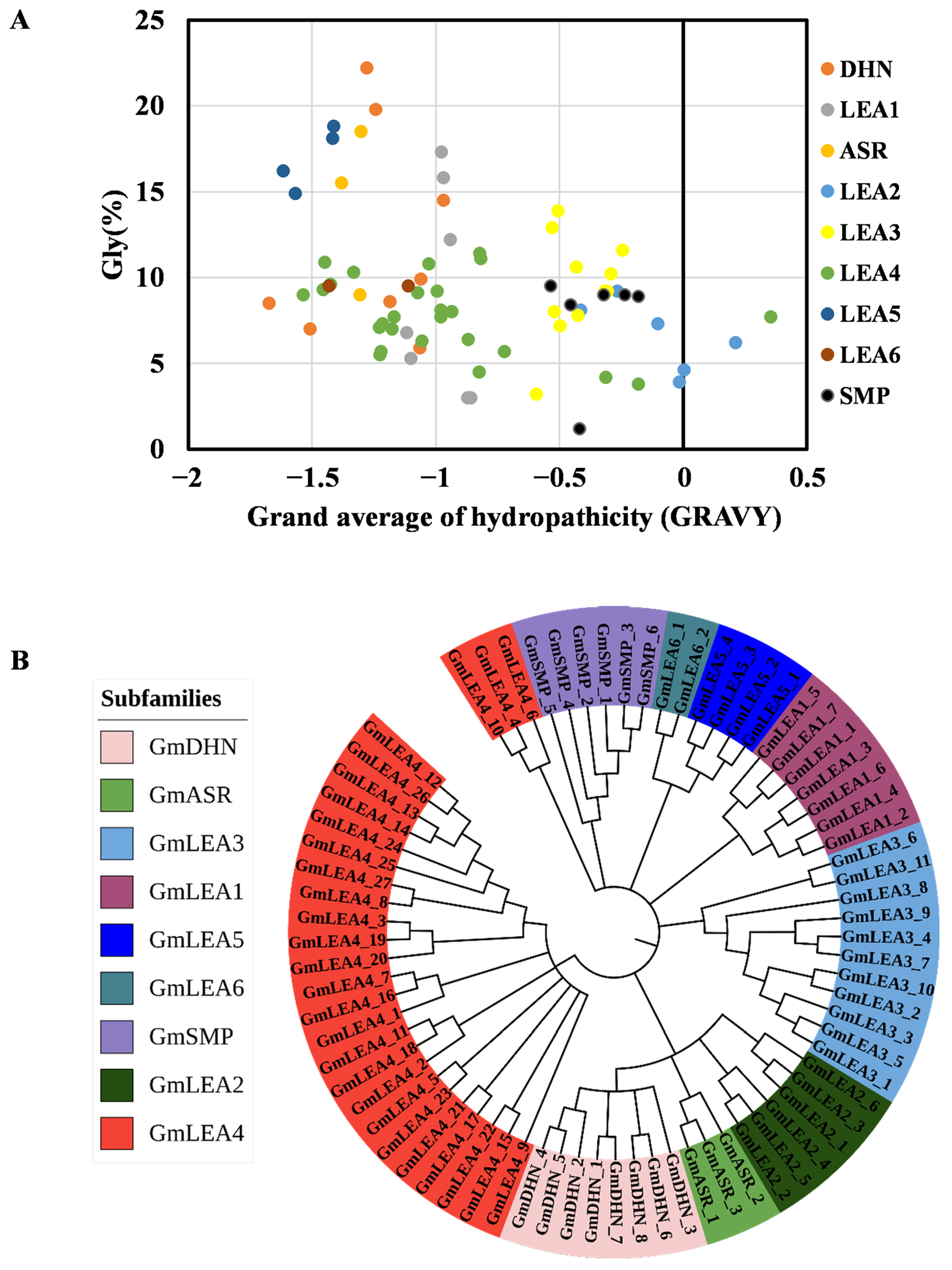
2.1. Identification and Characterization of the LEA Genes in Glycine Max
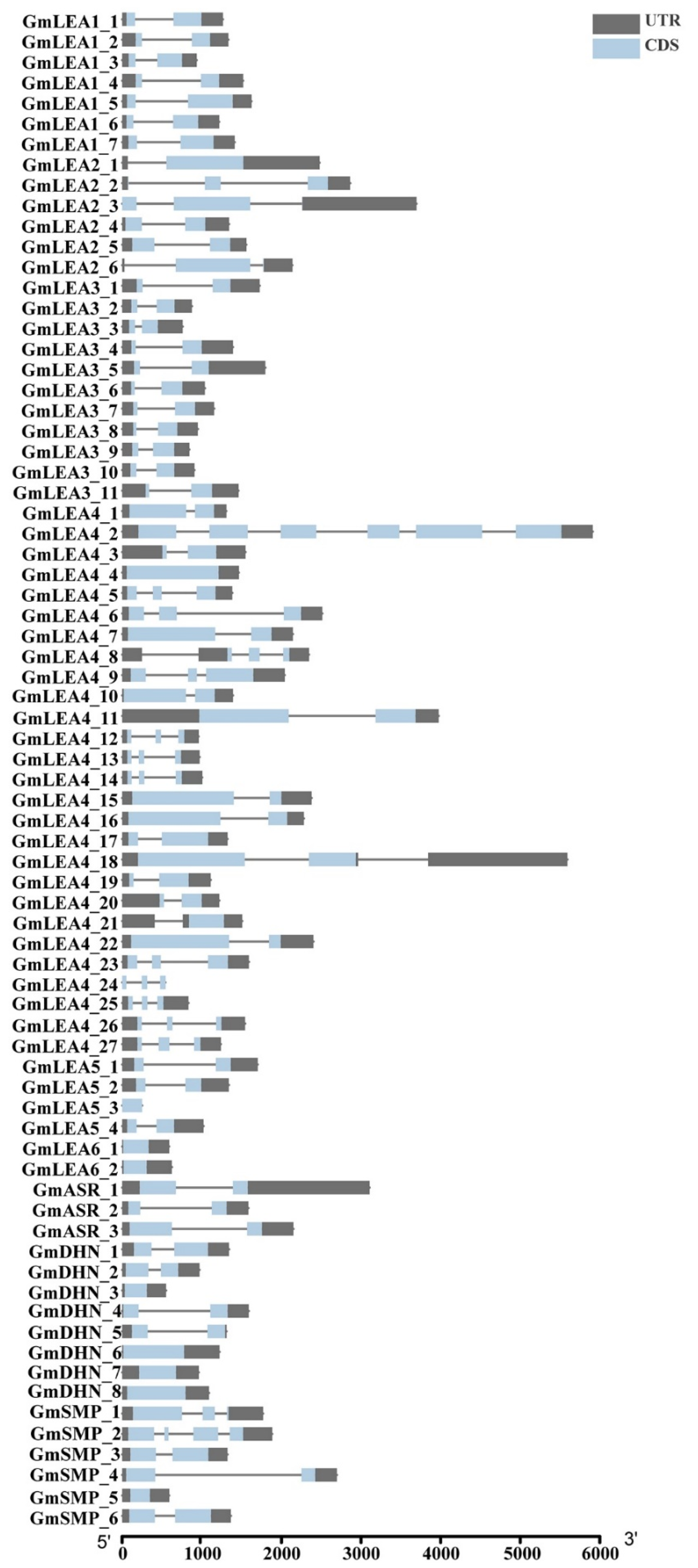
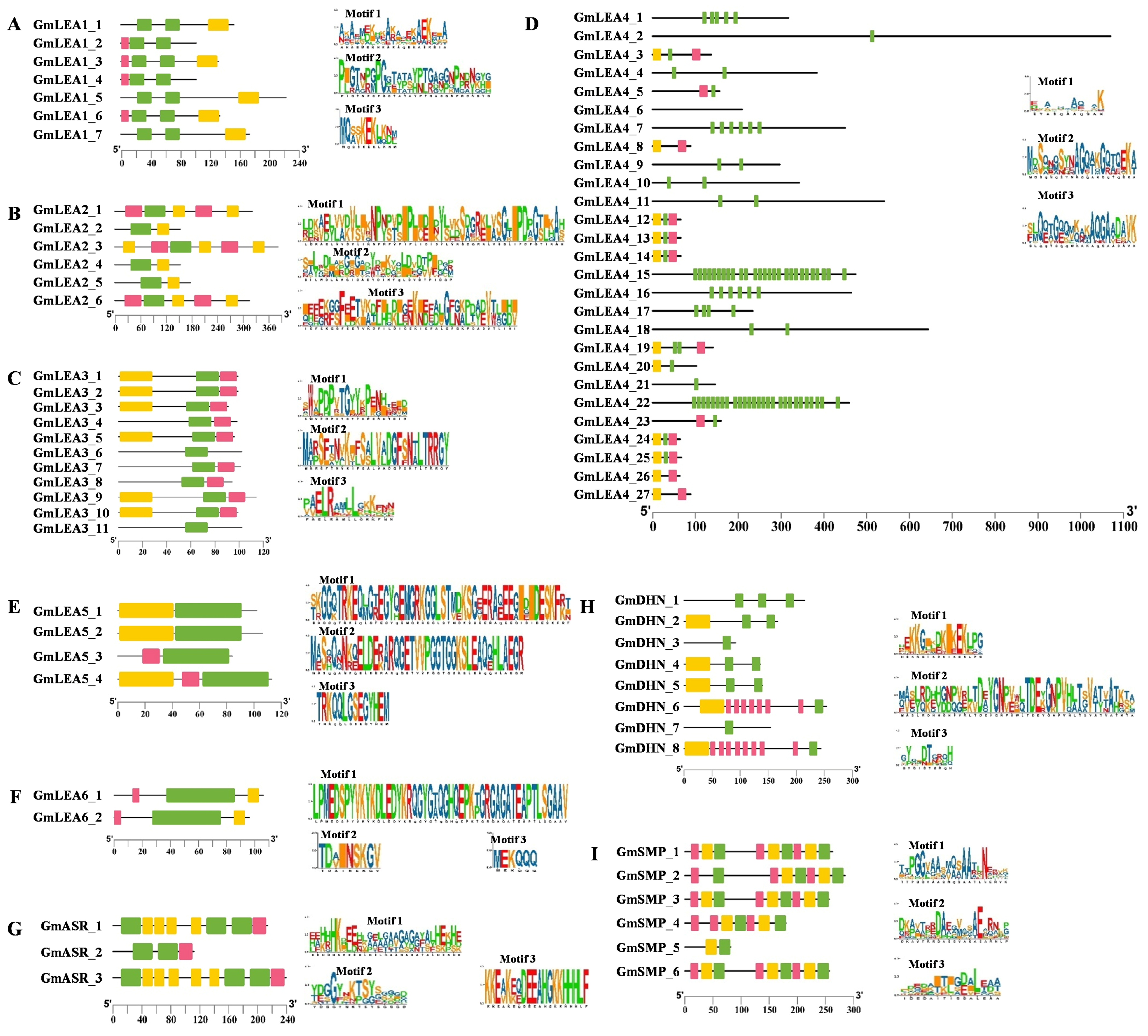
2.2. Phylogenetic Tree, Gene Structure, and Conserved Motifs Analysis of GmLEA Genes
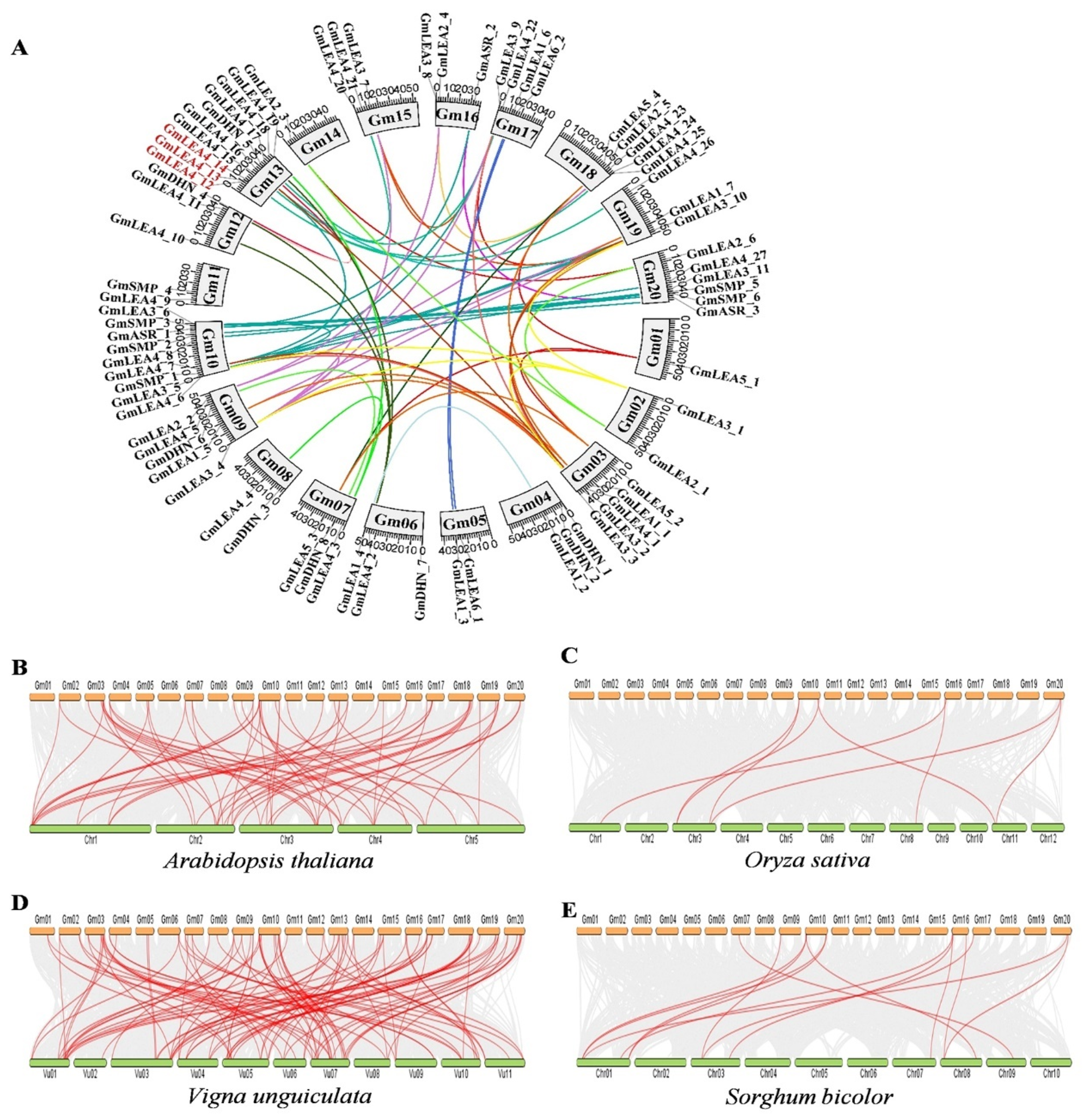
2.3. Chromosomal Distribution, Collinearity, and Ka/Ks Values of GmLEA Family Members
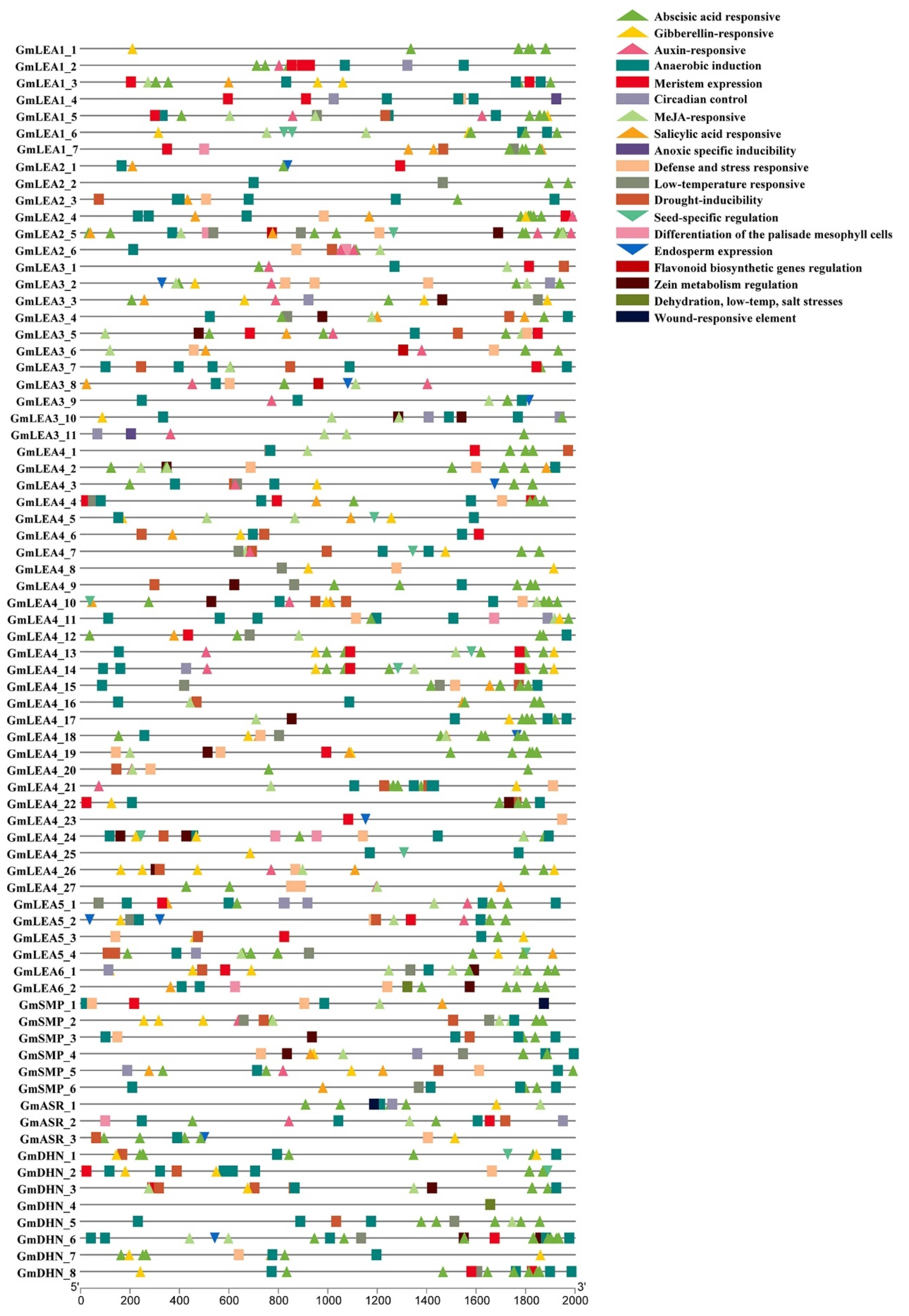
2.4. Cis-Elements Analysis in Promoters of GmLEA Genes
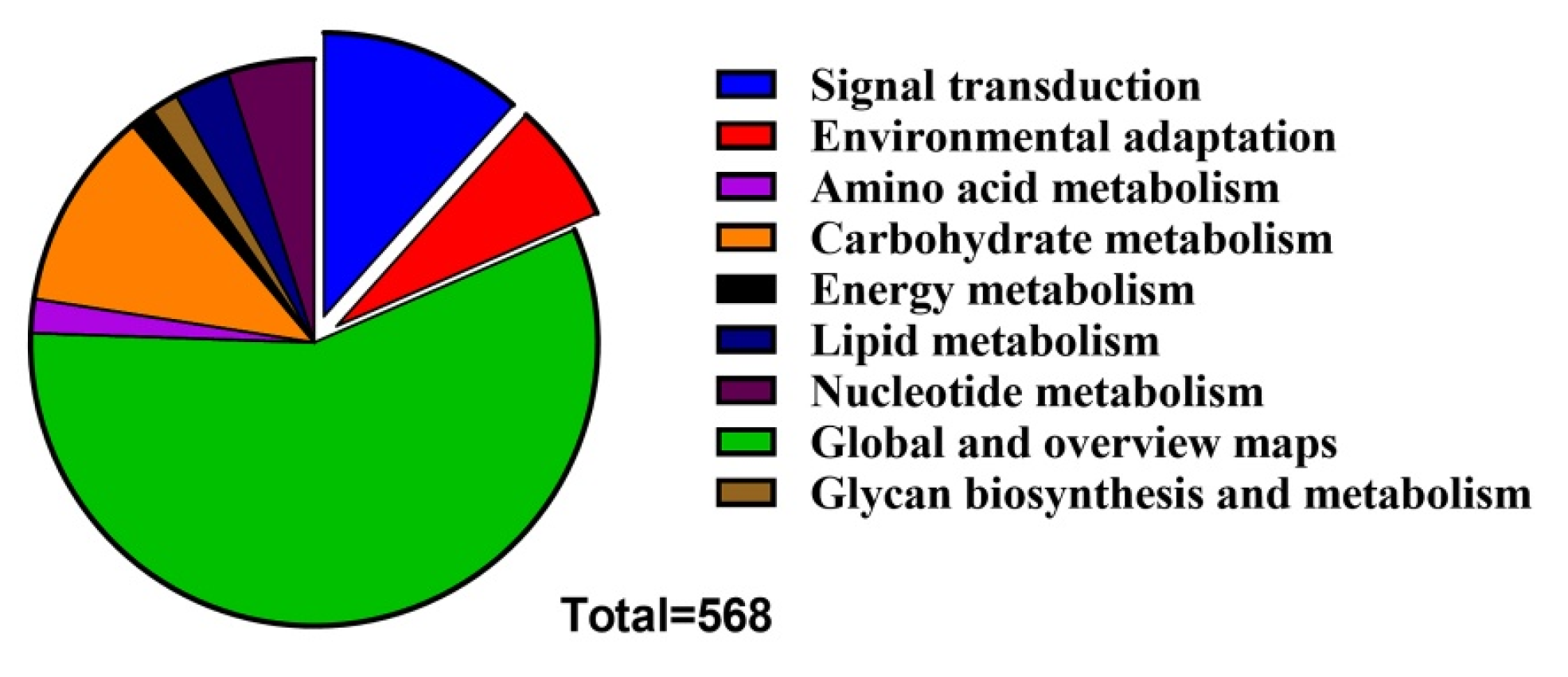
2.5. Prediction of Regulatory Factors and miRNA Targets on GmLEA Transcripts
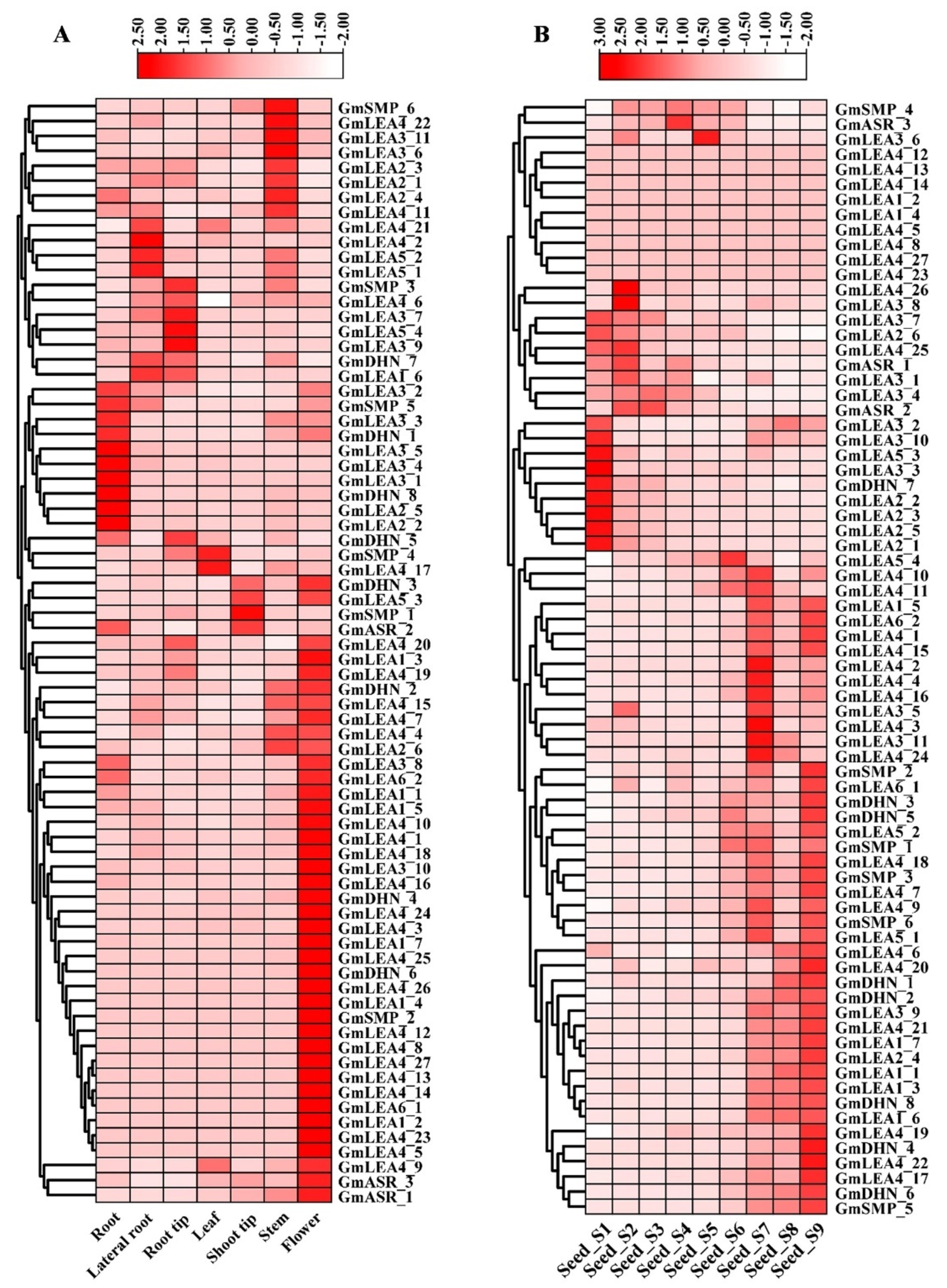
2.6. Expression Profiles Analysis of GmLEA Genes across Tissues
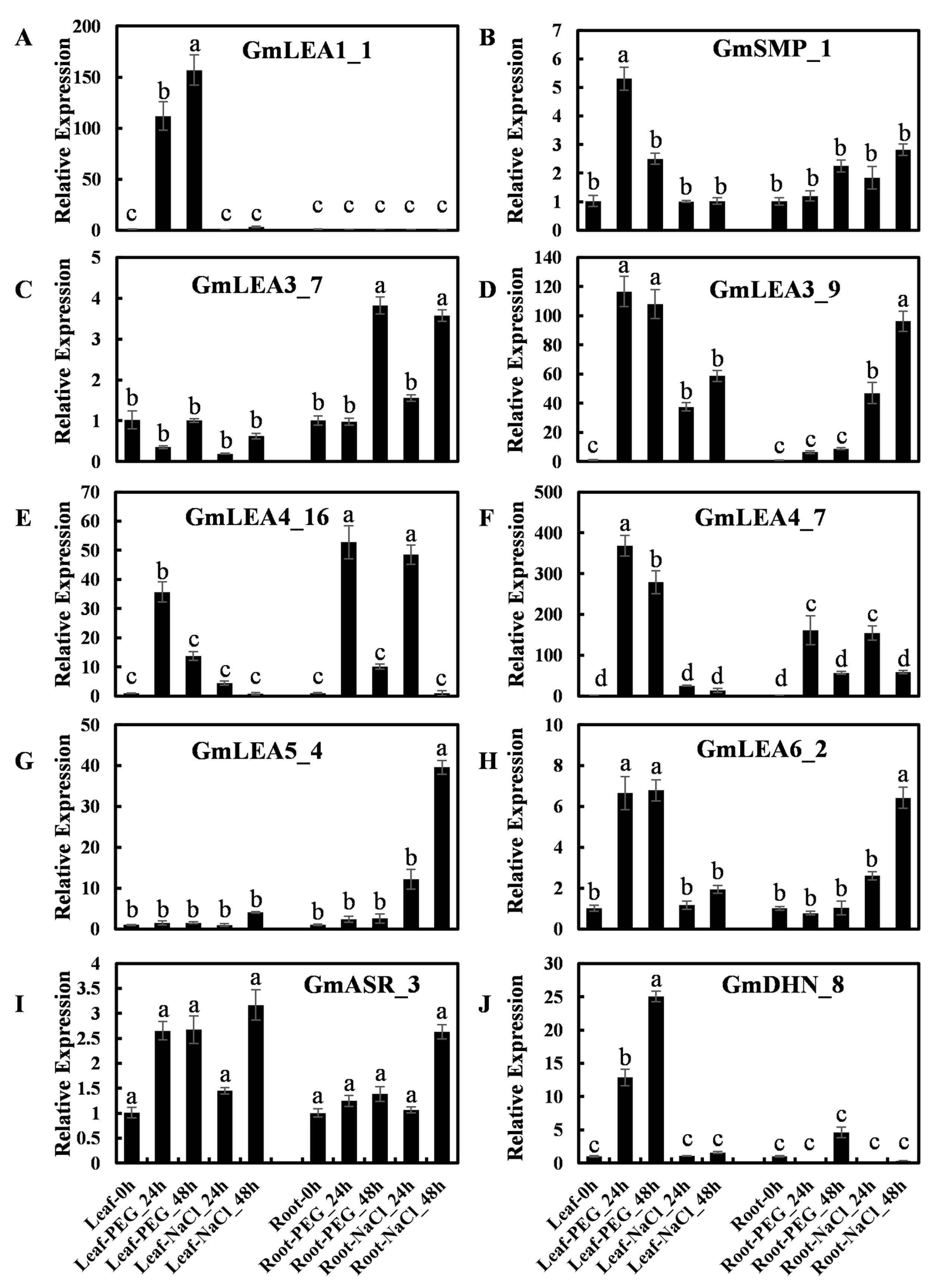
2.7. Expression of GmLEA Genes in Response to Abiotic Stress
2.8. Co-Expression-Based Gene Network Analysis of GmLEA Genes
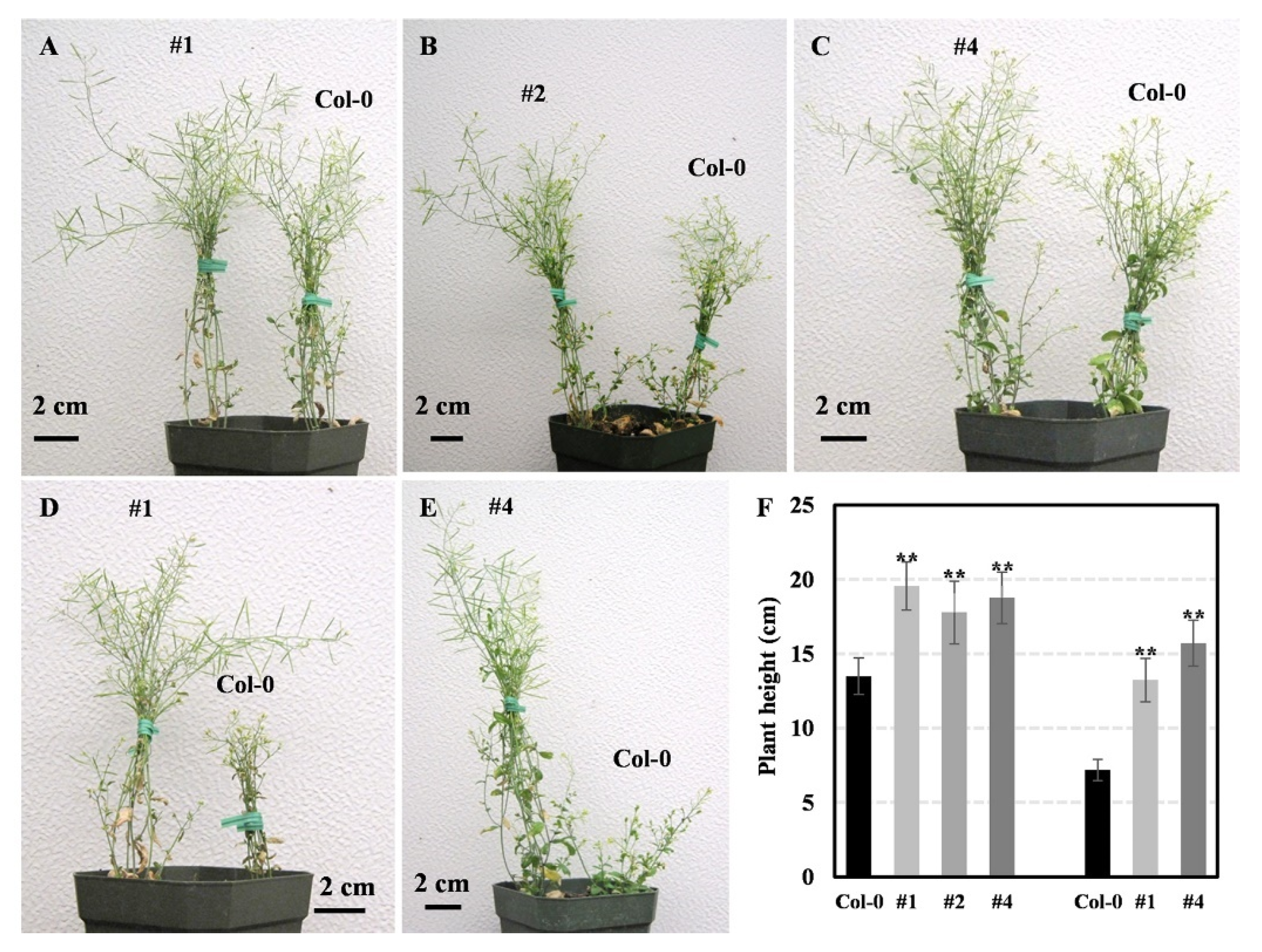
2.9. Overexpression of GnLEA4_19 Improved the Drought Tolerance in Arabidopsis and Soybean
3. Discussion
4. Materials and Methods
4.1. Identification of GmLEA Genes in Glycine Max
4.2. Analysis of GmLEA Protein Properties
4.3. Phylogenetic and Conserved Motifs Analysis of GmLEA Proteins
4.4. Chromosomal Location, Gene Structure, and Gene Duplication of GmLEA Genes
4.5. Regulatory Networks Analysis
4.6. Tissue Expression Pattern Analysis Based on RNA Sequencing Data
4.7. Agrobacterium-Mediated Soybean (Glycine Max) and Arabidopsis Transformation
4.8. Plant Materials Growth Conditions and Treatments
4.9. RNA Isolation, cDNA Synthesis, and qRT-PCR
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Shao, H.B.; Liang, Z.S.; Shao, M.A. LEA proteins in higher plants: Structure, function, gene expression and regulation. Colloid Surf. B Biointerfaces 2005, 45, 131–135. [Google Scholar] [CrossRef]
- Graether, S.P. Proteins Involved in Plant Dehydration Protection: The Late Embryogenesis Abundant Family. Biomolecules 2022, 12, 1380. [Google Scholar] [CrossRef] [PubMed]
- Jia, J.S.; Ge, N.; Wang, Q.Y.; Zhao, L.T.; Chen, C.; Chen, J.W. Genome-wide identification and characterization of members of the LEA gene family in Panax notoginseng and their transcriptional responses to dehydration of recalcitrant seeds. BMC Genom. 2023, 24, 126. [Google Scholar] [CrossRef] [PubMed]
- Bies-Ethève, N.; Gaubier-Comella, P.; Debures, A.; Lasserre, E.; Jobet, E.; Raynal, M.; Cooke, R.; Delseny, M. Inventory, evolution and expression profiling diversity of the LEA (late embryogenesis abundant) protein gene family in Arabidopsis thaliana. Plant Mol. Biol. 2008, 67, 107–124. [Google Scholar] [CrossRef]
- Hundertmark, M.; Hincha, D.K. LEA (late embryogenesis abundant) proteins and their encoding genes in Arabidopsis thaliana. BMC Genom. 2008, 9, 118. [Google Scholar] [CrossRef]
- Wang, X.S.; Zhu, H.B.; Jin, G.L.; Liu, H.L.; Wu, W.R.; Zhu, J. Genome-scale identification and analysis of LEA genes in rice (Oryza sativa L.). Plant Sci. 2007, 172, 414–420. [Google Scholar] [CrossRef]
- Li, X.; Cao, J. Late Embryogenesis Abundant (LEA) Gene Family in Maize: Identification, Evolution, and Expression Profiles. Plant Mol. Biol. Rep. 2016, 34, 15–28. [Google Scholar] [CrossRef]
- Liang, Y.; Xiong, Z.; Zheng, J.; Xu, D.; Zhu, Z.; Xiang, J.; Gan, J.; Raboanatahiry, N.; Yin, Y.; Li, M. Genome-wide identification, structural analysis and new insights into late embryogenesis abundant (LEA) gene family formation pattern in Brassica napus. Sci. Rep. 2016, 6, 24265. [Google Scholar] [CrossRef]
- Magwanga, R.O.; Lu, P.; Kirungu, J.N.; Lu, H.; Wang, X.; Cai, X.; Zhou, Z.; Zhang, Z.; Salih, H.; Wang, K.; et al. Characterization of the late embryogenesis abundant (LEA) proteins family and their role in drought stress tolerance in upland cotton. BMC Genet. 2018, 19, 6. [Google Scholar] [CrossRef]
- Nagaraju, M.; Kumar, S.A.; Reddy, P.S.; Kumar, A.; Rao, D.M.; Kavi Kishor, P.B. Genome-scale identification, classification, and tissue specific expression analysis of late embryogenesis abundant (LEA) genes under abiotic stress conditions in Sorghum bicolor L. PLoS ONE 2019, 14, e0209980. [Google Scholar] [CrossRef]
- Zan, T.; Li, L.; Li, J.; Zhang, L.; Li, X. Genome-wide identification and characterization of late embryogenesis abundant protein-encoding gene family in wheat: Evolution and expression profiles during development and stress. Gene 2020, 736, 144422. [Google Scholar] [CrossRef]
- Zhao, P.; Liu, F.; Ma, M.; Gong, J.; Wang, Q.; Jia, P.; Zheng, G.; Liu, H. Overexpression of AtLEA3-3 confers resistance to cold stress in Escherichia coli and provides enhanced osmotic stress tolerance and ABA sensitivity in Arabidopsis thaliana. Mol. Biol. 2011, 45, 851–862. [Google Scholar] [CrossRef]
- Xiao, B.; Huang, Y.; Tang, N.; Xiong, L. Over-expression of a LEA gene in rice improves drought resistance under the field conditions. TAG. Theor. Appl. Genet. 2007, 115, 35–46. [Google Scholar] [CrossRef] [PubMed]
- Duan, J.; Cai, W. OsLEA3-2, an abiotic stress induced gene of rice plays a key role in salt and drought tolerance. PLoS ONE 2012, 7, e45117. [Google Scholar] [CrossRef]
- Yu, Z.; Wang, X.; Tian, Y.; Zhang, D.; Zhang, L. The functional analysis of a wheat group 3 late embryogenesis abundant protein in Escherichia coli and Arabidopsis under abiotic stresses. Plant Signal. Behav. 2019, 14, 1667207. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Zhao, S.; Zhao, B.; Li, C. Overexpression of TaLEA3 induces rapid stomatal closure under drought stress in Phellodendron amurense Rupr. Plant Sci. 2018, 277, 100–109. [Google Scholar] [CrossRef] [PubMed]
- Luo, D.; Hou, X.; Zhang, Y.; Meng, Y.; Zhang, H.; Liu, S.; Wang, X.; Chen, R. CaDHN5, a Dehydrin Gene from Pepper, Plays an Important Role in Salt and Osmotic Stress Responses. Int. J. Mol. Sci. 2019, 20, 1989. [Google Scholar] [CrossRef]
- Chen, N.; Fan, X.; Wang, C.; Jiao, P.; Jiang, Z.; Ma, Y.; Guan, S.; Liu, S. Overexpression of ZmDHN15 Enhances Cold Tolerance in Yeast and Arabidopsis. Int. J. Mol. Sci. 2022, 24, 480. [Google Scholar] [CrossRef]
- Jia, H.; Wang, X.; Shi, Y.; Wu, X.; Wang, Y.; Liu, J.; Fang, Z.; Li, C.; Dong, K. Overexpression of Medicago sativa LEA4-4 can improve the salt, drought, and oxidation resistance of transgenic Arabidopsis. PLoS ONE 2020, 15, e0234085. [Google Scholar] [CrossRef]
- Lv, A.; Su, L.; Wen, W.; Fan, N.; Zhou, P.; An, Y. Analysis of the Function of the Alfalfa Mslea-D34 Gene in Abiotic Stress Responses and Flowering Time. Plant Cell Physiol. 2021, 62, 28–42. [Google Scholar] [CrossRef]
- Shiraku, M.L.; Magwanga, R.O.; Zhang, Y.; Hou, Y.; Kirungu, J.N.; Mehari, T.G.; Xu, Y.; Wang, Y.; Wang, K.; Cai, X.; et al. Late embryogenesis abundant gene LEA3 (Gh_A08G0694) enhances drought and salt stress tolerance in cotton. Int. J. Biol. Macromol. 2022, 207, 700–714. [Google Scholar] [CrossRef] [PubMed]
- Manavalan, L.P.; Guttikonda, S.K.; Tran, L.S.; Nguyen, H.T. Physiological and molecular approaches to improve drought resistance in soybean. Plant Cell Physiol. 2009, 50, 1260–1276. [Google Scholar] [CrossRef] [PubMed]
- Liu, G.; Xu, H.; Zhang, L.; Zheng, Y. Fe binding properties of two soybean (Glycine max L.) LEA4 proteins associated with antioxidant activity. Plant Cell Physiol. 2011, 52, 994–1002. [Google Scholar] [CrossRef] [PubMed]
- Li, R.H.; Liu, G.B.; Wang, H.; Zheng, Y.Z. Effects of Fe3+ and Zn2+ on the structural and thermodynamic properties of a soybean ASR protein. Biosci. Biotechnol. Biochem. 2013, 77, 475–481. [Google Scholar] [CrossRef]
- Wang, Z.; Yang, Q.; Shao, Y.; Zhang, B.; Feng, A.; Meng, F.; Li, W. GmLEA2-1, a late embryogenesis abundant protein gene isolated from soybean (Glycine max (L.) Merr.), confers tolerance to abiotic stress. Acta Biol. Hung. 2018, 69, 270–282. [Google Scholar] [CrossRef]
- Soulages, J.L.; Kim, K.; Arrese, E.L.; Walters, C.; Cushman, J.C. Conformation of a group 2 late embryogenesis abundant protein from soybean. Evidence of poly (L-proline)-type II structure. Plant Physiol. 2003, 131, 963–975. [Google Scholar] [CrossRef]
- Nekrutenko, A.; Makova, K.D.; Li, W.H. The K(A)/K(S) ratio test for assessing the protein-coding potential of genomic regions: An empirical and simulation study. Genome Res. 2002, 12, 198–202. [Google Scholar] [CrossRef]
- Lynch, M.; Conery, J.S. The Evolutionary Fate and Consequences of Duplicate Genes. Science 2000, 290, 1151–1155. [Google Scholar] [CrossRef]
- Xu, G.; Guo, C.; Shan, H.; Kong, H. Divergence of duplicate genes in exon-intron structure. Proc. Natl. Acad. Sci. USA 2012, 109, 1187–1192. [Google Scholar] [CrossRef]
- Holub, E.B. The arms race is ancient history in Arabidopsis, the wildflower. Nat. Rev. Genet. 2001, 2, 516–527. [Google Scholar] [CrossRef]
- Fuxreiter, M.; Simon, I.; Friedrich, P.; Tompa, P. Preformed structural elements feature in partner recognition by intrinsically unstructured proteins. J. Mol. Biol. 2004, 338, 1015–1026. [Google Scholar] [CrossRef] [PubMed]
- Patil, A.; Nakamura, H. Disordered domains and high surface charge confer hubs with the ability to interact with multiple proteins in interaction networks. FEBS Lett. 2006, 580, 2041–2045. [Google Scholar] [CrossRef] [PubMed]
- Abdul Aziz, M.; Sabeem, M.; Mullath, S.K.; Brini, F.; Masmoudi, K. Plant Group II LEA Proteins: Intrinsically Disordered Structure for Multiple Functions in Response to Environmental Stresses. Biomolecules 2021, 11, 1662. [Google Scholar] [CrossRef] [PubMed]
- Ali-Benali, M.A.; Alary, R.; Joudrier, P.; Gautier, M.F. Comparative expression of five Lea Genes during wheat seed development and in response to abiotic stresses by real-time quantitative RT-PCR. Biochim. Biophys. Acta 2005, 1730, 56–65. [Google Scholar] [CrossRef]
- Tolleter, D.; Jaquinod, M.; Mangavel, C.; Passirani, C.; Saulnier, P.; Manon, S.; Teyssier, E.; Payet, N.; Avelange-Macherel, M.H.; Macherel, D. Structure and function of a mitochondrial late embryogenesis abundant protein are revealed by desiccation. Plant Cell 2007, 19, 1580–1589. [Google Scholar] [CrossRef]
- Battaglia, M.; Olvera-Carrillo, Y.; Garciarrubio, A.; Campos, F.; Covarrubias, A.A. The enigmatic LEA proteins and other hydrophilins. Plant Physiol. 2008, 148, 6–24. [Google Scholar] [CrossRef]
- Wang, Z.; Zhang, Q.; Qin, J.; Xiao, G.; Zhu, S.; Hu, T. OsLEA1a overexpression enhances tolerance to diverse abiotic stresses by inhibiting cell membrane damage and enhancing ROS scavenging capacity in transgenic rice. Funct. Plant Biol. 2021, 48, 860–870. [Google Scholar] [CrossRef]
- Lim, J.; Lim, C.W.; Lee, S.C. The Pepper Late Embryogenesis Abundant Protein, CaDIL1, Positively Regulates Drought Tolerance and ABA Signaling. Front. Plant Sci. 2018, 9, 1301. [Google Scholar] [CrossRef]
- Battaglia, M.; Covarrubias, A.A. Late Embryogenesis Abundant (LEA) proteins in legumes. Front. Plant Sci. 2013, 4, 190. [Google Scholar] [CrossRef]
- Meinke, D.W.; Chen, J.; Beachy, R.N. Expression of storage-protein genes during soybean seed development. Planta 1981, 153, 130–139. [Google Scholar] [CrossRef]
- Jones, S.I.; Vodkin, L.O. Using RNA-Seq to profile soybean seed development from fertilization to maturity. PLoS ONE 2013, 8, e59270. [Google Scholar] [CrossRef] [PubMed]
- Hernández-Sánchez, I.E.; Maruri-López, I.; Martinez-Martinez, C.; Janis, B.; Jiménez-Bremont, J.F.; Covarrubias, A.A.; Menze, M.A.; Graether, S.P.; Thalhammer, A. LEAfing through literature: Late embryogenesis abundant proteins coming of age-achievements and perspectives. J. Exp. Bot. 2022, 73, 6525–6546. [Google Scholar] [CrossRef] [PubMed]
- Koubaa, S.; Brini, F. Functional analysis of a wheat group 3 late embryogenesis abundant protein (TdLEA3) in Arabidopsis thaliana under abiotic and biotic stresses. Plant Physiol. Biochem. 2020, 156, 396–406. [Google Scholar] [CrossRef] [PubMed]
- Guo, X.; Zhang, L.; Wang, X.; Zhang, M.; Xi, Y.; Wang, A.; Zhu, J. Overexpression of Saussurea involucrata dehydrin gene SiDHN promotes cold and drought tolerance in transgenic tomato plants. PLoS ONE 2019, 14, e0225090. [Google Scholar] [CrossRef] [PubMed]
- López-Cordova, A.; Ramírez-Medina, H.; Silva-Martinez, G.A.; González-Cruz, L.; Bernardino-Nicanor, A.; Huanca-Mamani, W.; Montero-Tavera, V.; Tovar-Aguilar, A.; Ramírez-Pimentel, J.G.; Durán-Figueroa, N.V.; et al. LEA13 and LEA30 Are Involved in Tolerance to Water Stress and Stomata Density in Arabidopsis thaliana. Plants 2021, 10, 1694. [Google Scholar] [CrossRef] [PubMed]
- Nida, H.; Girma, G.; Mekonen, M.; Tirfessa, A.; Seyoum, A.; Bejiga, T.; Birhanu, C.; Dessalegn, K.; Senbetay, T.; Ayana, G.; et al. Genome-wide association analysis reveals seed protein loci as determinants of variations in grain mold resistance in sorghum. TAG Theor. Appl. Genet. 2021, 134, 1167–1184. [Google Scholar] [CrossRef]
- Wang, X.; Zhang, M.; Xie, B.; Jiang, X.; Gai, Y. Functional Characteristics Analysis of Dehydrins in Larix kaempferi under Osmotic Stress. Int. J. Mol. Sci. 2021, 22, 1715. [Google Scholar] [CrossRef]
- Karpinska, B.; Razak, N.; Shaw, D.S.; Plumb, W.; Van De Slijke, E.; Stephens, J.; De Jaeger, G.; Murcha, M.W.; Foyer, C.H. Late Embryogenesis Abundant (LEA)5 Regulates Translation in Mitochondria and Chloroplasts to Enhance Growth and Stress Tolerance. Front. Plant Sci. 2022, 13, 875799. [Google Scholar] [CrossRef]
- Yang, Z.; Mu, Y.; Wang, Y.; He, F.; Shi, L.; Fang, Z.; Zhang, J.; Zhang, Q.; Geng, G.; Zhang, S. Characterization of a Novel TtLEA2 Gene from Tritipyrum and Its Transformation in Wheat to Enhance Salt Tolerance. Front. Plant Sci. 2022, 13, 830848. [Google Scholar] [CrossRef]
- Goodstein, D.M.; Shu, S.; Howson, R.; Neupane, R.; Hayes, R.D.; Fazo, J.; Mitros, T.; Dirks, W.; Hellsten, U.; Putnam, N.; et al. Phytozome: A comparative platform for green plant genomics. Nucleic Acids Res. 2012, 40, D1178–D1186. [Google Scholar] [CrossRef]
- Gasteiger, E.; Hoogland, C.; Gattiker, A.; Duvaud, S.E.; Wilkins, M.R.; Appel, R.D.; Bairoch, A. Protein Identification and Analysis Tools on the ExPASy Server; Springer: Berlin/Heidelberg, Germany, 2005. [Google Scholar]
- Hawkins, J.; Bodén, M. Detecting and sorting targeting peptides with neural networks and support vector machines. J. Bioinf. Comput. Biol. 2006, 4, 1–18. [Google Scholar] [CrossRef]
- Chou, K.C.; Shen, H.B. Plant-mPLoc: A top-down strategy to augment the power for predicting plant protein subcellular localization. PLoS ONE 2010, 5, e11335. [Google Scholar] [CrossRef] [PubMed]
- Katoh, K.; Standley, D.M. MAFFT multiple sequence alignment software version 7: Improvements in performance and usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef] [PubMed]
- Letunic, I.; Bork, P. Interactive Tree of Life (iTOL) v5: An online tool for phylogenetic tree display and annotation. Nucleic Acids Res. 2021, 49, W293–W296. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Chen, H.; Zhang, Y.; Thomas, H.R.; Frank, M.H.; He, Y.; Xia, R. TBtools: An Integrative Toolkit Developed for Interactive Analyses of Big Biological Data. Mol. Plant 2020, 13, 1194–1202. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Tang, H.; Debarry, J.D.; Tan, X.; Li, J.; Wang, X.; Lee, T.H.; Jin, H.; Marler, B.; Guo, H.; et al. MCScanX: A toolkit for detection and evolutionary analysis of gene synteny and collinearity. Nucleic Acids Res. 2012, 40, e49. [Google Scholar] [CrossRef]
- Koch, M.A.; Haubold, B.; Mitchell-Olds, T. Comparative evolutionary analysis of chalcone synthase and alcohol dehydrogenase loci in Arabidopsis, Arabis, and related genera (Brassicaceae). Mol. Biol. Evol. 2000, 17, 1483–1498. [Google Scholar] [CrossRef]
- Lescot, M.; Déhais, P.; Thijs, G.; Marchal, K.; Moreau, Y.; Van de Peer, Y.; Rouzé, P.; Rombauts, S. PlantCARE, a database of plant cis-acting regulatory elements and a portal to tools for in silico analysis of promoter sequences. Nucleic Acids Res. 2002, 30, 325–327. [Google Scholar] [CrossRef]
- Tian, F.; Yang, D.C.; Meng, Y.Q.; Jin, J.; Gao, G. PlantRegMap: Charting functional regulatory maps in plants. Nucleic Acids Res. 2020, 48, D1104–D1113. [Google Scholar] [CrossRef]
- Dai, X.; Zhuang, Z.; Zhao, P.X. psRNATarget: A plant small RNA target analysis server (2017 release). Nucleic Acids Res. 2018, 46, W49–W54. [Google Scholar] [CrossRef]
- Libault, M.; Farmer, A.; Brechenmacher, L.; Drnevich, J.; Langley, R.J.; Bilgin, D.D.; Radwan, O.; Neece, D.J.; Clough, S.J.; May, G.D.; et al. Complete transcriptome of the soybean root hair cell, a single-cell model, and its alteration in response to Bradyrhizobium japonicum infection. Plant Physiol. 2010, 152, 541–552. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Xing, A.; Staswick, P.; Clemente, T.E. The use of glufosinate as a selective agent in Agrobacterium-mediated transformation of soybean. Plant Cell Tissue Organ Cult. 1999, 56, 37–46. [Google Scholar] [CrossRef]
- Michel, B.E.; Kaufmann, M.R. The osmotic potential of polyethylene glycol 6000. Plant Physiol. 1973, 51, 914–916. [Google Scholar] [CrossRef] [PubMed]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | Gene ID | PFAM ID | PFAM | Motif # | # of Amino Acids | Molecular Weight | Theoretical pI | Instability Index |
|---|---|---|---|---|---|---|---|---|
| GmLEA1_1 | Glyma.03G144400 | PF03760 | LEA_1 | 1 | 152 | 15,581.23 | 9.66 | 21.04 |
| GmLEA1_2 | Glyma.04G128500 | PF03760 | LEA_1 | 1 | 101 | 11,069.41 | 6.85 | 24.22 |
| GmLEA1_3 | Glyma.05G112000 | PF03760 | LEA_1 | 1 | 131 | 14,628.44 | 9.07 | 48.27 |
| GmLEA1_4 | Glyma.06G310300 | PF03760 | LEA_1 | 1 | 101 | 11,006.42 | 6.92 | 9.7 |
| GmLEA1_5 | Glyma.09G112100 | PF03760 | LEA_1 | 1 | 222 | 23,228.42 | 6.17 | 30.92 |
| GmLEA1_6 | Glyma.17G155000 | PF03760 | LEA_1 | 1 | 133 | 14,680.46 | 9.27 | 49.32 |
| GmLEA1_7 | Glyma.19G147200 | PF03760 | LEA_1 | 1 | 173 | 17,606.29 | 9.58 | 13.11 |
| GmLEA2_1 | Glyma.02G277300 | PF03168 | LEA_2 | 1 | 321 | 35,813.72 | 4.92 | 25.83 |
| GmLEA2_2 | Glyma.09G254302 | PF03168 | LEA_2 | 1 | 152 | 16,551.14 | 4.85 | 17.14 |
| GmLEA2_3 | Glyma.14G037300 | PF03168 | LEA_2 | 1 | 381 | 42,615.88 | 4.75 | 24.75 |
| GmLEA2_4 | Glyma.16G031300 | PF03168 | LEA_2 | 1 | 152 | 16,688.34 | 5.16 | 21.68 |
| GmLEA2_5 | Glyma.18G238700 | PF03168 | LEA_2 | 1 | 176 | 18,933.17 | 5.83 | 17.91 |
| GmLEA2_6 | Glyma.20G044800 | PF03168 | LEA_2 | 1 | 314 | 34,616.47 | 4.79 | 16.24 |
| GmLEA3_1 | Glyma.02G017100 | PF03242 | LEA_3 | 1 | 98 | 10,299.65 | 9.7 | 33.61 |
| GmLEA3_2 | Glyma.03G215000 | PF03242 | LEA_3 | 1 | 98 | 10,513.96 | 9.43 | 51.92 |
| GmLEA3_3 | Glyma.03G253200 | PF03242 | LEA_3 | 1 | 90 | 9770.06 | 9.85 | 38.99 |
| GmLEA3_4 | Glyma.09G043400 | PF03242 | LEA_3 | 1 | 97 | 10,388.76 | 10.08 | 63.7 |
| GmLEA3_5 | Glyma.10G017600 | PF03242 | LEA_3 | 1 | 95 | 10,015.34 | 9.57 | 30.6 |
| GmLEA3_6 | Glyma.10G259200 | PF03242 | LEA_3 | 1 | 101 | 10,767.06 | 9.06 | 34.18 |
| GmLEA3_7 | Glyma.15G149600 | PF03242 | LEA_3 | 1 | 100 | 10,655.93 | 10.41 | 74.69 |
| GmLEA3_8 | Glyma.16G013200 | PF03242 | LEA_3 | 1 | 93 | 10,767.44 | 9.55 | 47.41 |
| GmLEA3_9 | Glyma.17G027400 | PF03242 | LEA_3 | 1 | 113 | 12,282.97 | 10.09 | 56.12 |
| GmLEA3_10 | Glyma.19G211600 | PF03242 | LEA_3 | 1 | 98 | 10,682.2 | 8.93 | 45.61 |
| GmLEA3_11 | Glyma.20G131700 | PF03242 | LEA_3 | 1 | 101 | 10,982.39 | 9.51 | 28.55 |
| GmLEA4_1 | Glyma.03G189200 | PF02987 | LEA_4 | 3 | 316 | 35,342.05 | 5.96 | 32.87 |
| GmLEA4_2 | Glyma.06G283900 | 1069 | 1069 | 8.55 | 42.09 | |||
| GmLEA4_3 | Glyma.07G032400 | LEA_4 | 0.00081 | 136 | 14,836.22 | 8.73 | 29.36 | |
| GmLEA4_4 | Glyma.08G239400 | PF13664 | 383 | 43,024.39 | 8.36 | 38.31 | ||
| GmLEA4_5 | Glyma.09G252700 | 155 | 16,701.6 | 9.65 | 30.8 | |||
| GmLEA4_6 | Glyma.10G014200 | 208 | 23,104.15 | 9.65 | 35.56 | |||
| GmLEA4_7 | Glyma.10G064400 | PF02987 | LEA_4 | 6 | 449 | 48,795.56 | 6.12 | 25.23 |
| GmLEA4_8 | Glyma.10G130600 | 88 | 9250.01 | 5.68 | 31.37 | |||
| GmLEA4_9 | Glyma.11G068900 | 296 | 32,056.4 | 5.63 | 30.6 | |||
| GmLEA4_10 | Glyma.12G001600 | 341 | 38,257.19 | 9.43 | 39.37 | |||
| GmLEA4_11 | Glyma.12G209500 | 540 | 57,273.13 | 5.43 | 32.19 | |||
| GmLEA4_12 | Glyma.13G050000 | 65 | 6712.32 | 6.06 | 42.02 | |||
| GmLEA4_13 | Glyma.13G050051 | 65 | 6668.26 | 8.1 | 26.48 | |||
| GmLEA4_14 | Glyma.13G050100 | 65 | 6682.29 | 8.1 | 30.1 | |||
| GmLEA4_15 | Glyma.13G119400 | 473 | 50,982.23 | 6.65 | 30.84 | |||
| GmLEA4_16 | Glyma.13G149000 | PF02987 | LEA_4 | 7 | 463 | 50,643.82 | 6.33 | 30.29 |
| GmLEA4_17 | Glyma.13G237700 | 233 | 25,630.03 | 5.5 | 32.31 | |||
| GmLEA4_18 | Glyma.13G291800 | 643 | 67,977.14 | 6.18 | 29.76 | |||
| GmLEA4_19 | Glyma.13G363300 | 140 | 15,097.38 | 8.95 | 37.62 | |||
| GmLEA4_20 | Glyma.15G010500 | 101 | 11,100.1 | 6.73 | 22.79 | |||
| GmLEA4_21 | Glyma.15G075700 | 145 | 16,464.96 | 5.03 | 25.52 | |||
| GmLEA4_22 | Glyma.17G040800 | 458 | 49,399.65 | 7.08 | 29.87 | |||
| GmLEA4_23 | Glyma.18G240000 | 159 | 16,983.91 | 9.45 | 17.5 | |||
| GmLEA4_24 | Glyma.18G278700 | 63 | 6593.26 | 9.05 | 24.83 | |||
| GmLEA4_25 | Glyma.18G279300 | 66 | 6808.39 | 7.92 | 28.75 | |||
| GmLEA4_26 | Glyma.19G040000 | 62 | 6450.99 | 4.72 | 40.22 | |||
| GmLEA4_27 | Glyma.20G081400 | 88 | 9258.04 | 6.71 | 29.55 | |||
| GmLEA5_1 | Glyma.01G119600 | PF00477 | LEA_5 | 2 | 101 | 11,141.06 | 6.31 | 42.97 |
| GmLEA5_2 | Glyma.03G056000 | PF00477 | LEA_5 | 2 | 105 | 11,505.35 | 5.53 | 44.21 |
| GmLEA5_3 | Glyma.07G152400 | PF00477 | LEA_5 | 1 | 83 | 9369.31 | 6.59 | 43.53 |
| GmLEA5_4 | Glyma.18G203500 | PF00477 | LEA_5 | 1 | 112 | 12,246.34 | 5.33 | 46.21 |
| GmLEA6_1 | Glyma.05G103100 | PF10714 | LEA_6 | 1 | 105 | 11,440.67 | 9.05 | 52.21 |
| GmLEA6_2 | Glyma.17G164200 | PF10714 | LEA_6 | 1 | 95 | 10,060.89 | 4.91 | 54.59 |
| GmASR_1 | Glyma.10G224300 | PF02496 | ABA_WDS | 1 | 213 | 23,058.3 | 5.7 | 38.31 |
| GmASR_2 | Glyma.16G166600 | PF02496 | ABA_WDS | 1 | 111 | 12,625.9 | 6.35 | 30.1 |
| GmASR_3 | Glyma.20G167500 | PF02496 | ABA_WDS | 1 | 238 | 25,353.7 | 5.58 | 37.27 |
| GmDHN_1 | Glyma.04G009400 | PF00257 | Dehydrin | 1 | 214 | 24,164.6 | 5.53 | 50 |
| GmDHN_2 | Glyma.04G009900 | PF00257 | Dehydrin | 1 | 166 | 17,319.92 | 9.22 | 34.1 |
| GmDHN_3 | Glyma.08G048900 | PF00257 | Dehydrin | 1 | 91 | 9917.88 | 6.64 | 30.22 |
| GmDHN_4 | Glyma.12G235800 | PF00257 | Dehydrin | 1 | 135 | 14,870.27 | 5.54 | 32.59 |
| GmDHN_5 | Glyma.13G201300 | PF00257 | Dehydrin | 1 | 139 | 15,133.43 | 5.52 | 38.07 |
| GmDHN_6 | Glyma.09G185500 | PF00257 | Dehydrin | 1 | 253 | 26,630.01 | 6.29 | 8.95 |
| GmDHN_7 | Glyma.06G009350 | PF00257 | Dehydrin | 1 | 153 | 17,521.21 | 5.56 | 46.52 |
| GmDHN_8 | Glyma.07G090400 | PF00257 | Dehydrin | 1 | 243 | 25,658.97 | 6.02 | 5.45 |
| GmSMP_1 | Glyma.10G027600 | PF04927 | SMP | 3 | 262 | 27,455.59 | 5.16 | 26.15 |
| GmSMP_2 | Glyma.10G159400 | PF04927 | SMP | 3 | 284 | 29,451.95 | 6.48 | 29.05 |
| GmSMP_3 | Glyma.10G247500 | PF04927 | SMP | 3 | 256 | 26,239 | 4.75 | 37.14 |
| GmSMP_4 | Glyma.11G158394 | PF04927 | SMP | 2 | 179 | 18,242.1 | 4.27 | 26.54 |
| GmSMP_5 | Glyma.20G147500 | PF04927 | SMP | 1 | 81 | 8806.94 | 6.13 | 77.43 |
| GmSMP_6 | Glyma.20G147600 | PF04927 | SMP | 3 | 256 | 26,058.06 | 4.9 | 34.86 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Guo, B.; Zhang, J.; Yang, C.; Dong, L.; Ye, H.; Valliyodan, B.; Nguyen, H.T.; Song, L. The Late Embryogenesis Abundant Proteins in Soybean: Identification, Expression Analysis, and the Roles of GmLEA4_19 in Drought Stress. Int. J. Mol. Sci. 2023, 24, 14834. https://doi.org/10.3390/ijms241914834
Guo B, Zhang J, Yang C, Dong L, Ye H, Valliyodan B, Nguyen HT, Song L. The Late Embryogenesis Abundant Proteins in Soybean: Identification, Expression Analysis, and the Roles of GmLEA4_19 in Drought Stress. International Journal of Molecular Sciences. 2023; 24(19):14834. https://doi.org/10.3390/ijms241914834
Chicago/Turabian StyleGuo, Binhui, Jianhua Zhang, Chunhong Yang, Lu Dong, Heng Ye, Babu Valliyodan, Henry T. Nguyen, and Li Song. 2023. "The Late Embryogenesis Abundant Proteins in Soybean: Identification, Expression Analysis, and the Roles of GmLEA4_19 in Drought Stress" International Journal of Molecular Sciences 24, no. 19: 14834. https://doi.org/10.3390/ijms241914834