
Facilitation of Ovarian Response by Mechanical Force—Latest Insight on Fertility Improvement in Women with Poor Ovarian Response or Primary Ovarian Insufficiency

{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
1.1. The Root of Reduced Fertility in Aged Women
1.2. A Myriad of Approaches Have Been Attempted to Prevent or Slow down the Process of Ovarian Aging
2. Reproductive Characteristics of Aging IVF Patients: Ovarian Aging and Poor Ovarian Response
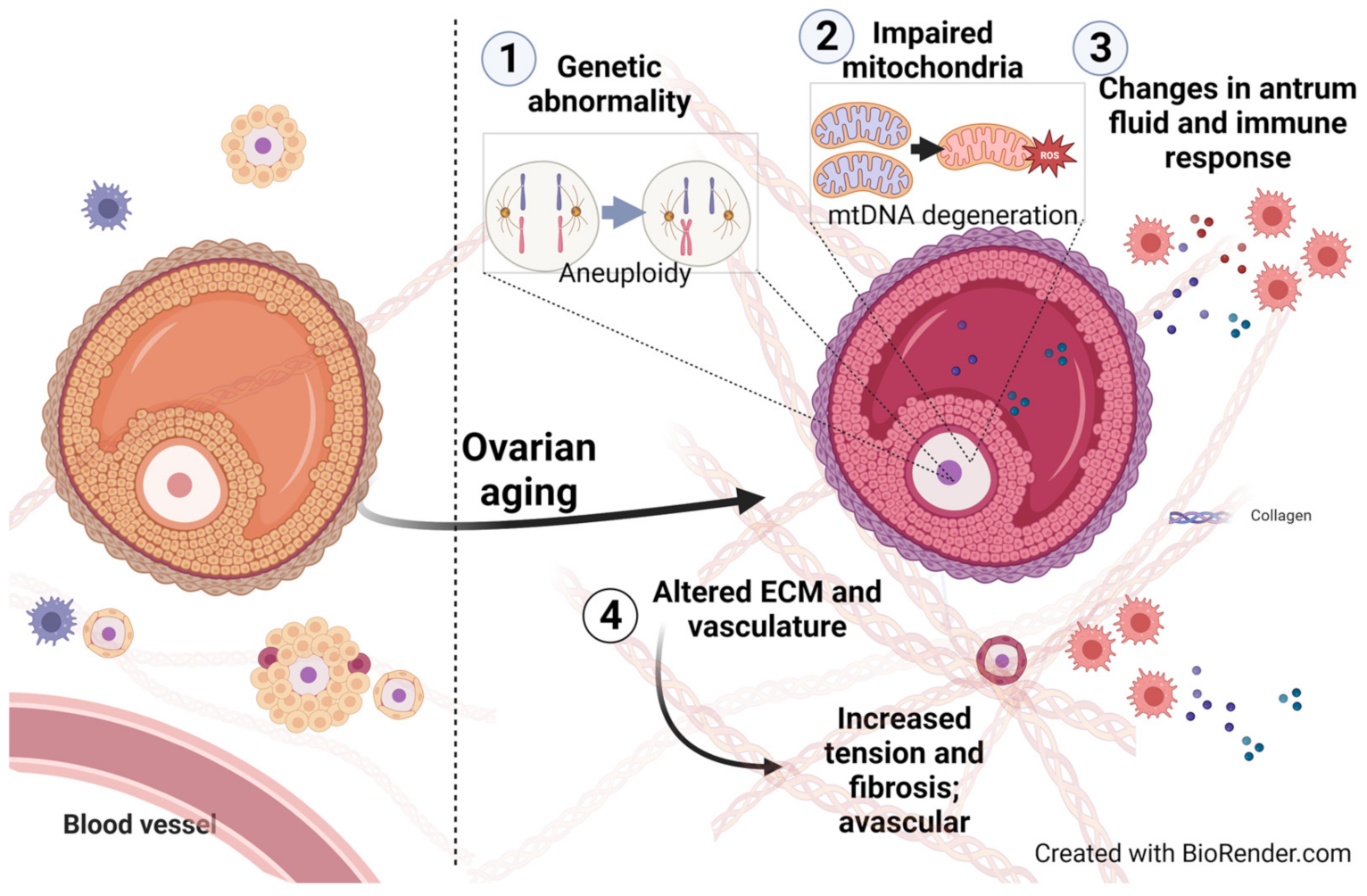
2.1. The Process of Ovarian Decay in Aging Women Is a Multi-Factorial Phenomenon
2.2. Molecular and Functional Changes in Aging Ovaries
2.2.1. Genetic Abnormality
2.2.2. Changes of Mitochondria in the Oocyte and Somatic Cells
2.2.3. Altered Extracellular Matrix (ECM) in the Aging Ovary
2.2.4. Alterations of Vascularization in Aging Ovaries
2.2.5. Shifts in Follicular Fluid Composition
2.2.6. Alterations in the Ovarian Immune Response
3. Potential Treatments to Avert Infertility in Women with POR or POI: Adjunct Intervention, Gamete Engineering, and Strategies That Incorporate Mechanical Manipulation
3.1. Adjunct Intervention
3.2. Gamete Engineering
3.3. Strategies That Incorporate Mechanical Force for Stimulating Follicular Growth in Women with POR or POI
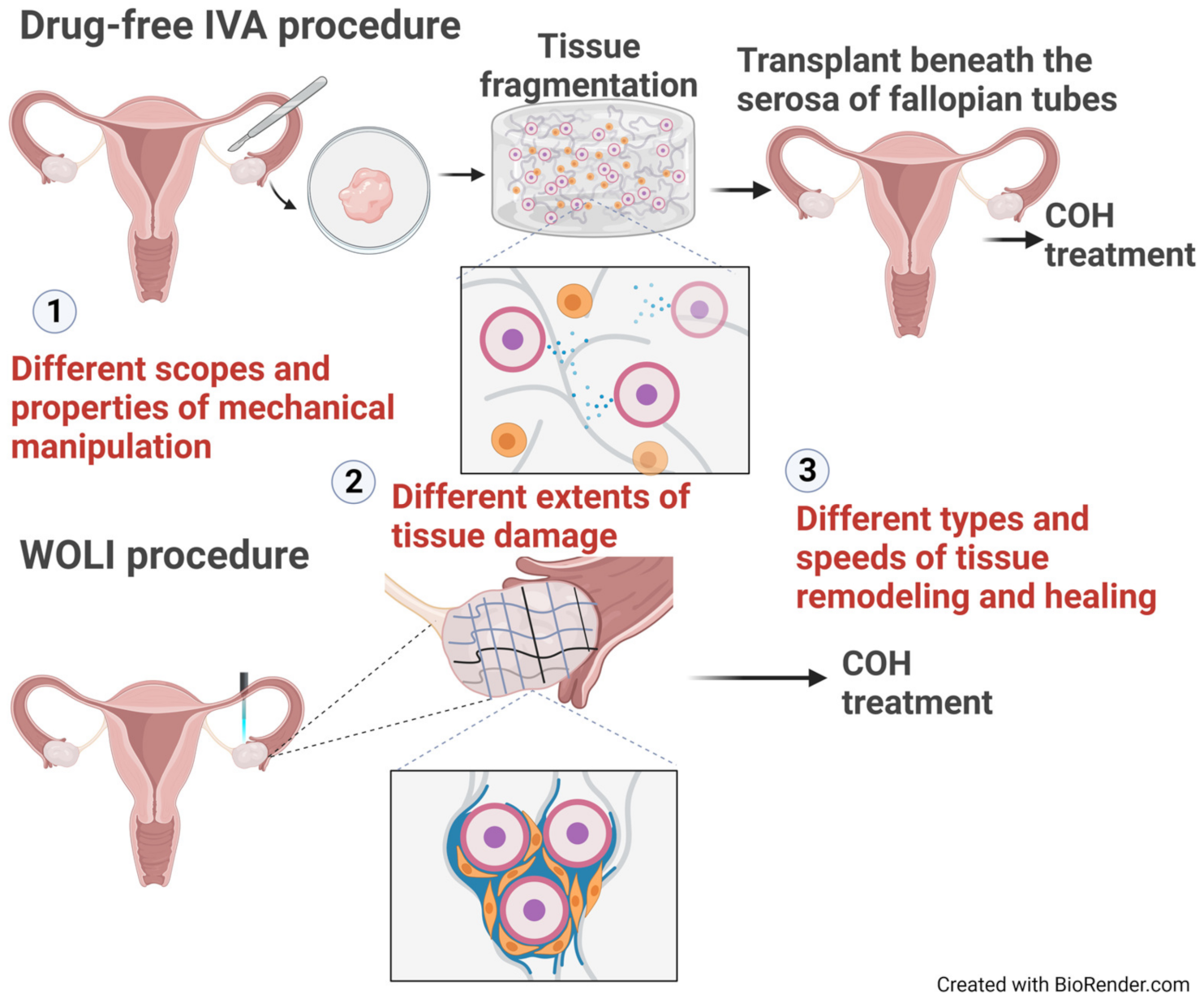
3.3.1. IVA: An Evolving Experimental Technique Involving Pharmacological Stimulation and Mechanical Manipulation
3.3.2. Drug-Free IVA: Mechanical Force Alone Is Sufficient to Activate Dormant Follicles
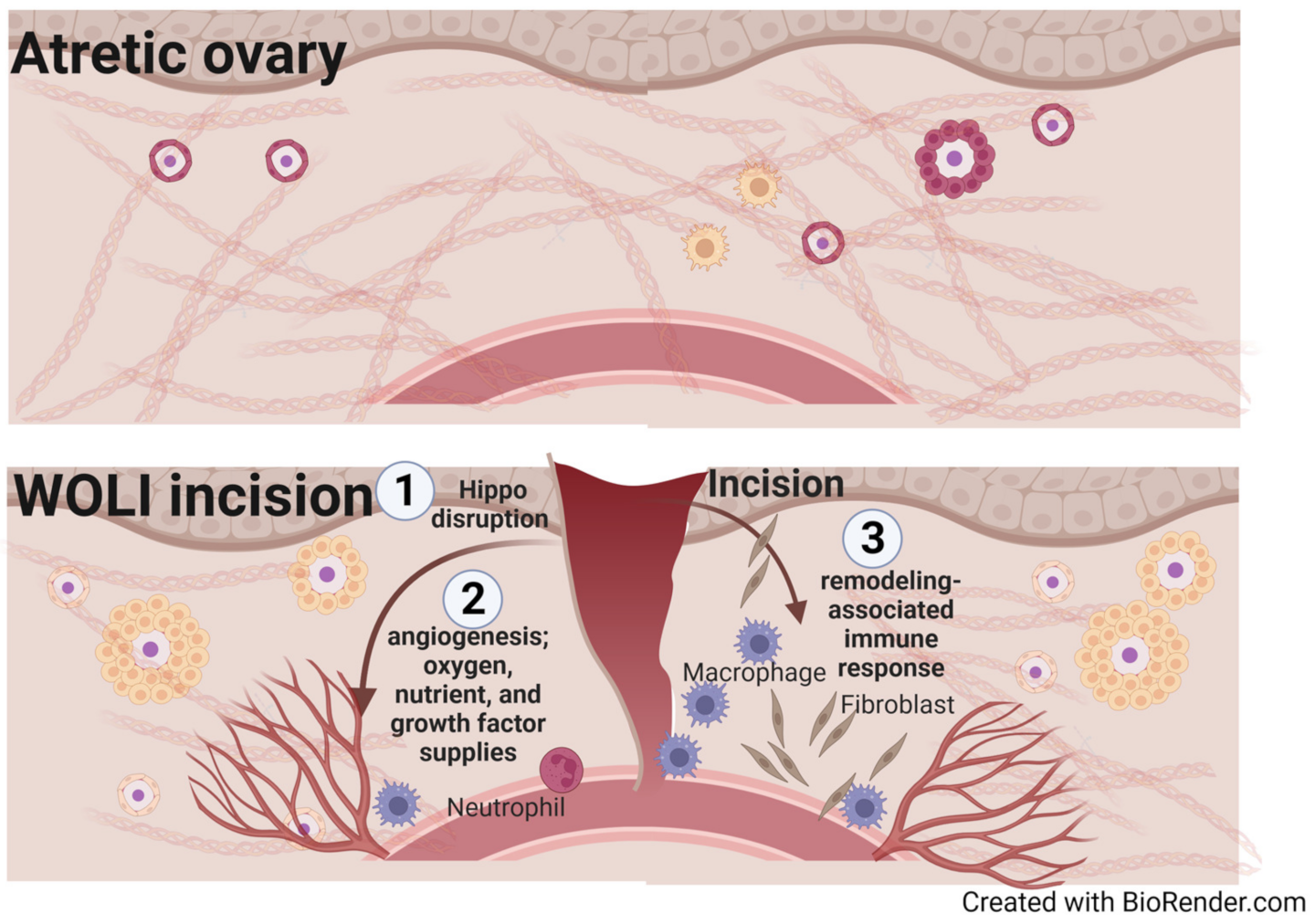
3.3.3. Whole Ovary Laparoscopic Incision (WOLI) Procedure: Rescuing Aging Ovaries via Surgical Intervention
3.3.4. Ovarian Surface Cutting Improves Follicular Development in Aging gcNrg1KO Mice
4. Potential Mechanisms That Underlie Mechanical-Manipulation-Mediated Stimulation of Folliculogenesis
4.1. Changes of Mechanobiological Signaling in the ECM Environment
4.2. Disruption of the Hypoxic Microenvironment and the Fibrotic Signal Barrier
4.3. Stimulation of Angiogenesis and Aseptic Inflammation
5. WOLI vs. Drug-Free IVA: Potential Differences in Their Actions and Physiological Consequences
6. Ways to Improve and Validate the WOLI Procedure
7. Perspectives and Future Directions
Funding
Data Availability Statement
Conflicts of Interest
References
- Choi, S.K.Y.; Venetis, C.; Ledger, W.; Havard, A.; Harris, K.; Norman, R.J.; Jorm, L.R.; Chambers, G.M. Population-wide contribution of medically assisted reproductive technologies to overall births in Australia: Temporal trends and parental characteristics. Hum. Reprod. 2022, 37, 1047–1058. [Google Scholar] [CrossRef] [PubMed]
- Schlegel, P.N. Evaluation of male infertility. Minerva Ginecol. 2009, 61, 261–283. [Google Scholar] [PubMed]
- Reig, A.; Herraiz, S.; Pellicer, A.; Seli, E. Emerging follicular activation strategies to treat women with poor ovarian response and primary ovarian insufficiency. Curr. Opin. Obstet. Gynecol. 2021, 33, 241–248. [Google Scholar] [CrossRef] [PubMed]
- Roest, J.; van Heusden, A.M.; Mous, H.; Zeilmaker, G.H.; Verhoeff, A. The ovarian response as a predictor for successful in vitro fertilization treatment after the age of 40 years. Fertil. Steril. 1996, 66, 969–973. [Google Scholar] [CrossRef] [PubMed]
- Drakopoulos, P.; Bardhi, E.; Boudry, L.; Vaiarelli, A.; Makrigiannakis, A.; Esteves, S.C.; Tournaye, H.; Blockeel, C. Update on the management of poor ovarian response in IVF: The shift from Bologna criteria to the Poseidon concept. Ther. Adv. Reprod. Health 2020, 14, 2633494120941480. [Google Scholar] [CrossRef] [PubMed]
- Kirshenbaum, M.; Orvieto, R. Premature ovarian insufficiency (POI) and autoimmunity-an update appraisal. J. Assist. Reprod. Genet. 2019, 36, 2207–2215. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, N.; Yoshioka, N.; Takae, S.; Sugishita, Y.; Tamura, M.; Hashimoto, S.; Morimoto, Y.; Kawamura, K. Successful fertility preservation following ovarian tissue vitrification in patients with primary ovarian insufficiency. Hum. Reprod. 2015, 30, 608–615. [Google Scholar] [CrossRef] [PubMed]
- Ishizuka, B. Current Understanding of the Etiology, Symptomatology, and Treatment Options in Premature Ovarian Insufficiency (POI). Front. Endocrinol. 2021, 12, 626924. [Google Scholar] [CrossRef]
- Szeliga, A.; Calik-Ksepka, A.; Maciejewska-Jeske, M.; Grymowicz, M.; Smolarczyk, K.; Kostrzak, A.; Smolarczyk, R.; Rudnicka, E.; Meczekalski, B. Autoimmune Diseases in Patients with Premature Ovarian Insufficiency—Our Current State of Knowledge. Int. J. Mol. Sci. 2021, 22, 2594. [Google Scholar] [CrossRef]
- Fiorentino, G.; Cimadomo, D.; Innocenti, F.; Soscia, D.; Vaiarelli, A.; Ubaldi, F.M.; Gennarelli, G.; Garagna, S.; Rienzi, L.; Zuccotti, M. Biomechanical forces and signals operating in the ovary during folliculogenesis and their dysregulation: Implications for fertility. Hum. Reprod. Update 2023, 29, 1–23. [Google Scholar] [CrossRef]
- Qiao, J.; Wang, Z.-B.; Feng, H.-L.; Miao, Y.-L.; Wang, Q.; Yu, Y.; Wei, Y.-C.; Yan, J.; Wang, W.-H.; Shen, W.; et al. The root of reduced fertility in aged women and possible therapentic options: Current status and future perspects. Mol. Asp. Med. 2014, 38, 54–85. [Google Scholar] [CrossRef] [PubMed]
- Ubaldi, F.M.; Cimadomo, D.; Vaiarelli, A.; Fabozzi, G.; Venturella, R.; Maggiulli, R.; Mazzilli, R.; Ferrero, S.; Palagiano, A.; Rienzi, L. Advanced Maternal Age in IVF: Still a Challenge? The Present and the Future of Its Treatment. Front. Endocrinol. 2019, 10, 94. [Google Scholar] [CrossRef] [PubMed]
- Mumusoglu, S.; Turan, V.; Uckan, H.; Suzer, A.; Sokmensuer, L.K.; Bozdag, G. The Impact of a Long-Acting Oral Sphingosine-1-Phosphate Analogue on Ovarian Aging in a Rat Model. Reprod. Sci. 2018, 25, 1330–1335. [Google Scholar] [CrossRef] [PubMed]
- Younis, J.S. Ovarian aging: Latest thoughts on assessment and management. Curr. Opin. Obstet. Gynecol. 2011, 23, 427–434. [Google Scholar] [CrossRef] [PubMed]
- Song, C.; Peng, W.; Yin, S.; Zhao, J.; Fu, B.; Zhang, J.; Mao, T.; Wu, H.; Zhang, Y. Melatonin improves age-induced fertility decline and attenuates ovarian mitochondrial oxidative stress in mice. Sci. Rep. 2016, 6, 35165. [Google Scholar] [CrossRef] [PubMed]
- Tamura, H.; Kawamoto, M.; Sato, S.; Tamura, I.; Maekawa, R.; Taketani, T.; Aasada, H.; Takaki, E.; Nakai, A.; Reiter, R.J.; et al. Long-term melatonin treatment delays ovarian aging. J. Pineal Res. 2017, 62, e12381. [Google Scholar] [CrossRef] [PubMed]
- Liu, M.; Yin, Y.; Ye, X.; Zeng, M.; Zhao, Q.; Keefe, D.L.; Liu, L. Resveratrol protects against age-associated infertility in mice. Hum. Reprod. 2013, 28, 707–717. [Google Scholar] [CrossRef] [PubMed]
- Okamoto, N.; Sato, Y.; Kawagoe, Y.; Shimizu, T.; Kawamura, K. Short-term resveratrol treatment restored the quality of oocytes in aging mice. Aging 2022, 14, 5628–5640. [Google Scholar] [CrossRef]
- Du, D.; Tang, X.; Li, Y.; Gao, Y.; Chen, R.; Chen, Q.; Wen, J.; Wu, T.; Zhang, Y.; Lu, H.; et al. Senotherapy Protects against Cisplatin-Induced Ovarian Injury by Removing Senescent Cells and Alleviating DNA Damage. Oxid. Med. Cell. Longev. 2022, 2022, 9144644. [Google Scholar] [CrossRef]
- Esencan, E.; Beroukhim, G.; Seifer, D.B. Age-related changes in Folliculogenesis and potential modifiers to improve fertility outcomes—A narrative review. Reprod. Biol. Endocrinol. 2022, 20, 156. [Google Scholar] [CrossRef]
- Secomandi, L.; Borghesan, M.; Velarde, M.; Demaria, M. The role of cellular senescence in female reproductive aging and the potential for senotherapeutic interventions. Hum. Reprod. Update 2022, 28, 172–189. [Google Scholar] [CrossRef] [PubMed]
- Hikabe, O.; Hamazaki, N.; Nagamatsu, G.; Obata, Y.; Hirao, Y.; Hamada, N.; Shimamoto, S.; Imamura, T.; Nakashima, K.; Saitou, M.; et al. Reconstitution in vitro of the entire cycle of the mouse female germ line. Nature 2016, 539, 299–303. [Google Scholar] [CrossRef] [PubMed]
- Yoshino, T.; Suzuki, T.; Nagamatsu, G.; Yabukami, H.; Ikegaya, M.; Kishima, M.; Kita, H.; Imamura, T.; Nakashima, K.; Nishinakamura, R.; et al. Generation of ovarian follicles from mouse pluripotent stem cells. Science 2021, 373, eabe0237. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.-J.; Ge, W.; Liu, J.-C.; Klinger, F.G.; Dyce, P.W.; De Felici, M.; Shen, W. Complete in vitro oogenesis: Retrospects and prospects. Cell Death Differ. 2017, 24, 1845–1852. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Ding, B.; Wu, L.; Qiu, J.; Li, Q.; Ye, Z.; Yang, J. Transcriptome Analysis Reveals the Mechanism of Natural Ovarian Ageing. Front. Endocrinol. 2022, 13, 918212. [Google Scholar] [CrossRef] [PubMed]
- Gleicher, N.; Kushnir, V.A.; Albertini, D.F.; Barad, D.H. Improvements in IVF in women of advanced age. J. Endocrinol. 2016, 230, F1–F6. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Kawamura, K.; Cheng, Y.; Liu, S.; Klein, C.; Liu, S.; Duan, E.K.; Hsueh, A.J. Activation of dormant ovarian follicles to generate mature eggs. Proc. Natl. Acad. Sci. USA 2010, 107, 10280–10284. [Google Scholar] [CrossRef] [PubMed]
- Hsueh, A.J.; Kawamura, K. Hippo signaling disruption and ovarian follicle activation in infertile patients. Fertil. Steril. 2020, 114, 458–464. [Google Scholar] [CrossRef]
- Lee, H.N.; Chang, E.M. Primordial follicle activation as new treatment for primary ovarian insufficiency. Clin. Exp. Reprod. Med. 2019, 46, 43–49. [Google Scholar] [CrossRef]
- Terren, C.; Munaut, C. Molecular Basis Associated with the Control of Primordial Follicle Activation During Transplantation of Cryopreserved Ovarian Tissue. Reprod. Sci. 2021, 28, 1257–1266. [Google Scholar] [CrossRef]
- Liu, K.; Rajareddy, S.; Liu, L.; Jagarlamudi, K.; Boman, K.; Selstam, G.; Reddy, P. Control of mammalian oocyte growth and early follicular development by the oocyte PI3 kinase pathway: New roles for an old timer. Dev. Biol. 2006, 299, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Feng, H.; Zhang, Y.; Zhang, J.V.; Wang, X.; Liu, D.; Wang, T.; Li, R.H.W.; Ng, E.H.Y.; Yeung, W.S.B.; et al. Current Understandings of Core Pathways for the Activation of Mammalian Primordial Follicles. Cells 2021, 10, 1491. [Google Scholar] [CrossRef] [PubMed]
- Reddy, P.; Liu, L.; Adhikari, D.; Jagarlamudi, K.; Rajareddy, S.; Shen, Y.; Du, C.; Tang, W.; Ha, T.; Peng, S.L.; et al. Oocyte-specific deletion of Pten causes premature activation of the primordial follicle pool. Science 2008, 319, 611–613. [Google Scholar] [CrossRef] [PubMed]
- Kawamara, K.; Kelsey, T.; Hiraike, O. Editorial: Ovarian Ageing: Pathophysiology and Recent Development of Maintaining Ovarian Reserve. Front. Endocrinol. 2020, 11, 591764. [Google Scholar] [CrossRef] [PubMed]
- Nagamatsu, G.; Shimamoto, S.; Hamazaki, N.; Nishimura, Y.; Hayashi, K. Mechanical stress accompanied with nuclear rotation is involved in the dormant state of mouse oocytes. Sci. Adv. 2019, 5, eaav9960. [Google Scholar] [CrossRef] [PubMed]
- Chang, C.L.; Chin, T.H.; Hsu, Y.C.; Hsueh, A.J. Whole-Ovary Laparoscopic Incisions Improve Hormonal Response and Fertility in Patients with Extremely Poor Ovarian Response. J. Minim. Invasive Gynecol. 2022, 29, 905–914. [Google Scholar] [CrossRef]
- Tanaka, Y.; Hsueh, A.J.; Kawamura, K. Surgical approaches of drug-free in vitro activation and laparoscopic ovarian incision to treat patients with ovarian infertility. Fertil. Steril. 2020, 114, 1355–1357. [Google Scholar] [CrossRef] [PubMed]
- Devenutto, L.; Quintana, R.; Quintana, T. In vitro activation of ovarian cortex and autologous transplantation: A novel approach to primary ovarian insufficiency and diminished ovarian reserve. Hum. Reprod. Open 2020, 2020, hoaa046. [Google Scholar] [CrossRef]
- Hansen, K.R.; Knowlton, N.S.; Thyer, A.C.; Charleston, J.S.; Soules, M.R.; Klein, N.A. A new model of reproductive aging: The decline in ovarian non-growing follicle number from birth to menopause. Hum. Reprod. 2008, 23, 699–708. [Google Scholar] [CrossRef]
- Harlev, A.; Walfisch, A.; Oran, E.; Har-Vardi, I.; Friger, M.; Lunenfeld, E.; Levitas, E. The effect of fertility treatment on adverse perinatal outcomes in women aged at least 40 years. Int. J. Gynecol. Obstet. 2018, 140, 98–104. [Google Scholar] [CrossRef]
- Velde, E.R.T.; Pearson, P.L. The variability of female reproductive ageing. Hum. Reprod. Update 2002, 8, 141–154. [Google Scholar] [CrossRef] [PubMed]
- Vollenhoven, B.; Hunt, S. Ovarian ageing and the impact on female fertility. F1000Research 2018, 7, 1835. [Google Scholar] [CrossRef] [PubMed]
- Li, C.-J.; Lin, L.-T.; Tsai, H.-W.; Chern, C.-U.; Wen, Z.-H.; Wang, P.-H.; Tsui, K.-H. The Molecular Regulation in the Pathophysiology in Ovarian Aging. Aging Dis. 2021, 12, 934–949. [Google Scholar] [CrossRef] [PubMed]
- Kupesic, S.; Kurjak, A.; Bjelos, D.; Vujisic, S. Three-dimensional ultrasonographic ovarian measurements and in vitro fertilization outcome are related to age. Fertil. Steril. 2003, 79, 190–197. [Google Scholar] [CrossRef] [PubMed]
- Cimadomo, D.; Fabozzi, G.; Vaiarelli, A.; Ubaldi, N.; Ubaldi, F.M.; Rienzi, L. Impact of Maternal Age on Oocyte and Embryo Competence. Front. Endocrinol. 2018, 9, 327. [Google Scholar] [CrossRef]
- Choi, J.K.; Ahn, J.I.; Park, J.H.; Lim, J.M. Derivation of developmentally competent oocytes by in vitro culture of preantral follicles retrieved from aged mice. Fertil. Steril. 2011, 95, 1487–1489. [Google Scholar] [CrossRef] [PubMed]
- Landry, D.A.; Bellefleur, A.-M.; Labrecque, R.; Grand, F.-X.; Vigneault, C.; Blondin, P.; Sirard, M.-A. Effect of cow age on the in vitro developmental competence of oocytes obtained after FSH stimulation and coasting treatments. Theriogenology 2016, 86, 1240–1246. [Google Scholar] [CrossRef] [PubMed]
- Bozkurt, B.; Erdem, M.; Mutlu, M.F.; Erdem, A.; Guler, I.; Mutlu, I.; Oktem, M. Comparison of age-related changes in anti-Müllerian hormone levels and other ovarian reserve tests between healthy fertile and infertile population. Hum. Fertil. 2016, 19, 192–198. [Google Scholar] [CrossRef]
- Iliodromiti, S.; Sanchez, C.I.; Messow, C.-M.; Cruz, M.; Velasco, J.G.; Nelson, S.M. Excessive age-related decline in functional ovarian reserve in infertile women: Prospective cohort of 15,500 women. J. Clin. Endocrinol. Metab. 2016, 101, 3548–3554. [Google Scholar] [CrossRef]
- Gruijters, M.J.; Visser, J.A.; Durlinger, A.L.; Themmen, A.P. Anti-Müllerian hormone and its role in ovarian function. Mol. Cell. Endocrinol. 2003, 211, 85–90. [Google Scholar] [CrossRef]
- Park, M.J.; Ahn, J.-W.; Kim, K.H.; Bang, J.; Kim, S.C.; Jeong, J.Y.; Choi, Y.E.; Kim, C.-W.; Joo, B.S. Prediction of ovarian aging using ovarian expression of BMP15, GDF9, and C-KIT. Exp. Biol. Med. 2020, 245, 711–719. [Google Scholar] [CrossRef] [PubMed]
- Depmann, M.; Broer, S.L.; Van Der Schouw, Y.T.; Tehrani, F.R.; Eijkemans, M.J.; Mol, B.W.; Broekmans, F.J. Can we predict age at natural menopause using ovarian reserve tests or mother’s age at menopause? A systematic literature review. Menopause 2016, 23, 224–232. [Google Scholar] [CrossRef] [PubMed]
- Kedem, A.; Yung, Y.; Yerushalmi, G.M.; Haas, J.; Maman, E.; Hanochi, M.; Hemi, R.; Orvieto, R.; Dor, J.; Hourvitz, A. Anti Müllerian Hormone (AMH) level and expression in mural and cumulus cells in relation to age. J. Ovarian Res. 2014, 7, 113. [Google Scholar] [CrossRef] [PubMed]
- Dewailly, D.; Andersen, C.Y.; Balen, A.; Broekmans, F.; Dilaver, N.; Fanchin, R.; Griesinger, G.; Kelsey, T.W.; La Marca, A.; Lambalk, C.; et al. The physiology and clinical utility of anti-Mullerian hormone in women. Hum. Reprod. Update 2014, 20, 370–385. [Google Scholar] [CrossRef] [PubMed]
- Honnma, H.; Baba, T.; Sasaki, M.; Hashiba, Y.; Oguri, H.; Fukunaga, T.; Endo, T.; Asada, Y. Different ovarian response by age in an anti-Müllerian hormone-matched group undergoing in vitro fertilization. J. Assist. Reprod. Genet. 2012, 29, 117–125. [Google Scholar] [CrossRef] [PubMed]
- Adhikari, D.; Lee, I.-W.; Yuen, W.S.; Carroll, J. Oocyte mitochondria—Key regulators of oocyte function and potential therapeutic targets for improving fertility. Biol. Reprod. 2022, 106, 366–377. [Google Scholar] [CrossRef] [PubMed]
- Meldrum, D.R. Female reproductive aging—Ovarian and uterine factors. Fertil. Steril. 1993, 59, 1–5. [Google Scholar] [CrossRef]
- Sauer, M.V.; Paulson, R.J.; Ary, B.A.; Lobo, R.A. Three hundred cycles of oocyte donation at the University of Southern California: Assessing the effect of age and infertility diagnosis on pregnancy and implantation rates. J. Assist. Reprod. Genet. 1994, 11, 92–96. [Google Scholar] [CrossRef]
- Ng, E.H.Y.; Tang, O.S.; Ho, P.C. The significance of the number of antral follicles prior to stimulation in predicting ovarian responses in an IVF programme. Hum. Reprod. 2000, 15, 1937–1942. [Google Scholar] [CrossRef]
- Muhammad, F.; Yivgi-Ohana, N.; Shveiky, D.; Orly, J.; Alexander, S.; Laufer, N. Levels of steroidogenic acute regulatory protein and mitochondrial membrane potential in granulosa cells of older poor-responder women. Fertil. Steril. 2009, 91, 220–225. [Google Scholar]
- Tsafrir, A.; Simon, A.; Revel, A.; Reubinoff, B.; Lewin, A.; Laufer, N. Retrospective analysis of 1217 IVF cycles in women aged 40 years and older. Reprod. Biomed. Online 2007, 14, 348–355. [Google Scholar] [CrossRef] [PubMed]
- Saldeen, P.; Källen, K.; Sundström, P. The probability of successful IVF outcome after poor ovarian response. Acta Obstet. Gynecol. Scand. 2007, 86, 457–461. [Google Scholar] [CrossRef] [PubMed]
- Gleicher, N.; Kushnir, V.A.; Barad, D.H. Prospectively assessing risk for premature ovarian senescence in young females: A new paradigm. Reprod. Biol. Endocrinol. 2015, 13, 34. [Google Scholar] [CrossRef] [PubMed]
- Golezar, S.; Tehrani, F.R.; Khazaei, S.; Ebadi, A.; Keshavarz, Z. The global prevalence of primary ovarian insufficiency and early menopause: A meta-analysis. Climacteric 2019, 22, 403–411. [Google Scholar] [CrossRef] [PubMed]
- Ke, H.; Tang, S.; Guo, T.; Hou, D.; Jiao, X.; Li, S.; Luo, W.; Xu, B.; Zhao, S.; Li, G.; et al. Landscape of pathogenic mutations in premature ovarian insufficiency. Nat. Med. 2023, 29, 483–492. [Google Scholar] [CrossRef] [PubMed]
- Subramanian, G.N.; Lavin, M.; Homer, H.A. Premature ovarian ageing following heterozygous loss of Senataxin. Mol. Hum. Reprod. 2021, 27, gaaa080. [Google Scholar] [CrossRef] [PubMed]
- Kushnir, V.A.; Yu, Y.; Barad, D.H.; Weghofer, A.; Himaya, E.; Lee, H.-J.; Wu, Y.-G.; Shohat-Tal, A.; Lazzaroni-Tealdi, E.; Gleicher, N. Utilizing FMR1 Gene Mutations as Predictors of Treatment Success in Human In Vitro Fertilization. PLoS ONE 2014, 9, e102274. [Google Scholar] [CrossRef]
- Murray, A.; Schoemaker, M.J.; Bennett, C.E.; Ennis, S.; Macpherson, J.N.; Jones, M.; Morris, D.H.; Orr, N.; Ashworth, A.; Jacobs, P.A.; et al. Population-based estimates of the prevalence of FMR1 expansion mutations in women with early menopause and primary ovarian insufficiency. Anesth. Analg. 2014, 16, 19–24. [Google Scholar] [CrossRef]
- Ruth, K.S.; Day, F.R.; Hussain, J.; Martínez-Marchal, A.; Aiken, C.E.; Azad, A.; Thompson, D.J.; Knoblochova, L.; Abe, H.; Tarry-Adkins, J.L.; et al. Genetic insights into biological mechanisms governing human ovarian ageing. Nature 2021, 596, 393–397. [Google Scholar] [CrossRef]
- Smits, M.A.J.; Janssens, G.E.; Goddijn, M.; Hamer, G.; Houtkooper, R.H.; Mastenbroek, S. Longevity pathways are associated with human ovarian ageing. Hum. Reprod. Open 2021, 2021, hoab020. [Google Scholar] [CrossRef]
- Louwers, Y.V.; Visser, J.A. Shared Genetics Between Age at Menopause, Early Menopause, POI and Other Traits. Front. Genet. 2021, 12, 676546. [Google Scholar] [CrossRef] [PubMed]
- Homer, H.A. Senataxin: A New Guardian of the Female Germline Important for Delaying Ovarian Aging. Front. Genet. 2021, 12, 647996. [Google Scholar] [CrossRef] [PubMed]
- McGrath, I.M.; Mortlock, S.; Montgomery, G.W. Genetic Regulation of Physiological Reproductive Lifespan and Female Fertility. Int. J. Mol. Sci. 2021, 22, 2556. [Google Scholar] [CrossRef] [PubMed]
- Pelosi, E.; Simonsick, E.; Forabosco, A.; Garcia-Ortiz, J.E.; Schlessinger, D. Dynamics of the Ovarian Reserve and Impact of Genetic and Epidemiological Factors on Age of Menopause. Biol. Reprod. 2015, 92, 130. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Zhang, X.; Jiao, J.; Zhang, F.; Pan, Y.; Wang, Q.; Chen, Q.; Cai, B.; Tang, S.; Zhou, Z.; et al. Rare variants in FANCA induce premature ovarian insufficiency. Hum. Genet. 2019, 138, 1227–1236. [Google Scholar] [CrossRef] [PubMed]
- Wasielak-Politowska, M.; Kordowitzki, P. Chromosome Segregation in the Oocyte: What Goes Wrong during Aging. Int. J. Mol. Sci. 2022, 23, 2880. [Google Scholar] [CrossRef] [PubMed]
- Handyside, A.H.; Montag, M.; Magli, M.C.; Repping, S.; Harper, J.; Schmutzler, A.; Vesela, K.; Gianaroli, L.; Geraedts, J. Multiple meiotic errors caused by predivision of chromatids in women of advanced maternal age undergoing in vitro fertilisation. Eur. J. Hum. Genet. 2012, 20, 742–747. [Google Scholar] [CrossRef]
- Aubert, G.; Lansdorp, P.M. Telomeres and aging. Physiol. Rev. 2008, 88, 557–579. [Google Scholar] [CrossRef]
- Tire, B.; Ozturk, S. Potential effects of assisted reproductive technology on telomere length and telomerase activity in human oocytes and early embryos. J. Ovarian Res. 2023, 16, 130. [Google Scholar] [CrossRef]
- Kohlrausch, F.B.; Wang, F.; Chamani, I.; Keefe, D.L. Telomere Shortening and Fusions: A Link to Aneuploidy in Early Human Embryo Development. Obstet. Gynecol. Surv. 2021, 76, 429–436. [Google Scholar] [CrossRef]
- Zhang, D.; Zhang, X.; Zeng, M.; Yuan, J.; Liu, M.; Yin, Y.; Wu, X.; Keefe, D.L.; Liu, L. Increased DNA damage and repair deficiency in granulosa cells are associated with ovarian aging in rhesus monkey. J. Assist. Reprod. Genet. 2015, 32, 1069–1078. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.; Bohlin, J.; Page, C.M.; Nustad, H.E.; Harris, J.R.; Magnus, P.; Jugessur, A.; Magnus, M.C.; Håberg, S.E.; Hanevik, H.I. Associations between epigenetic age acceleration and infertility. Hum. Reprod. 2022, 37, 2063–2074. [Google Scholar] [CrossRef] [PubMed]
- Cheng, E.-H.; Chen, S.-U.; Lee, T.-H.; Pai, Y.-P.; Huang, L.-S.; Huang, C.-C.; Lee, M.-S. Evaluation of telomere length in cumulus cells as a potential biomarker of oocyte and embryo quality. Hum. Reprod. 2013, 28, 929–936. [Google Scholar] [CrossRef] [PubMed]
- Keefe, D.L. Telomeres, Reproductive Aging, and Genomic Instability During Early Development. Reprod. Sci. 2016, 23, 1612–1615. [Google Scholar] [CrossRef] [PubMed]
- Dell’aversana, C.; Cuomo, F.; Longobardi, S.; D’hooghe, T.; Caprio, F.; Franci, G.; Santonastaso, M.; Colacurci, N.; Barone, S.; Pisaturo, V.; et al. Age-related miRNome landscape of cumulus oophorus cells during controlled ovarian stimulation protocols in IVF cycles. Hum. Reprod. 2021, 36, 1310–1325. [Google Scholar] [CrossRef] [PubMed]
- Caponnetto, A.; Battaglia, R.; Ferrara, C.; Vento, M.E.; Borzì, P.; Paradiso, M.; Scollo, P.; Purrello, M.; Longobardi, S.; D’Hooghe, T.; et al. Down-regulation of long non-coding RNAs in reproductive aging and analysis of the lncRNA-miRNA-mRNA networks in human cumulus cells. J. Assist. Reprod. Genet. 2022, 39, 919–931. [Google Scholar] [CrossRef] [PubMed]
- Sun, X.; Lu, J.; Li, H.; Huang, B. The Role of m6A on Female Reproduction and Fertility: From Gonad Development to Ovarian Aging. Front. Cell Dev. Biol. 2022, 10, 884295. [Google Scholar] [CrossRef] [PubMed]
- Sasaki, H.; Hamatani, T.; Kamijo, S.; Iwai, M.; Kobanawa, M.; Ogawa, S.; Miyado, K.; Tanaka, M. Impact of Oxidative Stress on Age-Associated Decline in Oocyte Developmental Competence. Front. Endocrinol. 2019, 10, 811. [Google Scholar] [CrossRef]
- Kasapoğlu, I.; Seli, E. Mitochondrial Dysfunction and Ovarian Aging. Endocrinology 2020, 161, bqaa001. [Google Scholar] [CrossRef]
- Yang, L.; Lin, X.; Tang, H.; Fan, Y.; Zeng, S.; Jia, L.; Li, Y.; Shi, Y.; He, S.; Wang, H.; et al. Mitochondrial DNA mutation exacerbates female reproductive aging via impairment of the NADH/NAD+ redox. Aging Cell 2020, 19, e13206. [Google Scholar] [CrossRef]
- Cecchino, G.N.; Seli, E.; da Motta, E.L.A.; García-Velasco, J.A. The role of mitochondrial activity in female fertility and assisted reproductive technologies: Overview and current insights. Reprod. Biomed. Online 2018, 36, 686–697. [Google Scholar] [CrossRef] [PubMed]
- May-Panloup, P.; Boucret, L.; de la Barca, J.M.C.; Desquiret-Dumas, V.; Ferre-L’Hotellier, V.; Moriniere, C.; Descamps, P.; Procaccio, V.; Reynier, P. Ovarian ageing: The role of mitochondria in oocytes and follicles. Hum. Reprod. Update 2016, 22, 725–743. [Google Scholar] [CrossRef] [PubMed]
- Meldrum, D.R.; Casper, R.F.; Diez-Juan, A.; Simon, C.; Domar, A.D.; Frydman, R. Aging and the environment affect gamete and embryo potential: Can we intervene? Fertil. Steril. 2016, 105, 548–559. [Google Scholar] [CrossRef] [PubMed]
- Tatone, C.; Carbone, M.; Falone, S.; Aimola, P.; Giardinelli, A.; Caserta, D.; Marci, R.; Pandolfi, A.; Ragnelli, A.; Amicarelli, F. Age-dependent changes in the expression of superoxide dismutases and catalase are associated with ultrastructural modifications in human granulosa cells. Mol. Hum. Reprod. 2006, 12, 655–660. [Google Scholar] [CrossRef] [PubMed]
- Bowolaksono, A.; Sundari, A.M.; Fauzi, M.; Maidarti, M.; Wiweko, B.; Mutia, K.; Iffanolida, P.A.; Febri, R.R.; Dwiranti, A.; Funahashi, H. Anti-Müllerian hormone independently affect mtDNA copy number in human granulosa cells. J. Ovarian Res. 2022, 15, 111. [Google Scholar] [CrossRef] [PubMed]
- Molinari, E.; Bar, H.; Pyle, A.; Patrizio, P. Transcriptome analysis of human cumulus cells reveals hypoxia as the main determinant of follicular senescence. Mol. Hum. Reprod. 2016, 22, 866–876. [Google Scholar] [CrossRef] [PubMed]
- Tatone, C.; Amicarelli, F.; Carbone, M.C.; Monteleone, P.; Caserta, D.; Marci, R.; Artini, P.G.; Piomboni, P.; Focarelli, R. Cellular and molecular aspects of ovarian follicle ageing. Hum. Reprod. Update 2008, 14, 131–142. [Google Scholar] [CrossRef] [PubMed]
- Mobarak, H.; Heidarpour, M.; Tsai, P.-S.J.; Rezabakhsh, A.; Rahbarghazi, R.; Nouri, M.; Mahdipour, M. Autologous mitochondrial microinjection: A strategy to improve the oocyte quality and subsequent reproductive outcome during aging. Cell Biosci. 2019, 9, 95. [Google Scholar] [CrossRef]
- Jiang, Z.; Shen, H. Mitochondria: Emerging therapeutic strategies for oocyte rescue. Reprod. Sci. 2022, 29, 711–722. [Google Scholar] [CrossRef]
- Cozzolino, M.; Marin, D.; Sisti, G. New Frontiers in IVF: mtDNA and autologous germline mitochondrial energy transfer. Reprod. Biol. Endocrinol. 2019, 17, 55. [Google Scholar] [CrossRef]
- Camaioni, A.; Ucci, M.A.; Campagnolo, L.; De Felici, M.; Klinger, F.G. The process of ovarian aging: It is not just about oocytes and granulosa cells. J. Assist. Reprod. Genet. 2022, 39, 783–792. [Google Scholar] [CrossRef]
- Ahmed, T.A.; Ahmed, S.M.; El-Gammal, Z.; Shouman, S.; Ahmed, A.; Mansour, R.; El-Badri, N. Oocyte Aging: The Role of Cellular and Environmental Factors and Impact on Female Fertility. Adv. Exp. Med. Biol. 2020, 1247, 109–123. [Google Scholar] [PubMed]
- Ouni, E.; Bouzin, C.; Dolmans, M.M.; Marbaix, E.; Ruys, S.P.D.; Vertommen, D.; Amorim, C.A. Spatiotemporal changes in mechanical matrisome components of the human ovary from prepuberty to menopause. Hum. Reprod. 2020, 35, 1391–1410. [Google Scholar] [CrossRef] [PubMed]
- Landry, D.A.; Vaishnav, H.T.; Vanderhyden, B.C. The significance of ovarian fibrosis. Oncotarget 2020, 11, 4366–4370. [Google Scholar] [CrossRef]
- Laronda, M.M.; Jakus, A.E.; Whelan, K.A.; Wertheim, J.A.; Shah, R.N.; Woodruff, T.K. Initiation of puberty in mice following decellularized ovary transplant. Biomaterials 2015, 50, 20–29. [Google Scholar] [CrossRef]
- Pennarossa, G.; De Iorio, T.; Gandolfi, F.; Brevini, T.A.L. Impact of Aging on the Ovarian Extracellular Matrix and Derived 3D Scaffolds. Nanomaterials 2022, 12, 345. [Google Scholar] [CrossRef]
- Amargant, F.; Manuel, S.L.; Tu, Q.; Parkes, W.S.; Rivas, F.; Zhou, L.T.; Rowley, J.E.; Villanueva, C.E.; Hornick, J.E.; Shekhawat, G.S.; et al. Ovarian stiffness increases with age in the mammalian ovary and depends on collagen and hyaluronan matrices. Aging Cell 2020, 19, e13259. [Google Scholar] [CrossRef] [PubMed]
- Salwowska, N.M.; Bebenek, K.A.; Żądło, D.A.; Wcisło-Dziadecka, D.L. Physiochemical properties and application of hyaluronic acid: A systematic review. J. Cosmet. Dermatol. 2016, 15, 520–526. [Google Scholar] [CrossRef]
- Rowley, J.E.; Amargant, F.; Zhou, L.T.; Galligos, A.; Simon, L.E.; Pritchard, M.T.; Duncan, F.E. Low Molecular Weight Hyaluronan Induces an Inflammatory Response in Ovarian Stromal Cells and Impairs Gamete Development In Vitro. Int. J. Mol. Sci. 2020, 21, 1036. [Google Scholar] [CrossRef]
- Marei, W.; Ghafari, F.; Fouladi-Nashta, A. Role of hyaluronic acid in maturation and further early embryo development of bovine oocytes. Theriogenology 2012, 78, 670–677. [Google Scholar] [CrossRef]
- Zhao, G.; Yan, G.; Cheng, J.; Zhou, X.; Fang, T.; Sun, H.; Hou, Y.; Hu, Y. Hyaluronic acid prevents immunosuppressive drug-induced ovarian damage via up-regulating PGRMC1 expression. Sci. Rep. 2015, 5, 7647. [Google Scholar] [CrossRef] [PubMed]
- Oksjoki, S.; Sallinen, S.; Vuorio, E.; Anttila, L. Cyclic expression of mRNA transcripts for connective tissue components in the mouse ovary. Mol. Hum. Reprod. 1999, 5, 803–808. [Google Scholar] [CrossRef] [PubMed]
- Birch, H.L. Extracellular Matrix and Ageing. Subcell. Biochem. 2018, 90, 169–190. [Google Scholar] [PubMed]
- Fessel, G.; Li, Y.; Diederich, V.; Guizar-Sicairos, M.; Schneider, P.; Sell, D.R.; Monnier, V.M.; Snedeker, J.G. Advanced Glycation End-Products Reduce Collagen Molecular Sliding to Affect Collagen Fibril Damage Mechanisms but Not Stiffness. PLoS ONE 2014, 9, e110948. [Google Scholar] [CrossRef] [PubMed]
- Vijg, J.; Campisi, J. Puzzles, promises and a cure for ageing. Nature 2008, 454, 1065–1071. [Google Scholar] [CrossRef] [PubMed]
- Wick, G.; Grundtman, C.; Mayerl, C.; Wimpissinger, T.F.; Feichtinger, J.; Zelger, B.; Sgonc, R.; Wolfram, D. The immunology of fibrosis. Annu. Rev. Immunol. 2013, 31, 107–135. [Google Scholar] [CrossRef] [PubMed]
- Statzer, C.; Park, J.Y.C.; Ewald, C.Y. Extracellular Matrix Dynamics as an Emerging yet Understudied Hallmark of Aging and Longevity. Aging Dis. 2023, 14, 670–693. [Google Scholar] [CrossRef]
- Sharma, D.; Bhartiya, D. Aged mice ovaries harbor stem cells and germ cell nests but fail to form follicles. J. Ovarian Res. 2022, 15, 37. [Google Scholar] [CrossRef]
- Silvestris, E.; Cafforio, P.; D’oronzo, S.; Felici, C.; Silvestris, F.; Loverro, G. In vitro differentiation of human oocyte-like cells from oogonial stem cells: Single-cell isolation and molecular characterization. Hum. Reprod. 2018, 33, 464–473. [Google Scholar] [CrossRef]
- Niikura, Y.; Niikura, T.; Tilly, J.L. Aged mouse ovaries possess rare premeiotic germ cells that can generate oocytes following transplantation into a young host environment. Aging 2009, 1, 971–978. [Google Scholar] [CrossRef]
- Guilak, F.; Cohen, D.M.; Estes, B.T.; Gimble, J.M.; Liedtke, W.; Chen, C.S. Control of stem cell fate by physical interactions with the extracellular matrix. Cell Stem Cell 2009, 5, 17–26. [Google Scholar] [CrossRef] [PubMed]
- MacDonald, J.A.; Takai, Y.; Ishihara, O.; Seki, H.; Woods, D.C.; Tilly, J.L. Extracellular matrix signaling activates differentiation of adult ovary-derived oogonial stem cells in a species-specific manner. Fertil. Steril. 2019, 111, 794–805. [Google Scholar] [CrossRef] [PubMed]
- Younis, J.S.; Haddad, S.; Matilsky, M.; Radin, O.; Ben-Ami, M. Undetectable basal ovarian stromal blood flow in infertile women is related to low ovarian reserve. Gynecol. Endocrinol. 2007, 23, 284–289. [Google Scholar] [CrossRef] [PubMed]
- Meirow, D.; Dor, J.; Kaufman, B.; Shrim, A.; Rabinovici, J.; Schiff, E.; Raanani, H.; Levron, J.; Fridman, E. Cortical fibrosis and blood-vessels damage in human ovaries exposed to chemotherapy. Potential mechanisms of ovarian injury. Hum. Reprod. 2007, 22, 1626–1633. [Google Scholar] [CrossRef] [PubMed]
- Kinnear, H.M.; Tomaszewski, C.E.; Chang, F.L.; Moravek, M.B.; Xu, M.; Padmanabhan, V.; Shikanov, A. The ovarian stroma as a new frontier. Reproduction 2020, 160, R25–R39. [Google Scholar] [CrossRef] [PubMed]
- Kizuka-Shibuya, F.; Tokuda, N.; Takagi, K.; Adachi, Y.; Lee, L.; Tamura, I.; Maekawa, R.; Tamura, H.; Suzuki, T.; Owada, Y.; et al. Locally existing endothelial cells and pericytes in ovarian stroma, but not bone marrow-derived vascular progenitor cells, play a central role in neovascularization during follicular development in mice. J. Ovarian Res. 2014, 7, 10. [Google Scholar] [CrossRef]
- Van Blerkom, J. Epigenetic Influences on Oocyte Developmental Competence: Perifollicular Vascularity and Intrafollicular Oxygen. J. Assist. Reprod. Genet. 1998, 15, 226–234. [Google Scholar] [CrossRef] [PubMed]
- Borini, A.; Maccolini, A.; Tallarini, A.; Bonu, M.A.; Sciajno, R.; Flamigni, C. Perifollicular vascularity and its relationship with oocyte maturity and IVF outcome. Ann. N. Y. Acad. Sci. 2001, 943, 64–67. [Google Scholar] [CrossRef]
- Naredi, N.; Singh, S.K.; Sharma, R. Does Perifollicular Vascularity on the Day of Oocyte Retrieval Affect Pregnancy Outcome in an In Vitro Fertilization Cycle? J. Hum. Reprod. Sci. 2017, 10, 281–287. [Google Scholar] [CrossRef]
- Vengetesh, P.M.; Ramachandran, A.; Kumar, P. Choosing GnRH Antagonist Protocol Shows Improved Oocyte and Embryo Quality, Coherent with the Perifollicular Vascularity (PFV) in Assisted Reproductive Techniques. J. Clin. Diagn. Res. 2015, 9, QC24–QC28. [Google Scholar] [CrossRef]
- Huyghe, S.; Verest, A.; Thijssen, A.; Ombelet, W. The prognostic value of perifollicular blood flow in the outcome after assisted reproduction: A systematic review. Facts Views Vis. Obgyn. 2017, 9, 153–156. [Google Scholar] [PubMed]
- Salvatore, G.; De Felici, M.; Dolci, S.; Tudisco, C.; Cicconi, R.; Campagnolo, L.; Camaioni, A.; Klinger, F.G. Human adipose-derived stromal cells transplantation prolongs reproductive lifespan on mouse models of mild and severe premature ovarian insufficiency. Stem Cell Res. Ther. 2021, 12, 537. [Google Scholar] [CrossRef] [PubMed]
- Cho, J.; Kim, T.-H.; Seok, J.; Jun, J.H.; Park, H.; Kweon, M.; Lim, J.-Y.; Kim, G.J. Vascular remodeling by placenta-derived mesenchymal stem cells restores ovarian function in ovariectomized rat model via the VEGF pathway. Lab. Investig. 2021, 101, 304–317. [Google Scholar] [CrossRef] [PubMed]
- Chan, K.A.; Bernal, A.B.; Vickers, M.H.; Gohir, W.; Petrik, J.J.; Sloboda, D.M. Early Life Exposure to Undernutrition Induces ER Stress, Apoptosis, and Reduced Vascularization in Ovaries of Adult Rat Offspring. Biol. Reprod. 2015, 92, 110. [Google Scholar] [CrossRef]
- Xu, X.; Mu, L.; Li, L.; Liang, J.; Zhang, S.; Jia, L.; Yang, X.; Dai, Y.; Zhang, J.; Wang, Y.; et al. Imaging and tracing the pattern of adult ovarian angiogenesis implies a strategy against female reproductive aging. Sci. Adv. 2022, 8, eabi8683. [Google Scholar] [CrossRef]
- Huang, Y.; Tu, M.; Qian, Y.; Ma, J.; Chen, L.; Liu, Y.; Wu, Y.; Chen, K.; Liu, J.; Ying, Y.; et al. Age-Dependent Metabolomic Profile of the Follicular Fluids from Women Undergoing Assisted Reproductive Technology Treatment. Front. Endocrinol. 2022, 13, 818888. [Google Scholar] [CrossRef]
- Cordeiro, F.B.; Montani, D.A.; Pilau, E.J.; Gozzo, F.C.; Fraietta, R.; Turco, E.G.L. Ovarian environment aging: Follicular fluid lipidomic and related metabolic pathways. J. Assist. Reprod. Genet. 2018, 35, 1385–1393. [Google Scholar] [CrossRef]
- de la Barca, J.M.C.; Boueilh, T.; Simard, G.; Boucret, L.; Ferré-L’hotellier, V.; Tessier, L.; Gadras, C.; Bouet, P.E.; Descamps, P.; Procaccio, V.; et al. Targeted metabolomics reveals reduced levels of polyunsaturated choline plasmalogens and a smaller dimethylarginine/arginine ratio in the follicular fluid of patients with a diminished ovarian reserve. Hum. Reprod. 2017, 32, 2269–2278. [Google Scholar] [CrossRef]
- Bouet, P.-E.; Boueilh, T.; de la Barca, J.M.C.; Boucret, L.; Blanchard, S.; Ferré-L’hotellier, V.; Jeannin, P.; Descamps, P.; Procaccio, V.; Reynier, P.; et al. The cytokine profile of follicular fluid changes during ovarian ageing. J. Gynecol. Obstet. Hum. Reprod. 2020, 49, 101704. [Google Scholar] [CrossRef]
- Southcombe, J.H.; Ledee, N.; Perrier d’Hauterive, S.; Turner, K.; Child, T.; Snider, J.V.; Redman, C.W.G.; Sargent, I.L.; Granne, I. Detection of soluble ST2 in human follicular fluid and luteinized granulosa cells. PLoS ONE 2013, 8, e74385. [Google Scholar] [CrossRef]
- Ledee, N.; Gridelet, V.; Ravet, S.; Jouan, C.; Gaspard, O.; Wenders, F.; Thonon, F.; Hincourt, N.; Dubois, M.; Foidart, J.M.; et al. Impact of follicular G-CSF quantification on subsequent embryo transfer decisions: A proof of concept study. Hum. Reprod. 2013, 28, 406–413. [Google Scholar] [CrossRef] [PubMed]
- Tournaye, H.; D’Hooghe, T.; Verheyen, G.; Devreker, K.F.; Perrier d’Hauterive, S.; Nisolle, M.; Foidart, J.-M.; Munaut, C.; Noel, L. Clinical performance of a specific granulocyte colony stimulating factor ELISA to determine its concentration in follicular fluid as a predictor of implantation success during in vitro fertilization. Gynecol. Endocrinol. 2020, 36, 44–48. [Google Scholar] [CrossRef] [PubMed]
- Ranjbaran, A.; Nejabati, H.R.; Ghasemnejad, T.; Latifi, Z.; Hamdi, K.; Hajipour, H.; Raffel, N.; Bahrami-Asl, Z.; Hakimi, P.; Mihanfar, A.; et al. Follicular Fluid Levels of Adrenomedullin 2, Vascular Endothelial Growth Factor and its Soluble Receptors Are Associated with Ovarian Response During ART Cycles. Geburtshilfe Frauenheilkd. 2019, 79, 86–93. [Google Scholar] [CrossRef] [PubMed]
- Marinoni, E.; Feliciani, E.; Muzzonigro, F.; Letizia, C.; Tranquilli, A.; Tranquilli, D.; Aragona, C.; Moscarini, M.; Di Iorio, R. Intrafollicular concentration of adrenomedullin is associated with IVF outcome. Gynecol. Endocrinol. 2010, 26, 435–439. [Google Scholar] [CrossRef] [PubMed]
- Nacak, G.B.; Ozkaya, E.; Abide, C.Y.; Bilgic, B.E.; Devranoglu, B.; Iscan, R.G. The impact of autoimmunity-related early ovarian aging on ICSI cycle outcome. Gynecol. Endocrinol. 2018, 34, 940–943. [Google Scholar] [CrossRef] [PubMed]
- Engelmann, F.; Barron, A.; Urbanski, H.; Neuringer, M.; Kohama, S.G.; Park, B.; Messaoudi, I. Accelerated immune senescence and reduced response to vaccination in ovariectomized female rhesus macaques. Age 2011, 33, 275–289. [Google Scholar] [CrossRef] [PubMed]
- Lliberos, C.; Liew, S.H.; Zareie, P.; La Gruta, N.L.; Mansell, A.; Hutt, K. Evaluation of inflammation and follicle depletion during ovarian ageing in mice. Sci. Rep. 2021, 11, 278. [Google Scholar] [CrossRef]
- Machlin, J.H.; Barishansky, S.J.; Kelsh, J.; Larmore, M.J.; Johnson, B.W.; Pritchard, M.T.; Pavone, M.E.; Duncan, F.E. Fibroinflammatory Signatures Increase with Age in the Human Ovary and Follicular Fluid. Int. J. Mol. Sci. 2021, 22, 4902. [Google Scholar] [CrossRef]
- Malizia, B.A.; Wook, Y.S.; Penzias, A.S.; Usheva, A. The human ovarian follicular fluid level of interleukin-8 is associated with follicular size and patient age. Fertil. Steril. 2010, 93, 537–543. [Google Scholar] [CrossRef]
- Navarro-Pando, J.M.; Alcocer-Gómez, E.; Castejón-Vega, B.; Navarro-Villarán, E.; Condés-Hervás, M.; Mundi-Roldan, M.; Muntané, J.; Pérez-Pulido, A.J.; Bullon, P.; Wang, C.; et al. Inhibition of the NLRP3 inflammasome prevents ovarian aging. Sci. Adv. 2021, 7, eabc7409. [Google Scholar] [CrossRef]
- Ramirez, J.; Bitterman, P.; Basu, S.; Barua, A. Changes in IL-16 Expression in the Ovary during Aging and Its Potential Consequences to Ovarian Pathology. J. Immunol. Res. 2022, 2022, 2870389. [Google Scholar] [CrossRef] [PubMed]
- Blumenfeld, Z. What Is the Best Regimen for Ovarian Stimulation of Poor Responders in ART/IVF? Front. Endocrinol. 2020, 11, 192. [Google Scholar] [CrossRef] [PubMed]
- Regan, S.L.; Knight, P.G.; Yovich, J.L.; Arfuso, F.; Dharmarajan, A. Growth hormone during in vitro fertilization in older women modulates the density of receptors in granulosa cells, with improved pregnancy outcomes. Fertil. Steril. 2018, 110, 1298–1310. [Google Scholar] [CrossRef] [PubMed]
- Hart, R.J. Use of Growth Hormone in the IVF Treatment of Women with Poor Ovarian Reserve. Front. Endocrinol. 2019, 10, 500. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Zhang, C.; Shu, J.; Guo, J.; Chang, H.-M.; Leung, P.C.K.; Sheng, J.-Z.; Huang, H. Adjuvant treatment strategies in ovarian stimulation for poor responders undergoing IVF: A systematic review and network meta-analysis. Hum. Reprod. Update 2020, 26, 247–263. [Google Scholar] [CrossRef] [PubMed]
- Jiang, L.; Cui, J.; Zhang, C.; Xie, J.; Zhang, S.; Fu, D.; Duo, W. Sigma-1 receptor is involved in diminished ovarian reserve possibly by influencing endoplasmic reticulum stress-mediated granulosa cells apoptosis. Aging 2020, 12, 9041–9065. [Google Scholar] [CrossRef] [PubMed]
- Cakiroglu, Y.; Saltik, A.; Yuceturk, A.; Karaosmanoglu, O.; Kopuk, S.Y.; Scott, R.T.; Tiras, B.; Seli, E. Effects of intraovarian injection of autologous platelet rich plasma on ovarian reserve and IVF outcome parameters in women with primary ovarian insufficiency. Aging 2020, 12, 10211–10222. [Google Scholar] [CrossRef] [PubMed]
- Sills, E.S. Ovarian recovery via autologous platelet-rich plasma: New benchmarks for condensed cytokine applications to reverse reproductive aging. Aging Med. 2022, 5, 63–67. [Google Scholar] [CrossRef]
- Sills, E.S.; Wood, S.H. Autologous activated platelet-rich plasma injection into adult human ovary tissue: Molecular mechanism, analysis, and discussion of reproductive response. Biosci. Rep. 2019, 39, BSR20190805. [Google Scholar] [CrossRef]
- Cakiroglu, Y.; Yuceturk, A.; Karaosmanoglu, O.; Kopuk, S.Y.; Korun, Z.E.U.; Herlihy, N.; Scott, R.T.; Tiras, B.; Seli, E. Ovarian reserve parameters and IVF outcomes in 510 women with poor ovarian response (POR) treated with intraovarian injection of autologous platelet rich plasma (PRP). Aging 2022, 14, 2513–2523. [Google Scholar] [CrossRef]
- Pantos, K.; Simopoulou, M.; Pantou, A.; Rapani, A.; Tsioulou, P.; Nitsos, N.; Syrkos, S.; Pappas, A.; Koutsilieris, M.; Sfakianoudis, K. A Case Series on Natural Conceptions Resulting in Ongoing Pregnancies in Menopausal and Prematurely Menopausal Women Following Platelet-Rich Plasma Treatment. Cell Transplant. 2019, 28, 1333–1340. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Cheung, H.-H.; Zhang, C.; Wu, J.; Chan, W.-Y. Melatonin as Potential Targets for Delaying Ovarian Aging. Curr. Drug Targets 2019, 20, 16–28. [Google Scholar] [CrossRef] [PubMed]
- Isola, J.V.; Zanini, B.M.; Hense, J.D.; Alvarado-Rincón, J.A.; Garcia, D.N.; Pereira, G.C.; Vieira, A.D.; Oliveira, T.L.; Collares, T.; Gasperin, B.G.; et al. Mild calorie restriction, but not 17α-estradiol, extends ovarian reserve and fertility in female mice. Exp. Gerontol. 2022, 159, 111669. [Google Scholar] [CrossRef] [PubMed]
- Garcia, D.N.; Saccon, T.D.; Pradiee, J.; Rincón, J.A.A.; Andrade, K.R.S.; Rovani, M.T.; Mondadori, R.G.; Cruz, L.A.X.; Barros, C.C.; Masternak, M.M.; et al. Effect of caloric restriction and rapamycin on ovarian aging in mice. GeroScience 2019, 41, 395–408. [Google Scholar] [CrossRef] [PubMed]
- Lucas, V.; Cavadas, C.; Aveleira, C.A. Cellular Senescence: From Mechanisms to Current Biomarkers and Senotherapies. Pharmacol. Rev. 2023, 75, 675–713. [Google Scholar] [CrossRef] [PubMed]
- Schafer, M.J.; Zhang, X.; Kumar, A.; Atkinson, E.J.; Zhu, Y.; Jachim, S.; Mazula, D.L.; Brown, A.K.; Berning, M.; Aversa, Z.; et al. The senescence-associated secretome as an indicator of age and medical risk. J. Clin. Investig. 2020, 5, e133668. [Google Scholar] [CrossRef] [PubMed]
- Gao, Y.; Wu, T.; Tang, X.; Wen, J.; Zhang, Y.; Zhang, J.; Wang, S. Increased cellular senescence in doxorubicin-induced murine ovarian injury: Effect of senolytics. GeroScience 2023, 45, 1775–1790. [Google Scholar] [CrossRef] [PubMed]
- Hense, J.D.; Garcia, D.N.; Isola, J.V.; Alvarado-Rincón, J.A.; Zanini, B.M.; Prosczek, J.B.; Stout, M.B.; Mason, J.B.; Walsh, P.T.; Brieño-Enríquez, M.A.; et al. Senolytic treatment reverses obesity-mediated senescent cell accumulation in the ovary. GeroScience 2022, 44, 1747–1759. [Google Scholar] [CrossRef]
- Pascuali, N.; Scotti, L.; Di Pietro, M.; Oubiña, G.; Bas, D.; May, M.; Muñoz, A.G.; Cuasnicú, P.S.; Cohen, D.J.; Tesone, M.; et al. Ceramide-1-phosphate has protective properties against cyclophosphamide-induced ovarian damage in a mice model of premature ovarian failure. Hum. Reprod. 2018, 33, 844–859. [Google Scholar] [CrossRef]
- Bertoldo, M.J.; Listijono, D.R.; Ho, W.-H.J.; Riepsamen, A.H.; Goss, D.M.; Richani, D.; Jin, X.L.; Mahbub, S.; Campbell, J.M.; Habibalahi, A.; et al. NAD+ Repletion Rescues Female Fertility during Reproductive Aging. Cell Rep. 2020, 30, 1670–1681.e7. [Google Scholar] [CrossRef]
- Miao, Y.; Cui, Z.; Gao, Q.; Rui, R.; Xiong, B. Nicotinamide Mononucleotide Supplementation Reverses the Declining Quality of Maternally Aged Oocytes. Cell Rep. 2020, 32, 107987. [Google Scholar] [CrossRef] [PubMed]
- Sun, X.; Su, Y.; He, Y.; Zhang, J.; Liu, W.; Zhang, H.; Hou, Z.; Liu, J.; Li, J. New strategy for in vitro activation of primordial follicles with mTOR and PI3K stimulators. Cell Cycle 2015, 14, 721–731. [Google Scholar] [CrossRef] [PubMed]
- Adhikari, D.; Risal, S.; Liu, K.; Shen, Y. Pharmacological Inhibition of mTORC1 Prevents Over-Activation of the Primordial Follicle Pool in Response to Elevated PI3K Signaling. PLoS ONE 2013, 8, e53810. [Google Scholar] [CrossRef] [PubMed]
- Adhikari, D.; Zheng, W.; Shen, Y.; Gorre, N.; Hämäläinen, T.; Cooney, A.J.; Huhtaniemi, I.; Lan, Z.-J.; Liu, K. Tsc/mTORC1 signaling in oocytes governs the quiescence and activation of primordial follicles. Hum. Mol. Genet. 2009, 19, 397–410. [Google Scholar] [CrossRef] [PubMed]
- Grosbois, J.; Demeestere, I. Dynamics of PI3K and Hippo signaling pathways during in vitro human follicle activation. Hum. Reprod. 2018, 33, 1705–1714. [Google Scholar] [CrossRef] [PubMed]
- Yorino, S.; Kawamura, K. Rapamycin treatment maintains developmental potential of oocytes in mice and follicle reserve in human cortical fragments grafted into immune-deficient mice. Mol. Cell. Endocrinol. 2020, 504, 110694. [Google Scholar] [CrossRef] [PubMed]
- Dou, X.; Sun, Y.; Li, J.; Zhang, J.; Hao, D.; Liu, W.; Wu, R.; Kong, F.; Peng, X.; Li, J. Short-term rapamycin treatment increases ovarian lifespan in young and middle-aged female mice. Aging Cell 2017, 16, 825–836. [Google Scholar] [CrossRef] [PubMed]
- Cacciottola, L.; Courtoy, G.E.; Nguyen, T.Y.T.; Hossay, C.; Donnez, J.; Dolmans, M.-M. Adipose tissue–derived stem cells protect the primordial follicle pool from both direct follicle death and abnormal activation after ovarian tissue transplantation. J. Assist. Reprod. Genet. 2021, 38, 151–161. [Google Scholar] [CrossRef]
- El-Derany, M.O.; Said, R.S.; El-Demerdash, E. Bone Marrow-Derived Mesenchymal Stem Cells Reverse Radiotherapy-Induced Premature Ovarian Failure: Emphasis on Signal Integration of TGF-beta, Wnt/β-Catenin and Hippo Pathways. Stem. Cell Rev. Rep. 2021, 17, 1429–1445. [Google Scholar] [CrossRef]
- Volarevic, V.; Bojic, S.; Nurkovic, J.; Volarevic, A.; Ljujic, B.; Arsenijevic, N.; Lako, M.; Stojkovic, M. Stem Cells as New Agents for the Treatment of Infertility: Current and Future Perspectives and Challenges. BioMed Res. Int. 2014, 2014, 507234. [Google Scholar] [CrossRef]
- Huang, B.; Ding, C.; Zou, Q.; Lu, J.; Wang, W.; Li, H. Human Amniotic Fluid Mesenchymal Stem Cells Improve Ovarian Function During Physiological Aging by Resisting DNA Damage. Front. Pharmacol. 2020, 11, 272. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, A.; Yousif, A.; Hong, L.; Chefetz, I. Premature ovarian insufficiency: Pathogenesis and therapeutic potential of mesenchymal stem cell. J. Mol. Med. 2021, 99, 637–650. [Google Scholar] [CrossRef] [PubMed]
- Morohaku, K.; Tanimoto, R.; Sasaki, K.; Kawahara-Miki, R.; Kono, T.; Hayashi, K.; Hirao, Y.; Obata, Y. Complete in vitro generation of fertile oocytes from mouse primordial germ cells. Proc. Natl. Acad. Sci. USA 2016, 113, 9021–9026. [Google Scholar] [CrossRef] [PubMed]
- Woodruff, T.K. Women in Reproductive Science: Lessons from bioengineering the ovarian follicle: A personal perspective. Reproduction 2019, 158, F113–F126. [Google Scholar] [CrossRef] [PubMed]
- Woodruff, T.K.; Shea, L.D. The Role of the Extracellular Matrix in Ovarian Follicle Development. Reprod. Sci. 2007, 14 (Suppl. 8), 6–10. [Google Scholar] [CrossRef] [PubMed]
- Szymanska, K.J.; Tan, X.; Oktay, K. Unraveling the mechanisms of chemotherapy-induced damage to human primordial follicle reserve: Road to developing therapeutics for fertility preservation and reversing ovarian aging. Mol. Hum. Reprod. 2020, 26, 553–566. [Google Scholar] [CrossRef] [PubMed]
- Kawamura, K.; Cheng, Y.; Suzuki, N.; Deguchi, M.; Sato, Y.; Takae, S.; Ho, C.-H.; Kawamura, N.; Tamura, M.; Hashimoto, S.; et al. Hippo signaling disruption and Akt stimulation of ovarian follicles for infertility treatment. Proc. Natl. Acad. Sci. USA 2013, 110, 17474–17479. [Google Scholar] [CrossRef]
- Hayashi, K.; Shimamoto, S.; Nagamatsu, G. Environmental factors for establishment of the dormant state in oocytes. Dev. Growth Differ. 2020, 62, 150–157. [Google Scholar] [CrossRef]
- Zhang, L.; Zhai, J.; Liu, X.; Deng, W.; Wang, H.; Zhang, Z.; Liu, W.; Chen, B.; Wu, C.; Long, H.; et al. Autotransplantation of the ovarian cortex after in-vitro activation for infertility treatment: A shortened procedure. Hum. Reprod. 2021, 36, 2134–2147. [Google Scholar] [CrossRef]
- Kawamura, K.; Ishizuka, B.; Hsueh, A.J. Drug-free in-vitro activation of follicles for infertility treatment in poor ovarian response patients with decreased ovarian reserve. Reprod. Biomed. Online 2020, 40, 245–253. [Google Scholar] [CrossRef]
- Tinjić, S.; Abazović, D.; Ljubić, D.; Vojvodić, D.; Božanović, T.; Ibrišimović, M.; Marković, S.; Ljubić, A. Influence of Autologous In Vitro Activation of Ovaries by Stem Cells and Growth Factors on Endocrine and Reproductive Function of Patients with Ovarian Insufficiency-A Clinical Trial Study. Int. J. Fertil. Steril. 2021, 15, 178–188. [Google Scholar]
- Borreguero-Muñoz, N.; Fletcher, G.C.; Aguilar-Aragon, M.; Elbediwy, A.; Vincent-Mistiaen, Z.I.; Thompson, B.J. The Hippo pathway integrates PI3K–Akt signals with mechanical and polarity cues to control tissue growth. PLoS Biol. 2019, 17, e3000509. [Google Scholar] [CrossRef] [PubMed]
- Shah, J.S.; Sabouni, R.; Vaught, K.C.C.; Owen, C.M.; Albertini, D.F.; Segars, J.H. Biomechanics and mechanical signaling in the ovary: A systematic review. J. Assist. Reprod. Genet. 2018, 35, 1135–1148. [Google Scholar] [CrossRef] [PubMed]
- Maher, J.Y.; Islam, S.; Yin, O.; Brennan, J.; Gough, E.; Driggers, P.; Segars, J. The role of Hippo pathway signaling and A-kinase anchoring protein 13 in primordial follicle activation and inhibition. F&S Sci. 2022, 3, 118–129. [Google Scholar]
- Devos, M.; Grosbois, J.; Demeestere, I. Interaction between PI3K/AKT and Hippo pathways during in vitro follicular activation and response to fragmentation and chemotherapy exposure using a mouse immature ovary model. Biol. Reprod. 2020, 102, 717–729. [Google Scholar] [CrossRef] [PubMed]
- Hu, L.; Su, T.; Luo, R.; Zheng, Y.; Huang, J.; Zhong, Z.; Nie, J.; Zheng, L. Hippo pathway functions as a downstream effector of AKT signaling to regulate the activation of primordial follicles in mice. J. Cell. Physiol. 2019, 234, 1578–1587. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Risal, S.; Gorre, N.; Busayavalasa, K.; Li, X.; Shen, Y.; Bosbach, B.; Brännström, M.; Liu, K. Somatic Cells Initiate Primordial Follicle Activation and Govern the Development of Dormant Oocytes in Mice. Curr. Biol. 2014, 24, 2501–2508. [Google Scholar] [CrossRef]
- Hannezo, E.; Heisenberg, C.-P. Mechanochemical Feedback Loops in Development and Disease. Cell 2019, 178, 12–25. [Google Scholar] [CrossRef]
- Fabregues, F.; Ferreri, J.; Calafell, J.; Moreno, V.; Borrás, A.; Manau, D.; Carmona, F. Pregnancy after drug-free in vitro activation of follicles and fresh tissue autotransplantation in primary ovarian insufficiency patient: A case report and literature review. J. Ovarian Res. 2018, 11, 76. [Google Scholar] [CrossRef]
- Ferreri, J.; Fàbregues, F.; Calafell, J.M.; Solernou, R.; Borrás, A.; Saco, A.; Manau, D.; Carmona, F. Drug-free in-vitro activation of follicles and fresh tissue autotransplantation as a therapeutic option in patients with primary ovarian insufficiency. Reprod. Biomed. Online 2020, 40, 254–260. [Google Scholar] [CrossRef]
- Cheng, Y.; Feng, Y.; Jansson, L.; Sato, Y.; Deguchi, M.; Kawamura, K.; Hsueh, A.J. Actin polymerization-enhancing drugs promote ovarian follicle growth mediated by the Hippo signaling effector YAP. FASEB J. 2015, 29, 2423–2430. [Google Scholar] [CrossRef] [PubMed]
- Díaz-García, C.; Herraiz, S.; Pamplona, L.; Subirá, J.; Soriano, M.J.; Simon, C.; Seli, E.; Pellicer, A. Follicular activation in women previously diagnosed with poor ovarian response: A randomized, controlled trial. Fertil. Steril. 2022, 117, 747–755. [Google Scholar] [CrossRef] [PubMed]
- Lunding, S.A.; Pors, S.E.; Kristensen, S.G.; Landersoe, S.K.; Jeppesen, J.V.; Flachs, E.M.; Pinborg, A.; Macklon, K.T.; Pedersen, A.T.; Andersen, C.Y.; et al. Biopsying, fragmentation and autotransplantation of fresh ovarian cortical tissue in infertile women with diminished ovarian reserve. Hum. Reprod. 2019, 34, 1924–1936. [Google Scholar] [CrossRef] [PubMed]
- Lunding, S.A.; Andersen, A.N.; Hardardottir, L.; Olesen, H.; Kristensen, S.G.; Andersen, C.Y.; Pors, S.E. Hippo signaling, actin polymerization, and follicle activation in fragmented human ovarian cortex. Mol. Reprod. Dev. 2020, 87, 711–719. [Google Scholar] [CrossRef] [PubMed]
- Gavish, Z.; Gil Peer, G.; Hadassa, R.; Yoram, C.; Meirow, D. Follicle activation and ‘burn-out’ contribute to post-transplantation follicle loss in ovarian tissue grafts: The effect of graft thickness. Hum. Reprod. 2014, 29, 989–996. [Google Scholar] [CrossRef] [PubMed]
- Roness, H.; Meirow, D. Fertility Preservation: Follicle reserve loss in ovarian tissue transplantation. Reproduction 2019, 158, F35–F44. [Google Scholar] [CrossRef] [PubMed]
- Umehara, T.; Urabe, N.; Obata, T.; Yamaguchi, T.; Tanaka, A.; Shimada, M. Cutting the ovarian surface improves the responsiveness to exogenous hormonal treatment in aged mice. Reprod. Med. Biol. 2020, 19, 415–424. [Google Scholar] [CrossRef]
- Britsch, S. The neuregulin-I/ErbB signaling system in development and disease. Adv. Anat. Embryol. Cell Biol. 2007, 190, 1–65. [Google Scholar]
- Agarwal, A.; Zhang, M.; Trembak-Duff, I.; Unterbarnscheidt, T.; Radyushkin, K.; Dibaj, P. Dysregulated expression of neuregulin-1 by cortical pyramidal neurons disrupts synaptic plasticity. Cell Rep. 2014, 8, 1130–1145. [Google Scholar] [CrossRef]
- Banerjee, S.; Mishra, S.; Xu, W.; Thompson, W.E.; Chowdhury, I. Neuregulin-1 signaling regulates cytokines and chemokines expression and secretion in granulosa cell. J. Ovarian Res. 2022, 15, 86. [Google Scholar] [CrossRef]
- Kawashima, I.; Umehara, T.; Noma, N.; Kawai, T.; Shitanaka, M.; Richards, J.S. Targeted disruption of Nrg1 in granulosa cells alters the temporal progression of oocyte maturation. Mol. Endocrinol. 2014, 28, 706–721. [Google Scholar] [CrossRef] [PubMed]
- Umehara, T.; Kawai, T.; Kawashima, I.; Tanaka, K.; Okuda, S.; Kitasaka, H. The acceleration of reproductive aging in Nrg1 (flox/flox); Cyp19-Cre female mice. Aging Cell 2017, 16, 1288–1299. [Google Scholar] [CrossRef]
- Pan, H.A.; Wu, M.H.; Cheng, Y.C.; Wu, L.H.; Chang, F.M. Quantification of ovarian stromal Doppler signals in poor responders undergoing in vitro fertilization with three-dimensional power Doppler ultrasonography. Am. J. Obstet. Gynecol. 2004, 190, 338–344. [Google Scholar] [CrossRef] [PubMed]
- Feng, Y.; Cui, P.; Lu, X.; Hsueh, B.; Billig, F.M.; Yanez, L.Z.; Tomer, R.; Boerboom, D.; Carmeliet, P.; Deisseroth, K.; et al. CLARITY reveals dynamics of ovarian follicular architecture and vasculature in three-dimensions. Sci Rep. 2017, 7, 44810. [Google Scholar] [CrossRef]
- Amin, A.F.; Abd el-Aal, D.E.; Darwish, A.M.; Meki, A.R. Evaluation of the impact of laparoscopic ovarian drilling on Doppler indices of ovarian stromal blood flow, serum vascular endothelial growth factor, and insulin-like growth factor-1 in women with polycystic ovary syndrome. Fertil. Steril. 2003, 79, 938–941. [Google Scholar] [CrossRef] [PubMed]
- Mammoto, T.; Mammoto, A.; Ingber, D.E. Mechanobiology and Developmental Control. Annu. Rev. Cell Dev. Biol. 2013, 29, 27–61. [Google Scholar] [CrossRef] [PubMed]
- Evans, J.P.; Leppert, P.C. “Feeling the force” in reproduction: Mechanotransduction in reproductive processes. Connect. Tissue Res. 2016, 57, 236–244. [Google Scholar] [CrossRef]
- Merle, T.; Farge, E. Trans-scale mechanotransductive cascade of biochemical and biomechanical patterning in embryonic development: The light side of the force. Curr. Opin. Cell Biol. 2018, 55, 111–118. [Google Scholar] [CrossRef]
- Kreeger, P.K.; Deck, J.W.; Woodruff, T.K.; Shea, L.D. The in vitro regulation of ovarian follicle development using alginate-extracellular matrix gels. Biomaterials 2006, 27, 714–723. [Google Scholar] [CrossRef]
- Parandakh, A.; Anbarlou, A.; Tafazzoli-Shadpour, M.; Ardeshirylajimi, A.; Khani, M.-M. Substrate topography interacts with substrate stiffness and culture time to regulate mechanical properties and smooth muscle differentiation of mesenchymal stem cells. Colloids Surf. B Biointerfaces 2019, 173, 194–201. [Google Scholar] [CrossRef]
- Matsuzaki, S. Mechanobiology of the female reproductive system. Reprod. Med. Biol. 2021, 20, 371–401. [Google Scholar] [CrossRef] [PubMed]
- Biswas, A.; Ng, B.H.; Prabhakaran, V.S.; Chan, C.J. Squeezing the eggs to grow: The mechanobiology of mammalian folliculogenesis. Front. Cell Dev. Biol. 2022, 10, 1038107. [Google Scholar] [CrossRef] [PubMed]
- Heeren, A.M.; van Iperen, L.; Klootwijk, D.B.; Bernardo, A.d.M.; Roost, M.S.; Fernandes, M.M.G.; Louwe, L.A.; Hilders, C.G.; Helmerhorst, F.M.; van der Westerlaken, L.A.J.; et al. Development of the follicular basement membrane during human gametogenesis and early folliculogenesis. BMC Dev. Biol. 2015, 15, 4. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.Y.; Nair, M.G. Macrophages in wound healing: Activation and plasticity. Immunol. Cell Biol. 2019, 97, 258–267. [Google Scholar] [CrossRef] [PubMed]
- Hornick, J.; Duncan, F.; Shea, L.; Woodruff, T. Isolated primate primordial follicles require a rigid physical environment to survive and grow in vitro. Hum. Reprod. 2012, 27, 1801–1810. [Google Scholar] [CrossRef] [PubMed]
- Woodruff, T.K.; Shea, L.D. A new hypothesis regarding ovarian follicle development: Ovarian rigidity as a regulator of selection and health. J. Assist. Reprod. Genet. 2011, 28, 3–6. [Google Scholar] [CrossRef] [PubMed]
- Gargus, E.S.; Jakubowski, K.L.; Arenas, G.A.; Miller, S.J.; Lee, S.S.M.; Woodruff, T.K. Ultrasound Shear Wave Velocity Varies Across Anatomical Region in Ex Vivo Bovine Ovaries. Tissue Eng. Part A 2020, 26, 720–732. [Google Scholar] [CrossRef]
- Umehara, T.; Winstanley, Y.E.; Andreas, E.; Morimoto, A.; Williams, E.J.; Smith, K.M.; Carroll, J.; Febbraio, M.A.; Shimada, M.; Russell, D.L.; et al. Female reproductive life span is extended by targeted removal of fibrotic collagen from the mouse ovary. Sci. Adv. 2022, 8, eabn4564. [Google Scholar] [CrossRef]
- Kreeger, P.K.; Woodruff, T.K.; Shea, L.D. Murine granulosa cell morphology and function are regulated by a synthetic Arg–Gly–Asp matrix. Mol. Cell. Endocrinol. 2003, 205, 1–10. [Google Scholar] [CrossRef]
- Berkholtz, C.B.; Shea, L.D.; Woodruff, T.K. Extracellular matrix functions in follicle maturation. Semin. Reprod. Med. 2006, 24, 262–269. [Google Scholar] [CrossRef]
- Zalewski, A.; Cecchini, E.L.; Deroo, B.J. Expression of extracellular matrix components is disrupted in the immature and adult estrogen receptor beta-null mouse ovary. PLoS ONE 2012, 7, e29937. [Google Scholar] [CrossRef] [PubMed]
- Henning, N.F.; Jakus, A.E.; Laronda, M.M. Building Organs Using Tissue-Specific Microenvironments: Perspectives from a Bioprosthetic Ovary. Trends Biotechnol. 2021, 39, 824–837. [Google Scholar] [CrossRef] [PubMed]
- Wood, C.D.; Vijayvergia, M.; Miller, F.H.; Carroll, T.; Fasanati, C.; Shea, L.D.; Brinson, L.C.; Woodruff, T.K. Multi-modal magnetic resonance elastography for noninvasive assessment of ovarian tissue rigidity in vivo. Acta Biomater. 2015, 13, 295–300. [Google Scholar] [CrossRef] [PubMed]
- Briley, S.M.; Jasti, S.; McCracken, J.M.; Hornick, J.E.; Fegley, B.; Pritchard, M.T. Reproductive age-associated fibrosis in the stroma of the mammalian ovary. Reproduction 2016, 152, 245–260. [Google Scholar] [CrossRef] [PubMed]
- Chan, C.J.; Heisenberg, C.-P.; Hiiragi, T. Coordination of Morphogenesis and Cell-Fate Specification in Development. Curr. Biol. 2017, 27, R1024–R1035. [Google Scholar] [CrossRef] [PubMed]
- Chang, H.-M.; Wu, H.-C.; Sun, Z.-G.; Lian, F.; Leung, P.C.K. Neurotrophins and glial cell line-derived neurotrophic factor in the ovary: Physiological and pathophysiological implications. Hum. Reprod. Update 2019, 25, 224–242. [Google Scholar] [CrossRef] [PubMed]
- He, Y.; Peng, X.; Wu, T.; Yang, W.; Liu, W.; Zhang, J.; Su, Y.; Kong, F.; Dou, X.; Li, J. Restricting the induction of NGF in ovarian stroma engenders selective follicular activation through the mTOR signaling pathway. Cell Death Dis. 2017, 8, e2817. [Google Scholar] [CrossRef]
- Streiter, S.; Fisch, B.; Sabbah, B.; Ao, A.; Abir, R. The importance of neuronal growth factors in the ovary. Mol. Hum. Reprod. 2016, 22, 3–17. [Google Scholar] [CrossRef]
- De Roo, C.; Lierman, S.; Tilleman, K.; De Sutter, P. In-vitro fragmentation of ovarian tissue activates primordial follicles through the Hippo pathway. Hum. Reprod. Open 2020, 2020, hoaa048. [Google Scholar] [CrossRef]
- Berkholtz, C.B.; Lai, B.E.; Woodruff, T.K.; Shea, L.D. Distribution of extracellular matrix proteins type I collagen, type IV collagen, fibronectin, and laminin in mouse folliculogenesis. Histochem. 2006, 126, 583–592. [Google Scholar] [CrossRef]
- Abu Hashim, H.; Foda, O.; El Rakhawy, M. Unilateral or bilateral laparoscopic ovarian drilling in polycystic ovary syndrome: A meta-analysis of randomized trials. Arch. Gynecol. Obstet. 2018, 297, 859–870. [Google Scholar] [CrossRef] [PubMed]
- Ostrzenski, A. Endoscopic carbon dioxide laser ovarian wedge resection in resistant polycystic ovarian disease. Int. J. Fertil. 1992, 37, 295–299. [Google Scholar] [PubMed]
- Seyam, E.; Hefzy, E. Tumor necrosis factor alpha versus LH and androstendione as a reliable predictor of spontaneous ovulation after laparoscopic ovarian drilling for women with clomiphene citrate resistance polycystic ovarian disease. Eur. J. Obstet. Gynecol. Reprod. Biol. 2018, 222, 126–133. [Google Scholar] [CrossRef] [PubMed]
- Amer, S.A.; El Shamy, T.T.; James, C.; Yosef, A.H.; Mohamed, A.A. The impact of laparoscopic ovarian drilling on AMH and ovarian reserve: A meta-analysis. Reproduction 2017, 154, R13–R21. [Google Scholar] [CrossRef] [PubMed]
- De Roo, C.; Tilleman, K.; Vercruysse, C.; Declercq, H.; T’sjoen, G.; Weyers, S.; De Sutter, P. Texture profile analysis reveals a stiffer ovarian cortex after testosterone therapy: A pilot study. J. Assist. Reprod. Genet. 2019, 36, 1837–1843. [Google Scholar] [CrossRef] [PubMed]
- Tozawa, H.; Brännström, M.; Petrucco, O.; Walker, S.; Chambers, H.; Pascoe, V.; Norman, R. Immunology: Distribution of leukocyte subtypes in the sheep ovary after laser drilling. Hum. Reprod. 1995, 10, 544–550. [Google Scholar] [CrossRef] [PubMed]
- Giampaolino, P.; Morra, I.; Della Corte, L.; Sparice, S.; Di Carlo, C.; Nappi, C.; Bifulco, G. Serum anti-Mullerian hormone levels after ovarian drilling for the second-line treatment of polycystic ovary syndrome: A pilot-randomized study comparing laparoscopy and transvaginal hydrolaparoscopy. Gynecol. Endocrinol. 2017, 33, 26–29. [Google Scholar] [CrossRef] [PubMed]
- Shimamoto, S.; Nishimura, Y.; Nagamatsu, G.; Hamada, N.; Kita, H.; Hikabe, O.; Hamazaki, N.; Hayashi, K. Hypoxia induces the dormant state in oocytes through expression of Foxo3. Proc. Natl. Acad. Sci. USA 2019, 116, 12321–12326. [Google Scholar] [CrossRef]
- Nagamatsu, G. Regulation of primordial follicle formation, dormancy, and activation in mice. J. Reprod. Dev. 2021, 67, 189–195. [Google Scholar] [CrossRef]
- Aiken, C.; Tarry-Adkins, J.L.; Spiroski, A.-M.; Nuzzo, A.M.; Ashmore, T.J.; Rolfo, A.; Sutherland, M.J.; Camm, E.J.; Giussani, D.; Ozanne, S. Chronic gestational hypoxia accelerates ovarian aging and lowers ovarian reserve in next-generation adult rats. FASEB J. 2019, 33, 7758–7766. [Google Scholar] [CrossRef]
- Kawai, T.; Shimada, M. Pretreatment of ovaries with collagenase before vitrification keeps the ovarian reserve by maintaining cell-cell adhesion integrity in ovarian follicles. Sci. Rep. 2020, 10, 6841. [Google Scholar] [CrossRef] [PubMed]
- Tomaszewski, C.E.; DiLillo, K.M.; Baker, B.M.; Arnold, K.B.; Shikanov, A. Sequestered cell-secreted extracellular matrix proteins improve murine folliculogenesis and oocyte maturation for fertility preservation. Acta Biomater. 2021, 132, 313–324. [Google Scholar] [CrossRef] [PubMed]
- Tagler, D.; Makanji, Y.; Tu, T.; Bernabé, B.P.; Lee, R.; Zhu, J.; Kniazeva, E.; Hornick, J.E.; Woodruff, T.K.; Shea, L.D. Promoting extracellular matrix remodeling via ascorbic acid enhances the survival of primary ovarian follicles encapsulated in alginate hydrogels. Biotechnol. Bioeng. 2014, 111, 1417–1429. [Google Scholar] [CrossRef] [PubMed]
- Man, L.; Park, L.; Bodine, R.; Ginsberg, M.; Zaninovic, N.; Man, O.A.; Schattman, G.; Rosenwaks, Z.; James, D. Engineered endothelium provides angiogenic and paracrine stimulus to grafted human ovarian tissue. Sci. Rep. 2017, 7, 8203. [Google Scholar] [CrossRef] [PubMed]
- Man, L.; Park, L.; Bodine, R.; Ginsberg, M.; Zaninovic, N.; Schattman, G.; Schwartz, R.E.; Rosenwaks, Z.; James, D. Co-Transplantation of Human Ovarian Tissue with Engineered Endothelial Cells: A Cell-based Strategy Combining Accelerated Perfusion with Direct Paracrine Delivery. J. Vis. Exp. 2018, 135, e57472. [Google Scholar]
- Caley, M.P.; Martins, V.L.; O’Toole, E.A. Metalloproteinases and Wound Healing. Adv. Wound Care 2015, 4, 225–234. [Google Scholar] [CrossRef] [PubMed]
- Sridharan, R.; Cavanagh, B.; Cameron, A.R.; Kelly, D.J.; O’Brien, F.J. Material stiffness influences the polarization state, function and migration mode of macrophages. Acta Biomater. 2019, 89, 47–59. [Google Scholar] [CrossRef] [PubMed]
- Irving-Rodgers, H.F.; Rodgers, R.J. Extracellular Matrix of the Developing Ovarian Follicle. Semin. Reprod. Med. 2006, 24, 195–203. [Google Scholar] [CrossRef]
- Tingen, C.M.; Kiesewetter, S.E.; Jozefik, J.; Thomas, C.; Tagler, D.; Shea, L.; Woodruff, T.K. A macrophage and theca cell-enriched stromal cell population influences growth and survival of immature murine follicles in vitro. Reproduction 2011, 141, 809–820. [Google Scholar] [CrossRef]
- Diaz, A.A.; Kubo, H.; Handa, N.; Hanna, M.T.; Laronda, M.M. A Systematic Review of Ovarian Tissue Transplantation Outcomes by Ovarian Tissue Processing Size for Cryopreservation. Front. Endocrinol. 2022, 13, 918899. [Google Scholar] [CrossRef]
- McGee, E.A.; Hsueh, A.J.W. Initial and Cyclic Recruitment of Ovarian Follicles. Endocr. Rev. 2000, 21, 200–214. [Google Scholar]
- Nelson, S.; Telfer, E.; Anderson, R. The ageing ovary and uterus: New biological insights. Hum. Reprod. Update 2013, 19, 67–83. [Google Scholar] [CrossRef]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chang, C.L. Facilitation of Ovarian Response by Mechanical Force—Latest Insight on Fertility Improvement in Women with Poor Ovarian Response or Primary Ovarian Insufficiency. Int. J. Mol. Sci. 2023, 24, 14751. https://doi.org/10.3390/ijms241914751
Chang CL. Facilitation of Ovarian Response by Mechanical Force—Latest Insight on Fertility Improvement in Women with Poor Ovarian Response or Primary Ovarian Insufficiency. International Journal of Molecular Sciences. 2023; 24(19):14751. https://doi.org/10.3390/ijms241914751
Chicago/Turabian StyleChang, Chia Lin. 2023. "Facilitation of Ovarian Response by Mechanical Force—Latest Insight on Fertility Improvement in Women with Poor Ovarian Response or Primary Ovarian Insufficiency" International Journal of Molecular Sciences 24, no. 19: 14751. https://doi.org/10.3390/ijms241914751