


Technical Implications of the Chicken Embryo Chorioallantoic Membrane Assay to Elucidate Neuroblastoma Biology
 ,
,  ,
,
Abstract
:1. Introduction
2. Structure and Origin of the CAM
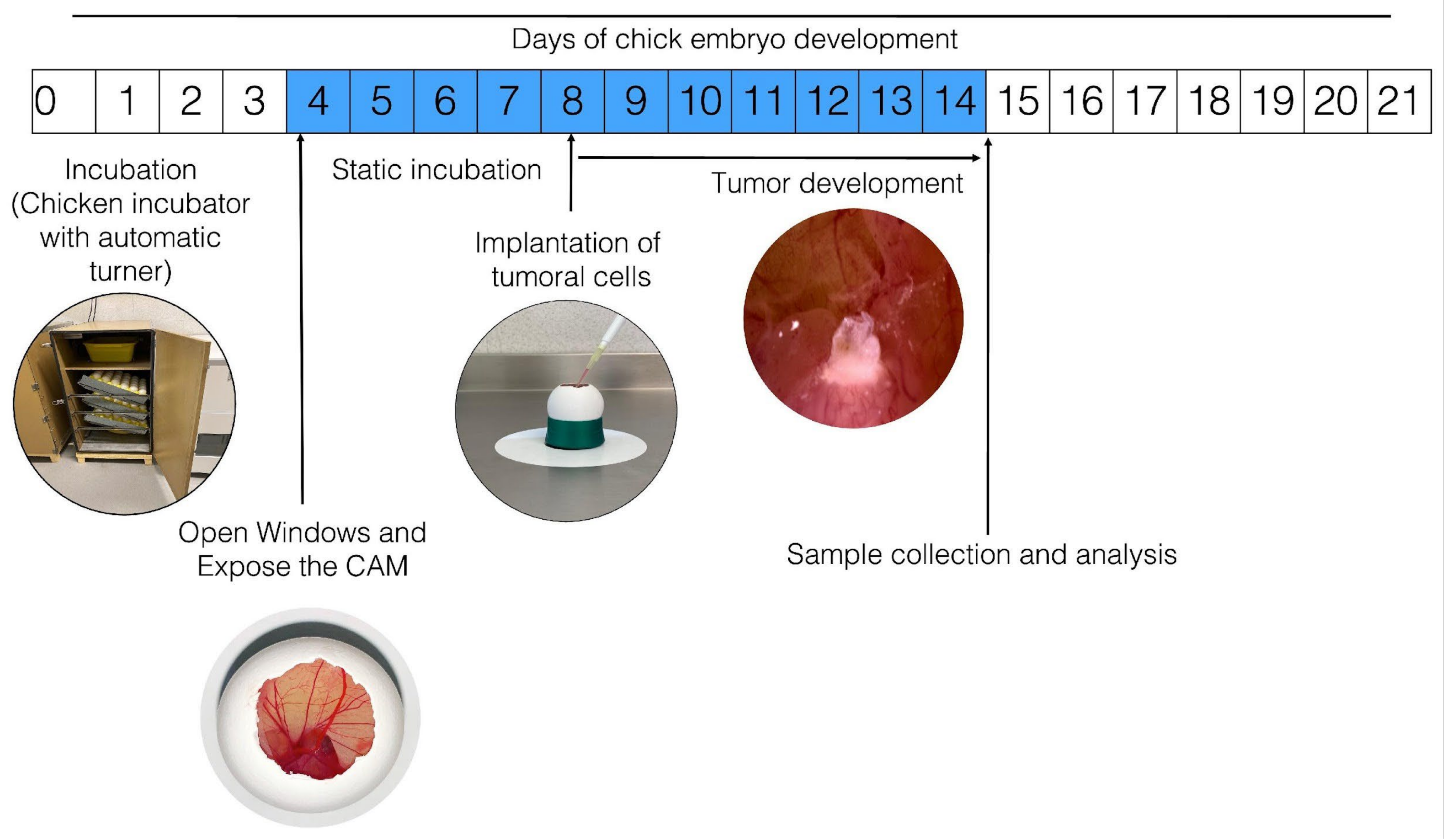
3. Technical Characteristics of the Use of CAM for In Ovo Tumor Generation
4. In Ovo and Ex Ovo CAM Assay
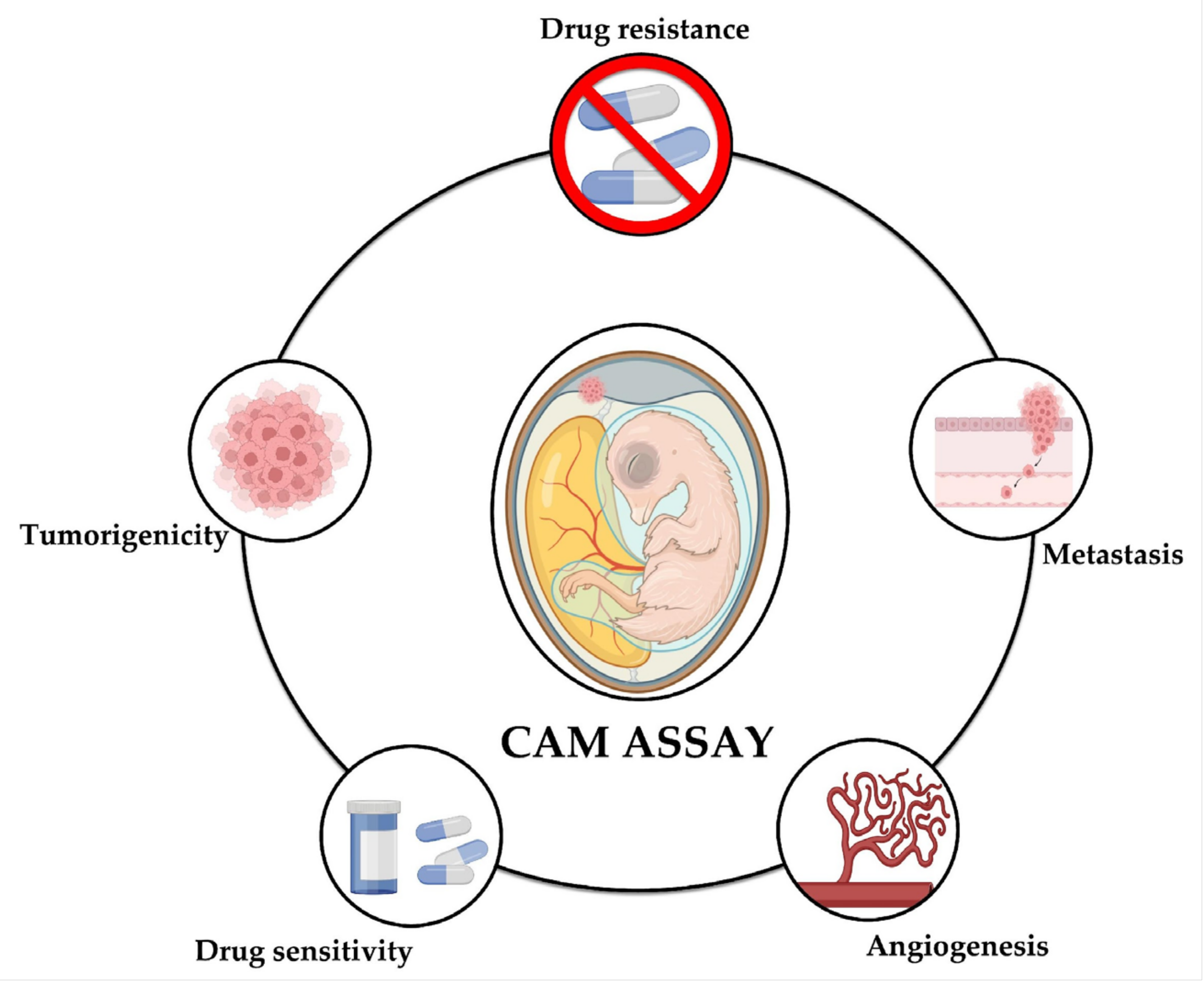
5. CAM Assay as an Alternative to the Use of Experimental Animals
6. The Importance of Studying NB
7. CAM and NB
8. Using CAM Assay to Study Metastasis and Antimetastatic Drugs
9. Using the CAM Assay to Study Chemoresistance and Molecules with Antitumor Effect in NB
10. Studying Angiogenesis in NB Using the CAM Assay

{kind=link}
{kind=link}
| Cells Used/TUMOR Type | Cell Number | Inoculation Technique | Target | Reference |
|---|---|---|---|---|
| Studies focused on metastasis and antimetastatic drugs | ||||
| UKF-NB-4 cell line | Ex ovo: 25 µL with 5 × 104 cells in serum-free medium (Iscove’s modified Dulbecco’s medium or IMDM). In ovo: 25 µL with 1 × 106 cells in serum-free medium (Iscove’s modified Dulbecco’s medium or IMDM). | Tumor was induced at day 10 and allowed to develop for 4 days. Tumor was induced on day 10. The tumor was allowed to develop for 6 days. | Ex ovo: To study the antiproliferative effect of CDDP (cisplatin) and Elli (ellipticin) as well as to evaluate their potential as inhibitors of cell extravasation into blood vessels. In ovo: Studying the efficiency of CDDP and Elli to inhibit metastasis of neuroblastoma (NB) cells to chick organs. | Merlos et al., 2021 [67] |
| SK-N-AS | 1 × 106 cells per microliter in serum-free medium and 2 microliters were inoculated. | Inoculation into chicken CAM was performed at day 7 cells were plated into CAM after a small laceration. Eggs were incubated until E14 | Evaluating the chick embryo CAM assay for the study of metastasis. | (A. Herrmann et al., 2015; Herrmann et al., 2016) [68,69] |
| SK-N-AS cells | 1 × 106 cells resuspended in 2–10 μL of serum-free minimal essential medium. | Cells were preconditioned in 1 or 21% O2 for 3 days, tumors were implanted at day 7 of incubation in the chick embryo CAM. Tumors were studied at day 14 postinoculation. | To elucidate the effect of hypoxic preconditioning on the metastatic phenotype of NB cells. | (Al-Mutawa et al., 2018) [70] |
| +SK-N-AS MPIO-Labeled- SK-N-AS GFP-Labeled- SK-N-AS | 1 × 106 cells resuspended in 2–10 μL of serum-free minimal essential medium. | Cells were implanted in the CAM on day 7. Before cell inoculation the membrane was carefully lacerated. Tumors were studied at days 11 and 14. | To determine the effectiveness of Magnetic Resonance Imaging (MRI) for the evaluation of tumor development and metastasis in a chick embryo model. | (Herrmann et al., 2018) [71] |
| SK-N-SH | Ten million SK-N-SH cells shNEO1, shNTN4 or shSCR. | Fertilized chicken eggs were incubated and on the second day of incubation (E2), 2 mL of albumin was extracted from the egg. On the tenth day of incubation (E10), cells were plated on the developing CAM. On incubation day 17 (E17). | To provide evidence for novel roles of the NTN4/NEO1 complex in the in vivo migration, survival, and metastasis of NB cells. | (Villanueva et al., 2017) [72] |
| Studies focused on chemoresistance and molecules with antitumor effect | ||||
| UKF-NB-4 UKF-NB-4CDDP | 1 × 105 cells were inoculated. | Fertilized hen eggs were incubated for 10 days following the cell inoculation, the tumor was allowed to grow for 5 days. | To investigate the impact of upregulation of human metallothionein 3 (hMT3) in NB cells and its contribution in resistance to cisplatin treatment. | (Rodrigo et al., 2021) [79] |
| SK-N-AS BE(2)C | A quantity of 2 × 106 GFP-labeled cells was inoculated. | Cells were inoculated on day 4 and incubated until day 14. | To investigate the potential of Palbociclib (CDK 4/6 inhibitor) and RO-3306 (CDK1 inhibitor) on NB cell differentiation, tumor progression and metastasis. | (Swadi et al., 2019) [80] |
| IMR32 and BE2C | 2 × 106 in 5 μL of DMEM or Matrigel was used. | Cells were implanted into the CAM on day 7 of embryo incubation, but before laying the cells the authors report that it is necessary to lacerate the CAM and add 5 μL of trypsin 0.05%–0.5 mM EDTA. The tumors were analyzed on day 14. | Optimize the CAM assay for the study of drugs with antitumor activity. | (Swadi et al., 2018) [81] |
| UKF-NB-3 | 5 × 106 cells were resuspended in 30 μL of ECM and then implanted in CAM. | Cells were implanted on day 11 of embryo development. Eggs were incubated for another 3 days to allow formation of a distinct tumor mass. On day 14, a small silicone ring was placed around the tumor mass. On day 18, the tumors were sampled with the surrounding CAM. | To evaluate the antitumor effect of flubendazole. | (Michaelis et al., 2015) [82] |
| Studies focused on angiogenesis | ||||
| LAN-5 and GI-LI-N | 3 µL suspension 6 × 106 cells per ml of RPMI-1640 medium absorbedon 1 mm3 gelatin sponges | Tumor was induced at day 8 and processed at day 12. | Investigated 2 human NB cell lines, LAN-5 and GI-LI-N, for the capacity to induce extracellular matrix-degrading enzymes and for their angiogenic potential both in vitro and in vivo. | (Ribatti et al., 1998) [87] |
| HTLA-230, GI-LI-N, ACN and SH-SY5Y | Fresh in sterile RPMI-1640 (1–2 mm3 size). These fragments were grafted onto the CAM. Biopsy fragments (1–2 mm3 size) obtained from xenografts derived from four cell lines injected into nude mice. | On day 3 of incubation, a square window was opened in the eggshell after removing 2–3 mL of albumin to separate the developing CAM from the shell. On day 8, implantation was performed and on day 12, sample analysis was performed. | To evaluate the role of MYCN in the regulation of angiogenesis in NB. | (Ribatti et al., 2002) [88] |
| STA-NB15 | 2 × 106 cells were transplanted. The cells were resuspended in 50 μL of collagen dissolved in cell culture medium and the mixture was solidified for 30 min at 37 °C. | On day 3 of embryonic development, 5–7 mL of egg white was removed and a 2 × 2 cm window was cut in the eggshell. On day 10, these cells were inoculated. The living embryos were incubated for another 5 days. | To understand the role of FOXO 3 as a promoter of cell migration and invasion. | (Hagenbuchner et al., 2016) [89] |
| Kelly | 5 × 106 Kelly cells were inoculated onto 15 μL Matrigel. | After 10 days, an opening was made in the eggshell and a plastic ring was placed in the CAM. Tumors were excised on day 17 of incubation. | To analyze the antitumor effect of Ang-2 upregulation in human NB. | (Klingenberg et al., 2014) [90] |
| UKF-NB-3 | 106 cells were resuspended in 50 μL of medium. | The cells were inoculated onto the chicken CAM at day 8. Vessel formation was evaluated at day 12. | To analyze the effect of chemoresistance on tumor angiogenesis. | (Michaelis et al., 2009) [91] |
| IMR-32 | Silicon rings were adsorbed with 2 μL of cell suspension of NB cells. | On day 3 of incubation a window was opened in the eggshell and 2–3 mL of albumen was removed. The angiogenic response was evaluated on day 12 of incubation after the implants. | To generate an alternative model for the study of NB angiogenesis. | (Mangieri et al., 2009) [92] |
| GI-LI-N | Cells were injected in the capsule of the left adrenal gland of mice. From these tumors a biopsy was taken and placed in the CAM. | On day 8 of incubation 2 mm3 biopsy fragments from xenograft tumors previously induced in mice, were placed onto the CAM. The CAM was analyzed at day 12. | To evaluate the antitumor effect of the administration of two combined drugs: bortezomib and fenretinide. | (Pegman et al., 2009) [93] |
| GI-LI-N | 1 × 106 cells were inoculated in the capsule of the left adrenal gland, of nude mice. A biopsy fragments taken from murine xenografts of NB. | On day 8 of incubation, biopsy fragments from xenografts derived from NB, lung, and ovarian carcinoma cells injected in athymic mice were grafted onto the CAM encapsulated in either Caelyx or in TVT-DOX. Vasculature was analyzed at day 12. | To evaluate the differences in the anti-angiogenic activity between TVT-DOX targeted liposomal doxorubicin (TVT-DOX) and Caelyx (untargeted liposomes). | (Pastorino et al., 2008) [94] |
| Fresh biopsy specimens from patients | 1- to 2-mm3 fragments fresh biopsy specimens. | Biopsy fragments from patients with NB were collected under sterile conditions in RPMI 1640. This biopsy was placed inside CAM at day 8 of incubation. Analysis was performed on day 12. | To study the antitumor activity of vinblastine and rapamycin against human NB. | (Marimpietri et al., 2007) [95] |
| HTLA-230 | 1–2 mm3 of mouse tumor biopsy fragments Fresh biopsies from NB patients in RPMI 1640 (1 to 2 mm3 fragments) | On day 3, a square window was opened in the cover and 2–3 mL of albumen was removed. The window was sealed, and the eggs were returned to the incubator. On day 8, the eggs were implanted with biopsies and left until day 12. | To evaluate the synergistic effect of low doses of vinblastine (VBL) and rapamycin (RAP) on anti-angiogenesis in NB (NB). | (Marimpietri et al., 2005) [96] |
| NB cell line | The cell lines were inoculated into mice to generate tumors. Tumors were frozen and crushed in RPMI 1640 (1 to 2 mm3 fragments) | Tumor fragments were grafted into the chicken embryo CAM at day 8 of incubation. The analysis was carried out on day 12. | To evaluate the angiogenesis inhibitory effect of IFN-γ in NB-derived cells. | (Ribatti et al., 2006) [97] |
| HTLA-230 | Grafting 1–2-mm3 fragments of xenograft tumors derived from cells injected into nude mice onto the CAM. Grafting fresh biopsy specimens from patients with NB in RPMI 1640 (1- to 2-mm3 fragments) | At day 8 of incubation, implantation of tumor fragments generated from mice was performed as tumor fragments obtained from patients were treated with PBS (control) or were treated with 20 nM bortezomib. | To analyze the effect of bortezomib on growth, apoptosis, and angiogenesis of human NB cells. | (Brignole et al., 2006) [98] |
11. Conclusions and Perspectives
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Chen, L.; Wang, S.; Feng, Y.; Zhang, J.; Du, Y.; Zhang, J.; Van Ongeval, C.; Ni, Y.; Li, Y. Utilisation of Chick Embryo Chorioallantoic Membrane as a Model Platform for Imaging-Navigated Biomedical Research. Cells 2021, 10, 463. [Google Scholar] [CrossRef]
- Victorelli, F.D.; Cardoso, V.M.d.O.; Ferreira, N.N.; Calixto, G.M.F.; Fontana, C.R.; Baltazar, F.; Gremião, M.P.D.; Chorilli, M. Chick embryo chorioallantoic membrane as a suitable in vivo model to evaluate drug delivery systems for cancer treatment: A review. Eur. J. Pharm. Biopharm. 2020, 153, 273–284. [Google Scholar] [CrossRef]
- Ahmed, T.A.E.; Cordeiro, C.M.M.; Elebute, O.; Hincke, M.T. Proteomic Analysis of Chicken Chorioallantoic Membrane (CAM) during Embryonic Development Provides Functional Insight. BioMed Res. Int. 2022, 2022, 7813921. [Google Scholar] [CrossRef] [PubMed]
- Ribatti, D. The chick embryo chorioallantoic membrane (CAM) assay. Reprod. Toxicol. 2017, 70, 97–101. [Google Scholar] [CrossRef] [PubMed]
- Kundeková, B.; Máčajová, M.; Meta, M.; Čavarga, I.; Bilčík, B. Chorioallantoic Membrane Models of Various Avian Species: Differences and Applications. Biology 2021, 10, 301. [Google Scholar] [CrossRef]
- Rous, P.; Murphy, J.B. Tumor implantations in the developing embryo. J. Med. Assoc. 1911, LVI, 742. [Google Scholar] [CrossRef]
- Komatsu, A.; Higashi, Y.; Matsumoto, K. Various CAM tumor models. Enzymes 2019, 46, 37–57. [Google Scholar] [CrossRef] [PubMed]
- Subauste, M.C.; Kupriyanova, T.A.; Conn, E.M.; Ardi, V.C.; Quigley, J.P.; Deryugina, E.I. Evaluation of metastatic and angiogenic potentials of human colon carcinoma cells in chick embryo model systems. Clin. Exp. Metastasis 2009, 26, 1033–1047. [Google Scholar] [CrossRef]
- Ho, B.-Y.; Wu, Y.-M.; Hsu, Y.-W.; Hsu, L.-C.; Kuo, Y.-H.; Chang, K.-J.; Pan, T.-M. Effects of Monascus-Fermented Rice Extract on Malignant Cell—Associated Neovascularization and Intravasation Determined Using the Chicken Embryo Chorioallantoic Membrane Model. Integr. Cancer Ther. 2010, 9, 204–212. [Google Scholar] [CrossRef]
- Kaufman, N.; Kinney, T.D.; Mason, E.J.; Prieto, L.C. Maintenance of human neoplasm on the chick chorioallantoic membrane. Am. J. Pathol. 1956, 32, 271–285. [Google Scholar]
- Lugassy, C.; Torres-Muñoz, J.E.; Kleinman, H.K.; Ghanem, G.; Vernon, S.; Barnhill, R.L. Overexpression of malignancy-associated laminins and laminin receptors by angiotropic human melanoma cells in a chick chorioallantoic membrane model. J. Cutan. Pathol. 2009, 36, 1237–1243. [Google Scholar] [CrossRef]
- Balčiūnienė, N.; Tamašauskas, A.; Valančiūtė, A.; Deltuva, V.; Vaitiekaitis, G.; Gudinavičienė, I.; Weis, J.; Von Keyserlingk, D.G. Histology of human glioblastoma transplanted on chicken chorioallantoic membrane. Medicina 2009, 45, 123. [Google Scholar] [CrossRef] [PubMed]
- Strojnik, T.; Kavalar, R.; Barone, T.A.; Plunkett, R.J. Experimental Model and Immunohistochemical Comparison of U87 Human Glioblastoma Cell Xenografts on the Chicken Chorioallantoic Membrane and in Rat Brains. Anticancer. Res. 2010, 30, 4851–4860. [Google Scholar] [PubMed]
- Komatsu, A.; Matsumoto, K.; Yoshimatsu, Y.; Sin, Y.; Kubota, A.; Saito, T.; Mizumoto, A.; Ohashi, S.; Muto, M.; Noguchi, R.; et al. The CAM Model for CIC-DUX4 Sarcoma and Its Potential Use for Precision Medicine. Cells 2021, 10, 2613. [Google Scholar] [CrossRef] [PubMed]
- Sys, G.M.L.; Lapeire, L.; Stevens, N.; Favoreel, H.; Forsyth, R.; Bracke, M.; De Wever, O. The In ovo CAM-assay as a Xenograft Model for Sarcoma. J. Vis. Exp. 2013, 77, e50522. [Google Scholar] [CrossRef]
- Lokman, N.A.; Elder, A.S.F.; Ricciardelli, C.; Oehler, M.K. Chick Chorioallantoic Membrane (CAM) Assay as an In Vivo Model to Study the Effect of Newly Identified Molecules on Ovarian Cancer Invasion and Metastasis. Int. J. Mol. Sci. 2012, 13, 9959–9970. [Google Scholar] [CrossRef] [PubMed]
- Vu, B.T.; Shahin, S.A.; Croissant, J.; Fatieiev, Y.; Matsumoto, K.; Doan, T.L.-H.; Yik, T.; Simargi, S.; Contreras, A.; Ratliff, L.; et al. Chick chorioallantoic membrane assay as an in vivo model to study the effect of nanoparticle-based anticancer drugs in ovarian cancer. Sci. Rep. 2018, 8, 8524. [Google Scholar] [CrossRef]
- Fergelot, P.; Bernhard, J.-C.; Soulet, F.; Kilarski, W.W.; Léon, C.; Courtois, N.; Deminière, C.; Herbert, J.M.J.; Antczak, P.; Falciani, F.; et al. The experimental renal cell carcinoma model in the chick embryo. Angiogenesis 2013, 16, 181–194. [Google Scholar] [CrossRef]
- Ferician, O.; Cimpean, A.M.; Avram, S.; Raica, M. Endostatin Effects on Tumor Cells and Vascular Network of Human Renal Cell Carcinoma Implanted on Chick Embryo Chorioallantoic Membrane. Anticancer. Res. 2015, 35, 6521–6528. [Google Scholar]
- Skowron, M.A.; Sathe, A.; Romano, A.; Hoffmann, M.J.; Schulz, W.A.; van Koeveringe, G.A.; Albers, P.; Nawroth, R.; Niegisch, G. Applying the chicken embryo chorioallantoic membrane assay to study treatment approaches in urothelial carcinoma. Urol. Oncol. 2017, 35, e11–e544. [Google Scholar] [CrossRef]
- Syrovets, T.; Gschwend, J.E.; Büchele, B.; Laumonnier, Y.; Zugmaier, W.; Genze, F.; Simmet, T. Inhibition of IκB Kinase Activity by Acetyl-boswellic Acids Promotes Apoptosis in Androgen-independent PC-3 Prostate Cancer Cells in Vitro and in Vivo. J. Biol. Chem. 2005, 280, 6170–6180. [Google Scholar] [CrossRef]
- Rudy, S.F.; Brenner, J.C.; Harris, J.L.; Liu, J.; Che, J.; Scott, M.V.; Owen, J.H.; Komarck, C.M.; Graham, M.P.; Bellile, E.L.; et al. In vivo Wnt pathway inhibition of human squamous cell carcinoma growth and metastasis in the chick chorioallantoic model. J. Otolaryngol.—Head. Neck Surg. 2016, 45, 26. [Google Scholar] [CrossRef]
- Xiao, X.; Zhou, X.; Ming, H.; Zhang, J.; Huang, G.; Zhang, Z.; Li, P. Chick Chorioallantoic Membrane Assay: A 3D Animal Model for Study of Human Nasopharyngeal Carcinoma. PLoS ONE 2015, 10, e0130935. [Google Scholar] [CrossRef] [PubMed]
- Uloza, V.; Kuzminienė, A.; Šalomskaitė-Davalgienė, S.; Palubinskienė, J.; Balnytė, I.; Ulozienė, I.; Šaferis, V.; Valančiūtė, A. Effect of Laryngeal Squamous Cell Carcinoma Tissue Implantation on the Chick Embryo Chorioallantoic Membrane: Morphometric Measurements and Vascularity. BioMed Res. Int. 2015, 2015, 629754. [Google Scholar] [CrossRef] [PubMed]
- Balke, M.; Neumann, A.; Szuhai, K.; Agelopoulos, K.; August, C.; Gosheger, G.; Hogendoorn, P.C.; Athanasou, N.; Buerger, H.; Hagedorn, M. A short-term in vivo model for giant cell tumor of bone. BMC Cancer 2011, 11, 241. [Google Scholar] [CrossRef] [PubMed]
- Karagianni, F.; Piperi, C.; Casar, B.; de la Fuente-Vivas, D.; García-Gómez, R.; Lampadaki, K.; Pappa, V.; Papadavid, E. Combination of Resminostat with Ruxolitinib Exerts Antitumor Effects in the Chick Embryo Chorioallantoic Membrane Model for Cutaneous T Cell Lymphoma. Cancers 2022, 14, 1070. [Google Scholar] [CrossRef] [PubMed]
- Gronau, S.; Thess, B.; Riechelmann, H.; Fischer, Y.; Schmitt, A.; Schmitt, M. An autologous system for culturing head and neck squamous cell carcinomas for the assessment of cellular therapies on the chorioallantois membrane. Eur. Arch. Otorhinolaryngol. 2006, 263, 308–312. [Google Scholar] [CrossRef] [PubMed]
- Marzullo, A.; Vacca, A.; Roncali, L.; Pollice, L.; Ribatti, D. Angiogenesis in hepatocellular carcinoma: An experimental study in the chick embryo chorioallantoic membrane. Int. J. Oncol. 1998, 13, 17–21. [Google Scholar] [CrossRef] [PubMed]
- Rovithi, M.; Avan, A.; Funel, N.; Leon, L.G.; Gomez, V.E.; Wurdinger, T.; Griffioen, A.W.; Verheul, H.M.W.; Giovannetti, E. Development of bioluminescent chick chorioallantoic membrane (CAM) models for primary pancreatic cancer cells: A platform for drug testing. Sci. Rep. 2017, 7, 44686. [Google Scholar] [CrossRef]
- Sudha, T.; Yalcin, M.; Lin, H.-Y.; Elmetwally, A.M.; Nazeer, T.; Arumugam, T.; Phillips, P.; Mousa, S.A. Suppression of pancreatic cancer by sulfated non-anticoagulant low molecular weight heparin. Cancer Lett. 2014, 350, 25–33. [Google Scholar] [CrossRef]
- Sugiura, Y.; Shimada, H.; Seeger, R.C.; Laug, W.E.; DeClerck, Y.A. Matrix metalloproteinases-2 and -9 are expressed in human neuroblastoma: Contribution of stromal cells to their production and correlation with metastasis. Cancer Res. 1998, 58, 2209–2216. [Google Scholar] [PubMed]
- Dumartin, L.; Quemener, C.; Laklai, H.; Herbert, J.; Bicknell, R.; Bousquet, C.; Pyronnet, S.; Castronovo, V.; Schilling, M.K.; Bikfalvi, A.; et al. Netrin-1 mediates early events in pancreatic adenocarcinoma progression, acting on tumor and endothelial cells. Gastroenterology 2010, 138, 1595–1606. [Google Scholar] [CrossRef] [PubMed]
- Ribatti, D. The chick embryo chorioallantoic membrane in the study of tumor angiogenesis. Rom. J. Morphol. Embryol. 2008, 49, 131–135. [Google Scholar] [PubMed]
- Dünker, N.; Jendrossek, V. Implementation of the Chick Chorioallantoic Membrane (CAM) Model in Radiation Biology and Experimental Radiation Oncology Research. Cancers 2019, 11, 1499. [Google Scholar] [CrossRef]
- Ribatti, D. The chick embryo chorioallantoic membrane (CAM). A multifaceted experimental model. Mech. Dev. 2016, 141, 70–77. [Google Scholar] [CrossRef]
- Ribatti, D. The chick embryo chorioallantoic membrane as a model for tumor biology. Exp. Cell Res. 2014, 328, 314–324. [Google Scholar] [CrossRef]
- Stern, C.D. The chicken; a large model system becomes even larger. Development cell. 2005, 8, 9–17. [Google Scholar] [CrossRef]
- Nowak-Sliwinska, P.; Segura, T.; Iruela-Arispe, M.L. The chicken chorioallantoic membrane model in biology, medicine and bioengineering. Angiogenesis 2014, 17, 779–804. [Google Scholar] [CrossRef]
- Vargas, A.; Zeisser-Labouèbe, M.; Lange, N.; Gurny, R.; Delie, F. The chick embryo and its chorioallantoic membrane (CAM) for the in vivo evaluation of drug delivery systems. Adv. Drug Deliv. Rev. 2007, 59, 1162–1176. [Google Scholar] [CrossRef]
- Tamanoi, F. Recent excitements in the study of the CAM assay. Enzymes 2019, 46, 1–9. [Google Scholar] [CrossRef]
- Chambers, A.F.; Shafir, R.; Ling, V. A model system for studying metastasis using the embryonic chick. Cancer Res. 1982, 42, 4018–4025. [Google Scholar] [PubMed]
- Nowak-Sliwinska, P.; Alitalo, K.; Allen, E.; Anisimov, A.; Aplin, A.C.; Auerbach, R.; Augustin, H.G.; Bates, D.O.; van Beijnum, J.R.; Bender, R.H.F.; et al. Consensus guidelines for the use and interpretation of angiogenesis assays. Angiogenesis 2018, 21, 425–532. [Google Scholar] [CrossRef] [PubMed]
- Naik, M.; Brahma, P.; Dixit, M. A Cost-Effective and Efficient Chick Ex-Ovo CAM Assay Protocol to Assess Angiogenesis. Methods Protoc. 2018, 31, 19. [Google Scholar] [CrossRef]
- Karnofsky, D.A.; Ridgway, L.P.; Patterson, P.A. Tumor transplantation to the chick embryo. Ann. N. Y. Acad. Sci. 1952, 55, 313–329. [Google Scholar] [CrossRef]
- Davison, T.F. The immunologists’ debt to the chicken. Br. Poult. Sci. 2003, 44, 6–21. [Google Scholar] [CrossRef] [PubMed]
- International Chicken Genome Sequencing Consortium. Sequence and comparative analysis of the chicken genome provide unique perspectives on vertebrate evolution. Nature 2004, 432, 695–716. [Google Scholar] [CrossRef]
- Mouse Genome Sequencing Consortium. Initial sequencing and comparative analysis of the mouse genome. Nature 2002, 420, 520–562. [Google Scholar] [CrossRef]
- International Human Genome Sequencing Consortium. Initial sequencing and analysis of the human genome. Nature 2001, 409, 860–921. [Google Scholar] [CrossRef]
- Cohn, S.L.; Pearson, A.D.J.; London, W.B.; Monclair, T.; Ambros, P.F.; Brodeur, G.M.; Faldum, A.; Hero, B.; Iehara, T.; Machin, D.; et al. The International Neuroblastoma Risk Group (INRG) Classification System: An INRG Task Force Report. J. Clin. Oncol. 2009, 27, 289–297. [Google Scholar] [CrossRef]
- Liang, W.H.; Federico, S.M.; London, W.B.; Naranjo, A.; Irwin, M.S.; Volchenboum, S.L.; Cohn, S.L. Tailoring Therapy for Children with Neuroblastoma on the Basis of Risk Group Classification: Past, Present, and Future. JCO Clin. Cancer Inform. 2020, 4, 895–905. [Google Scholar] [CrossRef]
- PDQ Pediatric Treatment Editorial Board. Neuroblastoma Treatment (PDQ®): Patient Version. In PDQ Cancer Information Summaries; National Cancer Institute: Bethesda, MD, USA, 2023. [Google Scholar]
- Chung, C.; Boterberg, T.; Lucas, J.; Panoff, J.; Valteau-Couanet, D.; Hero, B.; Bagatell, R.; Hill-Kayser, C.E. Neuroblastoma. Pediatr. Blood Cancer 2021, 68, e28473. [Google Scholar] [CrossRef]
- Schleiermacher, G.; Janoueix-Lerosey, I.; Ribeiro, A.; Klijanienko, J.; Couturier, J.; Pierron, G.; Mosseri, V.; Valent, A.; Auger, N.; Plantaz, D.; et al. Accumulation of Segmental Alterations Determines Progression in Neuroblastoma. J. Clin. Oncol. 2010, 28, 3122–3130. [Google Scholar] [CrossRef] [PubMed]
- Tomioka, N.; Oba, S.; Ohira, M.; Misra, A.; Fridlyand, J.; Ishii, S.; Nakamura, Y.; Isogai, E.; Hirata, T.; Yoshida, Y.; et al. Novel risk stratification of patients with neuroblastoma by genomic signature, which is independent of molecular signature. Oncogene 2008, 27, 441–449. [Google Scholar] [CrossRef]
- Nakagawara, A.; Li, Y.; Izumi, H.; Muramori, K.; Inada, H.; Nishi, M. Neuroblastoma. Jpn. J. Clin. Oncol. 2018, 48, 214–241. [Google Scholar] [CrossRef] [PubMed]
- Gatta, G.; Botta, L.; Rossi, S.; Aareleid, T.; Bielska-Lasota, M.; Clavel, J.; Dimitrova, N.; Jakab, Z.; Kaatsch, P.; Lacour, B.; et al. Childhood cancer survival in Europe 1999–2007: Results of EUROCARE-5—A population-based study. Lancet Oncol. 2014, 15, 35–47. [Google Scholar] [CrossRef] [PubMed]
- Mlakar, V.; Jurkovic Mlakar, S.; Lopez, G.; Maris, J.M.; Ansari, M.; Gumy-Pause, F. 11q deletion in neuroblastoma: A review of biological and clinical implications. Mol. Cancer. 2017, 16, 114. [Google Scholar] [CrossRef]
- Castel, V.; Cañete, A.; Navarro, S.; García-Miguel, P.; Melero, C.; Acha, T.; Navajas, A.; Badal, M.D. Outcome of high-risk neuroblastoma using a dose intensity approach: Improvement in initial but not in long-term results. Med. Pediatr. Oncol. 2001, 37, 537–542. [Google Scholar] [CrossRef] [PubMed]
- Lundberg, K.I.; Treis, D.; Johnsen, J.I. Neuroblastoma Heterogeneity, Plasticity, and Emerging Therapies. Curr. Oncol. Rep. 2022, 24, 1053–1062. [Google Scholar] [CrossRef]
- Azar, W.J.; Zivkovic, S.; Werther, G.A.; Russo, V.C. IGFBP-2 nuclear translocation is mediated by a functional NLS sequence and is essential for its pro-tumorigenic actions in cancer cells. Oncogene 2014, 33, 578–588. [Google Scholar] [CrossRef]
- Azar, W.J.; Azar, S.H.X.; Higgins, S.; Hu, J.-F.; Hoffman, A.R.; Newgreen, D.F.; Werther, G.A.; Russo, V.C. IGFBP-2 enhances VEGF gene promoter activity and consequent promotion of angiogenesis by neuroblastoma cells. Endocrinology 2011, 152, 3332–3342. [Google Scholar] [CrossRef]
- Hecht, M.; Schulte, J.H.; Eggert, A.; Wilting, J.; Schweigerer, L. The neurotrophin receptor TrkB cooperates with c-Met in enhancing neuroblastoma invasiveness. Carcinogenesis 2005, 26, 2105–2115. [Google Scholar] [CrossRef] [PubMed]
- Ribatti, D.; Tamma, R. The chick embryo chorioallantoic membrane as an in vivo experimental model to study human neuroblastoma. J. Cell Physiol. 2018, 234, 152–157. [Google Scholar] [CrossRef]
- Pizon, M.; Schott, D.; Pachmann, U.; Schobert, R.; Pizon, M.; Wozniak, M.; Bobinski, R.; Pachmann, K. Chick Chorioallantoic Membrane (CAM) Assays as a Model of Patient-Derived Xenografts from Circulating Cancer Stem Cells (cCSCs) in Breast Cancer Patients. Cancers 2022, 14, 1476. [Google Scholar] [CrossRef] [PubMed]
- Eckrich, J.; Kugler, P.; Buhr, C.R.; Ernst, B.P.; Mendler, S.; Baumgart, J.; Brieger, J.; Wiesmann, N. Monitoring of tumor growth and vascularization with repetitive ultrasonography in the chicken chorioallantoic-membrane-assay. Sci. Rep. 2020, 10, 18585. [Google Scholar] [CrossRef] [PubMed]
- Chu, P.Y.; Koh, A.P.; Antony, J.; Huang, R.Y. Applications of the Chick Chorioallantoic Membrane as an Alternative Model for Cancer Studies. Cells Tissues Organs. 2022, 211, 222–237. [Google Scholar] [CrossRef]
- Merlos, M.A.; Casar, B.; Michalkova, H.; Jimenez, A.M.; Heger, Z.; Adam, V. Extending the Applicability of In Ovo and Ex Ovo Chicken Chorioallantoic Membrane Assays to Study Cytostatic Activity in Neuroblastoma Cells. Front. Oncol. 2021, 11, 707366. [Google Scholar] [CrossRef]
- Herrmann, A.; Moss, D.; Sée, V. The Chorioallantoic Membrane of the Chick Embryo to Assess Tumor Formation and Metastasis. Methods Mol. Biol. 2016, 1464, 97–105. [Google Scholar] [CrossRef]
- Herrmann, A.; Rice, M.; Lévy, R.; Pizer, B.L.; Losty, P.D.; Moss, D.; Sée, V. Cellular memory of hypoxia elicits neuroblastoma metastasis and enables invasion by non-aggressive neighbouring cells. Oncogenesis 2015, 4, e138. [Google Scholar] [CrossRef]
- Al-Mutawa, Y.K.; Herrmann, A.; Corbishley, C.; Losty, P.D.; Phelan, M.; Sée, V. Effects of hypoxic preconditioning on neuroblastoma tumour oxygenation and metabolic signature in a chick embryo model. Biosci. Rep. 2018, 38, BSR20180185. [Google Scholar] [CrossRef]
- Herrmann, A.; Taylor, A.; Murray, P.; Poptani, H.; Sée, V. Magnetic Resonance Imaging for Characterization of a Chick Embryo Model of Cancer Cell Metastases. Mol. Imaging 2018, 17, 1536012118809585. [Google Scholar] [CrossRef]
- Villanueva, A.A.; Falcón, P.; Espinoza, N.; Luis, S.R.; Milla, L.A.; Hernandez-SanMiguel, E.; Torres, V.A.; Sanchez-Gomez, P.; Palma, V. The Netrin-4/Neogenin-1 axis promotes neuroblastoma cell survival and migration. Oncotarget. 2017, 8, 9767–9782. [Google Scholar] [CrossRef]
- Dagg, C.P.; Karnofsky, D.A.; Roddy, J. Growth of transplantable human tumors in the chick embryo and hatched chick. Cancer Res. 1956, 16, 589–594. [Google Scholar] [PubMed]
- Zijlstra, A.; Mellor, R.; Panzarella, G.; Aimes, R.T.; Hooper, J.D.; Marchenko, N.D.; Quigley, J.P. A quantitative analysis of rate-limiting steps in the metastatic cascade using human-specific real-time polymerase chain reaction. Cancer Res. 2002, 62, 7083–7092. [Google Scholar] [PubMed]
- Deryugina, E.I.; Quigley, J.P. Chick embryo chorioallantoic membrane model systems to study and visualize human tumor cell metastasis Histochem. Cell Biol. 2008, 130, 1119–1130. [Google Scholar] [CrossRef]
- Kim, J.; Yu, W.; Kovalski, K.; Ossowski, L. Requirement for specific proteases in cancer cell intravasation as revealed by a novel semiquantitative PCR-based assay. Cell 1998, 94, 353–362. [Google Scholar] [CrossRef] [PubMed]
- Endo, Y.; Sasaki, T.; Harada, F.; Noguchi, M. Specific detection of metastasized human tumor cells in embryonic chicks by the polymerase chain reaction. Jpn J. Cancer Res. 1990, 81, 723–726. [Google Scholar] [CrossRef] [PubMed]
- Mira, E.; Lacalle, R.A.; Gómez-Moutón, C.; Leonardo, E.; Mañes, S. Quantitative determination of tumor cell intravasation in a real-time polymerase chain reaction-based assay. Clin. Exp. Metastasis 2002, 19, 313–318. [Google Scholar] [CrossRef]
- Rodrigo, M.A.M.; Michalkova, H.; Strmiska, V.; Casar, B.; Crespo, P.; Rios, V.d.L.; Casal, J.I.; Haddad, Y.; Guran, R.; Eckschlager, T.; et al. Metallothionein-3 promotes cisplatin chemoresistance remodelling in neuroblastoma. Sci. Rep. 2021, 11, 5496. [Google Scholar] [CrossRef]
- Swadi, R.R.; Sampat, K.; Herrmann, A.; Losty, P.D.; See, V.; Moss, D.J. CDK inhibitors reduce cell proliferation and reverse hypoxia-induced metastasis of neuroblastoma tumours in a chick embryo model. Sci. Rep. 2019, 9, 9136. [Google Scholar] [CrossRef]
- Swadi, R.; Mather, G.; Pizer, B.L.; Losty, P.D.; See, V.; Moss, D. Optimising the chick chorioallantoic membrane xenograft model of neuroblastoma for drug delivery. BMC Cancer 2018, 18, 28. [Google Scholar] [CrossRef]
- Michaelis, M.; Agha, B.; Rothweiler, F.; Löschmann, N.; Voges, Y.; Mittelbronn, M.; Starzetz, T.; Harter, P.N.; Abhari, B.A.; Fulda, S.; et al. Identification of flubendazole as potential anti-neuroblastoma compound in a large cell line screen. Sci. Rep. 2015, 5, 8202. [Google Scholar] [CrossRef]
- Carter, R.; Mullassery, D.; See, V.; Theocharatos, S.; Pizer, B.; Losty, P.D.; Jesudason, E.; Moss, D.J. Exploitation of chick embryo environments to reprogram MYCN-amplified neuroblastoma cells to a benign phenotype, lacking detectable MYCN expression. Oncogenesis 2012, 1, e24. [Google Scholar] [CrossRef]
- Hammer-Wilson, M.J.; Cao, D.; Kimel, S.; Berns, M.W. Photodynamic parameters in the chick chorioallantoic membrane (CAM) bioassay for photosensitizers administered intraperitoneally (IP) into the chick embryo. Photochem. Photobiol. Sci. 2002, 1, 721–728. [Google Scholar] [CrossRef]
- Gottfried, V.; Davidi, R.; Averguj, C.; Kimel, S. In vivo damage to chorioallantoic membrane blood vessels by porphycene-induced photodynamic therapy. J. Photochem. Photobiol. B 1995, 30, 115–121. [Google Scholar] [CrossRef] [PubMed]
- Peterka, M.; Klepácek, I. Light irradiation increases embryotoxicity of photodynamic therapy sensitizers (5-aminolevulinic acid and protoporphyrin IX) in chick embryos. Reprod. Toxicol. 2001, 15, 111–116. [Google Scholar] [CrossRef]
- Ribatti, D.; Alessandri, G.; Vacca, A.; Iurlaro, M.; Ponzoni, M. Human neuroblastoma cells produce extracellular matrix-degrading enzymes, induce endothelial cell proliferation and are angiogenic in vivo. Int. J. Cancer 1998, 77, 449–454. [Google Scholar] [CrossRef]
- Ribatti, D.; Raffaghello, L.; Pastorino, F.; Nico, B.; Brignole, C.; Vacca, A.; Ponzoni, M. In vivo angiogenic activity of neuroblastoma correlates with MYCN oncogene overexpression. Int. J. Cancer 2002, 102, 351–354. [Google Scholar] [CrossRef] [PubMed]
- Hagenbuchner, J.; Rupp, M.; Salvador, C.; Meister, B.; Kiechl-Kohlendorfer, U.; Müller, T.; Geiger, K.; Sergi, C.; Obexer, P.; Ausserlechner, M.J. Nuclear FOXO3 predicts adverse clinical outcome and promotes tumor angiogenesis in neuroblastoma. Oncotarget 2016, 7, 77591–77606. [Google Scholar] [CrossRef] [PubMed]
- Klingenberg, M.; Becker, J.; Eberth, S.; Kube, D.; Wilting, J. The NADPH oxidase inhibitor imipramine-blue in the treatment of Burkitt lymphoma. Mol. Cancer Ther. 2014, 13, 833–841. [Google Scholar] [CrossRef] [PubMed]
- Michaelis, M.; Klassert, D.; Barth, S.; Suhan, T.; Breitling, R.; Mayer, B.; Hinsch, N.; Doerr, H.W.; Cinatl, J.; Cinatl, J. Chemoresistance acquisition induces a global shift of expression of aniogenesis-associated genes and increased pro-angogenic activity in neuroblastoma cells. Mol. Cancer 2009, 8, 80. [Google Scholar] [CrossRef]
- Mangieri, D.; Nico, B.; Coluccia, A.M.; Vacca, A.; Ponzoni, M.; Ribatti, D. An alternative in vivo system for testing angiogenic potential of human neuroblastoma cells. Cancer Lett. 2009, 277, 199–204. [Google Scholar] [CrossRef] [PubMed]
- Pagnan, G.; Di Paolo, D.; Carosio, R.; Pastorino, F.; Marimpietri, D.; Brignole, C.; Pezzolo, A.; Loi, M.; Galietta, L.J.; Piccardi, F.; et al. The combined therapeutic effects of bortezomib and fenretinide on neuroblastoma cells involve endoplasmic reticulum stress response. Clin. Cancer Res. 2009, 15, 1199–1209. [Google Scholar] [CrossRef] [PubMed]
- Pastorino, F.; Di Paolo, D.; Piccardi, F.; Nico, B.; Ribatti, D.; Daga, A.; Baio, G.; Neumaier, C.E.; Brignole, C.; Loi, M.; et al. Enhanced antitumor efficacy of clinical-grade vasculature-targeted liposomal doxorubicin. Clin. Cancer Res. 2008, 14, 7320–7329. [Google Scholar] [CrossRef] [PubMed]
- Marimpietri, D.; Brignole, C.; Nico, B.; Pastorino, F.; Pezzolo, A.; Piccardi, F.; Cilli, M.; Di Paolo, D.; Pagnan, G.; Longo, L.; et al. Combined therapeutic effects of vinblastine and rapamycin on human neuroblastoma growth, apoptosis, and angiogenesis. Clin. Cancer Res. 2007, 13, 3977–3988. [Google Scholar] [CrossRef]
- Marimpietri, D.; Nico, B.; Vacca, A.; Mangieri, D.; Catarsi, P.; Ponzoni, M.; Ribatti, D. Synergistic inhibition of human neuroblastoma-related angiogenesis by vinblastine and rapamycin. Oncogene 2005, 24, 6785–6795. [Google Scholar] [CrossRef]
- Ribatti, D.; Nico, B.; Pezzolo, A.; Vacca, A.; Meazza, R.; Cinti, R.; Carlini, B.; Parodi, F.; Pistoia, V.; Corrias, M.V. Angiogenesis in a human neuroblastoma xenograft model: Mechanisms and inhibition by tumour-derived interferon-gamma. Br. J. Cancer 2006, 94, 1845–1852. [Google Scholar] [CrossRef]
- Brignole, C.; Marimpietri, D.; Pastorino, F.; Nico, B.; Di Paolo, D.; Cioni, M.; Piccardi, F.; Cilli, M.; Pezzolo, A.; Corrias, M.V.; et al. Effect of bortezomib on human neuroblastoma cell growth, apoptosis, and angiogenesis. J. Natl. Cancer Inst. 2006, 98, 1142–1157. [Google Scholar] [CrossRef]
- Ribatti, D.; Vacca, A. Models for studying angiogenesis in vivo. Int. J. Biol. Markers 1999, 14, 207–213. [Google Scholar] [CrossRef]
- Ausprunk, D.H.; Folkman, J. Vascular injury in transplanted tissues. Fine structural changes in tumor, adult, and embryonic blood vessels. Virchows Arch. B Cell Pathol. 1976, 31–44. [Google Scholar] [CrossRef]
- DeBord, L.C.; Pathak, R.R.; Villaneuva, M.; Liu, H.-C.; Harrington, D.A.; Yu, W.; Lewis, M.T.; Sikora, A.G. The chick chorioallantoic membrane (CAM) as a versatile patient-derived xenograft (PDX) platform for precision medicine and preclinical research. Am. J. Cancer Res. 2018, 8, 1642–1660. [Google Scholar]


| FEATURES | CAM | MOUSE | CELL LINE |
|---|---|---|---|
| Ease of use [38] | Yes | No | Yes |
| Low cost [38] | Yes | No | No |
| Possibility of having a large sample size [39] | Yes | No | Yes |
| Real-time visualization of the assays [40] | Yes | Yes | Yes |
| Possibility of studying metastasis [41] | Yes | Yes | No |
| Possibility of studying angiogenesis [42,43] | Yes | Yes | No |
| Maintains tumor heterogeneity [44] | Yes | Yes | No |
| Replicates the tumor microenvironment [36] | Yes | Yes | No |
| Complete accessibility to the circulatory system [36] | Yes | No | No |
| Requirement for animal protocol approval (country dependent) [35] | No | Yes | No |
| Quickly tumoral formation [44] | Yes (3 and 4 days) | No (3–4 weeks) | Are not generated |
| Pain perception [35] | No (until day14) | Yes | No |
| Direct visibility of the tumor [44] | Yes | No | No |
| Availability of reagents (antibodies, primers, etc.) [36] | No | Yes | Yes |
| Immunodeficiency [45] | Yes | Yes | Yes |
| Genetic closeness to humans [46,47,48] | No | Yes | Yes |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Patiño-Morales, C.C.; Jaime-Cruz, R.; Ramírez-Fuentes, T.C.; Villavicencio-Guzmán, L.; Salazar-García, M. Technical Implications of the Chicken Embryo Chorioallantoic Membrane Assay to Elucidate Neuroblastoma Biology. Int. J. Mol. Sci. 2023, 24, 14744. https://doi.org/10.3390/ijms241914744
Patiño-Morales CC, Jaime-Cruz R, Ramírez-Fuentes TC, Villavicencio-Guzmán L, Salazar-García M. Technical Implications of the Chicken Embryo Chorioallantoic Membrane Assay to Elucidate Neuroblastoma Biology. International Journal of Molecular Sciences. 2023; 24(19):14744. https://doi.org/10.3390/ijms241914744
Chicago/Turabian StylePatiño-Morales, Carlos César, Ricardo Jaime-Cruz, Tania Cristina Ramírez-Fuentes, Laura Villavicencio-Guzmán, and Marcela Salazar-García. 2023. "Technical Implications of the Chicken Embryo Chorioallantoic Membrane Assay to Elucidate Neuroblastoma Biology" International Journal of Molecular Sciences 24, no. 19: 14744. https://doi.org/10.3390/ijms241914744