
The Genome-Wide Identification, Characterization, and Expression Analysis of the Strictosidine Synthase-like Family in Maize (Zea mays L.)


Abstract
:1. Introduction
2. Results
2.1. Identification of the Maize SSL Gene Family
2.2. Phylogenetic Analysis of the ZmSSL Genes’ Protein
2.3. Structural Analysis of ZmSSL Genes
2.4. Chromosomal Localization and Collinearity Analysis of ZmSSL Genes
2.5. The Cis-Elements Analysis of ZmSSL Gene Promoter Regions
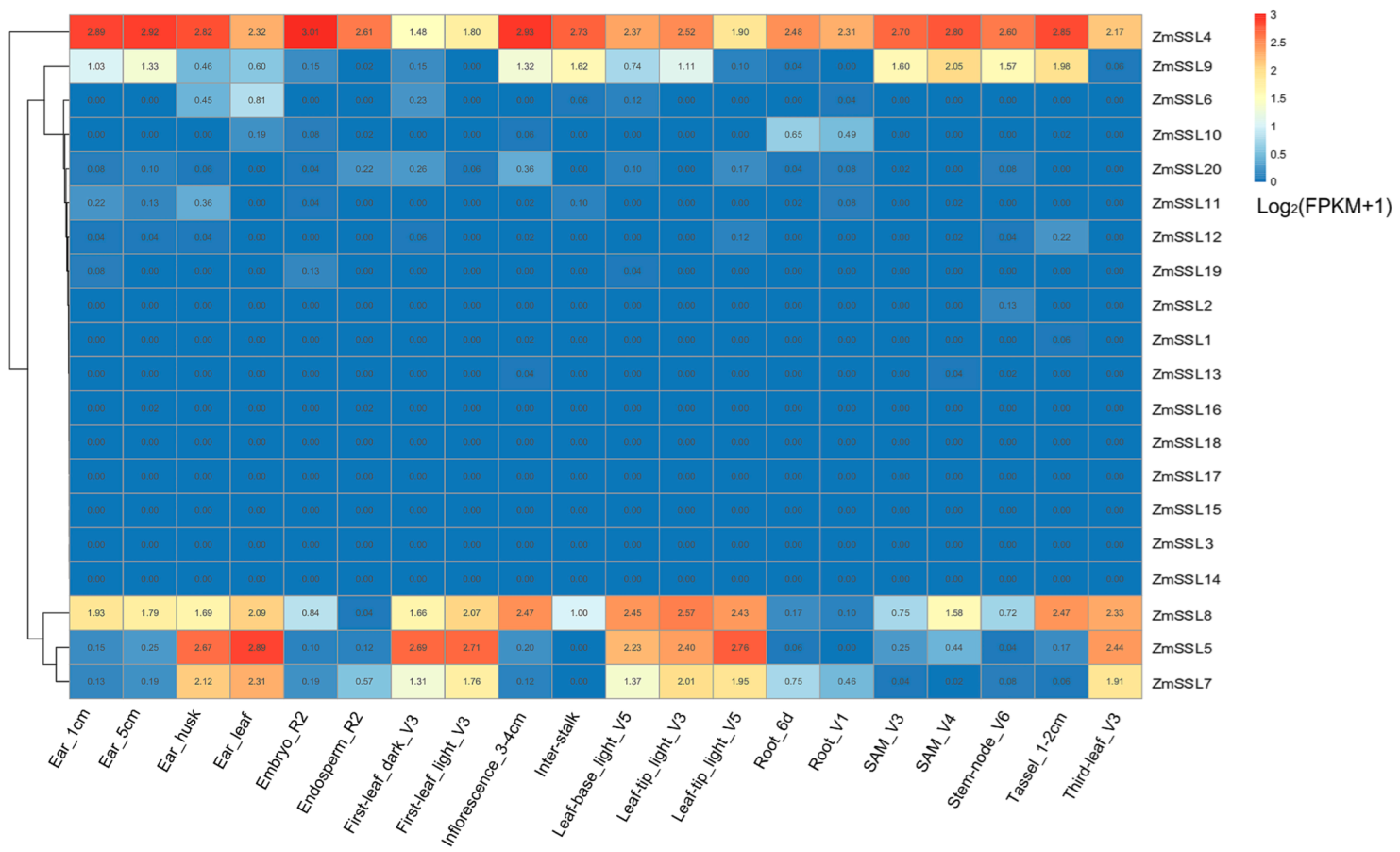
2.6. Expression Patterns of ZmSSL Genes in Different Maize Tissues under Normal Conditions
2.7. Expression Patterns of ZmSSL Genes under Abiotic/Biotic Stress Conditions
2.8. RT-qPCR Validation of ZmSSL Genes under Abiotic/Biotic Stresses
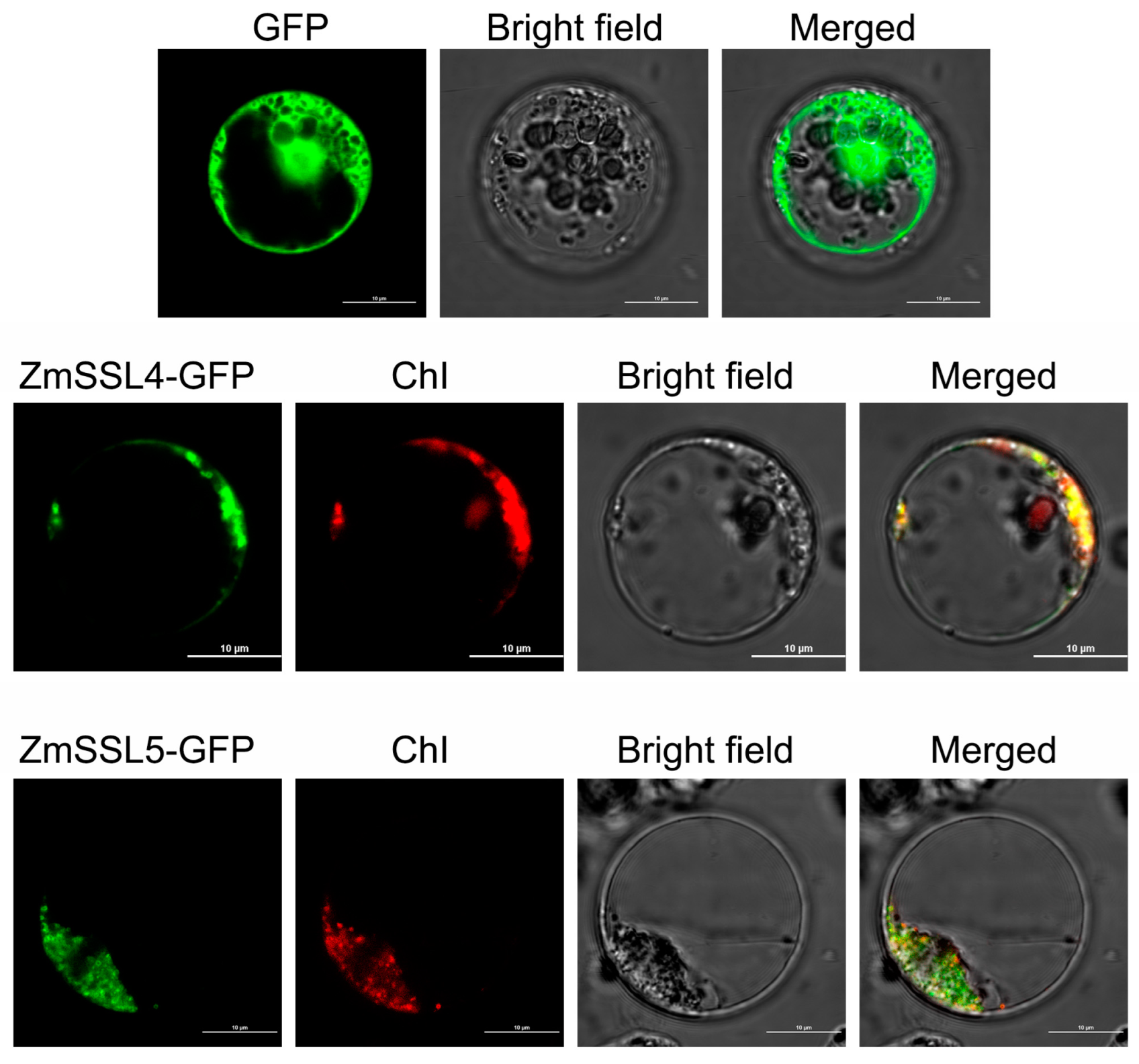
2.9. Subcellular Localization of ZmSSL4 and ZmSSL5
3. Discussion
4. Materials and Methods
4.1. Plant Materials and Stress Treatments
4.2. Identification of ZmSSL Genes
4.3. Phylogenetic and Structural Analysis of ZmSSL Genes
4.4. Chromosomal Localization and Collinearity Analysis
4.5. Cis-Acting Element Analysis
4.6. Transcriptome Data Analysis
4.7. RT-qPCR Analysis of Gene Expression
4.8. Subcellular Localization
4.9. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Yang, Z.; Cao, Y.; Shi, Y.; Qin, F.; Jiang, C.; Yang, S. Genetic and Molecular Exploration of Maize Environmental Stress Resilience: Towards Sustainable Agriculture. Mol. Plant 2023. [Google Scholar] [CrossRef]
- Zhu, J.K. Abiotic Stress Signaling and Responses in Plants. Cell 2016, 167, 313–324. [Google Scholar] [CrossRef] [PubMed]
- Chen, K.; Li, G.J.; Bressan, R.A.; Song, C.P.; Zhu, J.K.; Zhao, Y. Abscisic acid dynamics, signaling, and functions in plants. J. Integr. Plant Biol. 2020, 62, 25–54. [Google Scholar] [CrossRef] [PubMed]
- Xie, C.; Yang, L.; Jia, G.; Yan, K.; Zhang, S.; Yang, G.; Wu, C.; Gai, Y.; Zheng, C.; Huang, J. Maize HEAT UP-REGULATED GENE 1 plays vital roles in heat stress tolerance. J. Exp. Bot. 2022, 73, 6417–6433. [Google Scholar] [CrossRef]
- Pruitt, R.N.; Gust, A.A.; Nurnberger, T. Plant immunity unified. Nat. Plants. 2021, 7, 382–383. [Google Scholar] [CrossRef]
- Tang, D.; Wang, G.; Zhou, J.M. Receptor Kinases in Plant-Pathogen Interactions: More Than Pattern Recognition. Plant Cell 2017, 29, 618–637. [Google Scholar] [CrossRef] [PubMed]
- Deng, X.; Zhao, L.; Fang, T.; Xiong, Y.; Ogutu, C.; Yang, D.; Vimolmangkang, S.; Liu, Y.; Han, Y. Investigation of benzylisoquinoline alkaloid biosynthetic pathway and its transcriptional regulation in lotus. Hortic. Res. 2018, 5, 29. [Google Scholar] [CrossRef] [PubMed]
- Wang, R.; Zhao, W.; Yao, W.; Wang, Y.; Jiang, T.; Liu, H. Genome-Wide Analysis of Strictosidine Synthase-like Gene Family Revealed Their Response to Biotic/Abiotic Stress in Poplar. Int. J. Mol. Sci. 2023, 24, 10117. [Google Scholar] [CrossRef]
- Sohani, M.M.; Schenk, P.M.; Schultz, C.J.; Schmidt, O. Phylogenetic and transcriptional analysis of a strictosidine synthase-like gene family in Arabidopsis thaliana reveals involvement in plant defence responses. Plant Biol. 2009, 11, 105–117. [Google Scholar] [CrossRef]
- Ali, E.F.; El-Shehawi, A.M.; Ibrahim, O.H.M.; Abdul-Hafeez, E.Y.; Moussa, M.M.; Hassan, F.A.S. A vital role of chitosan nanoparticles in improvisation the drought stress tolerance in Catharanthus roseus (L.) through biochemical and gene expression modulation. Plant Physiol. Biochem. 2021, 161, 166–175. [Google Scholar] [CrossRef]
- Tang, W.; Liu, X.; He, Y.; Yang, F. Enhancement of Vindoline and Catharanthine Accumulation, Antioxidant Enzymes Activities, and Gene Expression Levels in Catharanthus roseus Leaves by Chitooligosaccharides Elicitation. Mar. Drugs 2022, 20, 188. [Google Scholar] [CrossRef]
- Liu, Y.; Yang, H.; Wen, F.; Bao, L.; Zhao, Z.; Zhong, Z. Chitooligosaccharide-induced plant stress resistance. Carbohydr. Polym. 2023, 302, 120344. [Google Scholar] [CrossRef] [PubMed]
- Kibble, N.A.J.; Sohani, M.M.; Shirley, N.; Byrt, C.; Roessner, U.; Bacic, A.; Schmidt, O.; Schultz, C.J. Phylogenetic analysis and functional characterisation of strictosidine synthase-like genes in Arabidopsis thaliana. Funct. Plant Biol. 2010, 36, 1098–1109. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Meng, J.; Luan, Y. miR1916 plays a role as a negative regulator in drought stress resistance in tomato and tobacco. Biochem. Biophys. Res. Commun. 2019, 508, 597–602. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Meng, J.; He, X.L.; Zhang, M.; Luan, Y.S. Solanum lycopersicum microRNA1916 targets multiple target genes and negatively regulates the immune response in tomato. Plant Cell Environ. 2019, 42, 1393–1407. [Google Scholar] [CrossRef]
- Zhu, W.; Miao, X.; Qian, J.; Chen, S.; Jin, Q.; Li, M.; Han, L.; Zhong, W.; Xie, D.; Shang, X.; et al. A translatome-transcriptome multi-omics gene regulatory network reveals the complicated functional landscape of maize. Genome Biol. 2023, 24, 60. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.T.; Ru, J.N.; Liu, Y.W.; Yang, J.F.; Li, M.; Xu, Z.S.; Fu, J.D. The Maize WRKY Transcription Factor ZmWRKY40 Confers Drought Resistance in Transgenic Arabidopsis. Int. J. Mol. Sci. 2018, 19, 2580. [Google Scholar] [CrossRef]
- Makarevitch, I.; Waters, A.J.; West, P.T.; Stitzer, M.; Hirsch, C.N.; Ross-Ibarra, J.; Springer, N.M. Transposable elements contribute to activation of maize genes in response to abiotic stress. PLoS Genet. 2015, 11, e1004915. [Google Scholar]
- Swart, V.; Crampton, B.G.; Ridenour, J.B.; Bluhm, B.H.; Olivier, N.A.; Meyer, J.J.M.; Berger, D.K. Complementation of CTB7 in the Maize Pathogen Cercospora zeina Overcomes the Lack of In Vitro Cercosporin Production. Mol. Plant Microbe Interact. 2017, 30, 710–724. [Google Scholar] [CrossRef]
- Hoopes, G.M.; Hamilton, J.P.; Wood, J.C.; Esteban, E.; Pasha, A.; Vaillancourt, B.; Provart, N.J.; Buell, C.R. An updated gene atlas for maize reveals organ-specific and stress-induced genes. Plant J. 2019, 97, 1154–1167. [Google Scholar] [CrossRef]
- Verma, P.; Sharma, A.; Khan, S.A.; Shanker, K.; Mathur, A.K. Over-expression of Catharanthus roseus tryptophan decarboxylase and strictosidine synthase in rol gene integrated transgenic cell suspensions of Vinca minor. Protoplasma 2015, 252, 373–381. [Google Scholar] [CrossRef]
- Cannon, S.B.; Mitra, A.; Baumgarten, A.; Young, N.D.; May, G. The roles of segmental and tandem gene duplication in the evolution of large gene families in Arabidopsis thaliana. BMC Plant Biol. 2004, 4, 10. [Google Scholar] [CrossRef] [PubMed]
- Hu, W.; Criscione, F.; Liang, S.; Tu, Z. MicroRNAs of two medically important mosquito species: Aedes aegypti and Anopheles stephensi. Insect Mol. Biol. 2015, 24, 240–252. [Google Scholar] [CrossRef]
- van Baren, M.J.; Bachy, C.; Reistetter, E.N.; Purvine, S.O.; Grimwood, J.; Sudek, S.; Yu, H.; Poirier, C.; Deerinck, T.J.; Kuo, A.; et al. Evidence-based green algal genomics reveals marine diversity and ancestral characteristics of land plants. BMC Genom. 2016, 17, 267. [Google Scholar] [CrossRef]
- Soltani, B.M.; Ehlting, J.; Hamberger, B.; Douglas, C.J. Multiple cis-regulatory elements regulate distinct and complex patterns of developmental and wound-induced expression of Arabidopsis thaliana 4CL gene family members. Planta 2006, 224, 1226–1238. [Google Scholar] [CrossRef]
- Walther, D.; Brunnemann, R.; Selbig, J. The regulatory code for transcriptional response diversity and its relation to genome structural properties in A. thaliana. PLoS Genet. 2007, 3, e11. [Google Scholar] [CrossRef] [PubMed]
- Abdullah, M.; Cheng, X.; Cao, Y.; Su, X.; Manzoor, M.A.; Gao, J.; Cai, Y.; Lin, Y. Zinc Finger-Homeodomain Transcriptional Factors (ZHDs) in Upland Cotton (Gossypium hirsutum): Genome-Wide Identification and Expression Analysis in Fiber Development. Front. Genet. 2018, 9, 357. [Google Scholar] [CrossRef]
- Ma, W.; Yang, J.; Ding, J.; Duan, C.; Zhao, W.; Peng, Y.L.; Bhadauria, V. CRISPR/Cas9-mediated deletion of large chromosomal segments identifies a minichromosome modulating the Colletotrichum graminicola virulence on maize. Int. J. Biol. Macromol. 2023, 245, 125462. [Google Scholar] [CrossRef]
- Cheng, Z.; Lv, X.; Duan, C.; Zhu, H.; Wang, J.; Xu, Z.; Yin, H.; Zhou, X.; Li, M.; Hao, Z.; et al. Pathogenicity Variation in Two Genomes of Cercospora Species Causing Gray Leaf Spot in Maize. Mol. Plant Microbe Interact. 2023, 36, 14–25. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Wang, R.; Yu, Y.; Gu, Y.; Wang, S.; Liao, S.; Xu, X.; Jiang, T.; Yao, W. Genome-Wide Analysis of SIMILAR TO RCD ONE (SRO) Family Revealed Their Roles in Abiotic Stress in Poplar. Int. J. Mol. Sci. 2023, 24, 4146. [Google Scholar] [CrossRef] [PubMed]
- Bailey, T.L.; Boden, M.; Buske, F.A.; Frith, M.; Grant, C.E.; Clementi, L.; Ren, J.; Li, W.W.; Noble, W.S. MEME SUITE: Tools for motif discovery and searching. Nucleic Acids Res. 2009, 37, W202–W208. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Chen, H.; Zhang, Y.; Thomas, H.R.; Frank, M.H.; He, Y.; Xia, R. TBtools: An Integrative Toolkit Developed for Interactive Analyses of Big Biological Data. Mol. Plant 2020, 13, 1194–1202. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Tang, H.; Debarry, J.D.; Tan, X.; Li, J.; Wang, X.; Lee, T.H.; Jin, H.; Marler, B.; Guo, H.; et al. MCScanX: A toolkit for detection and evolutionary analysis of gene synteny and collinearity. Nucleic Acids Res. 2012, 40, e49. [Google Scholar] [CrossRef]
- Higo, K.; Ugawa, Y.; Iwamoto, M.; Korenaga, T. Plant cis-acting regulatory DNA elements (PLACE) database: 1999. Nucleic Acids Res. 1999, 27, 297–300. [Google Scholar] [CrossRef] [PubMed]
- Gu, L.; Chen, X.; Hou, Y.; Wang, H.; Wang, H.; Zhu, B.; Du, X. ZmWRKY70 activates the expression of hypoxic responsive genes in maize and enhances tolerance to submergence in Arabidopsis. Plant Physiol. Biochem. 2023, 201, 107861. [Google Scholar] [CrossRef]
- Zuo, D.; Hu, M.; Zhou, W.; Lei, F.; Zhao, J.; Gu, L. EcAGL enhances cadmium tolerance in transgenic Arabidopsis thaliana through inhibits cadmium transport and ethylene synthesis pathway. Plant Physiol. Biochem. 2023, 201, 107900. [Google Scholar] [CrossRef]
- Gu, L.; Jiang, T.; Zhang, C.; Li, X.; Wang, C.; Zhang, Y.; Li, T.; Dirk, L.M.A.; Downie, A.B.; Zhao, T. Maize HSFA2 and HSBP2 antagonistically modulate raffinose biosynthesis and heat tolerance in Arabidopsis. Plant J. 2019, 100, 128–142. [Google Scholar] [CrossRef] [PubMed]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene NCBI ID | Gene Name | CDS (bp) | Protein Size (aa) | MW (kDa) | pI | Subcellular Protein Location |
|---|---|---|---|---|---|---|
| XM_008657285.2 | ZmSSL1 | 1017 | 338 | 36.31 | 6.08 | Extracellular/Cell wall |
| XM_008657302.2 | ZmSSL2 | 1020 | 339 | 36.46 | 5.89 | Extracellular/Cell wall |
| XM_008657311.2 | ZmSSL3 | 1017 | 338 | 36.33 | 6.08 | Extracellular/Cell wall |
| NM_001156536.2 | ZmSSL4 | 1197 | 398 | 44.10 | 6.34 | Plasma membrane |
| NM_001148541.1 | ZmSSL5 | 1104 | 367 | 39.46 | 5.25 | Chloroplast/Thylakoid |
| XM_008673664.2 | ZmSSL6 | 654 | 217 | 22.64 | 4.53 | Chloroplast/Thylakoid |
| NM_001139223.1 | ZmSSL7 | 1173 | 390 | 41.94 | 8.92 | Extracellular/Cell wall |
| NM_001157297.1 | ZmSSL8 | 1032 | 343 | 36.17 | 7.73 | Chloroplast/Thylakoid |
| XM_008654423.3 | ZmSSL9 | 1074 | 357 | 38.25 | 7.75 | Chloroplast/Thylakoid |
| XM_008658790.4 | ZmSSL10 | 1038 | 345 | 36.66 | 6.00 | Chloroplast/Thylakoid |
| NM_001157473.1 | ZmSSL11 | 1038 | 345 | 36.63 | 5.81 | Chloroplast/Thylakoid |
| XM_008659695.3 | ZmSSL12 | 594 | 197 | 20.81 | 7.73 | Chloroplast/Thylakoid |
| XM_008658833.3 | ZmSSL13 | 1038 | 345 | 36.78 | 7.01 | Chloroplast/Thylakoid |
| XM_008658834.2 | ZmSSL14 | 1038 | 345 | 36.74 | 6.61 | Chloroplast/Thylakoid |
| XM_008658837.2 | ZmSSL15 | 1038 | 345 | 36.81 | 6.61 | Chloroplast/Thylakoid |
| XM_008658836.3 | ZmSSL16 | 1038 | 345 | 36.78 | 7.02 | Chloroplast/Thylakoid |
| XM_020542371.1 | ZmSSL17 | 1050 | 349 | 37.09 | 7.01 | Chloroplast/Thylakoid |
| XM_008658839.2 | ZmSSL18 | 1050 | 349 | 37.05 | 6.09 | Chloroplast/Thylakoid |
| XM_008658840.2 | ZmSSL19 | 1038 | 345 | 36.75 | 7.01 | Chloroplast/Thylakoid |
| NM_001324231.1 | ZmSSL20 | 1239 | 412 | 46.54 | 6.00 | Endoplasmic reticulum |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gu, L.; Cao, Y.; Chen, X.; Wang, H.; Zhu, B.; Du, X.; Sun, Y. The Genome-Wide Identification, Characterization, and Expression Analysis of the Strictosidine Synthase-like Family in Maize (Zea mays L.). Int. J. Mol. Sci. 2023, 24, 14733. https://doi.org/10.3390/ijms241914733
Gu L, Cao Y, Chen X, Wang H, Zhu B, Du X, Sun Y. The Genome-Wide Identification, Characterization, and Expression Analysis of the Strictosidine Synthase-like Family in Maize (Zea mays L.). International Journal of Molecular Sciences. 2023; 24(19):14733. https://doi.org/10.3390/ijms241914733
Chicago/Turabian StyleGu, Lei, Yongyan Cao, Xuanxuan Chen, Hongcheng Wang, Bin Zhu, Xuye Du, and Yiyue Sun. 2023. "The Genome-Wide Identification, Characterization, and Expression Analysis of the Strictosidine Synthase-like Family in Maize (Zea mays L.)" International Journal of Molecular Sciences 24, no. 19: 14733. https://doi.org/10.3390/ijms241914733