

Structure-Function Characterisation of Eop1 Effectors from the Erwinia-Pantoea Clade Reveals They May Acetylate Their Defence Target through a Catalytic Dyad
 , , and
, , and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
2.1. YopJ Family Effectors from E. amylovora and Other Related Species from the “Erwinia-Pantoea” Clade Trigger HR-like Cell Death in Nicotiana tabacum
2.2. AlphaFold2-Predicted Tertiary Structures of the “Erwinia-Pantoea” Eop1 Effectors Mimic the HopZ1a Structure, Providing Insight into Their Functional and Structural Characteristics
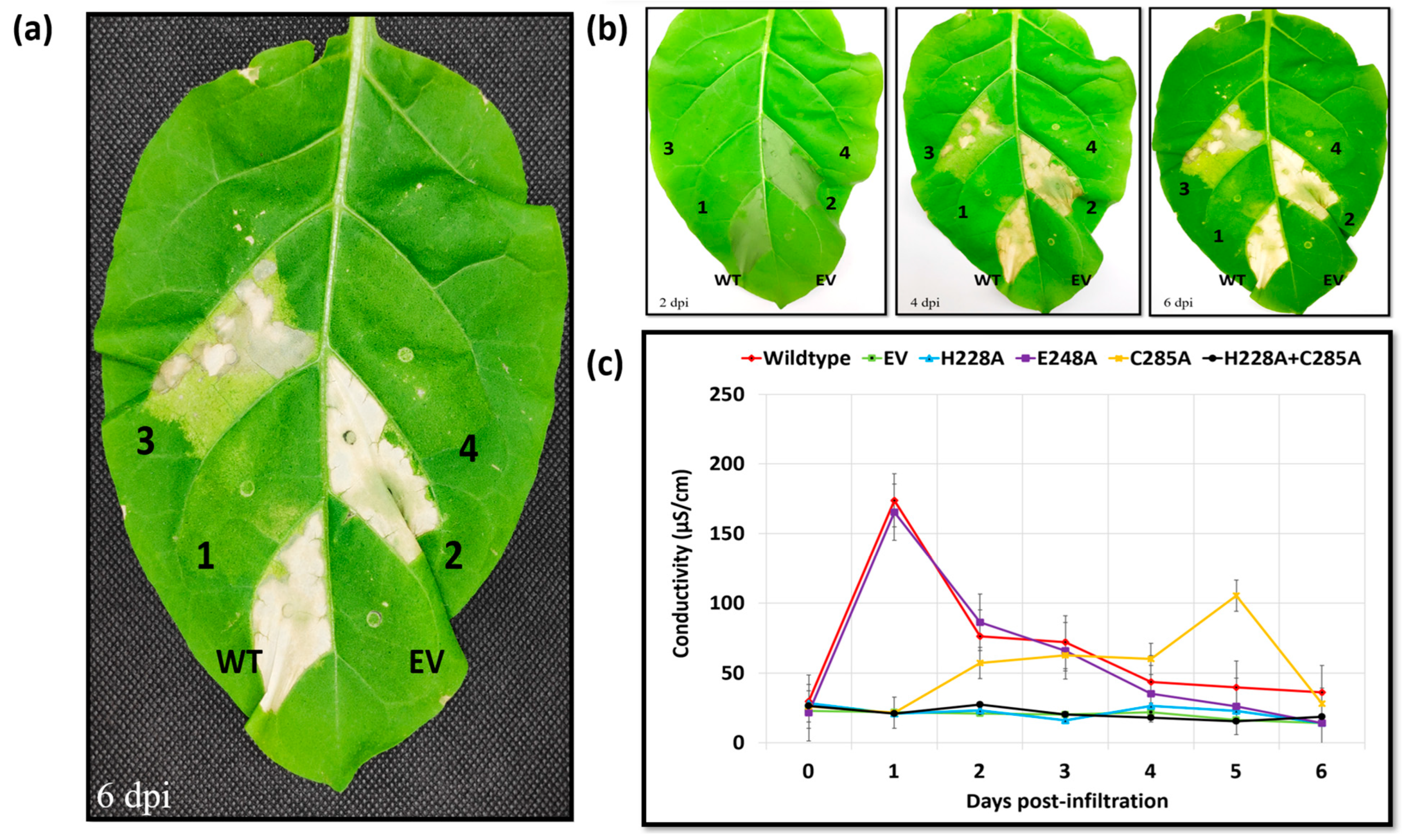
2.3. Erwinia amylovora Eop1 Effector (Ea246) Utilises a Catalytic Dyad with a Conserved Histidine Residue Required for Catalysis
- (a)
- One hypothesis predicts that C285 is not the “actual” nucleophile, but another amino acid residue functioning as the nucleophile in the catalysis is proximal to the mutated C285 residue. The C285A mutation could induce a conformation deformity in the catalytic pocket which interferes with, and impedes the catalytic activity, thus resulting in the slow catalysis, and consequently slower HR. However, AF2-predicted structures of the catalytic triad residue-mutated variants of Ea246 Eop1 exhibited no significant deviation from the wild-type structure (Supplementary Figure S9). Therefore, this possibility was deduced to be unlikely.
- (b)
- An alternate hypothesis predicts the presence of a “secondary nucleophile” that compensates for the loss of the “primary nucleophile” in Ea246 Eop1. Simply put, the proposal is that, whenever present, the proposed nucleophile, C285, functions as the “primary” nucleophile; however, in its absence (as is the case for the C285A mutant), another nearby “secondary nucleophile” with potentially weaker nucleophilic activity can function as a “substitute nucleophile” and still drive a slower rate of catalysis in this Eop1 variant.
2.4. Ea246 Eop1 Retains Its Ability to Function via a Substitute Nucleophile in the Absence of the Primary Nucleophile
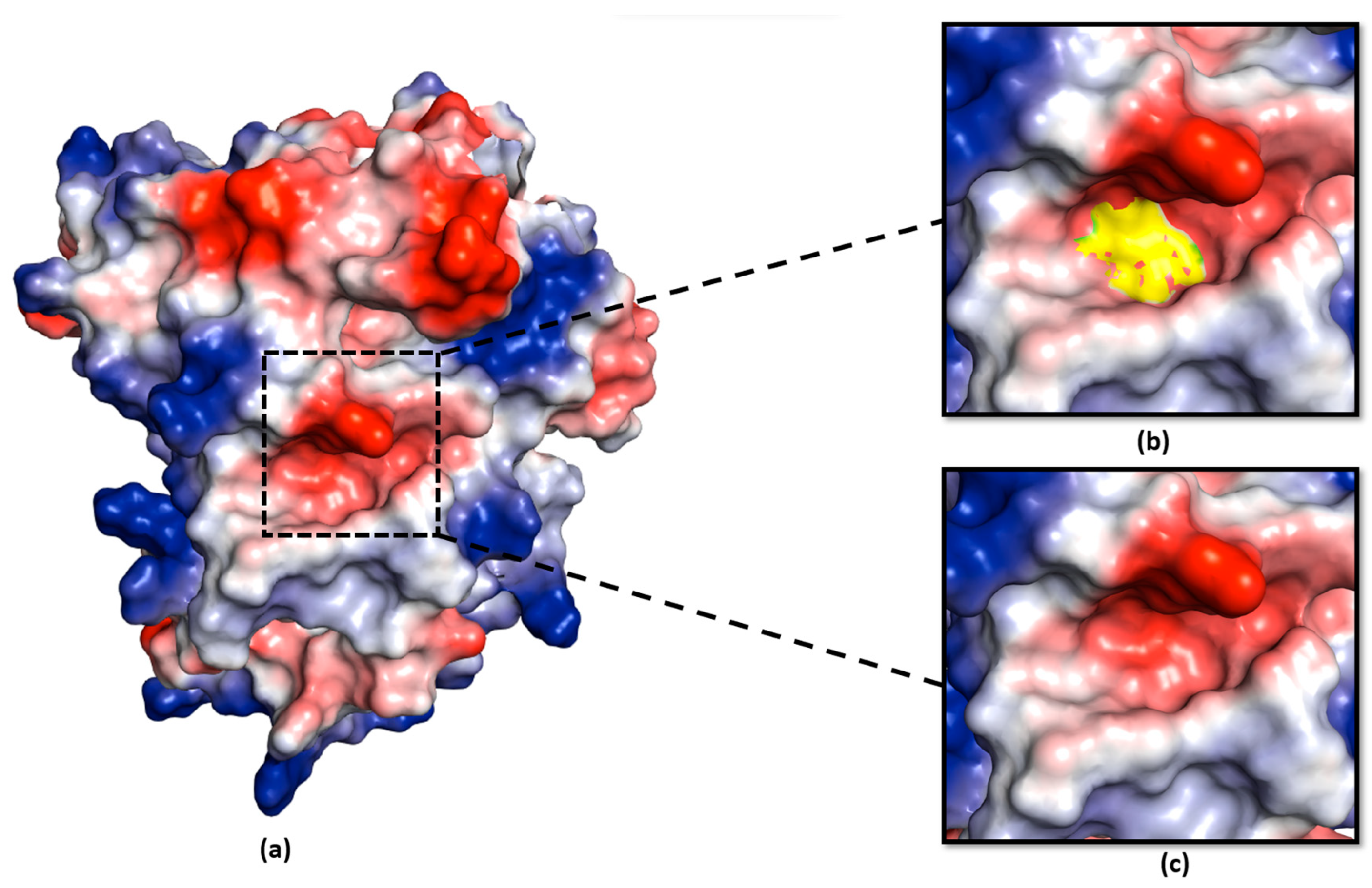
2.5. Negative Charge at the Catalytic Pocket Provides a Suitable Environment for the Serine Residue to Function as a Secondary Nucleophile in Ea246 Eop1
3. Materials and Methods
3.1. Bacterial Strains and Protocols
3.2. Plant Material
3.3. Agrobacterium tumefaciens Mediated Transient Expression Assays
3.4. Electrolyte Leakage Assays
3.5. Bioinformatic Methods
3.6. Protein Tertiary Structure Models
3.7. Site-Directed Mutagenesis
4. Conclusions
Novel Findings from the Analysis of the Eop1 Branch of YopJ Effectors
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Jones, J.D.; Dangl, J.L. The plant immune system. Nature 2006, 444, 323–329. [Google Scholar] [CrossRef] [PubMed]
- Boller, T.; Felix, G. A renaissance of elicitors: Perception of microbe-associated molecular patterns and danger signals by pattern-recognition receptors. Annu. Rev. Plant Biol. 2009, 60, 379–406. [Google Scholar] [CrossRef] [PubMed]
- Afzal, A.J.; da Cunha, L.; Mackey, D. Separable fragments and membrane tethering of Arabidopsis RIN4 regulate its suppression of PAMP-triggered immunity. Plant Cell 2011, 23, 3798–3811. [Google Scholar] [CrossRef]
- Rathore, J.S.; Ghosh, C. Pathogen-Associated Molecular Patterns and Their Perception in Plants. In Molecular Aspects of Plant-Pathogen Interaction; Singh, A., Singh, I.K., Eds.; Springer: Singapore, 2018. [Google Scholar]
- Głowacki, S.; Macioszek, V.K.; Kononowicz, A.K. R proteins as fundamentals of plant innate immunity. Cell Mol. Biol. Lett. 2011, 16, 1–24. [Google Scholar] [CrossRef] [PubMed]
- Kourelis, J.; van der Hoorn, R.A.L. Defended to the Nines: 25 Years of Resistance Gene Cloning Identifies Nine Mechanisms for R Protein Function. Plant Cell 2018, 30, 285–299. [Google Scholar] [CrossRef] [PubMed]
- Yang, S.; Li, J.; Zhang, X.; Zhang, Q.; Huang, J.; Chen, J.Q.; Hartl, D.L.; Tian, D. Rapidly evolving R genes in diverse grass species confer resistance to rice blast disease. Proc. Natl. Acad. Sci. USA 2013, 110, 18572–18577. [Google Scholar] [CrossRef] [PubMed]
- Lewis, J.D.; Lee, A.; Ma, W.; Zhou, H.; Guttman, D.S.; Desveaux, D. The YopJ superfamily in plant-associated bacteria. Mol. Plant Pathol. 2011, 12, 928–937. [Google Scholar] [CrossRef]
- Ma, K.W.; Ma, W. YopJ Family Effectors Promote Bacterial Infection through a Unique Acetyltransferase Activity. Microbiol. Mol. Biol. Rev. 2016, 80, 1011–1027. [Google Scholar] [CrossRef]
- Orth, K.; Xu, Z.; Mudgett, M.B.; Bao, Z.Q.; Palmer, L.E.; Bliska, J.B.; Mangel, W.F.; Staskawicz, B.; Dixon, J.E. Disruption of signaling by Yersinia effector YopJ, a ubiquitin-like protein protease. Science 2000, 290, 1594–1597. [Google Scholar] [CrossRef]
- Le Roux, C.; Huet, G.; Jauneau, A.; Camborde, L.; Trémousaygue, D.; Kraut, A.; Zhou, B.; Levaillant, M.; Adachi, H.; Yoshioka, H.; et al. A receptor pair with an integrated decoy converts pathogen disabling of transcription factors to immunity. Cell 2015, 161, 1074–1088. [Google Scholar] [CrossRef]
- Sarris, P.F.; Duxbury, Z.; Huh, S.U.; Ma, Y.; Segonzac, C.; Sklenar, J.; Derbyshire, P.; Cevik, V.; Rallapalli, G.; Saucet, S.B.; et al. A Plant Immune Receptor Detects Pathogen Effectors that Target WRKY Transcription Factors. Cell 2015, 161, 1089–1100. [Google Scholar] [CrossRef] [PubMed]
- Cheong, M.S.; Kirik, A.; Kim, J.G.; Frame, K.; Kirik, V.; Mudgett, M.B. AvrBsT acetylates Arabidopsis ACIP1, a protein that associates with microtubules and is required for immunity. PLoS Pathog. 2014, 10, e1003952. [Google Scholar] [CrossRef] [PubMed]
- Jiang, S.; Yao, J.; Ma, K.W.; Zhou, H.; Song, J.; He, S.Y.; Ma, W. Bacterial effector activates jasmonate signaling by directly targeting JAZ transcriptional repressors. PLoS Pathog. 2013, 9, e1003715. [Google Scholar] [CrossRef] [PubMed]
- Lee, A.H.; Hurley, B.; Felsensteiner, C.; Yea, C.; Ckurshumova, W.; Bartetzko, V.; Wang, P.W.; Quach, V.; Lewis, J.D.; Liu, Y.C.; et al. A bacterial acetyltransferase destroys plant microtubule networks and blocks secretion. PLoS Pathog. 2012, 8, e1002523. [Google Scholar] [CrossRef] [PubMed]
- Lewis, J.D.; Wu, R.; Guttman, D.S.; Desveaux, D. Allele-specific virulence attenuation of the Pseudomonas syringae HopZ1a type III effector via the Arabidopsis ZAR1 resistance protein. PLoS Genet. 2010, 6, e1000894. [Google Scholar] [CrossRef]
- Lewis, J.D.; Lee, A.H.; Hassan, J.A.; Wan, J.; Hurley, B.; Jhingree, J.R.; Wang, P.W.; Lo, T.; Youn, J.Y.; Guttman, D.S.; et al. The Arabidopsis ZED1 pseudokinase is required for ZAR1-mediated immunity induced by the Pseudomonas syringae type III effector HopZ1a. Proc. Natl. Acad. Sci. USA 2013, 110, 18722–18727. [Google Scholar] [CrossRef]
- Lee, J.; Manning, A.J.; Wolfgeher, D.; Jelenska, J.; Cavanaugh, K.A.; Xu, H.; Fernandez, S.M.; Michelmore, R.W.; Kron, S.J.; Greenberg, J.T. Acetylation of an NB-LRR Plant Immune-Effector Complex Suppresses Immunity. Cell Rep. 2015, 13, 1670–1682. [Google Scholar] [CrossRef]
- Mittal, R.; Peak-Chew, S.Y.; Sade, R.S.; Vallis, Y.; McMahon, H.T. The acetyltransferase activity of the bacterial toxin YopJ of Yersinia is activated by eukaryotic host cell inositol hexakisphosphate. J. Biol. Chem. 2010, 285, 19927–19934. [Google Scholar] [CrossRef]
- Zhang, Z.M.; Ma, K.W.; Yuan, S.; Luo, Y.; Jiang, S.; Hawara, E.; Pan, S.; Ma, W.; Song, J. Structure of a pathogen effector reveals the enzymatic mechanism of a novel acetyltransferase family. Nat. Struct. Mol. Biol. 2016, 23, 847–852. [Google Scholar] [CrossRef]
- Mukherjee, S.; Keitany, G.; Li, Y.; Wang, Y.; Ball, H.L.; Goldsmith, E.J.; Orth, K. Yersinia YopJ acetylates and inhibits kinase activation by blocking phosphorylation. Science 2006, 312, 1211–1214. [Google Scholar] [CrossRef]
- Trosky, J.E.; Li, Y.; Mukherjee, S.; Keitany, G.; Ball, H.; Orth, K. VopA inhibits ATP binding by acetylating the catalytic loop of MAPK kinases. J. Biol. Chem. 2007, 282, 34299–34305. [Google Scholar] [CrossRef] [PubMed]
- Adeolu, M.; Alnajar, S.; Naushad, S.; Gupta, R.S. Genome-based phylogeny and taxonomy of the ‘Enterobacteriales’: Proposal for Enterobacterales ord. nov. divided into the families Enterobacteriaceae, Erwiniaceae fam. nov., Pectobacteriaceae fam. nov., Yersiniaceae fam. nov., Hafniaceae fam. nov., Morganellaceae fam. nov., and Budviciaceae fam. nov. Int. J. Syst. Evol. Microbiol. 2016, 66, 5575–5599. [Google Scholar] [CrossRef] [PubMed]
- Janda, J.M.; Abbott, S.L. The Changing Face of the Family Enterobacteriaceae (Order: “Enterobacterales”): New Members, Taxonomic Issues, Geographic Expansion, and New Diseases and Disease Syndromes. Clin. Microbiol. Rev. 2021, 34, 46–76. [Google Scholar] [CrossRef] [PubMed]
- Yuan, X.; Hulin, M.T.; Sundin, G.W. Effectors, chaperones, and harpins of the Type III secretion system in the fire blight pathogen Erwinia amylovora: A review. J. Plant Pathol. 2021, 103, 25–39. [Google Scholar] [CrossRef]
- Zhao, Y. Genomics of Erwinia amylovora and Related Erwinia Species Associated with Pome Fruit Trees. In Genomics of Plant-Associated Bacteria; Gross, D.C., Lichens-Park, A., Kole, C., Eds.; Springer Berlin Heidelberg: Berlin, Heidelberg, 2014; pp. 1–36. [Google Scholar]
- Panstruga, R.; Moscou, M.J. What is the Molecular Basis of Nonhost Resistance? Mol. Plant Microbe Interact. 2020, 33, 1253–1264. [Google Scholar] [CrossRef]
- Toruño, T.Y.; Stergiopoulos, I.; Coaker, G. Plant-Pathogen Effectors: Cellular Probes Interfering with Plant Defenses in Spatial and Temporal Manners. Annu. Rev. Phytopathol. 2016, 54, 419–441. [Google Scholar] [CrossRef]
- Vleeshouwers, V.G.; Rietman, H.; Krenek, P.; Champouret, N.; Young, C.; Oh, S.K.; Wang, M.; Bouwmeester, K.; Vosman, B.; Visser, R.G.; et al. Effector genomics accelerates discovery and functional profiling of potato disease resistance and phytophthora infestans avirulence genes. PLoS ONE 2008, 3, e2875. [Google Scholar] [CrossRef]
- Lewis, J.D.; Abada, W.; Ma, W.; Guttman, D.S.; Desveaux, D. The HopZ family of Pseudomonas syringae type III effectors require myristoylation for virulence and avirulence functions in Arabidopsis thaliana. J. Bacteriol. 2008, 190, 2880–2891. [Google Scholar] [CrossRef]
- Jayaraman, J.; Choi, S.; Prokchorchik, M.; Choi, D.S.; Spiandore, A.; Rikkerink, E.H.; Templeton, M.D.; Segonzac, C.; Sohn, K.H. A bacterial acetyltransferase triggers immunity in Arabidopsis thaliana independent of hypersensitive response. Sci. Rep. 2017, 7, 3557. [Google Scholar] [CrossRef]
- Vinatzer, B.A.; Teitzel, G.M.; Lee, M.W.; Jelenska, J.; Hotton, S.; Fairfax, K.; Jenrette, J.; Greenberg, J.T. The type III effector repertoire of Pseudomonas syringae pv. syringae B728a and its role in survival and disease on host and non-host plants. Mol. Microbiol. 2006, 62, 26–44. [Google Scholar] [CrossRef]
- Olawole, O.I.; Liu, Q.; Chen, C.; Gleason, M.L.; Beattie, G.A. The Contributions to Virulence of the Effectors Eop1 and DspE Differ Between Two Clades of Erwinia tracheiphila Strains. Mol. Plant Microbe Interact. 2021, 34, 1399–1408. [Google Scholar] [CrossRef] [PubMed]
- Ma, W.; Dong, F.F.; Stavrinides, J.; Guttman, D.S. Type III effector diversification via both pathoadaptation and horizontal transfer in response to a coevolutionary arms race. PLoS Genet. 2006, 2, e209. [Google Scholar] [CrossRef] [PubMed]
- Nazareno, E.S.; Kersey, C.M.; Dumenyo, C.K. Characterization of the incompatible interaction between Erwinia tracheiphila and non-host tobacco (Nicotiana tabacum). Physiol. Mol. Plant Pathol. 2016, 96, 85–93. [Google Scholar] [CrossRef]
- Zhang, Z.M.; Ma, K.W.; Gao, L.; Hu, Z.; Schwizer, S.; Ma, W.; Song, J. Mechanism of host substrate acetylation by a YopJ family effector. Nat. Plants 2017, 3, 17115. [Google Scholar] [CrossRef]
- Polgár, L. The catalytic triad of serine peptidases. Cell Mol. Life Sci. 2005, 62, 2161–2172. [Google Scholar] [CrossRef]
- Catalano, C.; Al Mughram, M.H.; Guo, Y.; Kellogg, G.E. 3D interaction homology: Hydropathic interaction environments of serine and cysteine are strikingly different and their roles adapt in membrane proteins. Curr. Res. Struct. Biol. 2021, 3, 239–256. [Google Scholar] [CrossRef]
- Ballinger, P.; Long, F.A. Acid Ionization Constants of Alcohols. II. Acidities of Some Substituted Methanols and Related Compounds1,2. J. Am. Chem. Soc. 1960, 82, 795–798. [Google Scholar] [CrossRef]
- Gisdon, F.J.; Bombarda, E.; Ullmann, G.M. Serine and Cysteine Peptidases: So Similar, Yet Different. How the Active-Site Electrostatics Facilitates Different Reaction Mechanisms. J. Phys. Chem. B 2022, 126, 4035–4048. [Google Scholar] [CrossRef]
- Nozaki, Y.; Tanford, C. [84] Examination of titration behavior. In Methods in Enzymology; Academic Press: Cambridge, MA, USA, 1967; Volume 11, pp. 715–734. [Google Scholar]
- Thurlkill, R.L.; Grimsley, G.R.; Scholtz, J.M.; Pace, C.N. pK values of the ionizable groups of proteins. Protein Sci. 2006, 15, 1214–1218. [Google Scholar] [CrossRef]
- Patel, S. A critical review on serine protease: Key immune manipulator and pathology mediator. Allergol. Immunopathol. 2017, 45, 579–591. [Google Scholar] [CrossRef]
- Warshel, A.; Naray-Szabo, G.; Sussman, F.; Hwang, J.K. How do serine proteases really work? Biochemistry 1989, 28, 3629–3637. [Google Scholar] [CrossRef] [PubMed]
- Prah, A.; Frančišković, E.; Mavri, J.; Stare, J. Electrostatics as the Driving Force Behind the Catalytic Function of the Monoamine Oxidase A Enzyme Confirmed by Quantum Computations. ACS Catal. 2019, 9, 1231–1240. [Google Scholar] [CrossRef]
- Mladenovic, M.; Fink, R.F.; Thiel, W.; Schirmeister, T.; Engels, B. On the origin of the stabilization of the zwitterionic resting state of cysteine proteases: A theoretical study. J. Am. Chem. Soc. 2008, 130, 8696–8705. [Google Scholar] [CrossRef] [PubMed]
- Ishida, T.; Kato, S. Theoretical perspectives on the reaction mechanism of serine proteases: The reaction free energy profiles of the acylation process. J. Am. Chem. Soc. 2003, 125, 12035–12048. [Google Scholar] [CrossRef] [PubMed]
- Beveridge, A.J. A theoretical study of the active sites of papain and S195C rat trypsin: Implications for the low reactivity of mutant serine proteinases. Protein Sci. 1996, 5, 1355–1365. [Google Scholar] [CrossRef]
- Dardenne, L.E.; Werneck, A.S.; de Oliveira Neto, M.; Bisch, P.M. Electrostatic properties in the catalytic site of papain: A possible regulatory mechanism for the reactivity of the ion pair. Proteins 2003, 52, 236–253. [Google Scholar] [CrossRef]
- Asadi, M.; Oanca, G.; Warshel, A. Effect of Environmental Factors on the Catalytic Activity of Intramembrane Serine Protease. J. Am. Chem. Soc. 2022, 144, 1251–1257. [Google Scholar] [CrossRef]
- Madeira, F.; Park, Y.M.; Lee, J.; Buso, N.; Gur, T.; Madhusoodanan, N.; Basutkar, P.; Tivey, A.R.N.; Potter, S.C.; Finn, R.D.; et al. The EMBL-EBI search and sequence analysis tools APIs in 2019. Nucleic Acids Res. 2019, 47, W636–W641. [Google Scholar] [CrossRef]
- Jumper, J.; Evans, R.; Pritzel, A.; Green, T.; Figurnov, M.; Ronneberger, O.; Tunyasuvunakool, K.; Bates, R.; Žídek, A.; Potapenko, A.; et al. Highly accurate protein structure prediction with AlphaFold. Nature 2021, 596, 583–589. [Google Scholar] [CrossRef]
- Park, D.H.; Yu, J.G.; Oh, E.J.; Han, K.S.; Yea, M.C.; Lee, S.J.; Myung, I.S.; Shim, H.S.; Oh, C.S. First Report of Fire Blight Disease on Asian Pear Caused by Erwinia amylovora in Korea. Plant Dis. 2016, 100, 1946. [Google Scholar] [CrossRef]
- Jeleńska, J.; Lee, J.; Manning, A.J.; Wolfgeher, D.J.; Ahn, Y.; Walters-Marrah, G.; Lopez, I.E.; Garcia, L.; McClerklin, S.A.; Michelmore, R.W.; et al. Pseudomonas syringae effector HopZ3 suppresses the bacterial AvrPto1-tomato PTO immune complex via acetylation. PLoS Pathog. 2021, 17, e1010017. [Google Scholar] [CrossRef] [PubMed]
- Kawai, T.; Akira, S. The role of pattern-recognition receptors in innate immunity: Update on Toll-like receptors. Nat. Immunol. 2010, 11, 373–384. [Google Scholar] [CrossRef] [PubMed]
- Asselin, J.E.; Bonasera, J.M.; Kim, J.F.; Oh, C.S.; Beer, S.V. Eop1 from a Rubus strain of Erwinia amylovora functions as a host-range limiting factor. Phytopathology 2011, 101, 935–944. [Google Scholar] [CrossRef] [PubMed]
- Cheng, Y.; Oldfield, C.J.; Meng, J.; Romero, P.; Uversky, V.N.; Dunker, A.K. Mining α-Helix-Forming Molecular Recognition Features with Cross Species Sequence Alignments. Biochemistry 2007, 46, 13468–13477. [Google Scholar] [CrossRef]
- Disfani, F.M.; Hsu, W.-L.; Mizianty, M.J.; Oldfield, C.J.; Xue, B.; Dunker, A.K.; Kurgan, L. MoRFpred, a computational tool for sequence-based prediction and characterization of short disorder-to-order transitioning binding regions in proteins. Bioinformatics 2012, 28, i75–i83. [Google Scholar] [CrossRef]
- Kim, W.S.; Jock, S.; Paulin, J.P.; Rhim, S.L.; Geider, K. Molecular Detection and Differentiation of Erwinia pyrifoliae and Host Range Analysis of the Asian Pear Pathogen. Plant Dis. 2001, 85, 1183–1188. [Google Scholar] [CrossRef]
- Kube, M.; Migdoll, A.M.; Muller, I.; Kuhl, H.; Beck, A.; Reinhardt, R.; Geider, K. The genome of Erwinia tasmaniensis strain Et1/99, a non-pathogenic bacterium in the genus Erwinia. Environ. Microbiol. 2008, 10, 2211–2222. [Google Scholar] [CrossRef]
- Lee, G.M.; Ko, S.; Oh, E.J.; Song, Y.R.; Kim, D.; Oh, C.S. Comparative Genome Analysis Reveals Natural Variations in the Genomes of Erwinia pyrifoliae, a Black Shoot Blight Pathogen in Apple and Pear. Plant Pathol. J. 2020, 36, 428–439. [Google Scholar] [CrossRef]
- Rojas, E.S.; Batzer, J.C.; Beattie, G.A.; Fleischer, S.J.; Shapiro, L.R.; Williams, M.A.; Gleason, M.L. Bacterial Wilt of Cucurbits: Resurrecting a Classic Pathosystem. Plant Dis. 2015, 99, 564–574. [Google Scholar] [CrossRef]
- Shapiro, L.R.; Paulson, J.N.; Arnold, B.J.; Scully, E.D.; Zhaxybayeva, O.; Pierce, N.E.; Kolter, R. An Introduced Crop Plant Is Driving Diversification of the Virulent Bacterial Pathogen Erwinia tracheiphila. mBio 2018, 9, e01307–e01318. [Google Scholar] [CrossRef]
- Walterson, A.M.; Stavrinides, J. Pantoea: Insights into a highly versatile and diverse genus within the Enterobacteriaceae. FEMS Microbiol. Rev. 2015, 3, 968–984. [Google Scholar] [CrossRef] [PubMed]










Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tomar, V.; Rikkerink, E.H.A.; Song, J.; Sofkova-Bobcheva, S.; Bus, V.G.M. Structure-Function Characterisation of Eop1 Effectors from the Erwinia-Pantoea Clade Reveals They May Acetylate Their Defence Target through a Catalytic Dyad. Int. J. Mol. Sci. 2023, 24, 14664. https://doi.org/10.3390/ijms241914664
Tomar V, Rikkerink EHA, Song J, Sofkova-Bobcheva S, Bus VGM. Structure-Function Characterisation of Eop1 Effectors from the Erwinia-Pantoea Clade Reveals They May Acetylate Their Defence Target through a Catalytic Dyad. International Journal of Molecular Sciences. 2023; 24(19):14664. https://doi.org/10.3390/ijms241914664
Chicago/Turabian StyleTomar, Vishant, Erik H. A. Rikkerink, Janghoon Song, Svetla Sofkova-Bobcheva, and Vincent G. M. Bus. 2023. "Structure-Function Characterisation of Eop1 Effectors from the Erwinia-Pantoea Clade Reveals They May Acetylate Their Defence Target through a Catalytic Dyad" International Journal of Molecular Sciences 24, no. 19: 14664. https://doi.org/10.3390/ijms241914664